
Effects of ε-Poly-L-Lysine Combined with Wuyiencin as a Bio-Fungicide against Botryris cinerea

Abstract
:1. Introduction
2. Material and Methods
2.1. ε-PL and Wuyiencin Production and Assays
2.2. Pathogenic Fungi and Plants
2.3. Effect of ε-PL on Mycelial Growth
2.4. Synergistic Effects of ε-PL Combined with Wuyiencin on B. cinerea Mycelial Growth
2.5. The Effects of Treatment on B. cinerea Mycelial Morphology
2.6. Detection of the Spore Germination of B. cinerea
2.7. Antifungal Activity on Detached Leaves
2.8. Effects of the Combined Treatment on the Pathogen on Isolated Tomato Leaves under a Scanning Electron Microscope
2.9. Determination of Synergistic Effect of ε-PL Combined with Wuyiencin
2.10. Treatment Responses of the Tomato Antioxidant Enzyme System
2.10.1. Catalase (CAT) Activity Determination
2.10.2. Superoxide Dismutase (SOD) Activity Determination
2.10.3. Detection of Peroxidase (POD) Activity Determination
2.10.4. Phenylalanine Ammonia Lyase (PAL) Activity Detection
2.11. Statistical Analysis
3. Results
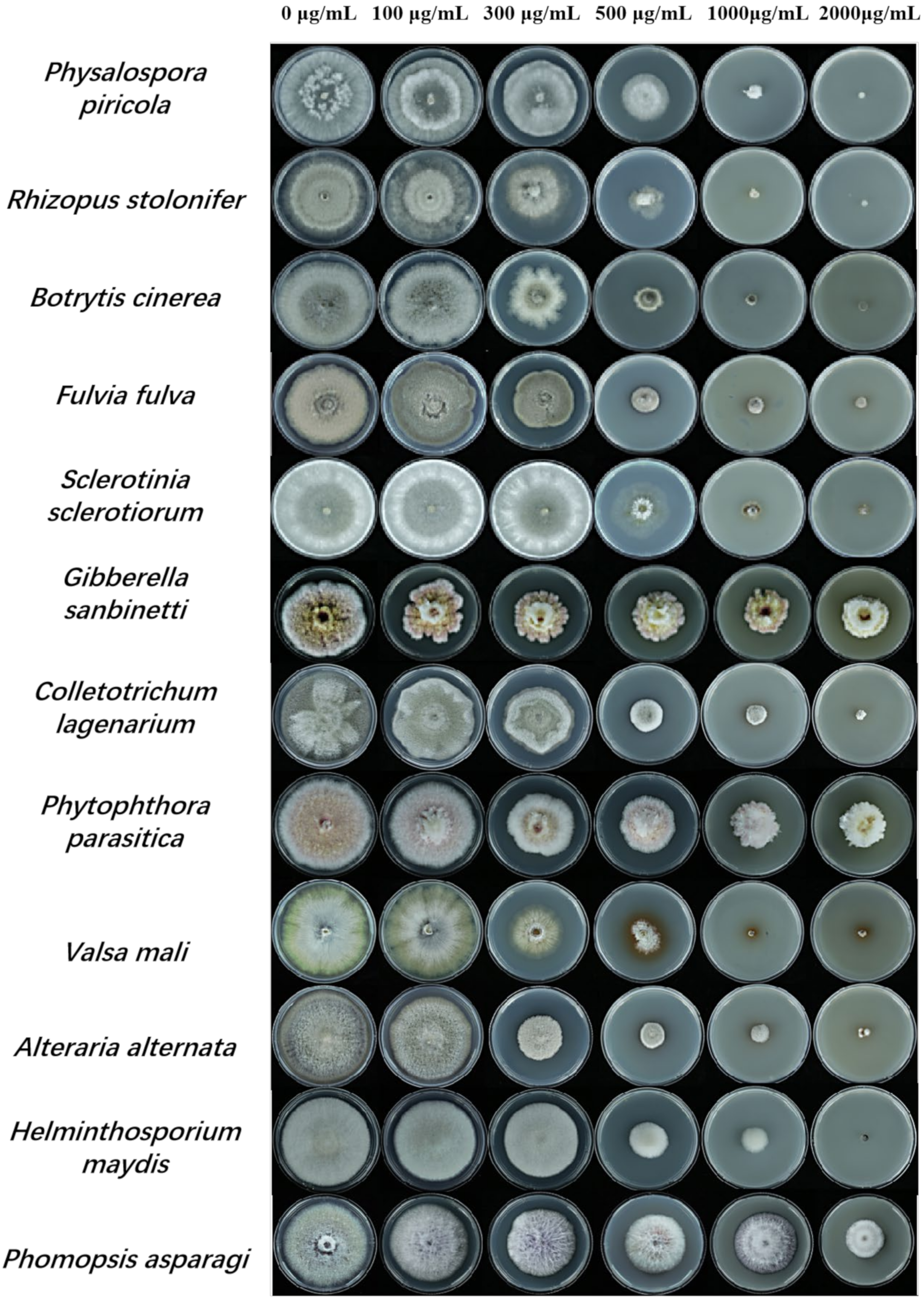
3.1. The Spectrum of ε-PL’s Antifungal Activity
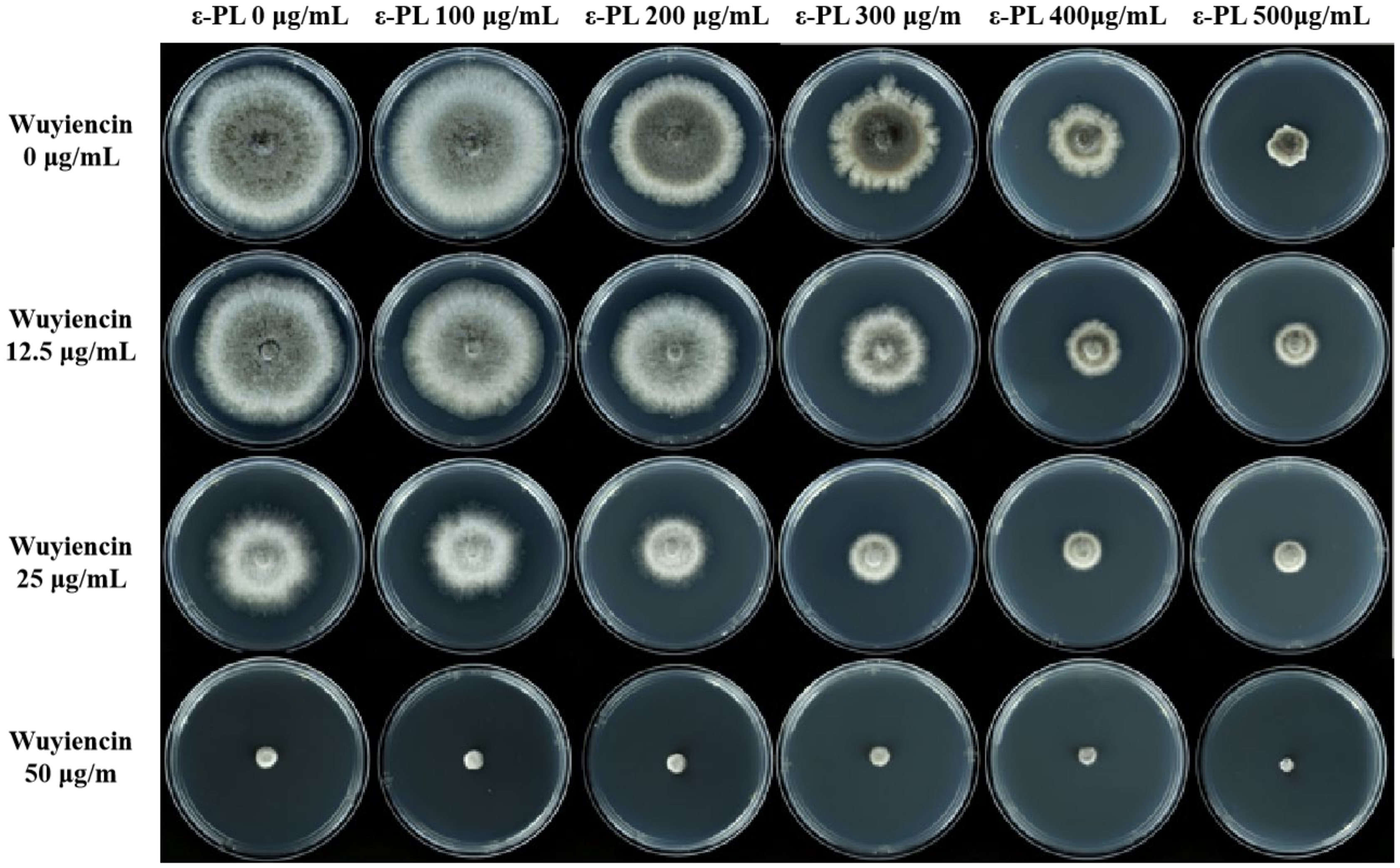
3.2. Effect of Combined Treatment on the Mycelial Growth of B. cinerea
3.3. Effect of Treatment on the Mycelial Morphology of B. cinerea
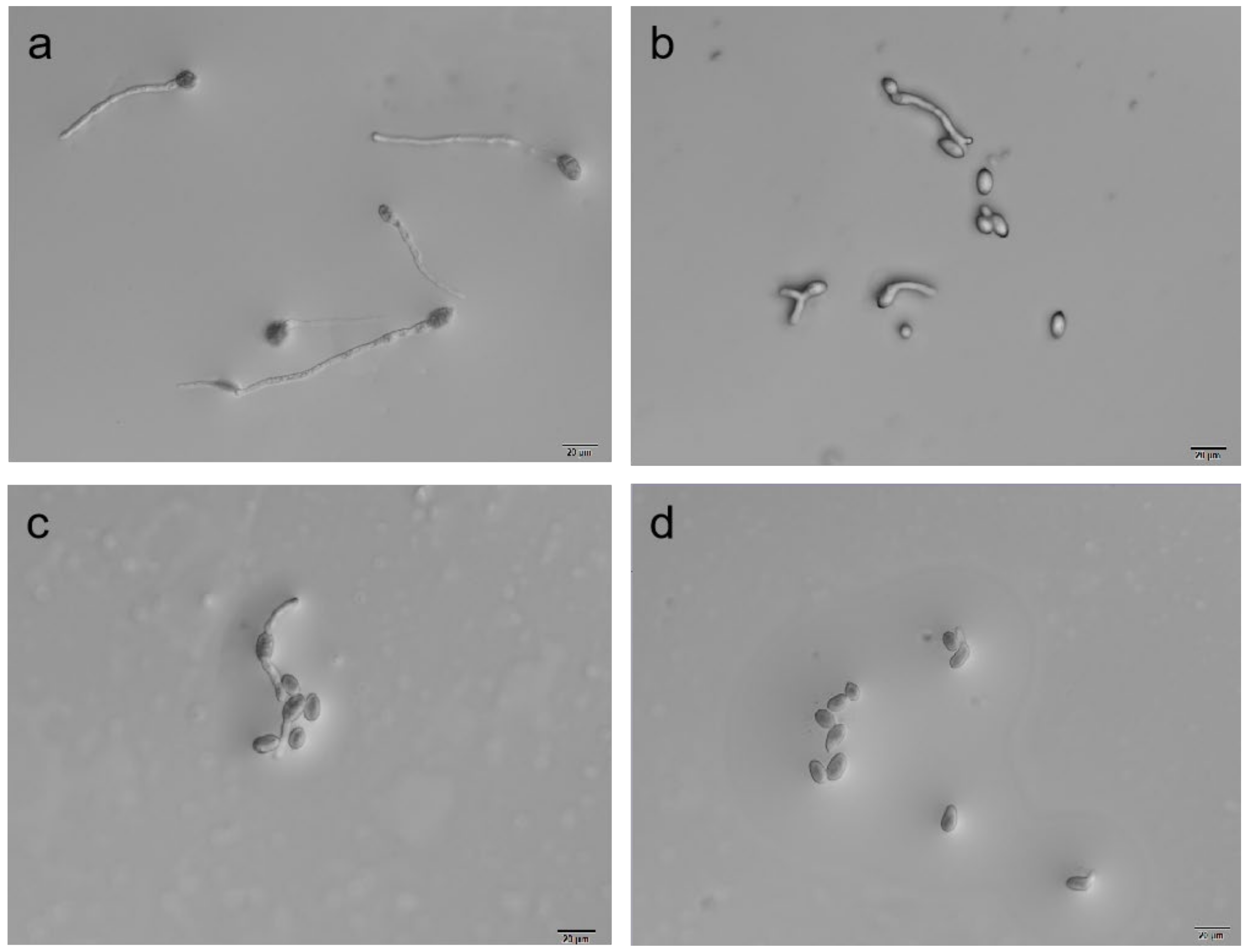
3.4. Detection of B. cinerea Spore Germination
3.5. Antifungal Activity on Detached Leaves
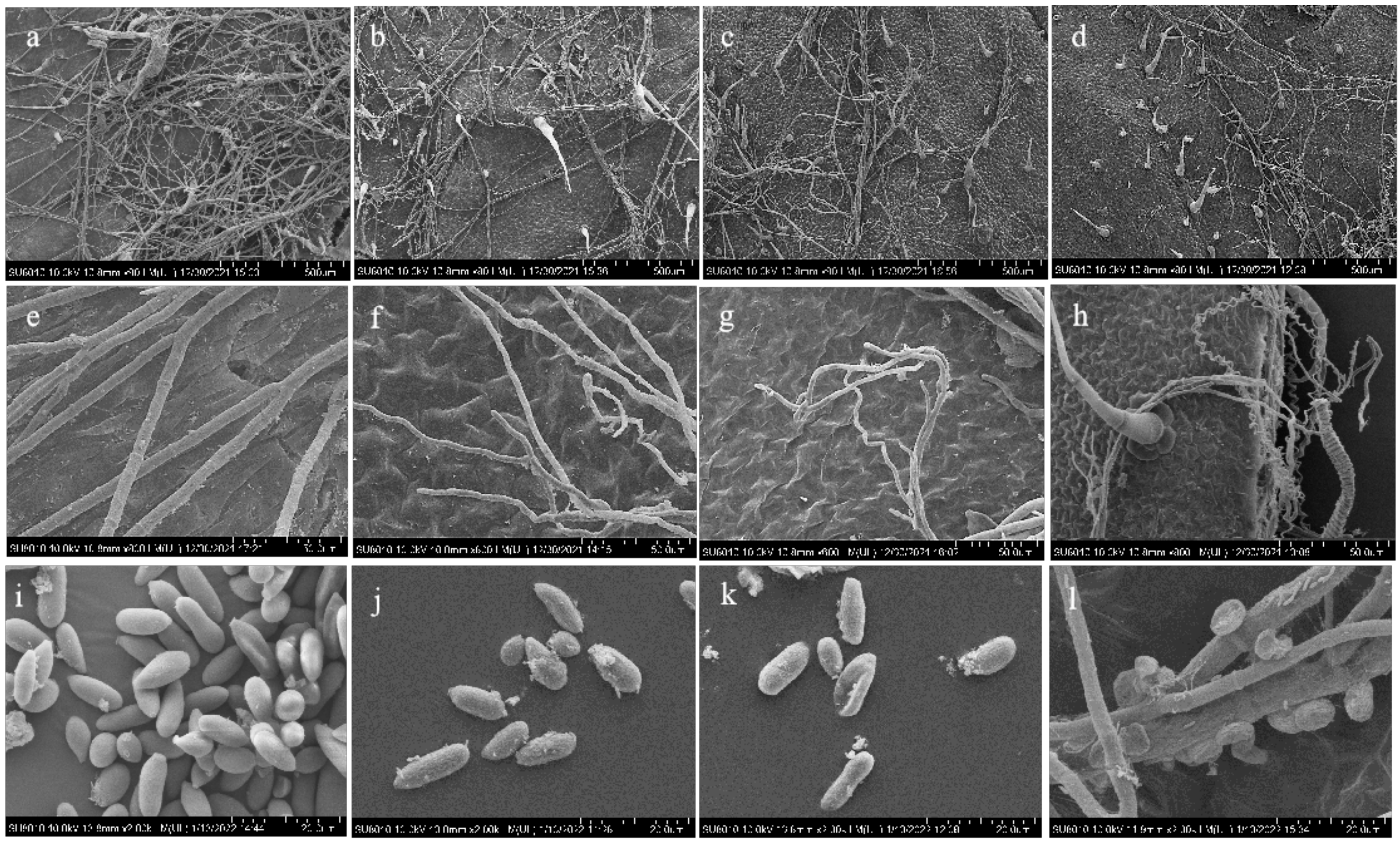
3.6. Scanning Electron Microscope Observation of the Effects of Combined Treatment on the Pathogen on Isolated Tomato Leaves
3.7. Determination of Synergistic Effect of ε-PL Combined with Wuyiencin
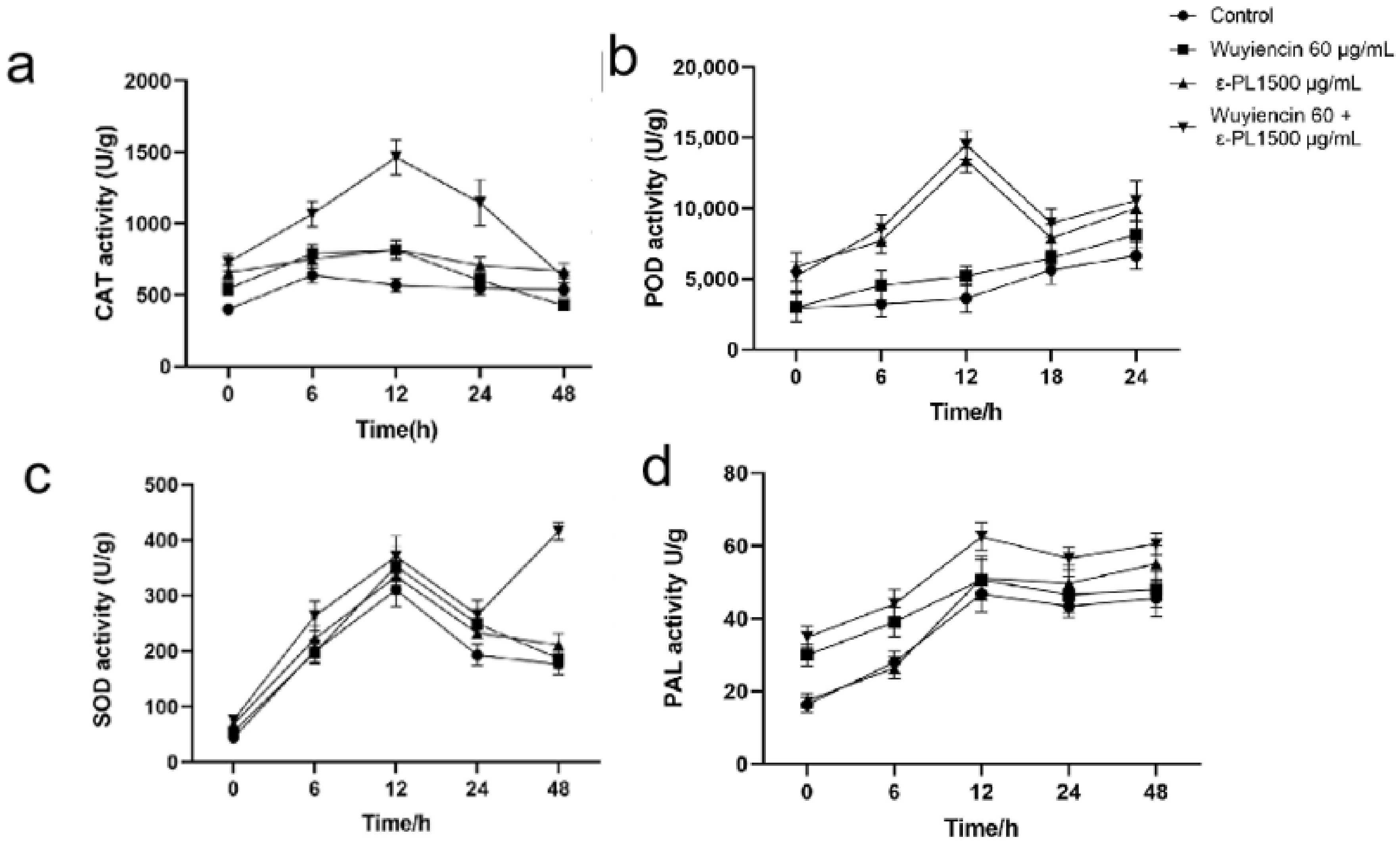
3.8. Defense-Related Enzyme Activities in Tomato
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L. Fungi vs. Fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.A. Biological control of plant diseases. Australas. Plant Pathol. 2017, 46, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Singh, H. Management of plant pathogens with microorganisms. Proc. Indian Natl. Sci. Acad. 2014, 80, 443–454. [Google Scholar] [CrossRef]
- Brimner, T.A.; Boland, G.J. A review of the non-target effects of fungi used to biologically control plant diseases. Agric. Ecosyst. Environ. 2003, 100, 3–16. [Google Scholar] [CrossRef]
- Najjar, M.B.; Kashtanov, D.; Chikindas, M.L. Natural antimicrobials ε-Poly-L-lysine and nisin a for control of oral microflora. Probiotics Antimicrob. Proteins 2009, 1, 143–147. [Google Scholar] [CrossRef]
- Shima, S.; Matsuoka, H.; Iwamoto, T. Antimicrobial action of ε-poly-L-lysine. J. Antibiot. 1984, 37, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Yao, H.; Xu, Z. Production of ε-poly-lysine by Streptomyces albulus PD-1 via solid-state fermentation. Bioresour. Technol. 2017, 223, 149–156. [Google Scholar] [CrossRef]
- Yoshida, T.; Nagasawa, T. ε-Poly-l-lysine: Microbial production, biodegradation and application potential. Appl. Microbiol. Biotechnol. 2003, 62, 21–26. [Google Scholar] [CrossRef]
- Chang, Y.; Mclandsborough, L.; Mcclements, D.J. Physicochemical properties and antimicrobial efficacy of electrostatic complexes based on cationic ε-Polylysine and anionic pectin. J. Agric. Food Chem. 2011, 59, 6776–6782. [Google Scholar] [CrossRef]
- Liu, K.; Zhou, X.; Fu, M. Inhibiting effects of epsilon-poly-lysine (ε-PL) on Pencillium digitatum and its involved mechanism. Postharvest Biol. Technol. 2017, 123, 94–101. [Google Scholar] [CrossRef]
- Wei, M.; Ge, Y.; Li, C. Antifungal activity of ε-poly-L-lysine on Trichothecium roseum in vitro and its mechanisms. Physiol. Mol. Plant Pathol. 2018, 103, 23–27. [Google Scholar] [CrossRef]
- Liu, H.; Chen, J.; Xia, Z. Effects of ε-poly-l-lysine on vegetative growth, pathogenicity and gene expression of Alternaria alternata infecting Nicotiana tabacum. Pestic. Biochem. Physiol. 2020, 163, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, X.; Yu, M. Transcriptomic and functional analyses indicate novel anti-viral mode of actions on tobacco mosaic virus of a microbial natural product ε-Poly-L-lysine. J. Agric. Food Chem. 2021, 69, 2076–2086. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.; Liu, X.; Chen, Q. Epsilon-poly-L-lysine (ε-PL) exhibits antifungal activity in vivo and in vitro against Botrytis cinerea and mechanism involved. Postharvest Biol. Technol. 2020, 168, 111270. [Google Scholar] [CrossRef]
- Sun, G.; Yang, Q.; Zhang, A. Synergistic effect of the combined bio-fungicides ε-poly- l -lysine and chitooligosaccharide in controlling grey mould (Botrytis cinerea) in tomatoes. Int. J. Food Microbiol. 2018, 276, 46–53. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, W.; Lv, Z. Evaluation of the inhibitory effects of wuyiencin, a secondary metabolite of streptomyces albulus CK-15, against Sclerotinia sclerotiorum In vitro. Plant Dis. 2022, 106, 156–164. [Google Scholar] [CrossRef]
- Romero, D.; De Vicente, A.; Olmos, J.L. Effect of lipopeptides of antagonistic strains of Bacillus subtilis on the morphology and ultrastructure of the cucurbit fungal pathogen Podosphaera fusca. J. Appl. Microbiol. 2007, 103, 969–976. [Google Scholar] [CrossRef]
- Shalit, I.; Shadkchan, Y.; Samra, Z. In vitro synergy of caspofungin and itraconazole against Aspergillus spp.: MIC versus minimal effective concentration end points. Antimicrob. Agents Chemother. 2003, 47, 1416–1418. [Google Scholar] [CrossRef] [Green Version]
- Righini, H.; Somma, A.; Cetrullo, S. Inhibitory activity of aqueous extracts from Anabaena minutissima, Ecklonia maxima and Jania adhaerens on the cucumber powdery mildew pathogen in vitro and in vivo. J. Appl. Phycol. 2020, 32, 3363–3375. [Google Scholar] [CrossRef]
- Pokora, W.; Reszka, J.; Tukaj, Z. Activities of superoxide dismutase (SOD) isoforms during growth of Scenedesmus (Chlorophyta) species and strains grown in batch-cultures. Acta Physiol. Plant. 2003, 25, 375–384. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, S.; Wang, C. Effects of Pb on the oxidative stress and antioxidant response in a Pb bioaccumulator plant Vallisneria natans. Ecotoxicol. Environ. Saf. 2012, 78, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; Hua, G.G.; Liu, H.X. Analysis of defence enzymes induced by antagonistic bacterium Bacillus subtilis strain AR12 towards Ralstonia solanacearum in tomato. Ann. Microbiol. 2008, 58, 573–578. [Google Scholar] [CrossRef]
- Dai, X.; An, J.; Wang, Y. Antibacterial amphiphiles based on ε-polylysine: Synthesis, mechanism of action, and cytotoxicity. RSC Adv. 2015, 5, 69325–69333. [Google Scholar] [CrossRef]
- Geng, W.; Yang, C.; Gu, Y. Cloning of ε-poly-L-lysine (ε-PL) synthetase gene from a newly isolated ε-PL-producing Streptomyces albulus NK 660 and its heterologous expression in Streptomyces lividans. Microb. Biotechnol. 2014, 7, 155–164. [Google Scholar] [CrossRef]
- Sun, G.; Wang, H.; Shi, B. Control efficiency and expressions of resistance genes in tomato plants treated with ε-poly-L-lysine against Botrytis cinerea. Pestic. Biochem. Physiol. 2017, 143, 191–198. [Google Scholar] [CrossRef]
- Zhou, T.; Liu, H.; Huang, Y. ε-poly-L-lysine affects the vegetative growth, pathogenicity and expression regulation of necrotrophic pathogen Sclerotinia sclerotiorum and Botrytis cinerea. J. Fungi 2021, 7, 821. [Google Scholar] [CrossRef]
- Bi, L.; Yang, L.; Bhunia, A.K. Emulsion stabilized with phytoglycogen octenyl succinate prolongs the antimicrobial efficacy of ε-poly-l-lysine against Escherichia coli O157: H7. LWT 2016, 70, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Liu, M.; Du, L. Synergistic antibacterial effect of the combination of epsilon-polylysine and nisin against Enterococcus faecalis. J. Food Prot. 2015, 78, 2200–2206. [Google Scholar] [CrossRef]
- Ning, H.Q.; Lin, H.; Wang, J.X. Synergistic effects of endolysin Lysqdvp001 and ε-poly-lysine in controlling Vibrio parahaemolyticus and its biofilms. Int. J. Food Microbiol. 2021, 343, 109112. [Google Scholar] [CrossRef]
- Zhang, Z.; Xia, G.; Yang, Q. Effects of chitosan-based coatings on storage quality of Chinese shrimp. Food Sci. Nutr. 2019, 7, 4085–4095. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Zhang, K.; She, G. Antimicrobial activity investigation on Wuyiencin fractions of different polarity. Molecules 2010, 15, 3179–3185. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.A.; Jones, J.D.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; El-Shetehy, M.; Shine, M.B. Free radicals mediate systemic acquired resistance. Cell Rep. 2014, 7, 348–355. [Google Scholar] [CrossRef]
- Choi, H.W.; Kim, Y.J.; Lee, S.C. Hydrogen peroxide generation by the pepper extracellular peroxidase CaPO2 activates local and systemic cell death and defense response to bacterial pathogens. Plant Physiol. 2007, 145, 890–904. [Google Scholar] [CrossRef] [Green Version]
- Hernández, J.A.; Gullner, G.; Clemente-Moreno, M.J. Oxidative stress and antioxidative responses in plant–virus interactions. Physiol. Mol. Plant Pathol. 2016, 94, 134–148. [Google Scholar] [CrossRef] [Green Version]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Xu, J.; Yang, J.; Duan, X. Increased expression of native cytosolic Cu/Zn superoxide dismutase and ascorbate peroxidase improves tolerance to oxidative and chilling stresses in cassava (Manihot esculenta Crantz). BMC Plant Biol. 2014, 14, 208. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Zhang, J.H.; Jiang, L.H.; Sun, C. Indole-3-acetic acid inhibits blue mold rot by inducing resistance in pear fruit wounds. Sci. Hortic. 2018, 231, 227–232. [Google Scholar] [CrossRef]
- Ngadze, E.; Icishahayo, D.; Coutinho, T.A. Role of polyphenol oxidase, peroxidase, phenylalanine ammonia lyase, chlorogenic acid, and total soluble phenols in resistance of potatoes to soft rot. Plant Dis. 2012, 96, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Ralton, J.E.; Howlett, B.J.; Clarke, A.E. Interaction of cowpea with Phytophthora vignae: Inheritance of resistance and production of phenylalanine ammonia-lyase as a resistance response. Physiol. Mol. Plant Pathol. 1988, 32, 89–103. [Google Scholar] [CrossRef]
- John, N.; Anjanadevi, I.; Suja, S. Biochemical changes induced in Amorphophallus in response to treatment with biocontrol agent and pathogen. Int. J. Biotechnol. Biochem. 2014, 10, 35–45. [Google Scholar]
- Christopher, D.J.; Raj, T.S.; Rani, S.U. Role of defense enzymes activity in tomato as induced by Trichoderma virens against Fusarium wilt caused by Fusarium oxysporum f sp. lycopersici. J. Biopestic. 2010, 3, 158–162. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogenic Fungi | Concertation of ε-PL (µg/mL) | Inhibition Rate (%) | Toxicity Equation | Correlation Coefficient | EC50 (µg/mL) |
|---|---|---|---|---|---|
| Physalospora piricola | 100 | 1.92 ± 0.33 e | y = 4.2465x − 6.1396 | R2 = 0.9115 | 419.99 |
| 300 | 16.35 ± 2.07 d | ||||
| 500 | 45.4 ± 3.18 c | ||||
| 1000 | 83.72 ± 6.29 b | ||||
| 2000 | 100 ± 0.00 a | ||||
| Rhizopus stolonifer | 100 | 0.63 ± 0.08 e | y = 4.6286x − 7.1749 | R2 = 0.9586 | 426.94 |
| 300 | 16.98 ± 2.34 d | ||||
| 500 | 50.44 ± 6.29 c | ||||
| 1000 | 89.31 ± 5.78 b | ||||
| 2000 | 100 ± 0.00 a | ||||
| Botrytis cinerea | 100 | 3.44 ± 0.47 d | y = 3.9943x − 5.0512 | R2 = 0.9393 | 328.39 |
| 300 | 36.5 ± 2.83 c | ||||
| 500 | 80.38 ± 5.48 b | ||||
| 1000 | 88.13 ± 7.29 b | ||||
| 2000 | 100 ± 0.00 a | ||||
| Fulvia fulva | 100 | 3.78 ± 0.63 e | y = 2.7349x − 2.0522 | R2 = 0.9789 | 378.96 |
| 300 | 48.14 ± 3.76 d | ||||
| 500 | 69.76 ± 5.29 c | ||||
| 1000 | 83.51 ± 6.68 b | ||||
| 2000 | 97.25 ± 3.29 a | ||||
| Sclerotinia sclerotiorum | 100 | 26.06 ± 2.24 b | y = 2.468x − 1.8093 | R2 = 0.9583 | 574.16 |
| 300 | 38.97 ± 2.83 a | ||||
| 500 | 38.97 ± 3.29 a | ||||
| 1000 | 43.23 ± 3.17 a | ||||
| 2000 | 43.28 ± 4.29 a | ||||
| Gibberella sanbinetti | 100 | 29.21 ± 2.13 d | y = 0.4513x + 3.5151 | R2 = 0.9207 | 1951.07 |
| 300 | 55.93 ± 6.83 c | ||||
| 500 | 70.59 ± 6.28 b | ||||
| 1000 | 83.35 ± 7.81 a | ||||
| 2000 | 91.23 ± 5.28 a | ||||
| Phytophthora infestans | 100 | 3.44 ± 0.59 d | y = 1.4789x + 1.5069 | R2 = 0.9979 | 430.12 |
| 300 | 18.44 ± 2.33 c | ||||
| 500 | 32.25 ± 2.39 b | ||||
| 1000 | 52.75 ± 4.74 a | ||||
| 2000 | 53.75 ± 4.29 a | ||||
| Colletotrichum lagenarium | 100 | 29.21 ± 2.13 d | y = 1.5575x + 0.1975 | R2 = 0.9454 | 1211.90 |
| 300 | 55.93 ± 6.83 c | ||||
| 500 | 70.59 ± 6.28 b | ||||
| 1000 | 83.35 ± 7.81 a | ||||
| 2000 | 91.23 ± 5.28 a | ||||
| Valsa mali | 100 | 2.22 ± 0.38 e | y = 4.1847x − 5.6559 | R2 = 0.9427 | 351.88 |
| 300 | 27.22 ± 1.38 d | ||||
| 500 | 80.13 ± 6.79 c | ||||
| 1000 | 87.97 ± 5.32 b | ||||
| 2000 | 100 ± 0.00 a | ||||
| Alteraria alternata | 100 | 10.3 ± 1.38 c | y = 2.4598x − 1.3359 | R2 = 0.9304 | 376.51 |
| 300 | 26.47 ± 3.29 b | ||||
| 500 | 75.29 ± 6.27 a | ||||
| 1000 | 79.41 ± 5.38 a | ||||
| 2000 | 83.17 ± 6.29 a | ||||
| Helminthosporium maydis | 100 | 2.91 ± 0.52 d | y = 4.1028x − 5.8397 | R2 = 0.8485 | 438.56 |
| 300 | 7.27 ± 0.81 d | ||||
| 500 | 55.75 ± 4.29 c | ||||
| 1000 | 71.93 ± 3.89 b | ||||
| 2000 | 100 ± 0.00 a | ||||
| Phomopsis asparagi | 100 | 3.29 ± 0.43 a | y = 1.0702x + 1.2992 | R2 = 0.8164 | 2871.08 |
| 300 | 22.63 ± 2.11 a | ||||
| 500 | 28.03 ± 1.39 a | ||||
| 1000 | 28.68 ± 1.93 a | ||||
| 2000 | 36.45 ± 1.29 a |
| Concentration (µg/mL) | Colony Diameter (cm) | Inhibition Rate (%) | |
|---|---|---|---|
| ε-PL | Wuyiencin | ||
| 0 | 0 | 7.73 ± 0.64 d | - |
| 12.5 | 6.62 ± 0.55 c | 14.36 ± 7.14 c | |
| 25 | 4.73 ± 0.39 b | 38.81 ± 5.10 b | |
| 50 | 0.92 ± 0.07 a | 88.10 ± 1.00 a | |
| 100 | 0 | 7.70 ± 0.64 d | 0.39 ± 0.22 d |
| 12.5 | 6.07 ± 0.51 c | 21.47 ± 3.78 c | |
| 25 | 3.80 ± 0.32 b | 50.84 ± 2.37 b | |
| 50 | 0.92 ± 0.08 a | 88.10 ± 0.57 a | |
| 200 | 0 | 6.20 ± 0.52 c | 19.79 ± 6.68 c |
| 12.5 | 6.05 ± 0.50 c | 21.73 ± 6.52 c | |
| 25 | 3.47 ± 0.29 b | 55.50 ± 3.74 b | |
| 50 | 0.37 ± 0.17 a | 95.21 ± 0.40 a | |
| 300 | 0 | 5.33 ± 0.70 d | 31.05 ± 5.75 d |
| 12.5 | 4.08 ± 0.20 c | 47.22 ± 4.40 c | |
| 25 | 2.07 ± 0.10 b | 73.22 ± 2.23 b | |
| 50 | 0.30 ± 0.15 a | 96.12 ± 3.23 a | |
| 400 | 0 | 3.20 ± 0.27 d | 58.60 ± 3.45 d |
| 12.5 | 2.78 ± 0.23 c | 64.04 ± 3.00 c | |
| 25 | 1.32 ± 0.11 b | 82.92 ± 1.42 b | |
| 50 | 0.18 ± 0.02 a | 97.67 ± 0.19 a | |
| 500 | 0 | 2.23 ± 0.19 c | 71.15 ± 2.40 |
| 12.5 | 2.02 ± 0.10 b | 73.87 ± 2.18 c | |
| 25 | 0.95 ± 0.79 a | 87.71 ± 5.91 b | |
| 50 | 0.00 ± 0.00 a | 100 ± 0.00 a | |
| Treatment | Concentration (μg/mL) | Germination Rate (%) | Inhibition Rate (%) |
|---|---|---|---|
| Control | - | 51.00 ± 2.50 | - |
| ε-PL | 50 | 47.00 ± 3.00 f | 7.84 ± 3.39 f |
| 100 | 26.00 ± 2.00 e | 49.02 ± 3.92 e | |
| 200 | 21.00 ± 2.00 d | 58.82 ± 3.92 d | |
| 300 | 15.00 ± 1.00 c | 70.59 ± 1.96 c | |
| 400 | 10.00 ± 1.50 b | 80.39 ± 2.94 b | |
| 500 | 4.00 ± 1.00 a | 92.16 ± 1.96 a | |
| wuyiencin | 10 | 42.00 ± 2.00 f | 17.65 ± 3.92 f |
| 20 | 35.00 ± 2.50 ef | 31.37 ± 4.90 e | |
| 40 | 30.00 ± 3.00 de | 41.18 ± 5.89 d | |
| 60 | 12.00 ± 1.00 cd | 76.47 ± 2.94 c | |
| 80 | 8.00 ± 1.50 bc | 84.31 ± 1.96 b | |
| 120 | 2.00 ± 0.50 ab | 96.08 ± 0.98 a | |
| ε-PL + wuyiencin | 200 + 10 | 17.50 ± 1.00 a | 65.68 ± 1.96 d |
| 200 + 20 | 11.00 ± 1.50 c | 80.00 ± 2.94 c | |
| 200 + 40 | 6.00 ± 1.00 b | 88.23 ± 1.96 b | |
| 200 + 60 | 0.00 ± 0.00 a | 100 ± 0.00 a | |
| 200 + 80 | 0.00 ± 0.00 a | 100 ± 0.00 a | |
| 200 + 120 | 0.00 ± 0.00 a | 100 ± 0.00 a |
| Treatment (μg/mL) | Colony Diameter (cm) | Inhibition Rate(%) |
|---|---|---|
| Control | 1.20 ± 0.20 a | - |
| Wuyiencin 60 | 0.73 ± 0.07 b | 38.89 ± 6.11 c |
| ε-PL 1500 | 0.33 ± 0.05 c | 72.22 ± 4.17 b |
| Wuyiencin 60 + ε-PL 1500 | 0.13 ± 0.02 d | 88.89 ± 1.67 a |
| ε-PL 2000 | 0.46 ± 0.06 cd | 61.67 ± 10.93 ab |
| Wuyiencin 60 + ε-PL 2000 | 0.20 ± 0.03 cd | 83.33 ± 1.67 ab |
| ε-PL 2500 | 0.60 ± 0.00 b | 50.00 ± 0.00 c |
| Wuyiencin 60 + ε-PL 2500 | 0.67 ± 0.05 b | 44.44 ± 4.17 c |
| ε-PL 3000 | 0.65 ± 0.04 b | 45.44 ± 3.33 c |
| Wuyiencin 60 + ε-PL 3000 | 0.63 ± 0.10 b | 47.78 ± 0.83 c |
| Treatment | MIC(μg/mL) | FIC | FICI | |
|---|---|---|---|---|
| Wuyiencin | ε-PL | |||
| Wuyiencin (alone) | 40 | 0.13 | 0.33 | 0.46 |
| ε-PL (alone) | 30 | |||
| Wuyiencin (in combination) | 5 | |||
| ε-PL (in combination) | 10 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Z.; Lu, Y.; Li, B.; Shi, L.; Zhang, K.; Ge, B. Effects of ε-Poly-L-Lysine Combined with Wuyiencin as a Bio-Fungicide against Botryris cinerea. Microorganisms 2022, 10, 971. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050971
Lv Z, Lu Y, Li B, Shi L, Zhang K, Ge B. Effects of ε-Poly-L-Lysine Combined with Wuyiencin as a Bio-Fungicide against Botryris cinerea. Microorganisms. 2022; 10(5):971. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050971
Chicago/Turabian StyleLv, Zhaoyang, Yanxuan Lu, Boya Li, Liming Shi, Kecheng Zhang, and Beibei Ge. 2022. "Effects of ε-Poly-L-Lysine Combined with Wuyiencin as a Bio-Fungicide against Botryris cinerea" Microorganisms 10, no. 5: 971. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050971