
Sheep Infection Trials with ‘Phase-Locked’ Vpma Expression Variants of Mycoplasma agalactiae—Towards Elucidating the Role of a Multigene Family Encoding Variable Surface Lipoproteins in Infection and Disease

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mycoplasma Strains, Animals and Experimental Infection
2.2. Ethic Statement
2.3. Clinical Examination
2.4. Collection of Milk
2.5. Udder Health, California Mastitis Test (CMT) and Single Cell Counts (SCC)
2.6. Quantitative Analysis of PLMs Recovered from Various Body Sites during Experimental Infection and at Necropsy
2.7. Euthanasia
2.8. Pathology
2.9. Immunohistochemistry
2.10. Statistical Analyses
3. Results
3.1. Comparison of Clinical Manifestations
3.2. Udder Health, Milk Yield and Quality
3.3. M. agalactiae Excretion in Milk: PLMU Is Impaired in Initial Host Colonization and Multiplication at the Site of Infection
3.4. Mycoplasma Re-Isolations from Organs and Lymph Nodes: PLMV Shows Maximum Numbers of Positive Tissues
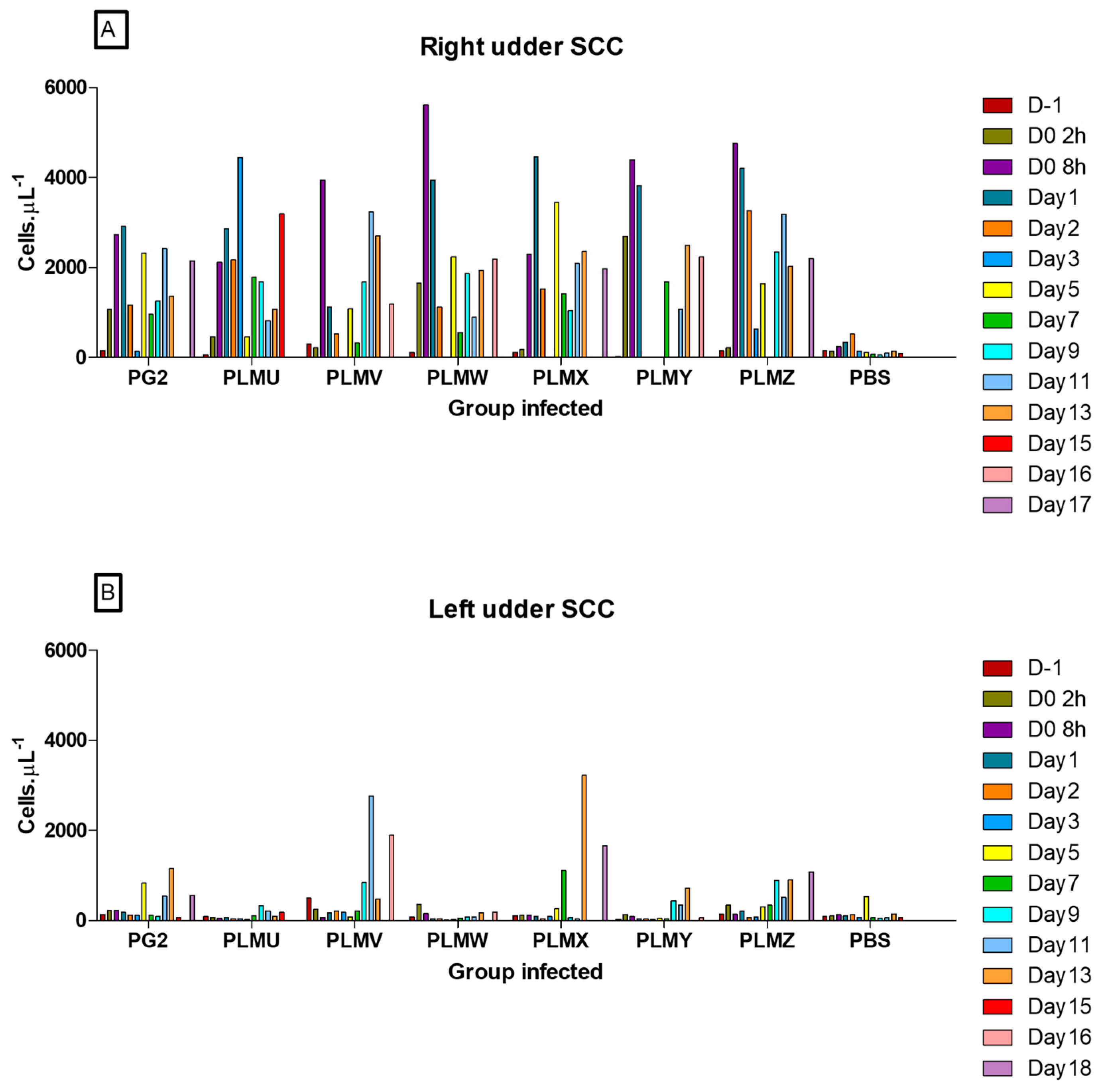
3.5. Immune Cells in the Milk
3.6. Pathological Anatomical Differences: PLMV and PLMX Show Significant Hyperplasia of the Left Udder Lymph Nodes
3.7. Histopathology: PLMV and PLMX Show Inflammatory Cells and Purulent Galactophoritis on the Left Side
3.8. Immunohistochemistry: PLMV Shows Significantly Higher Spread to Tissues and Organs Away from Inoculation Site
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosengarten, R.; Citti, C.; Glew, M.; Lischewski, A.; Droesse, M.; Much, P.; Winner, F.; Brank, M.; Spergser, J. Host-pathogen interactions in mycoplasma pathogenesis: Virulence and survival strategies of minimalist prokaryotes. Int. J. Med. Microbiol. 2000, 290, 15–25. [Google Scholar] [CrossRef]
- Nicholas, R.A.J.; Ayling, R.D.; Mcauliffe, L. Vaccines for Mycoplasma Diseases in Animals and Man. J. Comp. Pathol. 2009, 140, 85–96. [Google Scholar] [CrossRef]
- Calcutt, M.J.; Lysnyansky, I.; Sachse, K.; Fox, L.K.; Nicholas, R.A.J.; Ayling, R.D. Gap analysis of Mycoplasma bovis disease, diagnosis and control: An aid to identify future development requirements. Transbound. Emerg. Dis. 2018, 65, 91–109. [Google Scholar] [CrossRef] [Green Version]
- Cacciotto, C.; Addis, M.F.; Pagnozzi, D.; Coradduzza, E.; Pittau, M.; Alberti, A. Identification of conserved Mycoplasma agalactiae surface antigens by immunoproteomics. Vet. Immunol. Immunopathol. 2021, 236, 110239. [Google Scholar] [CrossRef]
- Arachchige, S.N.K.; Condello, A.K.; Zhu, L.; Shil, P.K.; Tivendale, K.A.; Underwood, G.J.; Noormohammadi, A.H.; Browning, G.F.; Wawegama, N.K. Effects of immunosuppression on the efficacy of vaccination against Mycoplasma gallisepticum infection in chickens. Vet. Microbiol. 2021, 260, 109182. [Google Scholar] [CrossRef]
- Shaw, B.M.; Simmons, W.L.; Dybvig, K. The vsa shield of Mycoplasma pulmonis is antiphagocytic. Infect. Immun. 2012, 80, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Gaurivaud, P.; Baranowski, E.; Pau-Roblot, C.; Sagné, E.; Citti, C.; Tardy, F. Mycoplasma agalactiae secretion of β-(1→6)-glucan, a rare polysaccharide in prokaryotes, is governed by high-frequency phase variation. Appl. Environ. Microbiol. 2016, 82, 3370–3383. [Google Scholar] [CrossRef] [Green Version]
- Czurda, S.; Hegde, S.M.; Rosengarten, R.; Chopra-Dewasthaly, R. Xer1-independent mechanisms of Vpma phase variation in Mycoplasma agalactiae are triggered by Vpma-specific antibodies. Int. J. Med. Microbiol. 2017, 307, 443–451. [Google Scholar] [CrossRef]
- Pflaum, K.; Tulman, E.R.; Canter, J.; Dhondt, K.V.; Reinoso-Perez, M.T.; Dhondt, A.A.; Geary, S.J. The influence of host tissue on M. gallisepticum vlhA gene expression. Vet. Microbiol. 2020, 251, 108891. [Google Scholar] [CrossRef]
- Chopra-Dewasthaly, R.; Spergser, J.; Zimmermann, M.; Citti, C.; Jechlinger, W.; Rosengarten, R.; Lewinsohn, D.M. Vpma phase variation is important for survival and persistence of Mycoplasma agalactiae in the immunocompetent host. PLoS Pathog. 2017, 13, e1006656. [Google Scholar] [CrossRef] [Green Version]
- Christodoulides, A.; Gupta, N.; Yacoubian, V.; Maithel, N.; Parker, J.; Kelesidis, T. The Role of Lipoproteins in Mycoplasma-Mediated Immunomodulation. Front. Microbiol. 2018, 9, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migliore, S.; Puleio, R.; Nicholas, R.A.J.; Loria, G.R.; Rodriguez, L.F.; Gómez-Martin, Á. Mycoplasma agalactiae: The Sole Cause of Classical Contagious Agalactia? Animals 2021, 11, 1782. [Google Scholar] [CrossRef] [PubMed]
- Chopra-Dewasthaly, R.; Baumgartner, M.; Gamper, E.; Innerebner, C.; Zimmermann, M.; Schilcher, F.; Tichy, A.; Winter, P.; Jechlinger, W.; Rosengarten, R.; et al. Role of Vpma phase variation in Mycoplasma agalactiae pathogenesis. FEMS Immunol. Med. Microbiol. 2012, 66, 307–322. [Google Scholar] [CrossRef] [Green Version]
- Glew, M.D.; Marenda, M.; Rosengarten, R.; Citti, C. Surface diversity in Mycoplasma agalactiae is driven by site-specific DNA inversions within the vpma multigene locus. J. Bacteriol. 2002, 184, 5987–5998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra-Dewasthaly, R.; Citti, C.; Glew, M.D.; Zimmermann, M.; Rosengarten, R.; Jechlinger, W. Phase-locked mutants of Mycoplasma agalactiae: Defining the molecular switch of high-frequency Vpma antigenic variation. Mol. Microbiol. 2008, 67, 1196–1210. [Google Scholar] [CrossRef] [Green Version]
- Czurda, S.; Jechlinger, W.; Rosengarten, R.; Chopra-Dewasthaly, R. Xer1-mediated site-specific DNA inversions and excisions in Mycoplasma agalactiae. J. Bacteriol. 2010, 192, 4462–4473. [Google Scholar] [CrossRef] [Green Version]
- Nouvel, L.X.; Marenda, M.; Sirand-Pugnet, P.; Sagné, E.; Glew, M.; Mangenot, S.; Barbe, V.; Barré, A.; Claverol, S.; Citti, C. Occurrence, plasticity, and evolution of the vpma gene family, a genetic system devoted to high-frequency surface variation in Mycoplasma agalactiae. J. Bacteriol. 2009, 191, 4111–4121. [Google Scholar] [CrossRef] [Green Version]
- Nouvel, L.X.; Sirand-Pugnet, P.; Marenda, M.S.; Sagné, E.; Barbe, V.; Mangenot, S.; Schenowitz, C.; Jacob, D.; Barré, A.; Claverol, S.; et al. Comparative genomic and proteomic analyses of two Mycoplasma agalactiae strains: Clues to the macro- and micro-events that are shaping mycoplasma diversity. BMC Genom. 2010, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Hegde, S.; Zimmermann, M.; Rosengarten, R.; Chopra-Dewasthaly, R. Novel role of Vpmas as major adhesins of Mycoplasma agalactiae mediating differential cell adhesion and invasion of Vpma expression variants. Int. J. Med. Microbiol. 2018, 308, 263–270. [Google Scholar] [CrossRef]
- Gonzalez, Y.R.C.; Bascunana, C.R.; Bölske, G.; Mattsson, J.G.; Molina, C.F.; Johansson, K.E. In vitro amplification of the 16S rRNA genes from Mycoplasma bovis and Mycoplasma agalactiae by PCR. Vet. Microbiol. 1995, 47, 183–190. [Google Scholar] [CrossRef]
- Baumgartner, W.; Aurich, C. Clinical Propaedeutics of Pets and Domestic Animals; Stuttgart, Ed.; Eighth Rev.; Enke: Stuttgart, Germany, 2014; p. 525. [Google Scholar]
- Lillie, R.D.; Fullmer, H.M. Histopathologic Technic and Practical Histochemistry, 4th ed.; McGraw Hill: New York, NY, USA, 1976; 942p, ISBN 0070378622. [Google Scholar]
- Hegde, S.; Hegde, S.; Spergser, J.; Brunthaler, R.; Rosengarten, R.; Chopra-Dewasthaly, R. In vitro and in vivo cell invasion and systemic spreading of Mycoplasma agalactiae in the sheep infection model. Int. J. Med. Microbiol. 2014, 304, 1024–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauf, A.C.W.; Rosenbusch, R.F.; Paape, M.J.; Bannerman, D.D. Innate immune response to intramammary Mycoplasma bovis infection. J. Dairy Sci. 2007, 90, 3336–3348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegde, S.; Hegde, S.; Zimmermann, M.; Flöck, M.; Spergser, J.; Rosengarten, R.; Chopra-Dewasthaly, R. Simultaneous identification of potential pathogenicity factors of Mycoplasma agalactiae in the natural ovine host by negative selection. Infect. Immun. 2015, 83, 2751–2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegde, S.; Zimmermann, M.; Flöck, M.; Brunthaler, R.; Spergser, J.; Rosengarten, R.; Chopra-Dewasthaly, R. Genetic loci of Mycoplasma agalactiae involved in systemic spreading during experimental intramammary infection of sheep. Vet. Res. 2016, 47, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-alonso, A.; Rodríguez, F.; De Fé, C.; Monteros, A.E.D.L.; Poveda, J.B.; Andrada, M.; Herráez, P. Correlating the immune response with the clinical–pathological course of persistent mastitis experimentally induced by Mycoplasma agalactiae in dairy goats. Res. Vet. Sci. 2009, 86, 274–280. [Google Scholar] [CrossRef]
- Castro-Alonso, A.; De la Fe, C.; de los Monteros, A.E.; Rodríguez, F.; Andrada, M.; Poveda, J.B.; Herráez, P. Chronological and immunohistochemical characterization of the mammary immunoinflammatory response in experimental caprine contagious agalactia. Vet. Immunol. Immunopathol. 2010, 136, 43–54. [Google Scholar] [CrossRef]
- Bergonier, D.; Berthelot, X.; Poumarat, F. Contagious agalactia of small ruminants: Current knowledge concerning epidemiology, diagnosis and control. Rev. Sci. Tech. Int. Off. Epizoot. 1997, 16, 848–873. [Google Scholar] [CrossRef]
- Kizil, O.; Ozdemir, H. Clinical, haematological and biochemical studies in goats naturally infected with Mycoplasma agalactiae. Bull. Vet. Inst. Pulawy 2006, 50, 325–328. [Google Scholar]
- Cacciotto, C.; Cubeddu, T.; Addis, M.F.; Anfossi, A.G.; Tedde, V.; Tore, G.; Carta, T.; Rocca, S.; Chessa, B.; Pittau, M.; et al. Mycoplasma lipoproteins are major determinants of neutrophil extracellular trap formation. Cell. Microbiol. 2016, 18, 1751–1762. [Google Scholar] [CrossRef]
- Flöck, M.; Brunthaler, R.; Spergser, J.; Chopra-Dewasthaly, R.; Wittek, T. Sonographic and histological udder parenchyma changes after intramammary infection of sheep with Mycoplasma agalactiae. Schweiz. Arch. Tierheilkd. 2021, 163, 851–858. [Google Scholar] [CrossRef]
- Kirchhoff, H.; Rosengarten, R. Isolation of a motile mycoplasma from fish. J. Gen. Microbiol. 1984, 130, 2439–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Groups | Positive Control Group | Negative Control Group | Mutant Group |
|---|---|---|---|
| (A) | |||
| Inoculum | M. agalactiae type strain PG2 | sterile & pyrogen free saline | M. agalactiae Vpma |
| phase-locked mutants | |||
| PLMU, PLMY and PLMW | |||
| Inoculum Size | 109 CFU | - | 109 CFU |
| Infection Route | intramammary | intramammary | intramammary |
| via right teat | via right teat | via right teat | |
| Animals | 3 lactating ewes | 3 lactating ewes | 3 lactating ewes/PLMU |
| 3 lactating ewes/PLMY | |||
| 3 lactating ewes/PLMW | |||
| =9 animals | |||
| (B) | |||
| Inoculum | M. agalactiae type strain PG2 | sterile & pyrogen free saline | M. agalactiae Vpma |
| phase-locked mutants | |||
| PLMX, PLMZ and PLMV | |||
| Inoculum Size | 109 CFU | - | 109 CFU |
| Infection Route | intramammary | intramammary | intramammary |
| via right teat | via right teat | via right teat | |
| Animals | 3 lactating ewes | 3 lactating ewes | 3 lactating ewes/PLMX |
| 3 lactating ewes/PLMZ | |||
| 3 lactating ewes/PLMV | |||
| =9 animals | |||
| Day p.i. 1 | Clinical Examination 2 | Feed Intake, Milk Yield | Body Temp 3 | Blood 4 | Milk 5 | Swabs 6 |
|---|---|---|---|---|---|---|
| D-5 | X | X | X | X | ||
| D-4 | X | X | X | |||
| D-3 | X | X | X | |||
| D-2 | X | X | X | |||
| D-1 | X | X | X | X | X | X |
| D0 | X | X | X | |||
| D0 2 h p.i. | X | X | X | |||
| D0 4 h p.i. | X | X | ||||
| D0 8 h p.i. | X | X | X | X | ||
| D0 12 h p.i. | X | X | ||||
| D1 | X | X | X | X | X | |
| D2 | X | X | X | X | X | |
| D3 | X | X | X | X | ||
| D4 | X | X | X | |||
| D5 | X | X | X | X | ||
| D6 | X | X | X | |||
| D7 | X | X | X | X | X | |
| D8 | X | X | X | |||
| D9 | X | X | X | X | X | |
| D10 | X | X | X | |||
| D11 | X | X | X | X | X | |
| D12 | X | X | X | |||
| D13 | X | X | X | X | ||
| D14 | X | X | X | |||
| D15 | X | X | X | X | X | X |
| D15–18 | Euthanasia and pathological examination | |||||
| Appetite | General Behavior | Conjunctiva | Pulse | Breathing | Auscultation Lungs | Points | General Condition |
|---|---|---|---|---|---|---|---|
| +++ | Calm and attentive | Pale pink–moderately red/anemic | 56–100 | 16–30 | Slightly a.v. | 4 | not affected |
| ++ | Slightly changed | moderately reddened | 101–124 | 21–45 | moderately a.v. | 3 | slightly impaired |
| + | Moderately changed | highly reddened | >125 | 46–60 | moderate to highly a.v. | 2 | moderately impaired |
| 0 | Highly changed | highly reddened | >125 | >60 | highly a.v. | 1 | highly impaired |
| Assessment | Test Pattern | Cells/mL of Milk |
|---|---|---|
| −/(+) | Liquid/Slight streaking | <150,000 cells/100,000–250,000 |
| + | High streaking | 200,000–700,000 |
| ++ | Gel formation | 500,000–1,500,000 |
| +++ | Gel and clog formation | >1,000,000 |
| CMT | Pain | Milk Secretion | Udder Consistency | Lymph Node Enlargement | Points | Mastitis |
|---|---|---|---|---|---|---|
| - | - | - | se; sebu | - | 4 | no |
| +/+++ | - | - | se; sebu | - | 3 | sub-clinical |
| +/+++ | - | slight to moderately changed | se; sebu | - | 2 | mild |
| +/+++ | −/+ | Slight to moderately changed | se; buco; seco | enlarged | 1 | moderate |
| ++/+++ | ++/+++ | Slight to highly changed | co | enlarged | 0 | severe |
| Part I | ||||||||||||
| Infection Group | PG2 | PLMW | PLMY | PLMU | ||||||||
| Organ/Tissue | Sheep 4 | Sheep 5 | Sheep 6 | Sheep 7 | Sheep 8 | Sheep 9 | Sheep 10 | Sheep 11 | Sheep 12 | Sheep 13 | Sheep 14 | Sheep 15 |
| Supramammary LN Left | + | + | + | + | + | + | + | + | + | |||
| Supramammary LN Right | + | + | + | + | + | + | + | + | + | + | + | + |
| Mandibular LN Left | + | |||||||||||
| Iliac LN Left | + | |||||||||||
| Iliac LN Right | + | + | + | + | + | |||||||
| Parotidial LN Left | + | |||||||||||
| Popliteal LN Right | + | |||||||||||
| Med retropharyngeal LN | + | |||||||||||
| Udder Left | + | + | + | + | + | + | + | + | + | + | + | + |
| Udder Right | + | + | + | + | + | + | + | + | + | + | + | + |
| Kidney Right | + | + | + | |||||||||
| Brain | + | |||||||||||
| Carpel Joint tissue Left | + | |||||||||||
| Knee joint Left | + | + | ||||||||||
| Subiliac LN Left | + | |||||||||||
| Subiliac LN Right | + | + | + | + | + | + | + | |||||
| Adrenal gland Left | + | |||||||||||
| Total Isolations | 8 | 5 | 4 | 7 | 8 | 6 | 6 | 4 | 5 | 6 | 5 | 7 |
| PG2 = 17 | PLMW = 21 | PLMY = 15 | PLMU = 18 | |||||||||
| Part II | ||||||||||||
| Infection Group | PLMV | PLMX | PLMZ | PG2 | ||||||||
| Organ/Tissue | Sheep 4 | Sheep 5 | Sheep 6 | Sheep 7 | Sheep 8 | Sheep 9 | Sheep 10 | Sheep 11 | Sheep 12 | Sheep 13 | Sheep 14 | Sheep 15 |
| Supramammary LN Left | + | + | + | + | + | + | ||||||
| Supramammary LN Right | + | + | + | + | + | + | + | + | + | + | + | + |
| Iliac LN Left | + | + | ||||||||||
| Iliac LN Right | + | + | + | + | ||||||||
| LN Cerv superficialis Left | + | |||||||||||
| LN Cerv superficialis Right | + | |||||||||||
| Mediastinal LN | + | |||||||||||
| Med retropharyngeal LN L | + | + | ||||||||||
| Med retropharyngeal LN R | + | + | ||||||||||
| Lateral retropharyngeal LN | + | |||||||||||
| Udder Left | + | + | + | + | + | |||||||
| Udder Right | + | + | + | + | + | + | + | + | + | + | + | |
| Kidney Left | + | + | ||||||||||
| Kidney Right | + | |||||||||||
| Carpel joint fluid right | + | |||||||||||
| Subiliac LN Right | + | + | + | |||||||||
| Adrenal gland Left | + | + | ||||||||||
| Adrenal gland Right | + | |||||||||||
| Total isolations | 8 | 6 | 9 | 5 | 4 | 7 | 5 | 2 | 5 | 3 | 2 | 2 |
| PLMV = 23 | PLMX = 16 | PLMZ = 12 | PG2 = 7 | |||||||||
- Part I: +: positive for M. agalactiae isolation. The following LN/tissue samples were negative for M. agalactaie isolation in all above tested sheep and hence not placed in this part: Mandibular LN R, Mesenterial LN, Mediastinal LN, Cervicalis superficialis LN L/R, Parotidial LN R, Jejunal LN, Lateral retropharyngeal LN, Popliteal LN L, Carpel Joint Tissue R; Carpel Joint Fluid L/R, Knee joint R, Knee Joint Fluid L/R, Adrenal gland R, Heart, liver, spleen, Kidney L, Lung L/R.
- Part II: +: positive for M. agalactiae isolation. The following LN/tissue samples were negative for M agalactaie isolation in all above tested sheep and hence not placed in this part: Mesenterial LN, Parotidial LN L/R Mandibular LN L/R, Popliteal LN L/R, Lung L/R, Subiliac LN L, Liver, Heart, Brain, Spleen, Uterus/fallopian tubes; Jejunal LN; Carpel Joint tissue L/R; Carpel Joint Fluid L, Knee Joint L/R; Knee Joint Fluid L/R.
| Group | Total Clearings of Secondary Follicles |
|---|---|
| Negative Control | 3 |
| PG2 (positive control) | 5 |
| PLM W | 1 |
| PLM Y | 2 |
| PLM U | 3 |
| PLM V | 14 |
| PLM X | 6 |
| PLM Z | 11 |
| Tissue | Udder Right | Udder Left | Supramammary LN Right | Supramammary LN Left | Subiliacus LN Right | Iliacus LN Right |
|---|---|---|---|---|---|---|
| Total positive detections | 18 | 3 | 19 | 6 | 10 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chopra-Dewasthaly, R.; Dagn, A.; Lohinger, C.; Brunthaler, R.; Flöck, M.; Kargl, M.; Hegde, S.; Spergser, J.; Rosengarten, R. Sheep Infection Trials with ‘Phase-Locked’ Vpma Expression Variants of Mycoplasma agalactiae—Towards Elucidating the Role of a Multigene Family Encoding Variable Surface Lipoproteins in Infection and Disease. Microorganisms 2022, 10, 815. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040815
Chopra-Dewasthaly R, Dagn A, Lohinger C, Brunthaler R, Flöck M, Kargl M, Hegde S, Spergser J, Rosengarten R. Sheep Infection Trials with ‘Phase-Locked’ Vpma Expression Variants of Mycoplasma agalactiae—Towards Elucidating the Role of a Multigene Family Encoding Variable Surface Lipoproteins in Infection and Disease. Microorganisms. 2022; 10(4):815. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040815
Chicago/Turabian StyleChopra-Dewasthaly, Rohini, Andreas Dagn, Christian Lohinger, René Brunthaler, Martina Flöck, Munkhtsetseg Kargl, Shrilakshmi Hegde, Joachim Spergser, and Renate Rosengarten. 2022. "Sheep Infection Trials with ‘Phase-Locked’ Vpma Expression Variants of Mycoplasma agalactiae—Towards Elucidating the Role of a Multigene Family Encoding Variable Surface Lipoproteins in Infection and Disease" Microorganisms 10, no. 4: 815. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040815