
Root-Associated Bacteria Are Biocontrol Agents for Multiple Plant Pests


1
Agricultural Solutions, BASF Korea Ltd., Seoul 04518, Korea
2
Department of Biological Engineering, Utah State University, Logan, UT 84322, USA
3
Department of Applied Biology, College of Agriculture and Life Sciences, Chonnam National University, Gwangju 61186, Korea
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(5), 1053; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051053
Submission received: 23 April 2022
/
Revised: 15 May 2022
/
Accepted: 16 May 2022
/
Published: 19 May 2022
(This article belongs to the Special Issue Microorganisms as Biocontrol Agents)
Abstract
:Biological control is an important process for sustainable plant production, and this trait is found in many plant-associated microbes. This study reviews microbes that could be formulated into pesticides active against various microbial plant pathogens as well as damaging insects or nematodes. The focus is on the beneficial microbes that colonize the rhizosphere where, through various mechanisms, they promote healthy plant growth. Although these microbes have adapted to cohabit root tissues without causing disease, they are pathogenic to plant pathogens, including microbes, insects, and nematodes. The cocktail of metabolites released from the beneficial strains inhibits the growth of certain bacterial and fungal plant pathogens and participates in insect and nematode toxicity. There is a reinforcement of plant health through the systemic induction of defenses against pathogen attack and abiotic stress in the plant; metabolites in the beneficial microbial cocktail function in triggering the plant defenses. The review discusses a wide range of metabolites involved in plant protection through biocontrol in the rhizosphere. The focus is on the beneficial firmicutes and pseudomonads, because of the extensive studies with these isolates. The review evaluates how culture conditions can be optimized to provide formulations containing the preformed active metabolites for rapid control, with or without viable microbial cells as plant inocula, to boost plant productivity in field situations.
1. Introduction
Globally, there is a strong movement for sustainable and regenerative agriculture, where plant quality, yield, and soil health are key factors. Additionally, agricultural methods for disease control are under scrutiny by consumers. These factors increasingly favor biopesticides as an alternative to a strict synthetic chemical approach to pest control. The success of chemical pesticides reveals the high standards for efficacy that has to be matched by biopesticides. Chemical pesticides are formulated to consistently control plant diseases and pests at a relatively low cost to the user. The chemicals are often targeted toward one or a group of related pests. However, the use of these chemical pesticides poses risks such as environmental contamination with residues, development of resistance to the pesticide in the targets, and impacts on human health [1]. Regulation of the use of chemical pesticides is constantly changing, and it is challenging to develop and register new compounds [2]. Furthermore, because the industry has provided pesticides with specific targets for either microbial plant pathogens, insects, or nematodes, they are not broad-spectrum. Therefore, different applications, possibly at separate times, are required for complete plant protection. Multiple applications increase costs to the farmer. One recent exception is the synthetic pesticide fluopyram, commercialized as both a fungicide and nematicide, that has been well received by growers for its reliable performance [3]. Fluopyram is a succinate dehydrogenase inhibitor that targets various plant fungal pathogens and kills pathogenic nematodes [3,4]. However, there are nontarget effects at higher concentrations, including the induction of mammalian tumors [5].
Successful biopesticides will protect the plants while maintaining crop quality and yield, and nurture soil health while minimizing their hazardous effects on consumers and the soil environment. However, application of biopesticides in agricultural fields has drawbacks that research involved in formulation, storage, and applications must overcome [6]. In the field, the survival and efficacy of microbial inocula may be limited by environmental factors [7]. For example, agricultural chemicals may negatively influence the activity and survival of beneficial microbes in the field. The formulation of current chemical pesticides maximizes their efficacy through the development of a stable shelf life and optimal methods of field application. A long shelf life and stability under storage are problems for live cell formulations. Compared with the rapid killing effects of some pesticides, time is required to amass beneficial microbial communities in the rhizosphere. The early observations of biological control in the field of the take-all fungus, Gaeumannomyces graminis, on cereals demonstrate how time and repetitive cropping are required to establish a suppressive soil effective for this pathogen [8].
Laboratory studies have firmly established that certain plant-associated microbes afford biocontrol of bacterial and fungal plant pathogens as well as plant-consuming insects and nematodes. Table 1 lists some of the plant-associated microbes where the microbial metabolites active in biocontrol of multiple plant pests are characterized. Formulations of such biocontrol agents are attractive because they could simultaneously deter microbial disease and insect/nematode damage to plants. Although endophytic fungal entomopathogens also limit microbial disease pressure in plants, this review focuses only on biocontrol bacteria [9,10].
The review is structured into four information sections. Section 2 introduces major bacterial genera and species for which biocontrol of multiple species by characterized metabolites are established. The different classes of biocides characterized from these bacteria are introduced and reveal that some active metabolites are shared between genera. The findings illustrate widespread variability in the susceptibility of the targets, something that is true for both different plant fungal pathogens and insect predators. Section 3 focuses on how certain microbial metabolites with direct biocontrol activity also induce systemic tolerance in the plant to pathogens, damaging insects, or nematode pests. Section 4 addresses the integration of the academic knowledge of biocontrol with industrial expertise and mechanization for scale-up to formulate commercial preparations for field applications as biopesticides. The conclusions section summarizes the highlights of the review, showing the potential for biocontrol of multiple targets by rhizosphere microbes.

{kind=link}
{kind=link}
Table 1.
Rhizosphere bacteria that protect plants against multiple pests (i.e., microbes, insects, and nematodes) through characterized metabolites and enzymes.
Table 1.
Rhizosphere bacteria that protect plants against multiple pests (i.e., microbes, insects, and nematodes) through characterized metabolites and enzymes.
| Bacterial Strains | Pathogens | Pests | Active Metabolites | Reference | |
|---|---|---|---|---|---|
| Antimicrobial | Insecticidal | ||||
| Gram-positive bacteria | |||||
| Bacillus amyloliquefaciens AG1 | Various fungal pathogens | Tuta absoluta | Crude protein extract | Biosurfactant | [11,12] |
| Bacillus atrophaeus L193 | Botrytis cinerea, Monilinia laxa | Rhopalosiphum padi | 2,3-Butanediol | Biosurfactant | [13,14] |
| Bacillus subtilis PTB185 | Various fungal pathogens | Aulacorthum solani Aphis gossypii | Lipopeptides | Chitinase | [15,16] |
| Bacillus subtilis SPB1 | Fusarium solani Rhizoctonia bataticola Rhizoctonia solani | Ectomyelois ceratoniae Spodoptera littoralis | Biosurfactant | Biosurfactant | [17,18,19,20] |
| Bacillus subtilis V26 | Botrytis cinerea | Tuta absoluta | Biosurfactant | Biosurfactant | [21] |
| Bacillus thuringiensis strains | Sclerotinia sclerotiorum | Plutella xylostella | Salicylic acid, ethylene, and jasmonic acid | BT toxin | [22] |
| Bacillus thuringiensis CMB26 | Colletotrichum gloeosporioides | Pieris rapae crucivora | Lipopeptide | Lipopeptide | [23] |
| Brevibacillus laterosporus Lak1210 | Fusarium equiseti | Plutella xylostella | Chitinase | Chitinase | [24] |
| Paenibacillus elgii HOA73 | Botrytis cinerea Cladosporiumsphaerospermum | Plutella xylostella Meloidogyne incognita | Chitinase | Crude enzyme, gelatinase, chitinase | [25,26,27] |
| Paenibacillus elgii HOA73 | Botrytis cinerea, Rhizoctonia solani, Fusarium oxysporum f. sp. lycopersici | Methyl-2,3 dihydroxybenzoate (phenolic compound) | [28] | ||
| Paenibacillus elgii HOA73 | Botrytis cinerea, Rhizoctonia solani | Protocatechuic acid | [29] | ||
| Paenibacillus polymyxa BMP-11 | Various fungal pathogens | Tribolium castaneum | 1-Octen-3-ol benzothiazole | 1-Octen-3-ol benzothiazole | [30] |
| Streptomyces hydrogenans DH16 | Various fungal pathogens | Spodoptera litura Meloidogyne incognita | NA | NA | [31,32] |
| Streptomyces tanaschiensis | Saccharomyces sp. Penicillium sp. | Musca domestica Locusta migratoria | Flavensomycin | Flavensomycin | [33] |
| Gram-negative bacteria | |||||
| Photorhabdus temperata M1021 | Phytophthora capsica, Rhizoctonia solani, Corynespora cassiicola | Galleria mellonella | Benzaldehyde | Benzaldehyde | [34] |
| Pseudomonas fluorescens CHA0 | Pythium ultimum | Spodoptera littoralis Heliothis virescens Plutella xylostella Meloidogyne javanica M. incognita | Pyoluteorin, 2,4-DAPG * | Pyoluteorin, 2,4-DAPG extracellular protease, and insect toxin (Fit **) | [35,36,37,38] |
| Pseudomonas chlororaphis PCL1391 | Fusarium oxysporum f. sp. radicis-lycopersici | Spodoptera littoralis Heliothis virescens Plutella xylostella Galleria mellonella | Phenazine-1-carboxamide | Potent insect toxin, HCN, lipopeptide, Fit toxin | [38,39,40] |
| Pseudomonas chlororaphis O6 | Rhizoctonia solani Fusarium graminearum Phytophthora infestans | Meloidogyne hapla Myzus persicae | Pyrrolnitrin, phenazines | Hydrogen cyanide (HCN), Cyclic lipopeptides | [41,42,43,44] |
| Pseudomonas chlororaphis PA23 | Sclerotinia sclerotiorum | Caenorhabditis elegans | Pyrrolnitrin | Pyrrolnitrin, HCN | [45,46] |
| Serratia entomophila AB2 | Aspergillus flavus Candida albicans Fusarium oxysporum | Heliothis armigera | NA | NA | [47] |
* 2,4-DAPG: 2,4-diacetylphloroglucinol, ** Fit: for P. fluorescens insecticidal toxin.
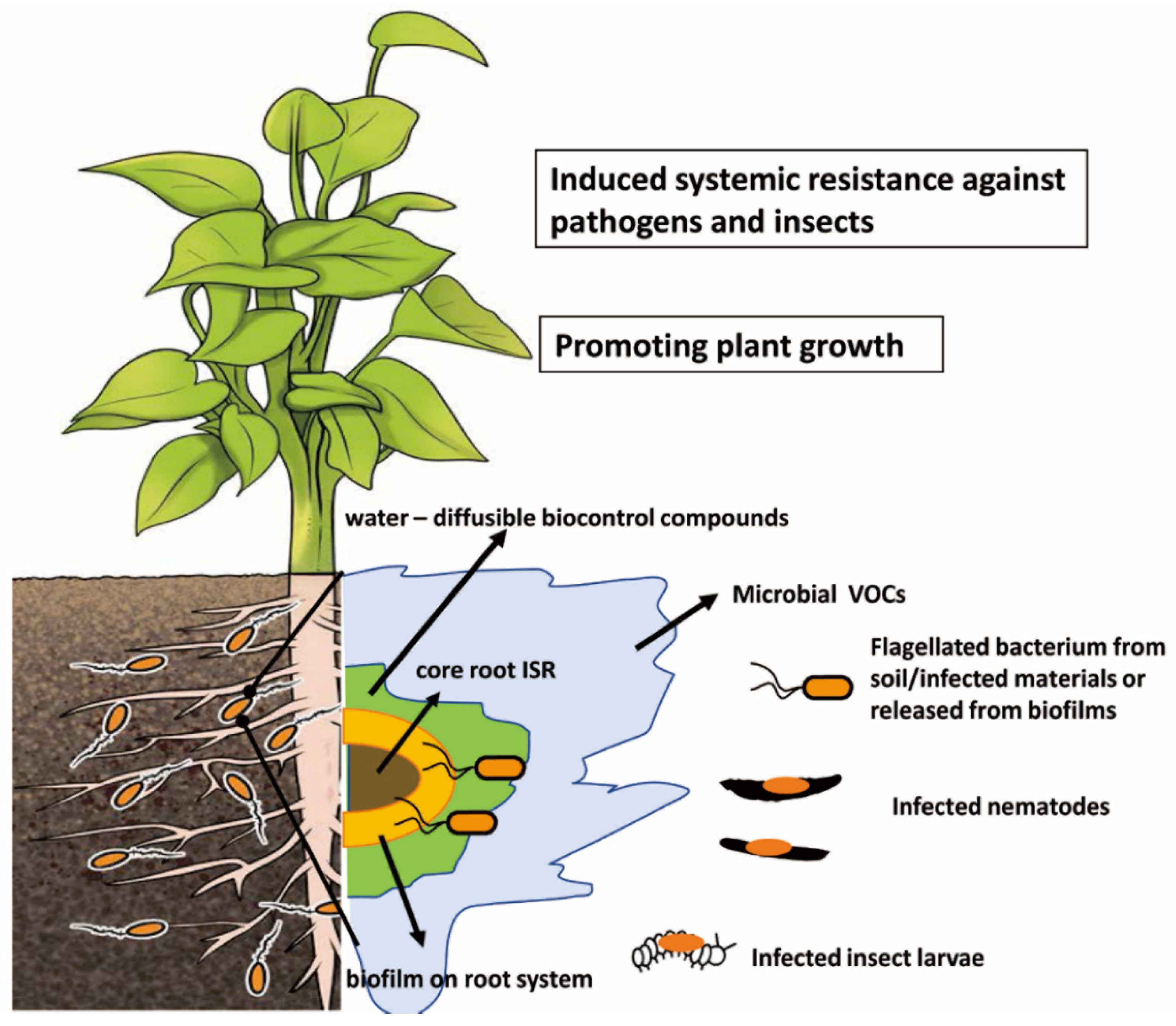
Targeted virulence against plant pests and other beneficial effects that promote plant vigor fits the description of these bacteria as probiotics, a term applied to microbes that improve the growth of their hosts [48,49,50]. The rhizosphere habitat of the beneficial microbes enables the plant to be surrounded by overlapping spheres of influence that depend on different mechanisms for plant protection. This concept is illustrated in Figure 1. Central and throughout the plant is the induction of systemic resistance through root colonization by certain beneficials. The formation of biofilms of the beneficial microbes in part physically protects the root surface. Protection is extended further into the rhizosphere through the secretion of antagonistic enzymes or metabolites by the biofilm cells. The production of antagonistic volatiles provides an even greater radius of protection.
2. The Array of Isolates and Their Products for Biocontrol of Multiple Targets
Bacteria with multiple biocontrol potential include genera classified as Gram-positive cells, such as the firmicutes Bacillus, Paenibacillus, and Brevibacillus; actinomycetes such as Streptomycete isolates; and Gram-negative isolates, including Pseudomonas, Photorhabdus, and Serratia (Table 1). For commercial formulation, Gram-positive isolates that sporulate are advantageous because the spores have an extended longevity over vegetative cells. Common habitats for these genera are soils. Many are documented to colonize root tissues, to which they are attracted by chemotaxis towards the gradient of plant root exudates. Certain isolates display very specific habitats, such as the symbiosis of Photorhabdus luminescens with entomopathogenic nematodes [51].
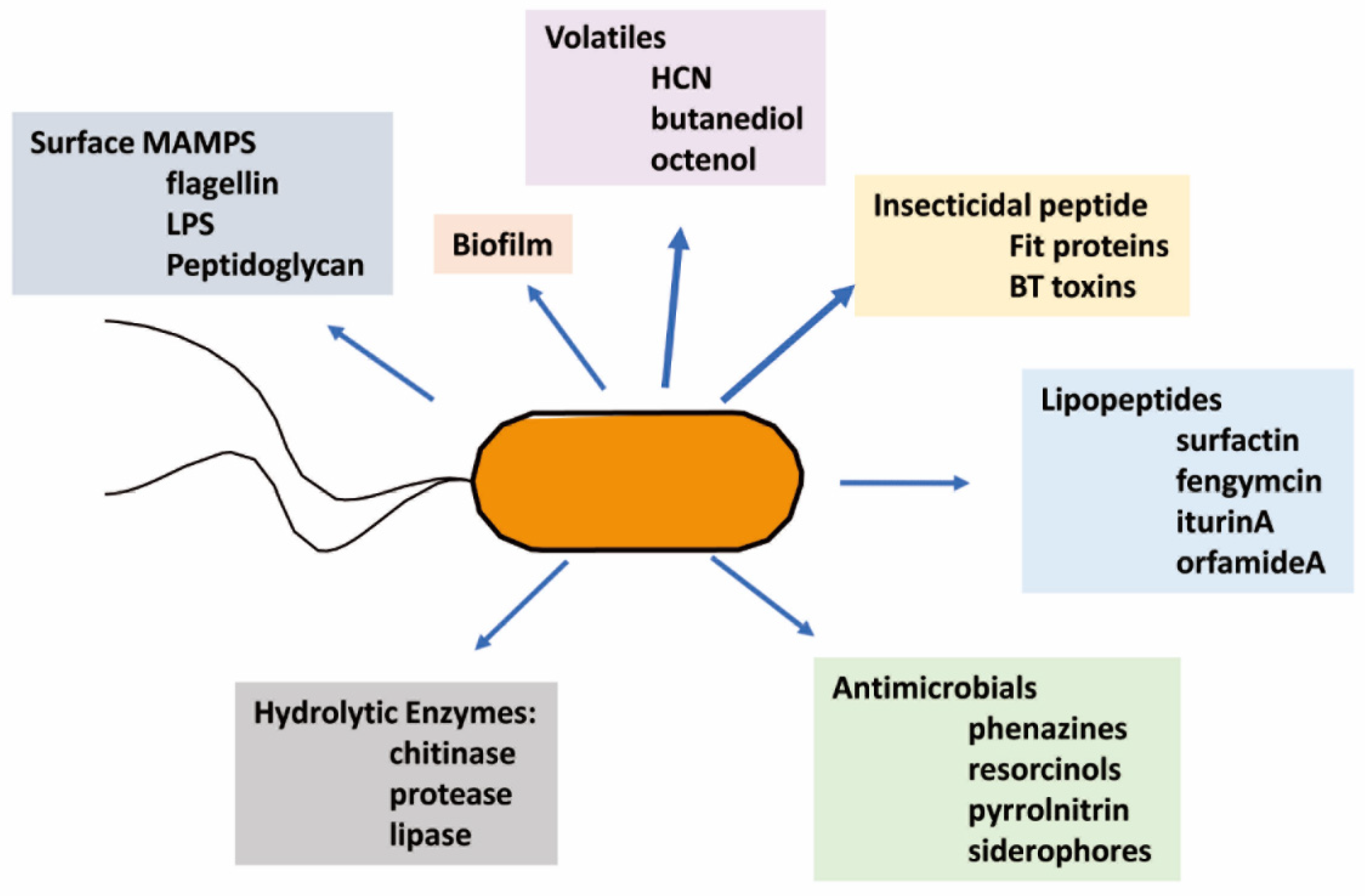
Biocontrol-active metabolites are diverse (Figure 2) but can be classified based on their structural similarity; for example, peptide toxins are implicated in insect and nematode control. Indeed, the most commercially relevant are the toxins produced by Bacillus thuringiensis, collectively termed BT toxins, which function by generating holes in the membranes of the insect’s digestive tract [52]. Fit proteins from the fit genes in the genomes of certain pseudomonads [53] and the related Mcf toxins from Photorhabdus and Xenorhabdus [54] that induce membrane disorganization also contribute to insecticidal activity. The lipopeptide group impacts membrane structures through their surfactant activity [55]. Phenolics such as the phenazine group may cause oxidative stress in the target leading to cell death, and some may act as iron chelators [56]. The hydrolytic enzymes chitinase, proteases, and lipases could damage cellular structures [37,57,58]. The combined activities of proteases and lipases could impair membrane structure. The chitinases could hydrolyze cell wall polymers in insects, nematode egg coats, and fungi [58].
The direct effects, revealed by dose-dependent in vitro studies with purified materials, generally require high active metabolite or enzyme levels. However, in addition to the potential direct effects of the above metabolites and secreted enzymes, there is also a strong involvement by the same active biocontrol product in activating systemic protection measures in the plant [59]. Characterized traits of the microbes and functions of the metabolites with potential biocontrol significance are summarized by type in Figure 2. The following discussion introduces the rhizosphere habitat for biocontrol agents and, then, with a focus on firmicutes and pseudomonads, the biocontrol potential of characterized metabolites produced by different isolates are compared.
2.1. The Rhizosphere Habitat: A Significant Trait of Microbes with Multiple Biocontrol Activities
The rhizosphere is a dynamic space and key to the health and productivity of the plant. Roots and their exudates supply nutrients to damaging microbial plant pathogens and a foothold for invasion by pathogenic nematodes and insect larvae. However, the plant counters these effects by nurturing root colonization by beneficial microbes that shift the balance towards root and shoot health. Firmicutes and Gamma-proteobacteria, such as the pseudomonads, are among the most-studied bacteria that are such probiotics for the plant [60]. Their consumption of simple metabolites, such as the sugars and amino acids in root exudates [60,61], reduces the ability of root exudation to support the growth of pathogenic microbes, insect larvae, and nematodes. Further, the microbe’s metabolism increases the level of protectant secondary microbial products in the soil pore waters. These protective metabolites may require specific catabolic mechanisms in order to have persistence and to influence microbial community composition [62]. One example by Stringlis et al. [63] shows how coumarins in the root exudates favor root colonization by biocontrol-active microbes with a tolerance to coumarin, whereas this root metabolite is toxic to pathogen growth. The pools of biocontrol-active metabolites will function to strengthen the niche of the beneficial microbes, and additionally may trigger systemically protective changes in the plant to further thwart potential pathogens.
The formation of biofilms by the probiotic cells on root cell surfaces is integral to the protection process [64]. The biofilm patches shield the root surface cells against direct attack and provide a sheltered environment for the cells of the beneficial bacterium. Additionally, the extracellular polymers forming the gel for the matrix encapsulating the cells could retain released microbial products, so that their concentrations are higher than in the soil pore water. When insect larvae and nematodes feed on the colonized plant root, they will ingest these biofilms that are loaded with toxic materials. Interestingly, both resorcinols and phenazines, identified as phenolics active in biocontrol, are correlated with improved biofilm formation [45,64,65]. The biofilm would also reduce the ingress of pathogenesis factors such as enzymes and toxins from any plant pathogens, so that contact with root cells is lessened.
Studies with a Paenibacillus polymyxa strain confirm that the matrix exopolysaccharides of the biofilm act as a rhizosphere nutrient source and as a structure that inhibits pathogen attack [66]. Chan et al. [67] indicate that reduced motility of nematodes within a Pseudomonas aeruginosa biofilm impairs their predatory behavior. Earlier work with the biocontrol agent Pseudomonas brassicacearum DF41 observed that biofilm formation over the head of the nematode contributes to the nematode’s demise [68]. This group cites the activity as being part of the mechanism that limits predation of the beneficial bacterium, i.e., it is part of the survival features of the biocontrol agent.
Invasion and feeding on root cells are part of the life cycle of the nematodes and insect larvae that damage plants. In this context, being able to kill these organisms would provide nutrients for the biocontrol organisms, thus augmenting the carbon-rich metabolites in the root exudates. A comparison of the root exudate composition shows that colonization of the roots by beneficial microbes alters the composition of amino acids [69,70,71]. Some amino acids presumably supply essential N for microbial growth, which is limited in the soil pore water. For instance, Boiteau et al. [70] found that the concentration of two N-containing amino acids in root exudates from Brachypodium decreases after colonization by Pseudomonas fluorescens. Microbes that are rhizosphere colonists have higher numbers of amino acid and sugar transporters than non-colonists [69]. Consequently, the presence of genes enabling the biocontrol agents to kill protein-rich larvae would be advantageous for bacterial multiplication, because of the rich supply of complex N-nutrients from the insect’s body mass [72,73]. Similarly, enzymatic digestion through proteases of the proteinaceous cuticle of nematodes or the chitin of their eggs would increase the N supply to the bacteria [74].
Infections with nematodes or insect larvae would result in biofilms on these hosts and the production of the biocontrol-active metabolites and enzymes, providing “hot spot” reservoirs additional to those at the plant root. Indeed, infection with insect larvae by biocontrol-active pseudomonads induces the expression of genes supporting antifungal metabolite production, boosting the value of such pockets of biocontrol microbes within the soil [40]. Further, any movement by the infected larvae and nematodes would aid in spreading the biocontrol agent (Figure 1). The concept that biocontrol microbes in the rhizosphere are spread by contact with and pathogenicity on insects is discussed well in a review by Pronk et al. [75]. Thus, the infectious ability on the soil fauna boosts the sphere of influence of the probiotics further from the root surface [60] (Figure 1). These potential roles by the biocontrol agents of the biofilm in plant protection add to the need to formulate viable cell preparations that are effective when introduced into agricultural settings.
2.2. Active Products from Bacillus spp.
Four classes of protectants essential in biocontrol by Bacillus spp. are peptide toxins, lipopeptides, enzymes, and volatile organic compounds. The most effective of these products are the insecticidal BT toxins, which have been successfully commercialized. The toxins are produced as sporulation of the Bacillus isolates commences and become concentrated as an inactive crystal structure within bacterial cells [52]. After ingestion of the crystals by the insect larvae, conversion to an active peptide dependent on the action of a larval protease occurs. The recognition of the active peptide by specific receptors on insect gut cells causes pore formation and loss of selective permeability. The efficacy of digestion is impaired, and the bacteria enter the circulatory systems to eventually kill the larvae. The specificity of the receptors in the gut cell membranes leads to products that target certain insects. The toxins vary in structure and activity; BT from B. thuringiensis shows an LC50 of 0.28 ppm against the second instar larvae of the diamondback moth Plutella xylostella, a value lower than the LC50 for the toxin from Bt subsp. kurstaki HD1 (LC50 0.47 ppm) [22]. However, BT toxins are not the only control mode, because additional toxic peptides can be formed even in vegetative cells [76]. Understanding the additive or synergistic effects of the array of metabolites in biocontrol requires further studies.
Lipopeptides are metabolites commonly secreted by B. thuringiensis and other Bacillus species [77]. Many lipopeptides have potent activity against insects and are also antifungal (Table 2). For example, the lipopeptide from B. thuringiensis CMB26 has insecticidal activity on the larvae of the cabbage white butterfly, Pieris rapae crucivora, and inhibits the growth of the pathogenic fungus, Colletotrichum gloeosporioides [23]. A single Bacillus spp. strain may produce several lipopeptides, each with a different potential against plant fungal pathogens. Two lipopeptides produced by B. subtilis EA-CB0015, iturin A and fengycin C, inhibit the anthracnose pathogen Colletotrichum acutatum, with a minimum inhibitory concentration (MIC) at 32 ppm and 124 ppm, respectively [78]. Other Bacillus species also produce iturin A, and this biocide also disrupts the membranes in cells of the fungal targets [79]. Studies of iturin from B. methylotrophicus TEB1 show a MIC of 100 ppm against Phoma tracheiphila [80], whereas iturin from B. amyloliquefaciens MG3 inhibits mycelial growth of C. gloeosporioides at concentrations less than 50 ppm but is ineffective in restricting spore germination [79]. These examples indicate that timing is crucial for fungal pathogen control on the plant, as it will depend on whether the spores or mycelial growth of a pathogen is targeted. Another lipopeptide from B. amyloliquefaciens BO5A inhibits mycelia of F. oxysporum at 10 ppm although there is no effect on the mycelium of Botrytis cinerea at 100 ppm [81].
Regarding insecticidal activity, a lipopeptide from B. amyloliquefaciens AG1 has an LC50 of 180 ng/cm2 against larvae of the leaf miner, Tuta absoluta, targeting the membranes in the larval midgut cells [12]. These same larvae are controlled at 278 ng/cm2 by the lipopeptide from B. subtilis V26, which is also an antifungal that halts mycelial growth of the gray mold fungus, B. cinerea, at 2 ppm [21].
The surfactin lipopolypeptide from B. subtilis Y9 shows aphicidal activity against the green peach aphid Myzus persicae at 20 ppm [82]. Similarly, active surfactant lipopeptides from Bacillus atrophaeus L193 cause the destruction of the cuticle of the aphid Rhopalosiphum padi [13]. B. atrophaeus L193 also controls fungal diseases caused by B. cinerea and Monilinia laxa in cherry fruits by producing 2,3,-butanediol, a weakly volatile metabolite [14]. Butanediol production is also observed in other plant-associated biocontrol bacteria [92], again showing the sharing of beneficial traits.
Other dual biocontrol lipopeptides synthesized by B. subtilis SPB1 kill larvae of the cotton leafworm Spodoptera littoralis, initially through the disruption of midgut function [17]. The lipopeptide also effectively controls two fungal pathogens of potato and tomato, Fusarium solani and Rhizoctonia solani, but at high MICs of 3000 ppm and 4000 ppm, respectively, compared with an MIC of 40 ppm for Rhizoctonia bataticola [20]. Of note is the finding that the B. subtilis SPB1 lipopeptides have preventative and curative effects [19]. These findings illustrate the strong potential for lipopeptides to combat plant loss due to insects and fungal pathogens, however, the sensitivities between targets are different.
Although the lipopeptide from B. subtilis PTB185 effectively controls the gray mold fungus B. cinerea, its chitinase is the major factor combating the aphids Aulacorthum solani and Aphis gossypii [15,16]. Chitinase of B. firmus also is highlighted as a seed treatment agent to control nematodes [93]. Several papers discuss the multiple roles of chitinase regarding its antifungal, insecticidal, and nematicidal activities [58,94]. However, chitinase is not implicated in the control of the nematode Meloidogyne incognita by a crude supernatant from B. firmus YBf-10. The efficacy in pot cultures is comparable to that of the chemical nematicide fenamiphos [95]. In this case, a serine protease and chemical metabolites seem to be functional in the nematicidal response [96].
Bacillus strains also produce biocontrol-active volatile organic compounds (VOCs). Two VOCs generated by B. velezensis CT32, 2,4-dimethyl-6-tert-butylphenol and benzothiazole, have strong antifungal activity against wilt in strawberries caused by Verticillium dahliae and F. oxysporum [97]. These VOCs are documented in the headspace when the bacillus is grown on nutrient agar. Unidentified VOCs produced by another B. velezensis strain, isolate VN10, inhibit mycelial growth of Sclerotinia sclerotium with high efficacy (MIC 1 ppm) and reduce the disease caused by this fungus through suppression of its production of oxalic acid [98].
2.3. Paenibacillus spp. Metabolites and Enzymes
The biocontrol arsenal of Paenibacillus elgii HOA73 displays the common finding that a multiplicity of metabolites formed by a single isolate is important in biocontrol. Hydrolytic enzymes (chitinase, protease, and gelatinase), along with the phenolic compounds benzothiazole, methyl 2,3-dihydroxybenzoate, protocatechuic acid, and the volatile 1-octen-3-ol, are among its biocontrol weapons. The phenolic compounds display promising MIC values for fungal pathogen control. Methyl 2,3-dihydroxybenzoate inhibits mycelial growth of B. cinerea, R. solani, and F. oxysporum f. sp. lycopersici at 32–64 ppm [28], and the MIC for protocatechuic acid is similar for B. cinerea and R. solani at 64 ppm. At 100 ppm, protocatechuic acid halts the disease progress of gray mold on strawberry fruits [29]. The lipopeptides from Paenibacillus strains also participate in biocontrol. A lipopeptide from P. polymyxa shows antimicrobial and anti-insect efficacy [66].
Active enzymes include chitinase, which inhibits spore germination of Cladosporium tenuissimum and B. cinerea at 100 ppm, whereas spores of Fulvia fulva and C. gloeosporioides are resilient [99]. The P. elgii HOA73 strain exhibits insecticidal and nematicidal performance correlating with chitinase and gelatinase (a type of protease) activities; mortality of M. incognita J2 juveniles and a reduction in egg mass and galling in tomatoes occur at 50–400 ppm. Other factors may participate in the nematicidal effects and the killing of second instar larvae of the diamondback moth, P. xylostella. Combining organic sulfur with the active enzymes acts synergistically [26]. It will be interesting to see how biocontrol microbes and their products can be combined with other tools to achieve better control.
VOCs from P. polymyxa BMP-11 are implicated in biocontrol. Two VOCs, 1-octen-3-ol (LC50, 16.75 ppm) and benzothiazole (LC50, 3.5 ppm), are active against the adult red flour beetle Tribolium castaneum [30]. In vitro work with authentic 1-octen-3-ol shows potent inhibition of bacteria and fungi at 1–2 ppm [100]. At 100 ppm, 1-octen-3-ol effectively inhibits mycelial growth of the brown rot fungus, Monilinia fructicola, but fruit treatment requires about 50 ppm to slow disease progress. Fumigation with 1-octen-3-ol (LD50, 27.7 mL/L air) reduces the effects of the maize weevil Sitophilus zeamais and the production of mycotoxin by the fungus Fusarium verticillioides (MIC 81.5 mL/L air) [101]. Citronellol, also produced by P. polymyxa strain BMP-11, inhibits F. oxysporum. Additionally, VOCs from P. polymyxa KM2501-1 can kill second-stage juveniles of M. incognita through mechanisms that include acting as a repellent and a fumigant active on contact [102].
These examples suggest that certain VOCs can be applied as fumigants. However, boosting the plant-associated bacteria in agricultural soils by inoculation would also promote beneficial volatile production in the rhizosphere. These methods could be valuable when raising crops in enclosed spaces, such as in greenhouse cultivation.
2.4. Pseudomonad Products
The weaponry of biocontrol pseudomonads uses the same metabolite classes discussed for the firmicutes in Section 2.2 and Section 2.3 for multiple control activity of nematodes, insect larvae, and microbial plant pathogens. Toxic peptides, lipopeptides, and volatiles are significant, as are several classes of phenolic-based structures (Figure 2).
Among the phenolic-based products with biocontrol activity, phenazines are well-known for their potential as antifungal compounds [103,104]. In general, phenazine-1-carboxylic-acid (PCA) has higher antifungal activity than phenazine-1-carboxamide (PCN) or 1-hydroxyphenazine (1HP) [105,106]. Phenazines of different structures can be produced simultaneously by pseudomonads, dependent on their gene pools [107].
The commercial product Shenqinmycin contains PCA as the major metabolite and has high efficacy in the field against Phoma infections [108]. The PCA produced by P. fluorescens 2–79 limits the mycelial growth of several plant pathogens; Cochliobolus sativus, G. graminis var. tritici, and R. solani are inhibited with MICs of 1 ppm, but Fusarium spp. requires 25–30 ppm. Species-dependent sensitivity is seen in the oomycete Pythium, with MIC values for PCA of 1 ppm for Pythium aristosporum, 25–30 ppm for Pythium ultimate, and 80–100 ppm for Pythium ultimum var. sporangiifurum [87].
Although mycelial exopolysaccharide production from B. cinerea is inhibited at 3 ppm PCA, the control of postharvest gray mold caused by this fungus requires 25 ppm [83]. The PCA synthesized by P. aeruginosa GC-B26 is active against Colletotrichum orbiculare as well as the oomycetes Pythium capsici (MIC 5 ppm) and P. ultimum (MIC 5 ppm) [84]. Studies with PCA from P. aeruginosa report activities against Sclerotium rolfsii (MIC 29 ppm), F. oxysporum (MIC 40 ppm), and Colletotrichum falcatum (MIC 50 ppm) [85]. The effective dose of PCA is often similar to the doses required for commercial pesticides, for example, for protection against Phytophthora blight on pepper and anthracnose on cucumber [84].
The control of tomato foot and root rot, caused by F. oxysporum sp. radices-lycopersici, with P. chlororaphis PCL1391 uses PCN as the major phenazine [39]. As for PCA, effective doses of PCN are similar to those of chemical products. Extracted PCN, produced by P. aeruginosa MML2212, effectively controls rice sheath blight caused by R. solani at 5 ppm, exceeding protection from the chemical fungicide carbendazim. Control of rice bacterial leaf blight, Xanthomonas oryzae pv. oryzae, by 5 ppm PCN is similar to that by the pesticide refamycin [89], and control of B. cinerea by PCN from P. aeruginosa at 108 ppm has almost the same inhibition rate as the chemical fungicide carbendazim. The effective dosages for PCN and chemical fungicides are higher for effects on spore germination and mycelial growth of Sclerotinia, requiring about 700 ppm for both PCN and carbendazim [88].
In contrast to their high efficacy for inhibiting various plant fungal pathogens, phenazines are not as efficient in suppressing insect or nematode larvae. They may impair egg hatching and promote increased J2 larval mortality of the root-knot nematode M. incognita [109,110]. Among the secondary metabolites produced by P. chlororaphis strain PA23, hydrogen cyanide (HCN) and the phenolic-based pyrrolnitrin (PRN) are cited as active agents in killing the nematode Caenorhabditis elegans, for which they also act as repellents. Growing P. chlororaphis strain PA23 with a nematode as the food source enhances the production of HCN and PRN [46]. However, mutational analysis of P. chlororaphis PA23 reveals that, among the secondary metabolites produced by this bacterium, pyrrolnitrin is the major metabolite involved in inhibiting S. sclerotiorum [45]. Studies with root-knot nematode juveniles found that HCN production is essential for the nematicidal effects of P. chlororaphis O6 [41,43].
Induced mortality of the root-knot nematode by P. fluorescens CHA0 requires another suite of products involving a protease and two phenolic-based structures, pyoluteorin and 2,4-diacetylphloroglucinol (2,4-DAPG), each with demonstrated antifungal activities [36,37]. The production of 2,4-DAPG by P. fluorescens CHA0 also suppresses populations of another nematode, Meloidogyne javanica [36], while 2,4-DAPG from P. fluorescens is toxic to Xiphinema americanum adults with an LC50 of 8.3 ppm. Apparently, 2,4-DAPG does not harm beneficial entomopathogenic nematodes [91].
The antifungal activity of 2,4-DAPG from pseudomonad isolates is well documented. It is effective at 50 ppm against the citrus postharvest fungi Penicillium digitatum and Penicillium italicum, a dose that compares favorably to 600 ppm for the chemical fungicide albesilate [111]. Production of 2,4-DAPG by P. fluorescens CHA0 is a major factor in suppressing the soil-borne disease black root rot in tobacco, caused by Thielaviopsis basicola, and against G. graminis var. tritici in wheat [90].
Other metabolites play roles in the insecticidal activities displayed by the multi-biocontrol pseudomonads [38,53]. Flury et al. [40] proposed that a mixture of the cyclic lipopeptide orfamide A and volatile HCN contributes to the insecticidal activity of P. fluorescens CHA0 and other pseudomonads [40]. Interestingly, the growth of P. fluorescens CHA0 on insect larvae promotes the expression of genes involved in producing an array of antifungal metabolites from this strain [40]. This finding suggests the to this rich nutrition of the bacterium helps its arsenal of metabolites provide security against competition with invasive microbes in the rhizosphere and bulk soil. Even though HCN is produced only at low levels by P. protegens Pf-5, the synthesis of analogs of rhizoxin adds to the power of orfamide A and a chitinase in the oral toxicity of this isolate to the larvae of the common fruit fly, Drosophila melanogaster [112].
Proteins termed Fit toxins are another part of the pathogenic mechanism of P. protegens and P. chlororaphis isolates on lepidopteran larvae [38,53,113]. Ruffner et al. [54] discussed how the fit gene clusters in the pseudomonads are related to those in two insect pathogens, Photorhabdus and Xenorhabdus, suggesting horizontal gene transfer of the clusters into limited pseudomonad genomes.
Hydrogen cyanide, HCN, is an important biocontrol volatile from the pseudomonads and other rhizobacteria including Bacillus isolates [114,115,116], contributing to a broad spectrum of inhibitory activities against microbes, insects, and nematodes [40,117]. Root exudates may exogenously supply the glycine that the microbes use as the substrate for HCN synthase; carbon dioxide is the other product of the synthase activity [117]. Production of HCN is observed in the airspace of closed growth boxes supporting plants with roots colonized by P. chlororaphis O6 [43,117]. Dependent on concentration, HCN may be toxic due to the disruption of cellular function through the inhibition of heme group function [118]. Indeed, root colonization and HCN production by certain isolates leads to herbicidal activity [119]. However, lower doses of HCN may also be important through synergistic interaction with other antimicrobial metabolites, such as lipopeptides or pyrrolnitrin, in enhancing the biocontrol effects.
The inhibition of fungal growth of the foliar plant pathogens Septoria tritici and Puccinia recondita f. sp. tritici on wheat is attributed to HCN production by P. putida BK8661 [120]. Furthermore, cyanide-producing pseudomonads suppress fungal diseases on canola and rice [121,122]. Mutation of P. fluorescens CHA0 revealed that HCN is a major player in controlling black root rot of tobacco, caused by the fungus Thielaviopsis basicola [123,124]. The combination of HCN and 2,4-DAPG syntheses by Pseudomonas sp. LBUM300 protects tomatoes against bacterial canker caused by Clavibacter michiganensis subsp. michiganensis [125].
Insecticidal effects of HCN production have been revealed by the observed mortality of second instar nymphs of the green peach aphid Myzus persicae when exposed to volatiles from P. chlororaphis O6 [44]. This study, and others with different pseudomonad isolates, showed that HCN production is controlled by quorum sensing [44]. Other studies extend the insecticidal effects of HCN produced by pseudomonads to flies and termites [126,127,128]. Combining HCN with lipopeptides of P. protegens CHA0 and P. chlororaphis PCL1391 shows activity against insects [40]. The nematicidal activities of HCN include killing of the nematode Caenorhabditis elegans by P. chlororaphis PA23, in combination with pyrrolnitrin [122]. A primary role for HCN is implicated in the mortality of J2 juveniles of the root-knot nematode, where for tomato, the efficacy of the biocontrol agent was as strong as that of the chemical nematicide fosthiazate in commercial greenhouses [41,43].
These studies illustrate that the biocontrol-active pseudomonads, like the firmicutes, strongly influence rhizosphere health, their strength being dependent upon a multiplicity of potential control mechanisms incited by different metabolites.
2.5. Streptomyces, Brevibacillus, Serratia, and Photorhabdus
As discussed above, secondary products from the firmicutes and pseudomonads produce many different structures with multiple biocontrol activities. Only a few highlights are provided in this section to illustrate the expansive diversity in structure and function. The Gram-positive Streptomyces hydrogenans DH16 secretes metabolites that kill M. incognita J2 juveniles at 100 ppm [32], inhibit fungal pathogens, and induce mortality to second instar larvae of Spodoptera litura [32]. A chitinase produced by Brevibacillus laterosporus Lak1210 has dual performance against larvae of the diamondback moth P. xylostella and the pathogenic fungi Fusarium equiseti [24].
For other Gram-negative isolates, the volatile benzaldehyde produced by Photorhabdus temperata M1021 contributes to the antimicrobial effects on Phytophthora capsici, R. solani, Corynespora cassiicola, and Bacillus spp., as well as insecticidal activity on the larvae of the greater wax moth Galleria mellonella [34].
Serratia species are a source of many deterrents for plant protection and represent an understudied resource for soil and plant associations [129]. Ordentlich et al. [130] reported that chitinase from Serratia marcescens is key for controlling Sclerotium rolfsii; direct lysis of mycelia of the pathogen was observed. Serratia entomophila AB2 shows a dual effect on pathogenic fungi and the pod borer Heliothis armigera [47]. Haterumalides, novel and complex metabolites from Serratia plymuthica A153, suppress the apothecial formation in sclerotia of Sclerotium with MIC of 0.5 ppm; additionally, oomycetes also are sensitive to these metabolites [131,132]. Serratia isolates are amongst the several bacterial genera that synthesize pyrrolnitrin, previously discussed as a pseudomonad product in Section 2. In combination, pyrrolnitrin and the haterumalides inhibit spore germination of various fungal plant pathogens at MICs of 0.4–50 ppm [131]. The array of volatiles synthesized from Serratia isolates also offers biological control potential [133]. Recent work on the plant-growth-promoting isolate of S. plymuthica A153 correlates impaired growth of several plant fungal pathogens with the volatiles for which ammonia is identified as an important component [133]. Activation of the WRKY18-plant stress pathway was noted with volatiles of Serratia [134].
These examples illustrate that the soil contains a diversity of microbial genera that can contribute to plant health by affecting the growth and metabolism of potentially harmful microbe, nematode, and insect challenges. The chosen examples highlight that many soil isolates thrive in the rhizosphere partly due to the carbon and nitrogen nutrition from plant metabolites in the root exudates. The examples, summarized in Figure 2, reveal a wide range of microbial metabolites are involved in the intricate interactions with cohabitors, and some directly reduce the effects of the plant predators. The sources of these metabolites, the biocontrol rhizobacteria, become the predators of the plant pests but retain balanced cohabitation with the plant.
3. Induction of Plant Systemic Resistance Mechanism by Biocontrol Agents
The observations of inhibited growth of fungal pathogens or the death of infested insect larvae or nematode juveniles established that plant-associated microbes produce compounds that directly inhibit pathogenic challenges. However, there is another valuable hidden trait: many microbes trigger the induction of systemic resistance mechanisms in the plant (Figure 1). The ability of beneficial microbes to induce resistance to both microbial plant pathogens and herbivorous insects/nematodes is summarized in several reviews [135,136].
In addition to causing direct pest antagonism, the microbial stimulation of plant tolerance also contributes to plant health. The onset of systemic plant resistance can be traced to the effects of discrete metabolites from the biological agents, indicating dual roles of these compounds as direct inhibitors of plant pests. This additional significance of biological control microbes to plant biotic resistance is discussed in Section 3.
Certain antifungal products from biocontrol-active pseudomonads and Bacillus isolates trigger plant resistance, as determined by treatment with the authentic compound. Additionally, this ability is detected from a comparison of responses that are induced in the plant by the wild type, but not by mutant cells defective in the metabolite. Resistance in the plant is seen at a distance from the site of microbial colonization; for example, although the stimulus is from a root colonizer, leaf tissues are protected against the pathogens. Thus, the term induced systemic resistance (ISR) is used for this process.
A cross-talking set of pathways involving different plant regulatory signals is involved. Identified pathways included the salicylic acid pathway, the jasmonate pathway, and an ethylene pathway [137]. Each pathway has a set of defense measures associated with the specific signal, such that one pathway offers protection against different sets of challenges [135]. For example, jasmonic acid pathway activation is associated with the plant’s responses to chewing insects [138], whereas plant cell necrosis by toxins from a plant pathogen triggers the salicylic acid pathway [139].
Studies with Pseudomonas sp. CMR12a find that mutants lacking phenazine production fail to induce systemic resistance in rice, whereas it is induced by the wild type cells and the phenazine metabolite, PCN [140]. Another phenazine, pyocyanin, produced by P. aeruginosa 7NSK2, stimulates ISR in tomatoes [141]. In tobacco, mutants of P. chlororaphis O6 lacking in phenazines are unable to induce systemic resistance to the soft-rot-causing pathogen Erwinia carotovora [142]. The ethylene defense pathway, rather than the salicylic acid pathway, is induced by P. chlororaphis O6 [143]. One common response to phenazines in plants is the production of reactive oxygen species, which may be an initiator for plant defense gene expression changes leading to ISR.
With P. chlororaphis PA23, the production of pyrrolnitrin is a possible activator for low-level systemic resistance to blackleg in canola [144]. Different compounds, an alkaloid, two amino lipids, three arylalkylamines, and a terpenoid, are active ISR activators from another root colonizer, P. fluorescens N 21.4 [145]. These were found by screening the metabolite pool from this bacterium. The release of 2,4-DAPG from other pseudomonads activates the jasmonic acid/ethylene pathway for ISR in Arabidopsis [146]. Resistance induction by 2,4-DAPG correlates with the suppression of take-all in soils [147]. Additionally, the suppression of root-knot nematode infections in tomato roots is linked to induced resistance by 2,4-DAPG produced from P. fluorescens CHA0 [36].
Volatiles produced by biocontrol bacteria also trigger resistance [148]. Kishimoto et al. [149] find that authentic 1-octen-3-ol induces systemic resistance through the jasmonate pathway. This VOC is produced by several biocontrol microbes, as discussed in Section 2. Ryu et al. [150] identified 2,3-butanediol among microbial volatiles from Bacillus subtilis GB03 and Bacillus amyloliquefaciens N937 that induce systemic resistance to gray mold, and B. cinerea in Arabidopsis, where the ethylene pathway, but not the salicylic acid or jasmonate pathway, is active. Butanediol production from P. chlororaphis O6 also explains the induction of systemic resistance to Erwinia in tobacco [92,151].
The lipopeptides produced by many Bacillus species also trigger ISR. Stimulation of ISR by iturin A and fengycin from indigenous Bacillus spp. offers protection in rice. This mechanism adds to the direct inhibition of spore germination and mycelial growth to achieve a high level of plant protection [152]. Park et al. [153] show that several analogs of iturin A from Bacillus vallismortis strain EXTN-1 activate ISR against Phytophthora capsici in chili pepper. Indeed, soft rot disease in cherry tomato caused by Rhizopus stolonifera is controlled by ISR activated by iturin A [154]. In both iturin A studies, an array of plant genes encoding enzymes associated with plant resistance is expressed.
The chitinases secreted by many of the biocontrol agents may also participate in enhanced plant resistance. Increased expression of plant chitinase genes is part of the changes in metabolism seen when ISR is activated in the plant; the induced proteins are termed the “pathogenesis-related proteins”. Differences in the array of chitinase isozymes expressed are dependent on the activation of the salicylic or ethylene/jasmonic acid pathways [155,156]. These hydrolytic enzymes degrade chitins/chitosans that are integral components of the walls of fungi and insects and the coating of nematode eggs [157,158]. However, chitin oligomers derived from such activities are among the group of activators for innate immune responses in plants [156]. Consequently, the secretion by the bioactive bacteria of chitinases, often a complex of different isoforms [25], is yet another avenue by which these microbes can indirectly protect the plant through their potential to activate plant defenses.
4. Secondary Metabolites and Cell Preparations for Commercial Biocontrol Formulations
Several biopesticides based on biocontrol metabolites with a control potential as effective as chemical pesticides are currently marketed for agricultural use. One such example is fludioxonil, a synthetic fungicide derived by using pyrrolnitrin as a base structure. Therefore, it belongs to the phenylpyrroles group of pesticides (FRAC code 12). Fludioxonil inhibits signal transduction in fungi and effectively controls Sclerotinia and Botrytis in various crops. Thirty years after its development, there have been few reports of resistance to fludioxonil [159]. Another commercial product is shenqinmycin, based on PCA. It has been in use since 2011 to control fungal diseases [160]. Metabolites from Streptomyces spp. are the foundation for many other chemical pesticides. Insecticides and nematicides that mimic the natural products from Streptomyces spp. include abamectin, emamectin benzoate, spinosad, and spinetoram. Blasticidin-S, kusagamycin, streptomycin, oxytetracycline, validamycin, and polyoxins are marketed to control various bacterial and fungal pathogens with stable efficacy [161].
Among the commercialized microbial biopesticides with live cells are the products Serifel®, based on B. amyloliquefaciens MBI600, and Serenade®, formulated with B. subtilis QST 713. These products control plant diseases caused by bacterial and fungal pathogens. Their ability to colonize plant surfaces through biofilm formation is one of the factors cited by the manufacturers for their activity. Another is that they will stimulate the plant’s resistance mechanisms. Indeed, some products are sold with such claims, such as LifeGard from Certis, based on the protective isolate Bacillus mycoides, a phyllosphere colonist [162]. However, findings suggest that they may be less effective than chemical pesticides in the field, perhaps due to poor survival of the microorganism after application, a factor that would lower the production of the bioactive metabolites [163,164,165]. The discussions below indicate the steps between the initial promising laboratory findings and successful commercialization.
As illustrated in Section 2 and Section 3, there is an abundant array of metabolites connected to the secondary metabolism of biocontrol isolates that could be exploited to control multiple targets through ISR or direct antagonism. Dose-dependency is shown in in vitro and in planta studies and with different pathosystems. Genome sequencing of increasing numbers of bacteria with soil habitats should promote the prediction of additional active molecules and identify the microbes that produce them. Determining LC/LD50 and MIC values using in vitro assays identifies the metabolites with strong direct effects on targets at low concentrations; these metabolites should be the most lucrative in commercial applications. The bioactive secondary metabolites may have a long shelf life in formulations and persistence in the environment. They can be applied as stand-alone products or to boost the effects of preparations containing appropriate bacterial biocontrol strains.
Maximizing the production of bioactive metabolites can involve different strategies. One approach is adjusting culture methods to optimize metabolite production. A second approach is engineering the genome to overproduce the product or express the biosynthetic genes under a defined productive promoter in an organism for which industrial scale growth is established. Using “smart” promoters with beneficial genes could adjust their production in inoculated plants to only occur under pest challenging conditions. If a pure compound is desired, methods to extract and purify the targeted molecule need to be developed. Shelf life and methods of delivery also should be considered during commercial formulation.
Basic studies reveal that the metabolites are often not constitutively released as cultures grow but are synthesized at specific development times or after a defined environmental stimulus. For example, the fit genes are not expressed until the bacterium is within the insect host, and BT toxin synthesis is withheld until the start of spore formation in the Bacillus producers [53]. With Pseudomonas and Serratia isolates, many of the compounds are only produced when quorum sensing is activated, so that there is both a cell density and nutrient dependency. Such understanding comes from basic science studies to establish the knowledge of the lifestyle of the microbe and nutritional effects.
The development of superior culture media is vital. For Pseudomonas chlororaphis O6, glucose represses the synthesis of pyrrolnitrin but promotes phenazine production [42]. The concentration of antifungal metabolites produced by B. subtilis V26 increases up to 93% using potato extract as a carbon source and soybean powder as a nitrogen source. The optimized formulations result in better control of gray mold in tomatoes [166]. The addition of ornithine increases the production of the lipopeptide surfactin by B. amyloliquefaciens HM618, an effective antagonist for the pathogenic fungi B. cinerea and R. solani [167]. The efficacy of P. elgii HOA73 in controlling gray mold on tomatoes is enhanced by adding chitin during cultivation [168], presumably because it triggers chitinase production. Polyamine additions to cultures of P. chlororaphis O6 promote phenazine and pyrrolnitrin levels, leading to greater protection against the plant fungal pathogens R. solani and Didymella bryoniae [169].
The media and any stimulants should be cost-effective. Ghribi et al. [170] find that B. subtilis SPB1 produce more than twice as many lipopeptides through easily available and cost-effective carbon, nitrogen, and inorganic nutrient sources; the increased lipopeptide levels correlate with better control of Prays oleae larvae [170]. Waste product use improves cost-efficiency; for example, potato peel and fish wastes enable the production of active lipopeptides with MICs for Mucor sp. at 6.25 ppm and Aspergillus niger at 12.5 ppm [171]. An extended culture time enhances yield [18] and the product’s potential to thwart pathogenic bacteria and fungi upon application [172].
Programs are under development to systematically modify media to optimize outcomes. The use of “One Factor at a Time” and “Two Factor at a Time” methods by Meena et al. [171] increased lipopeptide output by B. subtilis KLP2015 by more than four times. Ghribi et al. [170] approach optimization statistically, with suggestions that variable factors such as temperature, moisture, and inoculum age are important. Predicting optimal culture conditions using a statistical approach with the Plackett–Burman design (PBD) increased the production of chitinase and beta 1,4-endoglucanase by P. elgii PB1 and also enhanced antibacterial activity [173]. The use of PBD shows production of the insecticide avermectin B1 from Streptomyces avermitilis 14-12 to be positively influenced by carbon and nitrogen sources [174]. Media composition for producing avermectin B1 has also been optimized using another statistical tool, the Response Surface Method (RSM) [174]. Analysis with RSM additionally optimized the culture parameters for the production of the antimicrobial actinomycin D from Streptomyces hydrogenans IB310, an outcome also accompanied by lowered production costs [175].
Several studies have already demonstrated the benefit of genetic engineering in enhancing bioactive products. An early study with P. fluorescens CHA0 found that mutations in the gene rpoD, encoding the housekeeping sigma factor, enhances the production of the antifungals pyoluteorin, and 2,4-DAPG [35]. The genetic modification of ribosomal structure in P. protegens Pf-5 is another method for increasing 2,4-DAPG and pyoluteorin levels [176]. Introduction and expression of a cry gene enables the synthesis of a BT toxin in a transformed wild type B. velezensis strain to deliver its native strong antifungal activity and insecticidal efficacy, as demonstrated by its effects on third instar larvae of the diamondback moth P. xylostella [177]. The transformation of Pseudomonas synxantha 2–79, a strong producer of PCA, with genes enabling the production of pyrrolnitrin, extends the protection range of this biocontrol agent to more pathogens in cereals [178]. To heighten its antifungal potential, a combination of ten mutations in regulatory and synthesis pathway genes promotes 2-hydroxyphenazine synthesis in P. chlororaphis GP72 four-fold [179]. Changing glycerol utilization in this same microbe improves PCA production [160]. This effort is justified because of the success of the PCA-based pesticide “Shenqinmycin” and the availability of glycerol as a waste product from diesel formation [160]. Other groups have modified different P. chlororaphis isolates to produce PCN and 1-hydroxyphenazines in order to enhance the range of plant fungal pathogens that are targeted by one isolate [180,181].
Intact whole live or dead cells could also be important in formulations. Dead cells should still stimulate innate plant resistance mechanisms because the nonviable cells retain structures that act as microbe-associated molecular patterns (MAMPs) [135,182]. Live cells are essential for benefitting the biofilm formation as a root protectant layer and providing continuous sources of VOCs or other biocontrol-active metabolites. Basic research could show mixtures of beneficial organisms that are additive or even synergistic in providing crop protection. A mixture of Bacillus species that provides BT toxins and lipopeptides would be a potent weapon for insect and fungal control. Indeed, combined applications of B. pumilus PTB180 and B. subtilis PTB185 are compatible with insecticidal activity on aphids in the laboratory and greenhouse [16]. Additionally, a tripartite mixture of an arbuscular mycorrhizal fungus, Bacillus pumilus, and Pseudomonas alcaligenes enhances the antifungal properties and reduces the root disease caused by the root-knot nematode and a root rotting fungus in chickpea [183].
The studies discussed in Section 2, Section 3 and Section 4 illustrate how partnerships between basic research science on biocontrol microbes and industry can lead to products for agricultural applications that improve plant health. Exchanges of successful methods to produce commercial quantities of reliable cultures of pure or mixed microbes already achieved by the pharmaceutical industry and the agricultural groups are appropriate. For example, probiotics and prebiotics for human and animal health are readily available as “on the shelf” products to purchase.
5. Conclusions
It is clear that plants have engineered their own protection schemes by exploiting the intricacies of microbial activities from beneficial microbes that they nurture, especially in the rhizosphere. Emphasized in this review is the fact that rhizospheric biocontrol microbes have the weaponry to be pathogens themselves on multiple plant pests, i.e., fungal pathogens, insects, and nematodes, while having adapted to cohabit with the plant roots such that plant health is promoted. Development is needed to commercialize the varied direct and indirect mechanisms for biocontrol that is revealed by basic research. The current realization of the importance of nurturing soil health for regenerative agriculture aligns well with the use of biopesticides. Consumers are aware of the importance of probiotics for their health, and marketing could better convey their existence and value for plant health in agriculture. Furthermore, the microbes that can control multiple plant pests have a high potential to boost crop output regarding quality and yield at this time when global food security is a major issue.
Author Contributions
Conceptualization, J.H.L. and Y.C.K.; writing—review and editing, J.H.L., A.J.A. and Y.C.K.; funding acquisition, Y.C.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Korean Institute of Planning and Evaluation for Technology in Food, Agriculture, and Forestry (IPET) through the Agriculture Machinery/Equipment Localization Technology Development Program and funded by the Ministry of Agriculture, Food, and Rural Affairs (MAFRA) (321055-55).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design, execution, interpretation, or writing of the study.
References
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Handford, C.E.; Elliott, C.T.; Campbell, K. A review of the global pesticide legislation and the scale of challenge in reaching the global harmonization of food safety standards. Integr. Environ. Assess. Manag. 2015, 11, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Labourdette, G.; Lachaise, H.; Rieck, H.; Steiger, D. Fluopyram: Efficacy and beyond on problematic diseases. In Proceedings of the Modern Fungicides and Antifungal Compounds VI, 16th International Reinhardsbrunn Symposium, Friedrichroda, Germany, 25–29 April 2010; pp. 75–80. [Google Scholar]
- Faske, T.; Hurd, K. Sensitivity of Meloidogyne incognita and Rotylenchulus reniformis to fluopyram. J. Nematol. 2015, 47, 316. [Google Scholar] [PubMed]
- Rouquie, D.; Tinwell, H.; Blanck, O.; Schorsch, F.; Geter, D.; Wason, S.; Bars, R. Thyroid tumor formation in the male mouse induced by fluopyram is mediated by activation of hepatic CAR/PXR nuclear receptors. Regul. Toxicol. Pharmacol. 2014, 70, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.K.; Khare, E.; Maheshwari, D.K. Plant growth promoting rhizobacteria: Constraints in bioformulation, commercialization, and future strategies. In Plant Growth and Health Promoting Bacteria; Microbiology Monographs 18; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 97–116. [Google Scholar]
- Bashan, Y. Inoculants of plant growth-promoting bacteria for use in agriculture. Biotechnol. Adv. 1998, 16, 729–770. [Google Scholar] [CrossRef]
- Cook, R.J. Take-all of wheat. Physiol. Mol. Plant Pathol. 2003, 62, 73–86. [Google Scholar] [CrossRef]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Kang, B.R.; Han, J.H.; Kim, J.J.; Kim, Y.C. Dual biocontrol potential of the entomopathogenic fungus, Isaria javanica, for both aphids and plant fungal pathogens. Mycobiology 2018, 46, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Alfonzo, A.; Lo Piccolo, S.; Conigliaro, G.; Ventorino, V.; Burruano, S.; Moschetti, G. Antifungal peptides produced by Bacillus amyloliquefaciens AG1 active against grapevine fungal pathogens. Ann. Microbiol. 2012, 62, 1593–1599. [Google Scholar] [CrossRef]
- Khedher, S.B.; Boukedi, H.; Kilani-Feki, O.; Chaib, I.; Laarif, A.; Abdelkefi-Mesrati, L.; Tounsi, S. Bacillus amyloliquefaciens AG1 biosurfactant: Putative receptor diversity and histopathological effects on Tuta absoluta midgut. J. Invertebr. Pathol. 2015, 132, 42–47. [Google Scholar] [CrossRef]
- Rodríguez, M.; Marín, A.; Torres, M.; Béjar, V.; Campos, M.; Sampedro, I. Aphicidal activity of surfactants produced by Bacillus atrophaeus L193. Front. Microbiol. 2018, 9, 3114. [Google Scholar] [CrossRef] [Green Version]
- Navarro, L.T.; Rodríguez, M.; Martínez-Checa, F.; Montaño, A.; Cortés-Delgado, A.; Smolinska, A.; Llamas, I.; Sampedro, I. Identification of volatile organic compounds in extremophilic bacteria and their effective use in biocontrol of postharvest fungal phytopathogens. Front. Microbiol. 2021, 12, 3349. [Google Scholar]
- Cossus, L.; Roux-Dalvai, F.; Kelly, I.; Nguyen, T.T.A.; Antoun, H.; Droit, A.; Tweddell, R.J. Interactions with plant pathogens influence lipopeptides production and antimicrobial activity of Bacillus subtilis strain PTB185. Biol. Control 2021, 154, 104497. [Google Scholar] [CrossRef]
- Kahia, M.; Nguyen, T.T.A.; McCune, F.; Naasz, R.; Antoun, H.; Fournier, V. Insecticidal effect of Bacillus pumilus PTB180 and Bacillus subtilis PTB185 used alone and in combination against the foxglove aphid and the melon aphid (Hemiptera: Aphididae). Can. Entomol. 2021, 153, 726–740. [Google Scholar] [CrossRef]
- Ghribi, D.; Abdelkefi-Mesrati, L.; Boukedi, H.; Elleuch, M.; Ellouze-Chaabouni, S.; Tounsi, S. The impact of the Bacillus subtilis SPB1 biosurfactant on the midgut histology of Spodoptera littoralis (Lepidoptera: Noctuidae) and determination of its putative receptor. J. Invertebr. Pathol. 2012, 109, 183–186. [Google Scholar] [CrossRef]
- Mnif, I.; Elleuch, M.; Chaabouni, S.E.; Ghribi, D. Bacillus subtilis SPB1 biosurfactant: Production optimization and insecticidal activity against the carob moth Ectomyelois ceratoniae. Crop Prot. 2013, 50, 66–72. [Google Scholar] [CrossRef]
- Mnif, I.; Hammami, I.; Triki, M.A.; Azabou, M.C.; Ellouze-Chaabouni, S.; Ghribi, D. Antifungal efficiency of a lipopeptide biosurfactant derived from Bacillus subtilis SPB1 versus the phytopathogenic fungus, Fusarium solani. Environ. Sci. Pollut. Res. 2015, 22, 18137–18147. [Google Scholar] [CrossRef]
- Mnif, I.; Grau-Campistany, A.; Coronel-León, J.; Hammami, I.; Triki, M.A.; Manresa, A.; Ghribi, D. Purification and identification of Bacillus subtilis SPB1 lipopeptide biosurfactant exhibiting antifungal activity against Rhizoctonia bataticola and Rhizoctonia solani. Environ. Sci. Pollut. Res. 2016, 23, 6690–6699. [Google Scholar] [CrossRef]
- Ben Khedher, S.; Boukedi, H.; Laarif, A.; Tounsi, S. Biosurfactant produced by Bacillus subtilis V26: A potential biological control approach for sustainable agriculture development. Org. Agric. 2020, 10, 117–124. [Google Scholar] [CrossRef]
- Wang, M.; Geng, L.; Sun, X.; Shu, C.; Song, F.; Zhang, J. Screening of Bacillus thuringiensis strains to identify new potential biocontrol agents against Sclerotinia sclerotiorum and Plutella xylostella in Brassica campestris L. Biol. Control 2020, 145, 104262. [Google Scholar] [CrossRef]
- Kim, P.; Bai, H.; Bai, D.; Chae, H.; Chung, S.; Kim, Y.; Park, R.; Chi, Y.T. Purification and characterization of a lipopeptide produced by Bacillus thuringiensis CMB26. J. Appl. Microbiol. 2004, 97, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, L.; Eijsink, V.G.; Meadow, R.; Gåseidnes, S. A novel strain of Brevibacillus laterosporus produces chitinases that contribute to its biocontrol potential. Appl. Microbiol. Biotechnol. 2013, 97, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-C.; Kang, B.R.; Kim, Y.H.; Park, S.K. An effective and practical strategy for biocontrol of plant diseases using on-site mass cultivation of chitin-degrading bacteria. Res. Plant Dis. 2017, 23, 19–34. [Google Scholar] [CrossRef]
- Neung, S.; Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Kim, K.Y. Insecticidal potential of Paenibacillus elgii HOA73 and its combination with organic sulfur pesticide on diamondback moth, Plutella xylostella. J. Korean Soc. Appl. Biol. Chem. 2014, 57, 181–186. [Google Scholar] [CrossRef]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Jung, W.J.; Anees, M.; Kim, K.Y. Antagonistic potential of Paenibacillus elgii HOA73 against the root-knot nematode, Meloidogyne incognita. Nematology 2013, 15, 991–1000. [Google Scholar] [CrossRef]
- Lee, Y.S.; Nguyen, X.H.; Cho, J.-Y.; Moon, J.-H.; Kim, K.Y. Isolation and antifungal activity of methyl 2, 3-dihydroxybenzoate from Paenibacillus elgii HOA73. Microb. Pathog. 2017, 106, 139–145. [Google Scholar] [CrossRef]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Moon, J.H.; Lee, J.H.; Kim, K.Y. Isolation and characteristics of protocatechuic acid from Paenibacillus elgii HOA73 against Botrytis cinerea on strawberry fruits. J. Basic Microbiol. 2015, 55, 625–634. [Google Scholar] [CrossRef]
- Zhao, L.-j.; Yang, X.-n.; Li, X.-y.; Wei, M.; Feng, L. Antifungal, insecticidal and herbicidal properties of volatile components from Paenibacillus polymyxa strain BMP-11. Agric. Sci. China 2011, 10, 728–736. [Google Scholar] [CrossRef]
- Kaur, T.; Manhas, R.K. Antifungal, insecticidal, and plant growth promoting potential of Streptomyces hydrogenans DH16. J. Basic Microbiol. 2014, 54, 1175–1185. [Google Scholar] [CrossRef]
- Kaur, T.; Jasrotia, S.; Ohri, P.; Manhas, R.K. Evaluation of in vitro and in vivo nematicidal potential of a multifunctional streptomycete, Streptomyces hydrogenans strain DH16 against Meloidogyne incognita. Microbiol. Res. 2016, 192, 247–252. [Google Scholar] [CrossRef]
- Craveri, R.; Giolitti, G. An antibiotic with fungicidal and insecticidal activity produced by Streptomyces. Nature 1957, 179, 1307. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Khan, A.L.; Ali, L.; Khan, A.R.; Waqas, M.; Hussain, J.; Lee, I.-J.; Shin, J.-H. Benzaldehyde as an insecticidal, antimicrobial, and antioxidant compound produced by Photorhabdus temperata M1021. J. Microbiol. 2015, 53, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Schnider, U.; Keel, C.; Blumer, C.; Troxler, J.; Défago, G.; Haas, D. Amplification of the housekeeping sigma factor in Pseudomonas fluorescens CHA0 enhances antibiotic production and improves biocontrol abilities. J. Bacteriol. 1995, 177, 5387–5392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, I.A.; Shaukat, S.S. Suppression of root-knot disease by Pseudomonas fluorescens CHA0 in tomato: Importance of bacterial secondary metabolite, 2,4-diacetylpholoroglucinol. Soil Biol. Biochem. 2003, 35, 1615–1623. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Haas, D.; Heeb, S. Extracellular protease of Pseudomonas fluorescens CHA0, a biocontrol factor with activity against the root-knot nematode Meloidogyne incognita. Appl. Environ. Microbiol. 2005, 71, 5646–5649. [Google Scholar] [CrossRef] [Green Version]
- Ruffner, B.; Péchy-Tarr, M.; Ryffel, F.; Hoegger, P.; Obrist, C.; Rindlisbacher, A.; Keel, C.; Maurhofer, M. Oral insecticidal activity of plant-associated pseudomonads. Environ. Microbiol. 2013, 15, 751–763. [Google Scholar] [CrossRef]
- Chin-A-Woeng, T.F.; Bloemberg, G.V.; Mulders, I.H.; Dekkers, L.C.; Lugtenberg, B.J. Root colonization by phenazine-1-carboxamide-producing bacterium Pseudomonas chlororaphis PCL1391 is essential for biocontrol of tomato foot and root rot. Mol. Plant Microbe Interact. 2000, 13, 1340–1345. [Google Scholar] [CrossRef] [Green Version]
- Flury, P.; Vesga, P.; Péchy-Tarr, M.; Aellen, N.; Dennert, F.; Hofer, N.; Kupferschmied, K.P.; Kupferschmied, P.; Metla, Z.; Ma, Z. Antimicrobial and insecticidal: Cyclic lipopeptides and hydrogen cyanide produced by plant-beneficial Pseudomonas strains CHA0, CMR12a, and PCL1391 contribute to insect killing. Front. Microbiol. 2017, 8, 100. [Google Scholar] [CrossRef]
- Lee, J.H.; Ma, K.C.; Ko, S.J.; Kang, B.R.; Kim, I.S.; Kim, Y.C. Nematicidal activity of a nonpathogenic biocontrol bacterium, Pseudomonas chlororaphis O6. Curr. Microbiol. 2011, 62, 746–751. [Google Scholar] [CrossRef]
- Park, J.; Oh, S.; Anderson, A.; Neiswender, J.; Kim, J.C.; Kim, Y. Production of the antifungal compounds phenazine and pyrrolnitrin from Pseudomonas chlororaphis O6 is differentially regulated by glucose. Lett. Appl. Microbiol. 2011, 52, 532–537. [Google Scholar] [CrossRef]
- Kang, B.R.; Anderson, A.J.; Kim, Y.C. Hydrogen cyanide produced by Pseudomonas chlororaphis O6 exhibits nematicidal activity against Meloidogyne hapla. Plant Pathol. J. 2018, 34, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.R.; Anderson, A.J.; Kim, Y.C. Hydrogen cyanide produced by Pseudomonas chlororaphis O6 is a key aphicidal metabolite. Can. J. Microbiol. 2019, 65, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Selin, C.; Habibian, R.; Poritsanos, N.; Athukorala, S.N.; Fernando, D.; De Kievit, T.R. Phenazines are not essential for Pseudomonas chlororaphis PA23 biocontrol of Sclerotinia sclerotiorum, but do play a role in biofilm formation. FEMS Microbiol. Ecol. 2009, 71, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandi, M.; Selin, C.; Brassinga, A.K.C.; Belmonte, M.F.; Fernando, W.D.; Loewen, P.C.; De Kievit, T.R. Pyrrolnitrin and hydrogen cyanide production by Pseudomonas chlororaphis strain PA23 exhibits nematicidal and repellent activity against Caenorhabditis elegans. PLoS ONE 2015, 10, e0123184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyay, P.; Sen, S.K. Systemic infestation of Serratia entomophila AB2 through plant tissue inferred protection against insect pest and fungal pathogens. Afr. J. Microbiol. Res. 2013, 7, 2651–2655. [Google Scholar]
- Berlec, A. Novel techniques and findings in the study of plant microbiota: Search for plant probiotics. Plant Sci. 2012, 193, 96–102. [Google Scholar] [CrossRef]
- Spence, C.; Alff, E.; Shantharaj, D.; Bais, H. Probiotics for plants: Importance of rhizobacteria on aboveground fitness in plants. In Bacteria in Agrobiology: Plant Probiotics; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–14. [Google Scholar]
- Woo, S.L.; Pepe, O. Microbial consortia: Promising probiotics as plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1801. [Google Scholar] [CrossRef]
- Regaiolo, A.; Dominelli, N.; Andresen, K.; Heermann, R. The biocontrol agent and insect pathogen Photorhabdus luminescens interacts with plant roots. Appl. Environ. Microbiol. 2020, 86, e00891-20. [Google Scholar] [CrossRef]
- Melo, A.L.d.A.; Soccol, V.T.; Soccol, C.R. Bacillus thuringiensis: Mechanism of action, resistance, and new applications. A review. Crit. Rev. Biotechnol. 2016, 36, 317–326. [Google Scholar] [CrossRef]
- Keel, C. A look into the toolbox of multi-talents: Insect pathogenicity determinants of plant-beneficial pseudomonads. Environ. Microbiol. 2016, 18, 3207–3209. [Google Scholar] [CrossRef] [Green Version]
- Ruffner, B.; Péchy-Tarr, M.; Höfte, M.; Bloemberg, G.; Grunder, J.; Keel, C.; Maurhofer, M. Evolutionary patchwork of an insecticidal toxin shared between plant-associated pseudomonads and the insect pathogens Photorhabdus and Xenorhabdus. BMC Genom. 2015, 16, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mnif, I.; Ghribi, D. Review lipopeptides biosurfactants: Mean classes and new insights for industrial, biomedical, and environmental applications. Peptide Sci. 2015, 104, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Chin-A-Woeng, T.F.; Bloemberg, G.V.; Lugtenberg, B.J. Phenazines and their role in biocontrol by Pseudomonas bacteria. New Phytol. 2003, 157, 503–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuel, J.; Selin, C.; Fernando, W.D.; de Kievit, T. Stringent response mutants of Pseudomonas chlororaphis PA23 exhibit enhanced antifungal activity against Sclerotinia sclerotiorum in vitro. Microbiology 2012, 158, 207–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, G.; Arya, S.K. Antifungal and insecticidal potential of chitinases: A credible choice for the eco-friendly farming. Biocatal. Agric. Biotechnol. 2019, 20, 101289. [Google Scholar] [CrossRef]
- Sharp, R.G. A review of the applications of chitin and its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy 2013, 3, 757–793. [Google Scholar] [CrossRef]
- Kim, Y.C.; Anderson, A.J. Rhizosphere pseudomonads as probiotics improving plant health. Mol. Plant Pathol. 2018, 19, 2349–2359. [Google Scholar] [CrossRef] [Green Version]
- Dennis, P.G.; Miller, A.J.; Hirsch, P.R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities? FEMS Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, D.S.; Teixeira, P.J. Root-exuded coumarin shapes the root microbiome. Proc. Natl. Acad. Sci. USA 2018, 115, 5629–5631. [Google Scholar] [CrossRef] [Green Version]
- Stringlis, I.A.; Yu, K.; Feussner, K.; de Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.; Feussner, I.; Pieterse, C.M. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl. Acad. Sci. USA 2018, 115, E5213–E5222. [Google Scholar] [CrossRef] [Green Version]
- Maddula, V.; Pierson, E.; Pierson III, L. Altering the ratio of phenazines in Pseudomonas chlororaphis (aureofaciens) strain 30–84: Effects on biofilm formation and pathogen inhibition. J. Bacteriol. 2008, 190, 2759–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderón, C.E.; Tienda, S.; Heredia-Ponce, Z.; Arrebola, E.; Cárcamo-Oyarce, G.; Eberl, L.; Cazorla, F.M. The compound 2-hexyl, 5-propyl resorcinol has a key role in biofilm formation by the biocontrol rhizobacterium Pseudomonas chlororaphis PCL1606. Front. Microbiol. 2019, 10, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daud, N.S.; Rosli, M.A.; Azam, Z.M.; Othman, N.Z.; Sarmidi, M.R. Paenibacillus polymyxa bioactive compounds for agricultural and biotechnological applications. Biocatal. Agric. Biotechnol. 2019, 18, 101092. [Google Scholar] [CrossRef]
- Chan, S.Y.; Liu, S.Y.; Seng, Z.; Chua, S.L. Biofilm matrix disrupts nematode motility and predatory behavior. ISME J. 2021, 15, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Nandi, M.; Berry, C.; Brassinga, A.K.C.; Belmonte, M.F.; Fernando, W.D.; Loewen, P.C.; de Kievit, T.R. Pseudomonas brassicacearum strain DF41 kills Caenorhabditis elegans through biofilm-dependent and biofilm-independent mechanisms. Appl. Environ. Microbiol. 2016, 82, 6889–6898. [Google Scholar] [CrossRef] [Green Version]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Boiteau, R.M.; Markillie, L.M.; Hoyt, D.W.; Hu, D.; Chu, R.K.; Mitchell, H.D.; Pasa-Tolic, L.; Jansson, J.K.; Jansson, C. Metabolic interactions between Brachypodium and Pseudomonas fluorescens under controlled iron-limited conditions. mSystems 2021, 6, e00580-20. [Google Scholar] [CrossRef]
- Yahya, M.; ul Islam, E.; Rasul, M.; Farooq, I.; Mahreen, N.; Tawab, A.; Irfan, M.; Rajput, L.; Amin, I.; Yasmin, S. Differential root exudation and architecture for improved growth of wheat mediated by phosphate solubilizing bacteria. Front. Microbiol. 2021, 12, 744094. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Finke, M.D. Nutritional value of insects and ways to manipulate their composition. J. Insects Food Feed 2020, 7, 639–659. [Google Scholar] [CrossRef]
- Bukkens, S.G.F. The nutritional value of edible insects. Ecol. Food Nutr. 1997, 36, 287–319. [Google Scholar] [CrossRef]
- Cox, G.N.; Kusch, M.; Edgar, R.S. Cuticle of Caenorhabditis elegans: Its isolation and partial characterization. J. Cell Biol. 1981, 90, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Pronk, L.J.; Bakker, P.A.; Keel, C.; Maurhofer, M.; Flury, P. The secret life of plant-beneficial rhizosphere bacteria: Insects as alternative hosts. Environ. Microbiol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théatre, A.; Cano-Prieto, C.; Bartolini, M.; Laurin, Y.; Deleu, M.; Niehren, J.; Fida, T.; Gerbinet, S.; Alanjary, M.; Medema, M.H. The surfactin-like lipopeptides from Bacillus spp.: Natural biodiversity and synthetic biology for a broader application range. Front. Bioeng. Biotechnol. 2021, 9, 118. [Google Scholar] [CrossRef] [PubMed]
- Arroyave-Toro, J.J.; Mosquera, S.; Villegas-Escobar, V. Biocontrol activity of Bacillus subtilis EA-CB0015 cells and lipopeptides against postharvest fungal pathogens. Biol. Control 2017, 114, 195–200. [Google Scholar] [CrossRef]
- Yan, F.; Li, C.; Ye, X.; Lian, Y.; Wu, Y.; Wang, X. Antifungal activity of lipopeptides from Bacillus amyloliquefaciens MG3 against Colletotrichum gloeosporioides in loquat fruits. Biol. Control 2020, 146, 104281. [Google Scholar] [CrossRef]
- Kalai-Grami, L.; Karkouch, I.; Naili, O.; Slimene, I.B.; Elkahoui, S.; Zekri, R.B.; Touati, I.; Mnari-Hattab, M.; Hajlaoui, M.R.; Limam, F. Production and identification of iturin A lipopeptide from Bacillus methyltrophicus TEB1 for control of Phoma tracheiphila. J. Basic Microbiol. 2016, 56, 864–871. [Google Scholar] [CrossRef]
- Romano, A.; Vitullo, D.; Senatore, M.; Lima, G.; Lanzotti, V. Antifungal cyclic lipopeptides from Bacillus amyloliquefaciens strain BO5A. J. Nat. Prod. 2013, 76, 2019–2025. [Google Scholar] [CrossRef]
- Yang, S.Y.; Lim, D.J.; Noh, M.Y.; Kim, J.C.; Kim, Y.C.; Kim, I.S. Characterization of biosurfactants as insecticidal metabolites produced by Bacillus subtilis Y9. Entomol. Res. 2017, 47, 55–59. [Google Scholar] [CrossRef]
- Simionato, A.S.; Navarro, M.O.; de Jesus, M.L.; Barazetti, A.R.; da Silva, C.S.; Simões, G.C.; Balbi-Peña, M.I.; de Mello, J.C.; Panagio, L.A.; de Almeida, R.S. The effect of phenazine-1-carboxylic acid on mycelial growth of Botrytis cinerea produced by Pseudomonas aeruginosa LV strain. Front. Microbiol. 2017, 8, 1102. [Google Scholar] [CrossRef]
- Lee, J.Y.; Moon, S.S.; Hwang, B.K. Isolation and in vitro and in vivo activity against Phytophthora capsici and Colletotrichum orbiculare of phenazine-1-carboxylic acid from Pseudomonas aeruginosa strain GC-B26. Pest Manag. Sci. 2003, 59, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Rane, M.R.; Sarode, P.D.; Chaudhari, B.L.; Chincholkar, S.B. Detection, isolation and identification of phenazine-1-carboxylic acid produced by biocontrol strains of Pseudomonas aeruginosa. J. Sci. Ind. Res. 2007, 66, 627–631. [Google Scholar]
- Xu, Z.; Wang, M.; Du, J.; Huang, T.; Liu, J.; Dong, T.; Chen, Y. Isolation of Burkholderia sp. HQB-1, a promising biocontrol bacteria to protect banana against Fusarium wilt through phenazine-1-carboxylic acid secretion. Front. Microbiol. 2020, 11, 3156. [Google Scholar] [CrossRef]
- Gurusiddaiah, S.; Weller, D.; Sarkar, A.; Cook, R. Characterization of an antibiotic produced by a strain of Pseudomonas fluorescens inhibitory to Gaeumannomyces graminis var. tritici and Pythium spp. Antimicrob. Agents Chemother. 1986, 29, 488–495. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, C.; Su, P.; Liao, X. Control effect and possible mechanism of the natural compound phenazine-1-carboxamide against Botrytis cinerea. PLoS ONE 2015, 10, e0140380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugaiah, V.; Mathivanan, N.; Varghese, B. Purification, crystal structure and antimicrobial activity of phenazine-1-carboxamide produced by a growth-promoting biocontrol bacterium, Pseudomonas aeruginosa MML2212. J. Appl. Microbiol. 2010, 108, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Keel, C.; Schnider, U.; Maurhofer, M.; Voisard, C.; Laville, J.; Burger, U.; Wirthner, P.J.; Haas, D.; Défago, G. Suppression of root diseases by Pseudomonas fluorescens CHA0: Importance of the bacterial secondary metabolite 2,4-diacetylphloroglucinol. Mol. Plant Microbe Interact. 1992, 5, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Meyer, S.L.; Halbrendt, J.M.; Carta, L.K.; Skantar, A.M.; Liu, T.; Abdelnabby, H.M.; Vinyard, B.T. Toxicity of 2,4-diacetylphloroglucinol (DAPG) to plant-parasitic and bacterial-feeding nematodes. J. Nematol. 2009, 41, 274. [Google Scholar]
- Han, S.H.; Lee, S.J.; Moon, J.H.; Park, K.H.; Yang, K.Y.; Cho, B.H.; Kim, K.Y.; Kim, Y.W.; Lee, M.C.; Anderson, A.J. GacS-dependent production of 2R, 3R-butanediol by Pseudomonas chlororaphis O6 is a major determinant for eliciting systemic resistance against Erwinia carotovora but not against Pseudomonas syringae pv. tabaci in tobacco. Mol. Plant Microbe Interact. 2006, 19, 924–930. [Google Scholar] [CrossRef] [Green Version]
- Mendis, H.C.; Thomas, V.P.; Schwientek, P.; Salamzade, R.; Chien, J.-T.; Waidyarathne, P.; Kloepper, J.; De La Fuente, L. Strain-specific quantification of root colonization by plant growth promoting rhizobacteria Bacillus firmus I-1582 and Bacillus amyloliquefaciens QST713 in non-sterile soil and field conditions. PLoS ONE 2018, 13, e0193119. [Google Scholar] [CrossRef]
- Berini, F.; Katz, C.; Gruzdev, N.; Casartelli, M.; Tettamanti, G.; Marinelli, F. Microbial and viral chitinases: Attractive biopesticides for integrated pest management. Biotechnol. Adv. 2018, 36, 818–838. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Zhou, Q.; Luo, H.; Xia, L.; Li, L.; Sun, M.; Yu, Z. Systemic nematicidal activity and biocontrol efficacy of Bacillus firmus against the root-knot nematode Meloidogyne incognita. World J. Microbiol. Biotechnol. 2015, 31, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, G.; Horak, I.; Jansen van Rensburg, P.J.; Claassens, S. Bacillus-based bionematicides: Development, modes of action and commercialisation. Biocontrol Sci. Technol. 2018, 28, 629–653. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Shi, X.; Wang, B.; Li, M.; Wang, Q.; Zhang, S. Antifungal effect of volatile organic compounds from Bacillus velezensis CT32 against Verticillium dahliae and Fusarium oxysporum. Processes 2020, 8, 1674. [Google Scholar] [CrossRef]
- Massawe, V.C.; Hanif, A.; Farzand, A.; Mburu, D.K.; Ochola, S.O.; Wu, L.; Tahir, H.A.S.; Gu, Q.; Wu, H.; Gao, X. Volatile compounds of endophytic Bacillus spp. have biocontrol activity against Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1373–1385. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Park, S.K.; Hur, J.Y.; Kim, Y.C. Purification and characterization of a major extracellular chitinase from a biocontrol bacterium, Paenibacillus elgii HOA73. Plant Pathol. J. 2017, 33, 318–328. [Google Scholar] [CrossRef] [Green Version]
- Xiong, C.; Li, Q.; Li, S.; Chen, C.; Chen, Z.; Huang, W. In vitro antimicrobial activities and mechanism of 1-octen-3-ol against food-related bacteria and pathogenic fungi. J. Oleo Sci. 2017, 66, ess16196. [Google Scholar] [CrossRef] [Green Version]
- Herrera, J.M.; Pizzolitto, R.P.; Zunino, M.P.; Dambolena, J.S.; Zygadlo, J.A. Effect of fungal volatile organic compounds on a fungus and an insect that damage stored maize. J. Stored Prod. Res. 2015, 62, 74–80. [Google Scholar] [CrossRef]
- Cheng, W.; Yang, J.; Nie, Q.; Huang, D.; Yu, C.; Zheng, L.; Cai, M.; Thomashow, L.S.; Weller, D.M.; Yu, Z. Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies. Sci. Rep. 2017, 7, 16213. [Google Scholar] [CrossRef]
- Dahlstrom, K.M.; McRose, D.L.; Newman, D.K. Keystone metabolites of crop rhizosphere microbiomes. Curr. Biol. 2020, 30, R1131–R1137. [Google Scholar] [CrossRef]
- Biessy, A.; Filion, M. Phenazines in plant-beneficial Pseudomonas spp.: Biosynthesis, regulation, function and genomics. Environ. Microbiol. 2018, 20, 3905–3917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-Y.; Shi, X.-C.; Chen, X.; Laborda, P.; Zhao, Y.-Y.; Liu, F.-Q.; Laborda, P. Biocontrol ability of phenazine-producing strains for the management of fungal plant pathogens: A review. Biol. Control 2021, 155, 104548. [Google Scholar] [CrossRef]
- Mazzola, M.; Fujimoto, D.K.; Thomashow, L.S.; Cook, R.J. Variation in sensitivity of Gaeumannomyces graminis to antibiotics produced by fluorescent Pseudomonas spp. and effect on biological control of take-all of wheat. Appl. Environ. Microbiol. 1995, 61, 2554–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, A.J.; Kim, Y.C. Biopesticides produced by plant-probiotic Pseudomonas chlororaphis isolates. Crop Prot. 2018, 105, 62–69. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, Z.; Yu, L.; Hu, D.; Song, B. Investigating the antifungal activity and mechanism of a microbial pesticide Shenqinmycin against Phoma sp. Pestic. Biochem. Physiol. 2018, 147, 46–50. [Google Scholar] [CrossRef]
- Kavitha, K.; Mathiyazhagan, S.; Sendhilvel, V.; Nakkeeran, S.; Chandrasekar, G.; Dilantha Fernando, W. Broad spectrum action of phenazine against active and dormant structures of fungal pathogens and root knot nematode. Arch. Phytopathol. Plant Prot. 2005, 38, 69–76. [Google Scholar] [CrossRef]
- Meena, K.S.; Jonathan, E.; Ardhanareeswaran, N. Isolation of phenazine and its activity against root-knot nematode, Meloidogyne incognita. Indian J. Nematol. 2013, 43, 180–183. [Google Scholar]
- Gong, L.; Tan, H.; Chen, F.; Li, T.; Zhu, J.; Jian, Q.; Yuan, D.; Xu, L.; Hu, W.; Jiang, Y. Novel synthesized 2,4-DAPG analogues: Antifungal activity, mechanism and toxicology. Sci. Rep. 2016, 6, 32266. [Google Scholar] [CrossRef] [Green Version]
- Loper, J.E.; Henkels, M.D.; Rangel, L.I.; Olcott, M.H.; Walker, F.L.; Bond, K.L.; Kidarsa, T.A.; Hesse, C.N.; Sneh, B.; Stockwell, V.O. Rhizoxin analogs, orfamide A and chitinase production contribute to the toxicity of Pseudomonas protegens strain Pf-5 to Drosophila melanogaster. Environ. Microbiol. 2016, 18, 3509–3521. [Google Scholar] [CrossRef]
- Péchy-Tarr, M.; Bruck, D.J.; Maurhofer, M.; Fischer, E.; Vogne, C.; Henkels, M.D.; Donahue, K.M.; Grunder, J.; Loper, J.E.; Keel, C. Molecular analysis of a novel gene cluster encoding an insect toxin in plant-associated strains of Pseudomonas fluorescens. Environ. Microbiol. 2008, 10, 2368–2386. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ. Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Anand, A.; Chinchilla, D.; Tan, C.; Mène-Saffrané, L.; L’Haridon, F.; Weisskopf, L. Contribution of hydrogen cyanide to the antagonistic activity of Pseudomonas strains against Phytophthora infestans. Microorganisms 2020, 8, 1144. [Google Scholar] [CrossRef]
- Sehrawat, A.; Sindhu, S.S.; Glick, B.R. Hydrogen cyanide production by soil bacteria: Biological control of pests and promotion of plant growth in sustainable agriculture. Pedosphere 2022, 32, 15–38. [Google Scholar] [CrossRef]
- Petrikovics, I.; Budai, M.; Kovacs, K.; Thompson, D.E. Past, present and future of cyanide antagonism research: From the early remedies to the current therapies. World J. Methodol. 2015, 5, 88. [Google Scholar] [CrossRef] [Green Version]
- Zdor, R.E. Bacterial cyanogenesis: Impact on biotic interactions. J. Appl. Microbiol. 2015, 118, 267–274. [Google Scholar] [CrossRef]
- Flaishman, M.A.; Eyal, Z.; Zilberstein, A.; Voisard, C.; Haas, D. Suppression of Septoria tritici blotch and leaf rust of wheat by recombinant cyanide-producing strains of Pseudomonas putida. Mol. Plant Microbe Interact. 1996, 9, 642–645. [Google Scholar] [CrossRef]
- Spence, C.A.; Raman, V.; Donofrio, N.M.; Bais, H.P. Global gene expression in rice blast pathogen Magnaporthe oryzae treated with a natural rice soil isolate. Planta 2014, 239, 171–185. [Google Scholar] [CrossRef]
- Nandi, M.; Selin, C.; Brawerman, G.; Fernando, W.D.; de Kievit, T.R. The global regulator ANR is essential for Pseudomonas chlororaphis strain PA23 biocontrol. Microbiology 2016, 162, 2159–2169. [Google Scholar] [CrossRef]
- Keel, C.; Voisard, C.; Berling, C.-H.; Kahr, G.; Defago, G. Iron sufficiency, a prerequisite for the suppression of tobacco black root rot by Pseudomonas fluorescens strain CHA 0 under gnotobiotic conditions. Phytopathology 1989, 79, 584–589. [Google Scholar] [CrossRef]
- Voisard, C.; Keel, C.; Haas, D.; Dèfago, G. Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnotobiotic conditions. EMBO J. 1989, 8, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Lanteigne, C.; Gadkar, V.J.; Wallon, T.; Novinscak, A.; Filion, M. Production of DAPG and HCN by Pseudomonas sp. LBUM300 contributes to the biological control of bacterial canker of tomato. Phytopathology 2012, 102, 967–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broderick, K.E.; Chan, A.; Balasubramanian, M.; Feala, J.; Reed, S.L.; Panda, M.; Sharma, V.S.; Pilz, R.B.; Bigby, T.D.; Boss, G.R. Cyanide produced by human isolates of Pseudomonas aeruginosa contributes to lethality in Drosophila melanogaster. J. Infect. Dis. 2008, 197, 457–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devi, K.K.; Kothamasi, D. Pseudomonas fluorescens CHA0 can kill subterranean termite Odontotermes obesus by inhibiting cytochrome c oxidase of the termite respiratory chain. FEMS Microbiol. Lett. 2009, 300, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.-Y.; Jang, E.-J.; Park, S.-Y.; Son, H.-J. Production of HCN, weed control substance, by Pseudomonas koreensis and its plant growth-promoting and termiticidal activities. J. Environ. Sci. Int. 2018, 27, 771–780. [Google Scholar] [CrossRef]
- Soenens, A.; Imperial, J. Biocontrol capabilities of the genus Serratia. Phytochem. Rev. 2020, 19, 577–587. [Google Scholar] [CrossRef]
- Ordentlich, A.; Elad, Y.; Chet, I. The role of chitinase of Serratia marcescens in biocontrol of Sclerotium rolfsii. Phytopathology 1988, 78, 84–88. [Google Scholar]
- Levenfors, J.J.; Hedman, R.; Thaning, C.; Gerhardson, B.; Welch, C.J. Broad-spectrum antifungal metabolites produced by the soil bacterium Serratia plymuthica A 153. Soil Biol. Biochem. 2004, 36, 677–685. [Google Scholar] [CrossRef]
- Matilla, M.A.; Drew, A.; Udaondo, Z.; Krell, T.; Salmond, G.P. Genome sequence of Serratia plymuthica A153, a model rhizobacterium for the investigation of the synthesis and regulation of haterumalides, zeamine, and andrimid. Genome Announc. 2016, 4, e00373-16. [Google Scholar] [CrossRef] [Green Version]
- Das, P.; Effmert, U.; Baermann, G.; Quella, M.; Piechulla, B. Impact of bacterial volatiles on phytopathogenic fungi: An in vitro study on microbial competition and interaction. J. Exp. Bot. 2022, 73, 596–614. [Google Scholar] [CrossRef]
- Wenke, K.; Wanke, D.; Kilian, J.; Berendzen, K.; Harter, K.; Piechulla, B. Volatiles of two growth-inhibiting rhizobacteria commonly engage AtWRKY18 function. Plant J. 2012, 70, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kesel, J.; Conrath, U.; Flors, V.; Luna, E.; Mageroy, M.H.; Mauch-Mani, B.; Pastor, V.; Pozo, M.J.; Pieterse, C.M.; Ton, J. The induced resistance lexicon: Do’s and don’ts. Trends Plant Sci. 2021, 26, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, B.N.; Brooks, D.M. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol. 2002, 5, 325–331. [Google Scholar] [CrossRef]
- Ma, F.; Yang, X.; Shi, Z.; Miao, X. Novel crosstalk between ethylene-and jasmonic acid-pathway responses to a piercing–sucking insect in rice. New Phytol. 2020, 225, 474–487. [Google Scholar] [CrossRef] [Green Version]
- Vlot, A.C.; Dempsey, D.M.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Hua, G.K.H.; Ongena, M.; Höfte, M. Role of phenazines and cyclic lipopeptides produced by Pseudomonas sp. CMR12a in induced systemic resistance on rice and bean. Environ. Microbiol. Rep. 2016, 8, 896–904. [Google Scholar] [CrossRef]
- Audenaert, K.; Pattery, T.; Cornelis, P.; Höfte, M. Induction of systemic resistance to Botrytis cinerea in tomato by Pseudomonas aeruginosa 7NSK2: Role of salicylic acid, pyochelin, and pyocyanin. Mol. Plant Microbe Interact. 2002, 15, 1147–1156. [Google Scholar] [CrossRef] [Green Version]
- Kang, B.-R.; Han, S.-H.; Zdor, R.E.; Anderson, A.J.; Spencer, M.; Yang, K.-Y.; Kim, Y.-H.; Lee, M.-C.; Cho, B.-H.; Kim, Y.-C. Inhibition of seed germination and induction of systemic disease resistance by Pseudomonas chlororaphis O6 requires phenazine production regulated by the global regulator, GacS. J. Microbiol. Biotechnol. 2007, 17, 586–593. [Google Scholar]
- Spencer, M.; Ryu, C.-M.; Yang, K.-Y.; Kim, Y.C.; Kloepper, J.W.; Anderson, A.J. Induced defence in tobacco by Pseudomonas chlororaphis strain O6 involves at least the ethylene pathway. Physiol. Mol. Plant Pathol. 2003, 63, 27–34. [Google Scholar] [CrossRef]
- Ramarathnam, R.; Fernando, W.; de Kievit, T. The role of antibiosis and induced systemic resistance, mediated by strains of Pseudomonas chlororaphis, Bacillus cereus and B. amyloliquefaciens, in controlling blackleg disease of canola. BioControl 2011, 56, 225–235. [Google Scholar] [CrossRef]
- Martin-Rivilla, H.; Garcia-Villaraco, A.; Ramos-Solano, B.; Gutierrez-Manero, F.; Lucas, J. Improving flavonoid metabolism in blackberry leaves and plant fitness by using the bioeffector Pseudomonas fluorescens N 21.4 and its metabolic elicitors: A biotechnological approach for a more sustainable crop. J. Agric. Food Chem. 2020, 68, 6170–6180. [Google Scholar] [CrossRef] [PubMed]
- Chae, D.-H.; Kim, D.-R.; Cheong, M.S.; Lee, Y.B.; Kwak, Y.-S. Investigating the induced systemic resistance mechanism of 2,4-Diacetylphloroglucinol (DAPG) using DAPG hydrolase-transgenic Arabidopsis. Plant Pathol. J. 2020, 36, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, J.M.; Weller, D.M. Natural plant protection by 2,4-diacetylphloroglucinol-producing Pseudomonas spp. in take-all decline soils. Mol. Plant Microbe Interact. 1998, 11, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Cofer, T.M.; Seidl-Adams, I.; Tumlinson, J.H. From acetoin to (Z)-3-hexen-1-ol: The diversity of volatile organic compounds that induce plant responses. J. Agric. Food Chem. 2018, 66, 11197–11208. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Volatile 1-octen-3-ol induces a defensive response in Arabidopsis thaliana. J. Gen. Plant Pathol. 2007, 73, 35–37. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Kloepper, J.W.; Paré, P.W. Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.M.; Kang, B.R.; Han, S.H.; Anderson, A.J.; Park, J.-Y.; Lee, Y.-H.; Cho, B.H.; Yang, K.-Y.; Ryu, C.-M.; Kim, Y.C. 2R, 3R-butanediol, a bacterial volatile produced by Pseudomonas chlororaphis O6, is involved in induction of systemic tolerance to drought in Arabidopsis thaliana. Mol. Plant Microbe Interact. 2008, 21, 1067–1075. [Google Scholar] [CrossRef] [Green Version]
- Lam, V.B.; Meyer, T.; Arias, A.A.; Ongena, M.; Oni, F.E.; Höfte, M. Bacillus cyclic lipopeptides iturin and fengycin control rice blast caused by Pyricularia oryzae in potting and acid sulfate soils by direct antagonism and induced systemic resistance. Microorganisms 2021, 9, 1441. [Google Scholar] [CrossRef]
- Park, K.; Park, Y.-S.; Ahamed, J.; Dutta, S.; Ryu, H.; Lee, S.-H.; Balaraju, K.; Manir, M.; Moon, S.-S. Elicitation of induced systemic resistance of chili pepper by iturin A analogs derived from Bacillus vallismortis EXTN-1. Can. J. Plant Sci. 2016, 96, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Pang, X.; Liu, H.; Lin, F.; Lu, F.; Bie, X.; Lu, Z.; Lu, Y. Iturin A induces resistance and improves the quality and safety of harvested cherry tomato. Molecules 2021, 26, 6905. [Google Scholar] [CrossRef] [PubMed]
- Legrand, M.; Kauffmann, S.; Geoffroy, P.; Fritig, B. Biological function of pathogenesis-related proteins: Four tobacco pathogenesis-related proteins are chitinases. Proc. Natl. Acad. Sci. USA 1987, 84, 6750–6754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusztahelyi, T. Chitin and chitin-related compounds in plant–fungal interactions. Mycology 2018, 9, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Merzendorfer, H.; Zimoch, L. Chitin metabolism in insects: Structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 2003, 206, 4393–4412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Peng, D. Nematode chitin and application. Adv. Exp. Med. Biol. 2019, 1142, 209–219. [Google Scholar] [PubMed]
- Kilani, J.; Fillinger, S. Phenylpyrroles: 30 years, two molecules and (nearly) no resistance. Front. Microbiol. 2016, 7, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Yue, S.-J.; Liu, W.-H.; Zheng, Y.-F.; Zhang, C.-H.; Feng, T.-T.; Hu, H.-B.; Wang, W.; Zhang, X.-H. Engineering of glycerol utilization in Pseudomonas chlororaphis GP72 for enhancing phenazine-1-carboxylic acid production. World J. Microbiol. Biotechnol. 2020, 36, 49. [Google Scholar] [CrossRef]
- Aggarwal, N.; Thind, S.; Sharma, S. Role of secondary metabolites of Actinomycetes in crop protection. In Plant Growth Promoting Actinobacteria; Springer: Berlin/Heidelberg, Germany, 2016; pp. 99–121. [Google Scholar]
- Bargabus, R.; Zidack, N.; Sherwood, J.; Jacobsen, B. Characterisation of systemic resistance in sugar beet elicited by a non-pathogenic, phyllosphere-colonizing Bacillus mycoides, biological control agent. Physiol. Mol. Plant Pathol. 2002, 61, 289–298. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Jeffers, S.N. Efficacy of commercial formulation of two biofungicides for control of blue mold and gray mold of apples in cold storage. Crop Prot. 1997, 16, 629–633. [Google Scholar] [CrossRef]
- Abbey, J.A.; Percival, D.; Abbey, L.; Asiedu, S.K.; Prithiviraj, B.; Schilder, A. Biofungicides as alternative to synthetic fungicide control of grey mould (Botrytis cinerea)–prospects and challenges. Biocontrol Sci. Technol. 2019, 29, 207–228. [Google Scholar] [CrossRef]
- Matzen, N.; Heick, T.M.; Jørgensen, L.N. Control of powdery mildew (Blumeria graminis spp.) in cereals by Serenade® ASO (Bacillus amyloliquefaciens (former subtilis) strain QST 713). Biol. Control 2019, 139, 104067. [Google Scholar] [CrossRef]
- Kilani-Feki, O.; Khedher, S.B.; Dammak, M.; Kamoun, A.; Jabnoun-Khiareddine, H.; Daami-Remadi, M.; Tounsi, S. Improvement of antifungal metabolites production by Bacillus subtilis V26 for biocontrol of tomato postharvest disease. Biol. Control 2016, 95, 73–82. [Google Scholar] [CrossRef]
- Liu, S.; Tang, M.H.; Cheng, J.S. Fermentation optimization of surfactin production of Bacillus amyloliquefaciens HM618. Biotechnol. Appl. Biochem. 2022, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Hur, J.Y.; Park, S.K. Biocontrol of Botrytis cinerea by chitin-based cultures of Paenibacillus elgii HOA73. Euro. J. Plant Pathol. 2019, 155, 253–263. [Google Scholar] [CrossRef]
- Park, J.Y.; Kang, B.R.; Ryu, C.M.; Anderson, A.J.; Kim, Y.C. Polyamine is a critical determinant of Pseudomonas chlororaphis O6 for GacS-dependent bacterial cell growth and biocontrol capacity. Mol. Plant Pathol. 2018, 19, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- Ghribi, D.; Mnif, I.; Boukedi, H.; Kammoun, R.; Ellouze-Chaabouni, S. Statistical optimization of low-cost medium for economical production of Bacillus subtilis biosurfactant, a biocontrol agent for the olive moth Prays oleae. Afr. J. Microbiol. Res. 2011, 5, 4927–4936. [Google Scholar]
- Meena, K.R.; Sharma, A.; Kumar, R.; Kanwar, S.S. Two factor at a time approach by response surface methodology to aggrandize the Bacillus subtilis KLP2015 surfactin lipopeptide to use as antifungal agent. J. King Saud Univ. Sci. 2020, 32, 337–348. [Google Scholar] [CrossRef]
- Ghribi, D.; Abdelkefi-Mesrati, L.; Mnif, I.; Kammoun, R.; Ayadi, I.; Saadaoui, I.; Maktouf, S.; Chaabouni-Ellouze, S. Investigation of antimicrobial activity and statistical optimization of Bacillus subtilis SPB1 biosurfactant production in solid-state fermentation. J. Biomed. Biotechnol. 2012, 2012, 373682. [Google Scholar] [CrossRef] [Green Version]
- Philip, N.V.; Koteshwara, A.; Kiran, G.A.; Raja, S.; Subrahmanyam, V.; Chandrashekar, H.R. Statistical optimization for coproduction of chitinase and beta 1, 4-endoglucanase by chitinolytic Paenibacillus elgii PB1 having antifungal activity. Appl. Biochem. Biotechnol. 2020, 191, 135–150. [Google Scholar] [CrossRef]
- Gao, H.; Liu, M.; Liu, J.; Dai, H.; Zhou, X.; Liu, X.; Zhuo, Y.; Zhang, W.; Zhang, L. Medium optimization for the production of avermectin B1a by Streptomyces avermitilis 14-12A using response surface methodology. Bioresour. Technol. 2009, 100, 4012–4016. [Google Scholar] [CrossRef]
- Kulkarni, M.; Gorthi, S.; Banerjee, G.; Chattopadhyay, P. Production, characterization and optimization of actinomycin D from Streptomyces hydrogenans IB310, an antagonistic bacterium against phytopathogens. Biocatal. Agric. Biotechnol. 2017, 10, 69–74. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, Z.; Zhang, G.; Mo, X.; Ding, X.; Xia, L.; Hu, S. A rifampicin-resistant (rpoB) mutation in Pseudomonas protegens Pf-5 strain leads to improved antifungal activity and elevated production of secondary metabolites. Res. Microbiol. 2016, 167, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.-Y.; Liu, Q.; Choi, J.-Y.; Wang, Y.; Shim, H.-J.; Xu, H.G.; Choi, G.-J.; Kim, J.-C.; Je, Y.-H. Construction of a recombinant Bacillus velezensis strain as an integrated control agent against plant diseases and insect pests. J. Microbiol. Biotechnol. 2009, 19, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Mavrodi, D.V.; Yang, M.; Thomashow, L.S.; Mavrodi, O.V.; Kelton, J.; Weller, D.M. Pseudomonas synxantha 2-79 transformed with pyrrolnitrin biosynthesis genes has improved biocontrol activity against soilborne pathogens of wheat and canola. Phytopathology 2020, 110, 1010–1017. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Hu, H.; Wang, W.; Zhang, X. Genetic engineering of Pseudomonas chlororaphis GP72 for the enhanced production of 2-hydroxyphenazine. Microb. Cell Factories 2016, 15, 131. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, Z.; Yao, W.; Zhang, X.; Wang, R.; Li, P.; Yang, K.; Wang, T.; Liu, K. Metabolic engineering of Pseudomonas chlororaphis Qlu-1 for the enhanced production of phenazine-1-carboxamide. J. Agric. Food Chem. 2020, 68, 14832–14840. [Google Scholar] [CrossRef]
- Wan, Y.; Liu, H.; Xian, M.; Huang, W. Biosynthesis and metabolic engineering of 1-hydroxyphenazine in Pseudomonas chlororaphis H18. Microb. Cell Factories 2021, 20, 235. [Google Scholar] [CrossRef]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.-F. PTI-ETI crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef]
- Akhtar, M.S.; Siddiqui, Z.A. Glomus intraradices, Pseudomonas alcaligenes, and Bacillus pumilus: Effective agents for the control of root-rot disease complex of chickpea (Cicer arietinum L.). J. Gen. Plant Pathol. 2008, 74, 53–60. [Google Scholar] [CrossRef]
Figure 1.
The multiple impacts of biocontrol-active bacteria in the rhizosphere and extending out into bulk soil. Biocontrol microbes are accepted as plant root colonists while exerting antagonism against plant pests through several layers of control in the rhizosphere space. Defense by induced systemic resistance (ISR) may be triggered by metabolites from the biocontrol root colonists. The patchy biofilms on the root surface provide protective barriers. Aqueous diffusion in soil pore waters distributes antagonistic metabolites and enzymes secreted by the biocontrol colonists further into the rhizosphere and biocontrol-active volatiles will diffuse to bulk soil, even to the airspace. These mechanisms boost the survival of the biocontrol bacteria, with their protection within biofilms and spread through chemotaxis and swarming at the rhizoplane. Movement of infected nematodes and insect larvae transport inocula away from the root, and these structures act as hot spots for production of antagonistic products from the biocontrol microbes.
Figure 1.
The multiple impacts of biocontrol-active bacteria in the rhizosphere and extending out into bulk soil. Biocontrol microbes are accepted as plant root colonists while exerting antagonism against plant pests through several layers of control in the rhizosphere space. Defense by induced systemic resistance (ISR) may be triggered by metabolites from the biocontrol root colonists. The patchy biofilms on the root surface provide protective barriers. Aqueous diffusion in soil pore waters distributes antagonistic metabolites and enzymes secreted by the biocontrol colonists further into the rhizosphere and biocontrol-active volatiles will diffuse to bulk soil, even to the airspace. These mechanisms boost the survival of the biocontrol bacteria, with their protection within biofilms and spread through chemotaxis and swarming at the rhizoplane. Movement of infected nematodes and insect larvae transport inocula away from the root, and these structures act as hot spots for production of antagonistic products from the biocontrol microbes.

Figure 2.
Classes of metabolites involved in biocontrol of microbial, insect, and nematode pests for plants. The biocontrol-active rhizobacteria are highly selective pathogens causing death, or impaired growth, of plant pests through the production of soluble and volatile metabolites, enzymes, and the formation of biofilms. Additionally, microbially associated molecular patterns (MAMPs) and certain metabolites will trigger plant defense systems.
Figure 2.
Classes of metabolites involved in biocontrol of microbial, insect, and nematode pests for plants. The biocontrol-active rhizobacteria are highly selective pathogens causing death, or impaired growth, of plant pests through the production of soluble and volatile metabolites, enzymes, and the formation of biofilms. Additionally, microbially associated molecular patterns (MAMPs) and certain metabolites will trigger plant defense systems.

Table 2.
Minimum inhibitory concentration (MIC) and lethal dose (LD)/lethal concentration (LC) 50 of secondary metabolites from microbials against fungal pathogens; lethal dose or concentration against pathogenic insects or nematodes.
Table 2.
Minimum inhibitory concentration (MIC) and lethal dose (LD)/lethal concentration (LC) 50 of secondary metabolites from microbials against fungal pathogens; lethal dose or concentration against pathogenic insects or nematodes.
| Active Metabolite | Pathogen/Pest | MIC (ppm) | LD/LC 50 (ppm) | Inhibition | Source of Active Metabolite | Reference |
|---|---|---|---|---|---|---|
| Lipopeptide | Tuta absoluta | 180 ng/cm2 | 1st Instar larvae mortality | Bacillus amyloliquefaciens AG1 | [12] | |
| Lipopeptide | Fusarium oxysporum f. sp. lycopersici | 10 | Mycelial growth | Bacillus amyloliquefaciens BO5A | [81] | |
| Lipopeptide | Myzus persicae | 22.2 | 2nd Instar nymph mortality | Bacillus subtilis Y9 | [82] | |
| Lipopeptide | Ectomyelois ceratoniae | 152 | 3rd Instar larvae morality | Bacillus subtilis SPB1 | [18] | |
| Lipopeptide | Spodoptera littoralis | 251 ng/cm2 | 1st Instar larvae mortality | Bacillus subtilis SPB1 | [17] | |
| Lipopeptide | Fusarium solani | 3000 | Mycelial growth | Bacillus subtilis SPB1 | [19,20] | |
| Rhizoctonia bataticola | 40 | |||||
| Rhizoctonia solani | 4000 | |||||
| Lipopeptide | Colletotrichum acutatum | 32 | Mycelial growth | Bacillus subtilis EA-CB0015 | [78] | |
| Lipopeptide | Colletotrichum gloeosporioides | 36.47 | Mycelial growth | Bacillus amyloliquefaciens MG3 | [79] | |
| Lipopeptide | Phoma tracheiphila | 47.5 | Mycelial growth | Bacillus methyltrophicus TEB1 | [80] | |
| Phenazine-1-carboxylic acid (PCA) | Botrytis cinerea | 25 | Mycelial growth | Pseudomonas aeruginosa LV | [83] | |
| PCA | Botrytis cinerea | 50 | ||||
| Colletotrichum orbiculare | 5 | Mycelial growth | Pseudomonas aeruginosa GC-B26 | [84] | ||
| Phytophthora capsici | 5 | |||||
| Pythium ultimum | 5 | |||||
| PCA | Sclerotium rolfsii | 29 | Mycelial growth | Pseudomonas aeruginosa | [85] | |
| Fusarium oxysporum | 40 | |||||
| Colletotrichum falcatum | 50 | |||||
| PCA | Fusarium oxysporum | 1.56 | Mycelial growth | Burkholderia sp.HQB-1 | [86] | |
| Colletotrichum gloeosporioides | 6.13 | |||||
| Botrytis cinerea | 1.56 | |||||
| Curvularia fallax | 3.13 | |||||
| PCA | Gaeumannomyces graminis var. tritici | 1 | Mycelial growth | Pseudomonas fluorescens 2–79 | [87] | |
| Rhizoctonia solani | 1 | |||||
| Cochliobolus sativus | 1–3 | |||||
| Pythium aristosporum | 1 | |||||
| Pythium ultimum | 25–30 | |||||
| Pythium uttimum var. sporangiifurum | 80–100 | |||||
| Fusarium sp. | 25–30 | |||||
| Phenazine-1-carboxamide (PCN) | Botrytis cinerea | 108.12 * | Mycelial growth | Pseudomonas aeruginosa | [88] | |
| PCN | Rhizoctonia solani | 5 | Mycelial growth | Pseudomonas aeruginosa MML2212 | [89] | |
| 1-Octen-3-ol | Tribolium castaneum | 16.75 | Adult mortality | Paenibacillus polymyxa BMP-11 | [30] | |
| Benzothiazole | Tribolium castaneum | 3.5 | Adult mortality | Paenibacillus polymyxa BMP-11 | [30] | |
| 2,4-Diacetylphloroglucinol (2,4-DAPG) | Fusarium oxysporum f. sp. lycopersici | 16 | Mycelial growth | Pseudomonas fluorescens CHA0 | [90] | |
| Gaeumannomyces graminis var. tritici | 16–32 | |||||
| Pythium ultimum | 64 | |||||
| Rhizoctonia solani | 32–64 | |||||
| 2,4-DAPG | Xiphinema americanum | 8.3 | Adult mortality | Pseudomonas fluorescens | [91] |
* EC50 (ppm).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, J.H.; Anderson, A.J.; Kim, Y.C. Root-Associated Bacteria Are Biocontrol Agents for Multiple Plant Pests. Microorganisms 2022, 10, 1053. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051053
AMA Style
Lee JH, Anderson AJ, Kim YC. Root-Associated Bacteria Are Biocontrol Agents for Multiple Plant Pests. Microorganisms. 2022; 10(5):1053. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051053
Chicago/Turabian StyleLee, Jang Hoon, Anne J. Anderson, and Young Cheol Kim. 2022. "Root-Associated Bacteria Are Biocontrol Agents for Multiple Plant Pests" Microorganisms 10, no. 5: 1053. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051053
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.