
Harnessing Paenarthrobacter ureafaciens YL1 and Pseudomonas koreensis YL2 Interactions to Improve Degradation of Sulfamethoxazole


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Pure Strain Isolation and Culture Conditions
2.3. Identification of Isolates and Morphology Characterization
2.4. Construction of a SMX-Degrading Consortium
2.5. Determination of Optimal Conditions
2.6. Analysis Methods of Biotransformation
2.7. Quantitative PCR (qPCR) Analysis
2.8. Whole Genome Sequencing and Assembly
2.9. Genome Annotation
3. Results
3.1. Isolation and Identification Characterization of Strain YL2
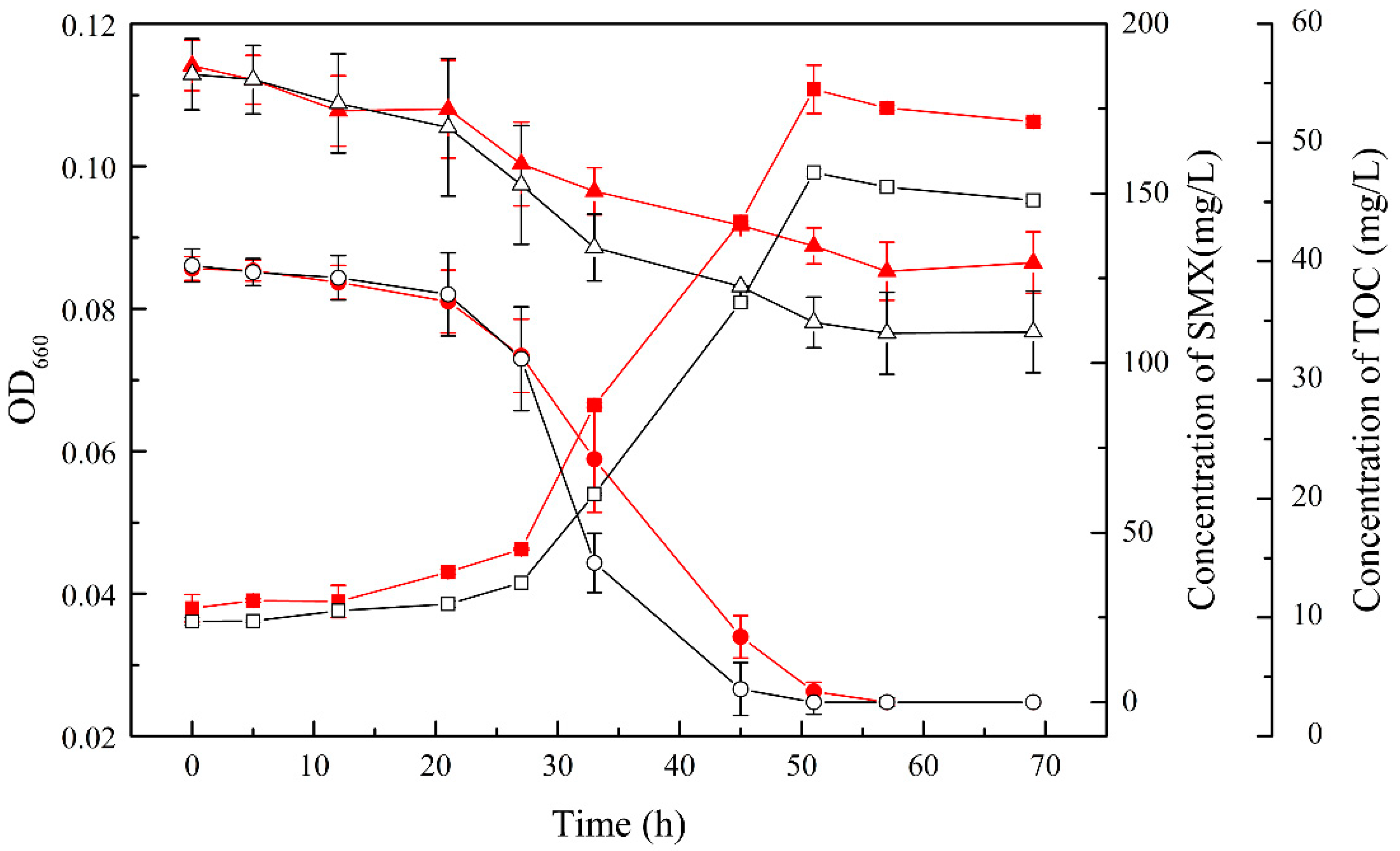
3.2. SMX Biodegradation Efficiency of the Consortium
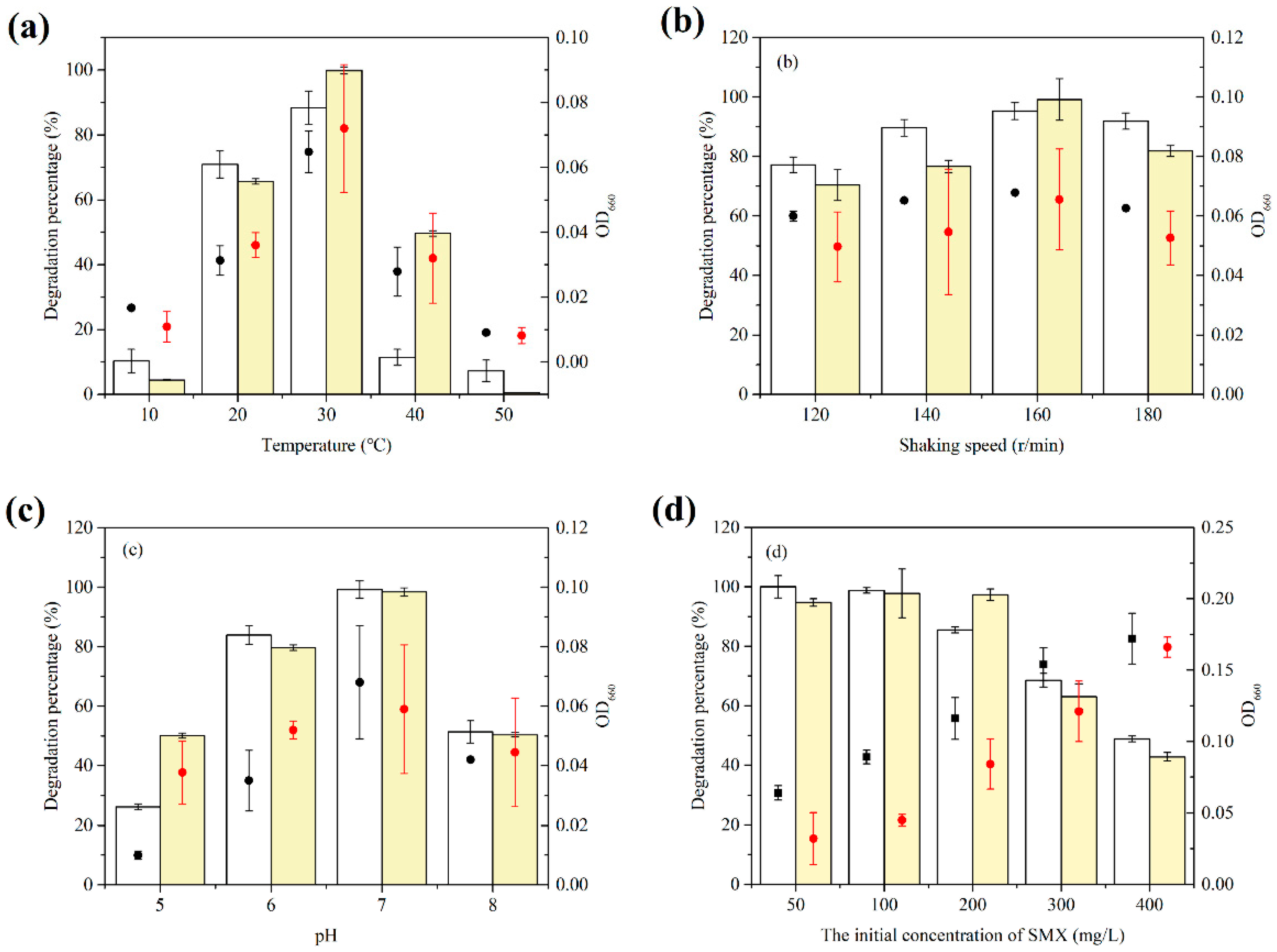
3.3. Effect of Different Growth Conditions on SMX Biodegradation by the Consortium
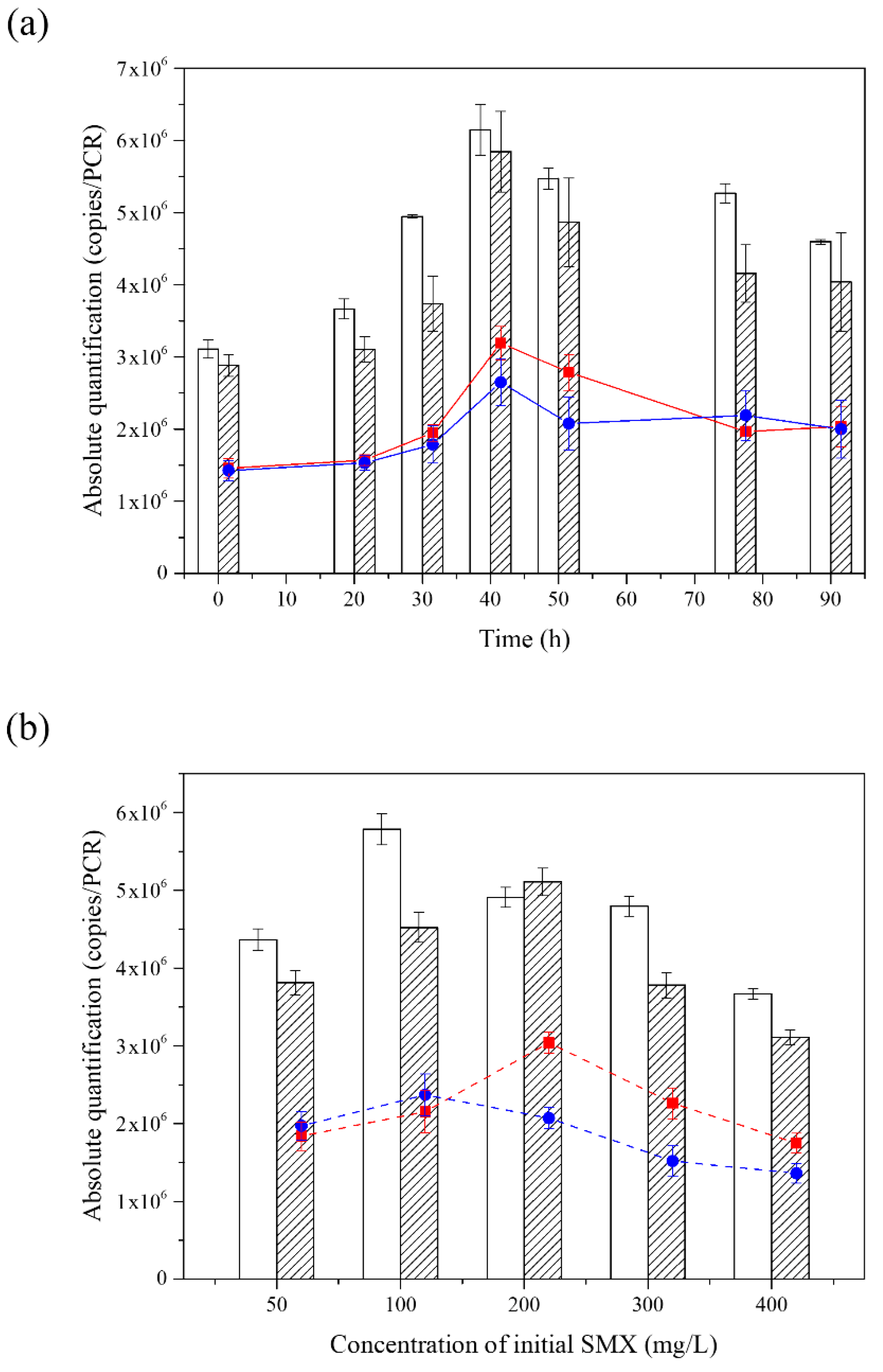
3.4. The Analysis of Composition in the Consortium during the Degradation of SMX
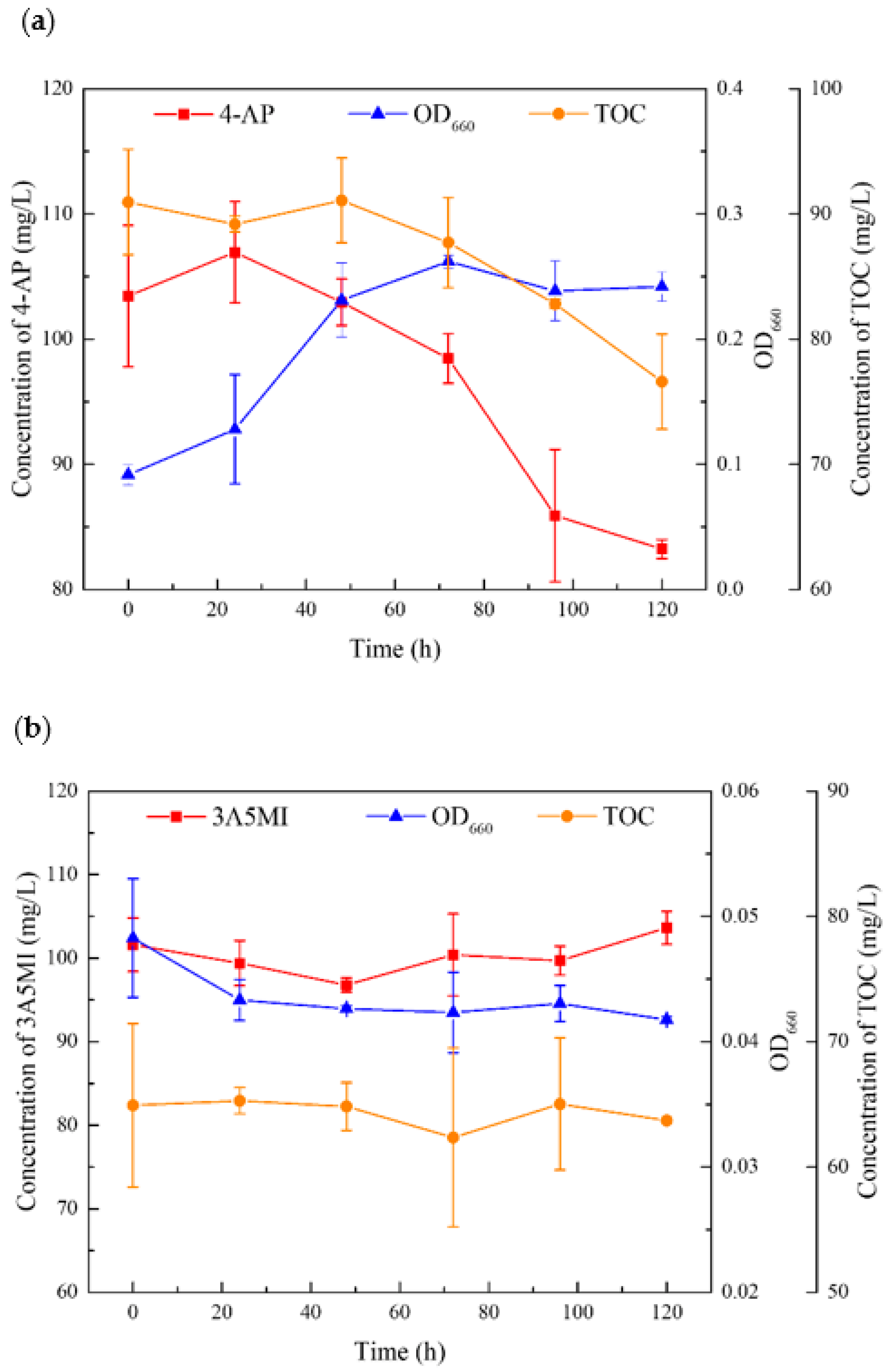
3.5. Utilization of the Major Intermediates by YL2 during the Degradation of SMX
3.6. Genomic Features and Putative Functional Classification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liang, D.H.; Hu, Y. Simultaneous sulfamethoxazole biodegradation and nitrogen conversion by achromobacter sp. Jl9 using with different carbon and nitrogen sources. Bioresour. Technol. 2019, 293, 122061. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.Q.; Kim, S.J.; Kurade, M.B.; Govindwar, S.; Abou-Shanab, R.A.I.; Kim, J.R.; Roh, H.S.; Khan, M.A.; Jeon, B.H. Combined effects of sulfamethazine and sulfamethoxazole on a freshwater microalga, scenedesmus obliquus: Toxicity, biodegradation, and metabolic fate. J. Hazard. Mater. 2019, 370, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Poch, M.; Garrido-Baserba, M.; Corominas, L.; Perello-Moragues, A.; Monclus, H.; Cermeron-Romero, M.; Melitas, N.; Jiang, S.C.; Rosso, D. When the fourth water and digital revolution encountered covid-19. Sci. Total Environ. 2020, 744, 140980. [Google Scholar] [CrossRef]
- Wang, J.; Zhuan, R.; Chu, L. The occurrence, distribution and degradation of antibiotics by ionizing radiation: An overview. Sci. Total Environ. 2019, 646, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Pelalak, R.; Alizadeh, R.; Ghareshabani, E.; Heidari, Z. Degradation of sulfonamide antibiotics using ozone-based advanced oxidation process: Experimental, modeling, transformation mechanism and dft study. Sci. Total Environ. 2020, 734, 139446. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Guo, M.; Wang, Q.; Li, Z.; Wang, C.; Chen, N.; Wang, C.-C.; Wan, C.; Chen, S. Controllable synthesis of cerium zirconium oxide nanocomposites and their application for photocatalytic degradation of sulfonamides. Appl. Catal. B Environ. 2019, 259, 118107. [Google Scholar] [CrossRef]
- Lei, K.; Zhu, Y.; Chen, W.; Pan, H.Y.; Cao, Y.X.; Zhang, X.; Guo, B.B.; Sweetman, A.; Lin, C.Y.; Ouyang, W.; et al. Spatial and seasonal variations of antibiotics in river waters in the haihe river catchment in china and ecotoxicological risk assessment. Environ. Int. 2019, 130, 104919. [Google Scholar] [CrossRef]
- Wang, J.; Chu, L.; Wojnarovits, L.; Takacs, E. Occurrence and fate of antibiotics, antibiotic resistant genes (args) and antibiotic resistant bacteria (arb) in municipal wastewater treatment plant: An overview. Sci. Total Environ. 2020, 744, 140997. [Google Scholar] [CrossRef]
- Fang, W.; Peng, Y.; Muir, D.; Lin, J.; Zhang, X. A critical review of synthetic chemicals in surface waters of the us, the eu and china. Environ. Int 2019, 131, 104994. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, F.; Yang, J.; Guo, H.; Su, D.; Yu, L. Adaption and degradation strategies of methylotrophic 1,4-dioxane degrading strain xanthobacter sp. Yn2 revealed by transcriptome-scale analysis. Int. J. Mol. Sci. 2021, 22, 10435. [Google Scholar] [CrossRef]
- Larcher, S.; Yargeau, V. Biodegradation of sulfamethoxazole: Current knowledge and perspectives. Appl. Microbiol. Biotechnol. 2012, 96, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Xie, B.; Tang, X.; Ng, H.Y.; Deng, S.; Shi, X.; Song, W.; Huang, S.; Li, G.; Liang, H. Biological sulfamethoxazole degradation along with anaerobically digested centrate treatment by immobilized microalgal-bacterial consortium: Performance, mechanism and shifts in bacterial and microalgal communities. Chem. Eng. J. 2020, 388, 124217. [Google Scholar] [CrossRef]
- Wang, S.; Wang, J. Biodegradation and metabolic pathway of sulfamethoxazole by a novel strain acinetobacter sp. Appl. Microbiol. Biotechnol. 2018, 102, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hu, Y.; Wang, J. Biodegradation of typical pharmaceutical compounds by a novel strain acinetobacter sp. J. Environ. Manag. 2018, 217, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Zhang, C.; Huang, D.; Wang, R.; Zeng, G.; Yan, M.; Xiong, W.; Zhou, C.; Cheng, M.; Xue, W.; et al. Recent progress in sustainable technologies for adsorptive and reactive removal of sulfonamides. Chem. Eng. J. 2020, 389, 123423. [Google Scholar] [CrossRef]
- Chen, K.L.; Liu, L.C.; Chen, W.R. Adsorption of sulfamethoxazole and sulfapyridine antibiotics in high organic content soils. Environ. Pollut. 2017, 231, 1163–1171. [Google Scholar] [CrossRef]
- Liu, Y.; Fan, Q.; Wang, J. Zn-fe-cnts catalytic in situ generation of h2o2 for fenton-like degradation of sulfamethoxazole. J. Hazard. Mater. 2018, 342, 166–176. [Google Scholar] [CrossRef]
- Chen, H.; Wang, J. Degradation of sulfamethoxazole by ozonation combined with ionizing radiation. J. Hazard. Mater. 2021, 407, 124377. [Google Scholar] [CrossRef]
- Wang, S.; Liu, Y.; Wang, J. Peroxymonosulfate activation by Fe-Co-O-Codoped graphite carbon nitride for degradation of sulfamethoxazole. Environ. Sci. Technol. 2020, 54, 10361–10369. [Google Scholar] [CrossRef]
- Wang, J.; Zhuan, R. Degradation of antibiotics by advanced oxidation processes: An overview. Sci. Total Environ. 2020, 701, 135023. [Google Scholar] [CrossRef]
- Song, H.; Yan, L.; Jiang, J.; Ma, J.; Pang, S.; Zhai, X.; Zhang, W.; Li, D. Enhanced degradation of antibiotic sulfamethoxazole by electrochemical activation of pds using carbon anodes. Chem. Eng. J. 2018, 344, 12–20. [Google Scholar] [CrossRef]
- Wang, J.; Chu, L. Irradiation treatment of pharmaceutical and personal care products (ppcps) in water and wastewater: An overview. Radiat. Phys. Chem. 2016, 125, 56–64. [Google Scholar] [CrossRef]
- Yu, T.H.; Lin, A.Y.; Panchangam, S.C.; Hong, P.K.; Yang, P.Y.; Lin, C.F. Biodegradation and bio-sorption of antibiotics and non-steroidal anti-inflammatory drugs using immobilized cell process. Chemosphere 2011, 84, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Pang, W.; Dou, C.; Yin, D. Sulfamethoxazole and cod increase abundance of sulfonamide resistance genes and change bacterial community structures within sequencing batch reactors. Chemosphere 2017, 175, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wang, Y.; Su, X.; Fu, Y.; Ma, F.; Guo, H. Biodiversity, isolation and genome analysis of sulfamethazine-degrading bacteria using high-throughput analysis. Bioprocess. Biosyst. Eng. 2020, 2020, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, S. Microbial degradation of sulfamethoxazole in the environment. Appl. Microbiol. Biotechnol. 2018, 102, 3573–3582. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Li, A.; Cui, D.; Cai, R.; Ma, F.; Wang, Y. Biodegradation and metabolic pathway of sulfamethoxazole by pseudomonas psychrophila ha-4, a newly isolated cold-adapted sulfamethoxazole-degrading bacterium. Appl. Microbiol. Biotechnol. 2014, 98, 4671–4681. [Google Scholar] [CrossRef]
- Reis, P.J.; Reis, A.C.; Ricken, B.; Kolvenbach, B.A.; Manaia, C.M.; Corvini, P.F.; Nunes, O.C. Biodegradation of sulfamethoxazole and other sulfonamides by achromobacter denitrificans pr1. J. Hazard. Mater. 2014, 280, 741–749. [Google Scholar] [CrossRef]
- Ricken, B.; Fellmann, O.; Kohler, H.P.; Schaffer, A.; Corvini, P.F.; Kolvenbach, B.A. Degradation of sulfonamide antibiotics by microbacterium sp. Strain br1-elucidating the downstream pathway. N. Biotechnol. 2015, 32, 710–715. [Google Scholar] [CrossRef]
- Cao, L.; Zhang, J.; Zhao, R.; Deng, Y.; Liu, J.; Fu, W.; Lei, Y.; Zhang, T.; Li, X.; Li, B. Genomic characterization, kinetics, and pathways of sulfamethazine biodegradation by Paenarthrobacter sp. A01. Environ. Int. 2019, 131, 104961. [Google Scholar] [CrossRef]
- Qi, M.; Ma, X.; Liang, B.; Zhang, L.; Kong, D.; Li, Z.; Wang, A. Complete genome sequences of the antibiotic sulfamethoxazole-mineralizing bacteria paenarthrobacter sp. P27 and Norcardiodes sp. N27. Environ. Res. 2022, 204, 112013. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.C.; Cvancarova, M.; Liu, Y.; Lenz, M.; Hettich, T.; Kolvenbach, B.A.; Corvini, P.F.; Nunes, O.C. Biodegradation of sulfamethoxazole by a bacterial consortium of achromobacter denitrificans pr1 and Leucobacter sp. Gp. Appl. Microbiol. Biotechnol. 2018, 102, 10299–10314. [Google Scholar] [CrossRef]
- Qi, M.; Liang, B.; Zhang, L.; Ma, X.; Yan, L.; Dong, W.; Kong, D.; Zhang, L.; Zhu, H.; Gao, S.H.; et al. Microbial interactions drive the complete catabolism of the antibiotic sulfamethoxazole in activated sludge microbiomes. Environ. Sci. Technol. 2021, 55, 3270–3282. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, S. Removal of pharmaceuticals and personal care products (ppcps) from wastewater: A review. J. Environ. Manag. 2016, 182, 620–640. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, Y.; Zheng, Y.; Liu, L.; Zhao, F. Efficient degradation of sulfamethoxazole and the response of microbial communities in microbial fuel cells. RSC Adv. 2015, 5, 56430–56437. [Google Scholar] [CrossRef]
- Liao, X.; Li, B.; Zou, R.; Xie, S. Antibiotic sulfanilamide biodegradation by acclimated microbial populations. Appl. Microbiol. Biotechnol. 2016, 100, 2439–2447. [Google Scholar] [CrossRef] [PubMed]
- Ricken, B.; Kolvenbach, B.A.; Bergesch, C.; Benndorf, D.; Kroll, K.; Strnad, H.; Vlcek, C.; Adaixo, R.; Hammes, F.; Shahgaldian, P.; et al. Fmnh2-dependent monooxygenases initiate catabolism of sulfonamides in microbacterium sp. Strain br1 subsisting on sulfonamide antibiotics. Sci. Rep. 2017, 7, 15783. [Google Scholar] [CrossRef]
- Uniyal, S.; Sharma, R.K.; Kondakal, V. New insights into the biodegradation of chlorpyrifos by a novel bacterial consortium: Process optimization using general factorial experimental design. Ecotoxicol. Environ. Saf. 2021, 209, 111799. [Google Scholar] [CrossRef]
- Ma, F.; Wang, Y.; Yang, J.; Guo, H.; Su, D.; Yu, L. Degradation of 1,4-dioxane by xanthobacter sp. Yn2. Curr. Microbiol. 2021, 78, 992–1005. [Google Scholar] [CrossRef]
- Liang, S.; Duan, J.; Hu, H.; Zhang, J.; Gao, S.; Jing, H.; Li, G.; Sun, Z. Comprehensive analysis of sinps on the genome-wide transcriptional changes in caenorhabditis elegans. Int. J. Nanomed. 2020, 15, 5227–5237. [Google Scholar] [CrossRef]
- Mulla, S.I.; Hu, A.; Sun, Q.; Li, J.; Suanon, F.; Ashfaq, M.; Yu, C.P. Biodegradation of sulfamethoxazole in bacteria from three different origins. J. Environ. Manag. 2018, 206, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Brankatschk, R.; Bodenhausen, N.; Zeyer, J.; Burgmann, H. Simple absolute quantification method correcting for quantitative pcr efficiency variations for microbial community samples. Appl. Environ. Microbiol. 2012, 78, 4481–4489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Ren, Y.; Jiang, N. Analysis of draft genome sequence of pseudomonas sp. Qtf5 reveals its benzoic acid degradation ability and heavy metal tolerance. Biomed. Res. Int. 2017, 2017, 4565960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The rast server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. Card 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef]
- Kertesz, M.A.; Schmidt-Larbig, K.; Wüest, T. A novel reduced flavin mononucleotide-dependent methanesulfonate sulfonatase encoded by the sulfur-regulated msu operon of pseudomonas aeruginosa. J. Bacteriol. 1999, 181, 1464–1473. [Google Scholar] [CrossRef] [Green Version]
- van Der Ploeg, J.R.; Iwanicka-Nowicka, R.; Bykowski, T.; Hryniewicz, M.M.; Leisinger, T. The escherichia coli ssueadcb gene cluster is required for the utilization of sulfur from aliphatic sulfonates and is regulated by the transcriptional activator cbl. J. Biol. Chem. 1999, 274, 29358–29365. [Google Scholar] [CrossRef] [Green Version]
- Mu, D.S.; Liang, Q.Y.; Wang, X.M.; Lu, D.C.; Shi, M.J.; Chen, G.J.; Du, Z.J. Metatranscriptomic and comparative genomic insights into resuscitation mechanisms during enrichment culturing. Microbiome 2018, 6, 230. [Google Scholar] [CrossRef]
- Bouju, H.; Ricken, B.; Beffa, T.; Corvini, P.F.; Kolvenbach, B.A. Isolation of bacterial strains capable of sulfamethoxazole mineralization from an acclimated membrane bioreactor. Appl. Environ. Microbiol. 2012, 78, 277–279. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Li, Y.; Li, X.; Huang, H.; Zheng, G.; Chen, Y. Biological removal of sulfamethoxazole enhanced by s. Oneidensis mr-1 via promoting nadh generation and electron transfer and consumption. J. Hazard. Mater. 2022, 426, 127839. [Google Scholar] [CrossRef]
- Afzal Khan, S.; Hamayun, M.; Ahmed, S. Degradation of 4-aminophenol by newly isolated pseudomonas sp. Strain st-4. Enzym. Microb. Technol. 2006, 38, 10–13. [Google Scholar] [CrossRef]
- Takenaka, S.; Murakami, S.; Shinke, R.; Hatakeyama, K.; Yukawa, H.; Aoki, K. Novel genes encoding 2-aminophenol 1,6-dioxygenase from pseudomonas species ap-3 growing on 2-aminophenol and catalytic properties of the purified enzyme. J. Biol. Chem. 1997, 272, 14727–14732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, K.; Shinke, R.; Nishira, H. Metabolism of aniline by rhodococcus erythropolis an-13. Agric. Biol. Chem. 1983, 47, 1611–1616. [Google Scholar] [CrossRef]
- Helm, V.; Reber, H. Investigation on the regulation of aniline utilization in pseudomonas multivorans strain an 1. Eur. J. Appl. Microbiol. Biotechnol. 1979, 7, 191–199. [Google Scholar] [CrossRef]
- Zhang, J.J.; Liu, H.; Xiao, Y.; Zhang, X.E.; Zhou, N.Y. Identification and characterization of catabolic para-nitrophenol 4-monooxygenase and para-benzoquinone reductase from pseudomonas sp. Strain wbc-3. J. Bacteriol. 2009, 191, 2703–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moonen, M.J.; Kamerbeek, N.M.; Westphal, A.H.; Boeren, S.A.; Janssen, D.B.; Fraaije, M.W.; van Berkel, W.J. Elucidation of the 4-hydroxyacetophenone catabolic pathway in pseudomonas fluorescens acb. J. Bacteriol. 2008, 190, 5190–5198. [Google Scholar] [CrossRef] [Green Version]
- Yonezawa, A.; Inui, K. Novel riboflavin transporter family rfvt/slc52: Identification, nomenclature, functional characterization and genetic diseases of rfvt/slc52. Mol. Aspects Med. 2013, 34, 693–701. [Google Scholar] [CrossRef]
- Zhou, C.; Wang, H.; Si, Y.; Wu, K.; Yousaf, A. Electron shuttles enhance the degradation of sulfamethoxazole coupled with fe(iii) reduction by shewanella oneidensis mr-1. Environ. Toxicol. Pharmacol. 2018, 62, 156–163. [Google Scholar] [CrossRef]
- Nanjani, S.; Paul, D.; Keharia, H. Genome analysis to decipher syntrophy in the bacterial consortium ‘scp’ for azo dye degradation. BMC Microbiol. 2021, 21, 177. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | The Names of Quantitative Primers | The Sequences of Quantitative Primers |
|---|---|---|
| 16S rRNA gene of strain YL1 | YL1 16S rRNA-F | TCCTCAGCGTCAGTTACA |
| YL1 16S rRNA-R | TGGTGTAGCGGTGAAATG | |
| YL1 16S rRNA-probe | FAM-AGAGACCTGCCTTCGCCATCGG-MGB | |
| 16S rRNA gene of strain YL2 | YL2 16S rRNA-F | ATGCGTAGATATAGGAAGGAA |
| YL2 16S rRNA-R | CAGGCGGTCAACTTAATG | |
| YL2 16S rRNA-probe | FAM-ACCACCTGGACTGATACTGACACTGA-MGB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Wang, Y.; Shan, X.; Ma, F.; Guo, H. Harnessing Paenarthrobacter ureafaciens YL1 and Pseudomonas koreensis YL2 Interactions to Improve Degradation of Sulfamethoxazole. Microorganisms 2022, 10, 648. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10030648
Yu L, Wang Y, Shan X, Ma F, Guo H. Harnessing Paenarthrobacter ureafaciens YL1 and Pseudomonas koreensis YL2 Interactions to Improve Degradation of Sulfamethoxazole. Microorganisms. 2022; 10(3):648. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10030648
Chicago/Turabian StyleYu, Lan, Yingning Wang, Xiaoqing Shan, Fang Ma, and Haijuan Guo. 2022. "Harnessing Paenarthrobacter ureafaciens YL1 and Pseudomonas koreensis YL2 Interactions to Improve Degradation of Sulfamethoxazole" Microorganisms 10, no. 3: 648. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10030648