
Root-Knot Disease Suppression in Eggplant Based on Three Growth Ages of Ganoderma lucidum

 ,
,  , , ,
, , ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection, Identification and Microscopic Analysis of G. lucidum
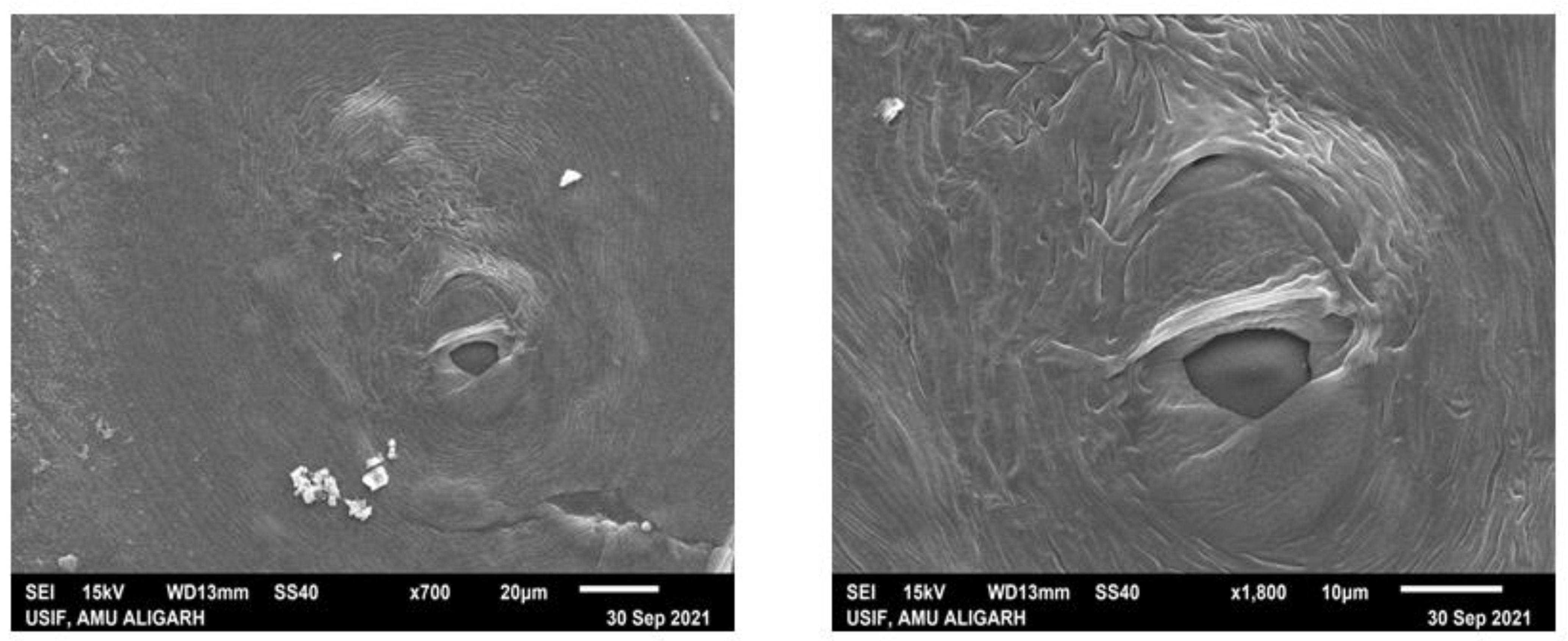
2.2. Scanning Electron Microscopy of Basidiocarp (Pileal Surface and Stipe) of G. lucidum
2.3. Cultivation and Maintenance of G. lucidum
2.4. GC-MS Analysis of G. lucidum
2.4.1. Sample Preparation
2.4.2. Gas Chromatography-Mass Spectrometry (GC-MS) Conditions
2.5. Inoculums Maintenance and Species Identification of M. incognita
2.6. Inoculum Preparation of M. incognita Juveniles
2.7. Toxicity Testing of G. lucidum against M. incognita Juveniles: In Vitro
2.7.1. Preparation of Culture Filtrates (CFs) of G. lucidum
2.7.2. Inhibition in Hatching of M. incognita Eggs Exposed to the CFs of G. lucidum
2.7.3. Mortality and Immobility of J2 of M. incognita Exposed to the CFs of G. lucidum
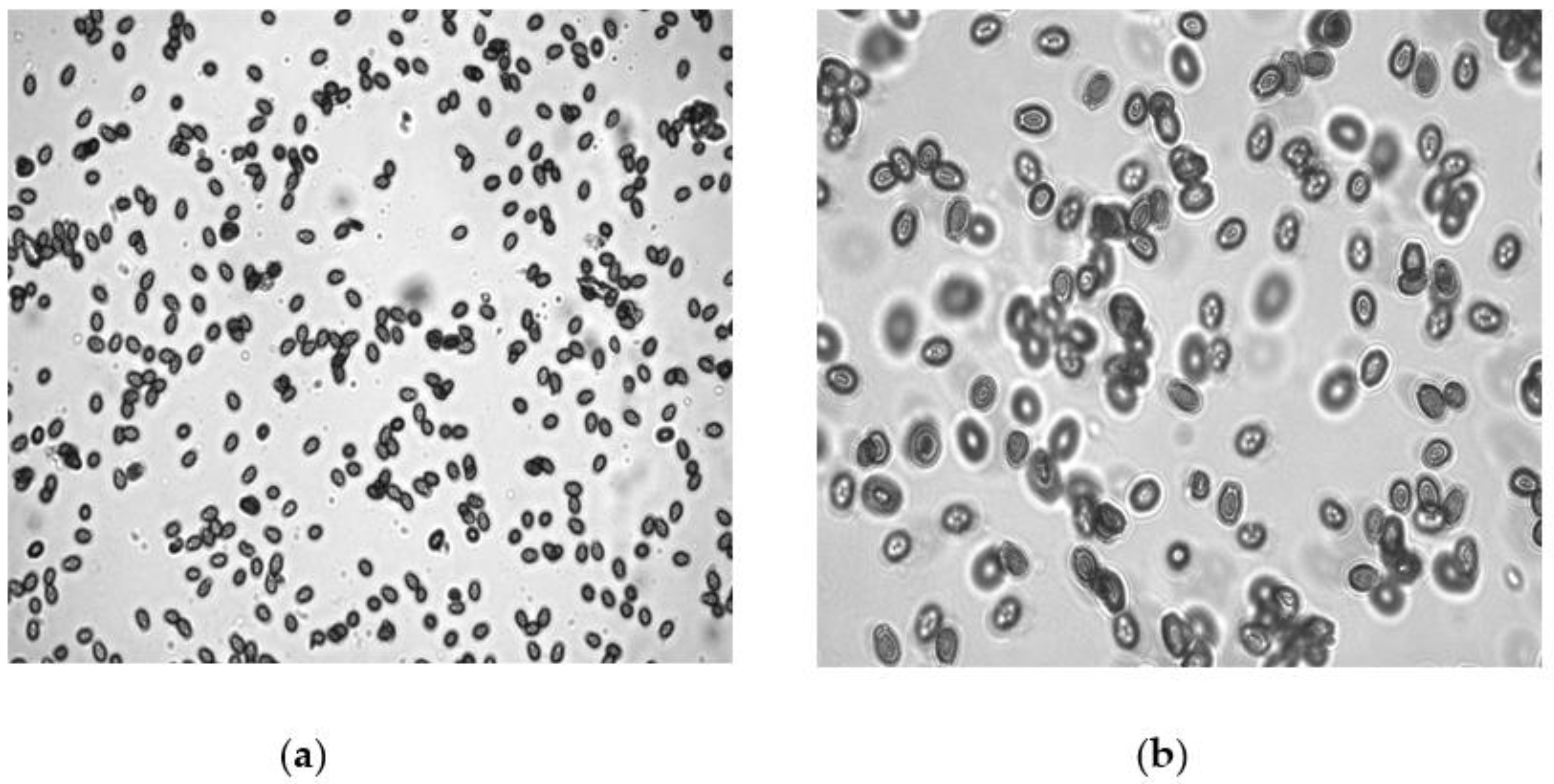
2.8. SEM of Treated Juveniles (TJ) and Untreated Juveniles (UTJ) of M. incognita
2.9. In Planta Study (Pot Experiment)
2.10. Data Collection and Observations
2.11. Statistical Analysis
3. Results
3.1. Effect of Different Concentrations of G. lucidum CFs on the Egg Hatching of M. incognita
3.2. Effect of Different Concentrations of G. lucidum CFs on the Mortality of M. incognita
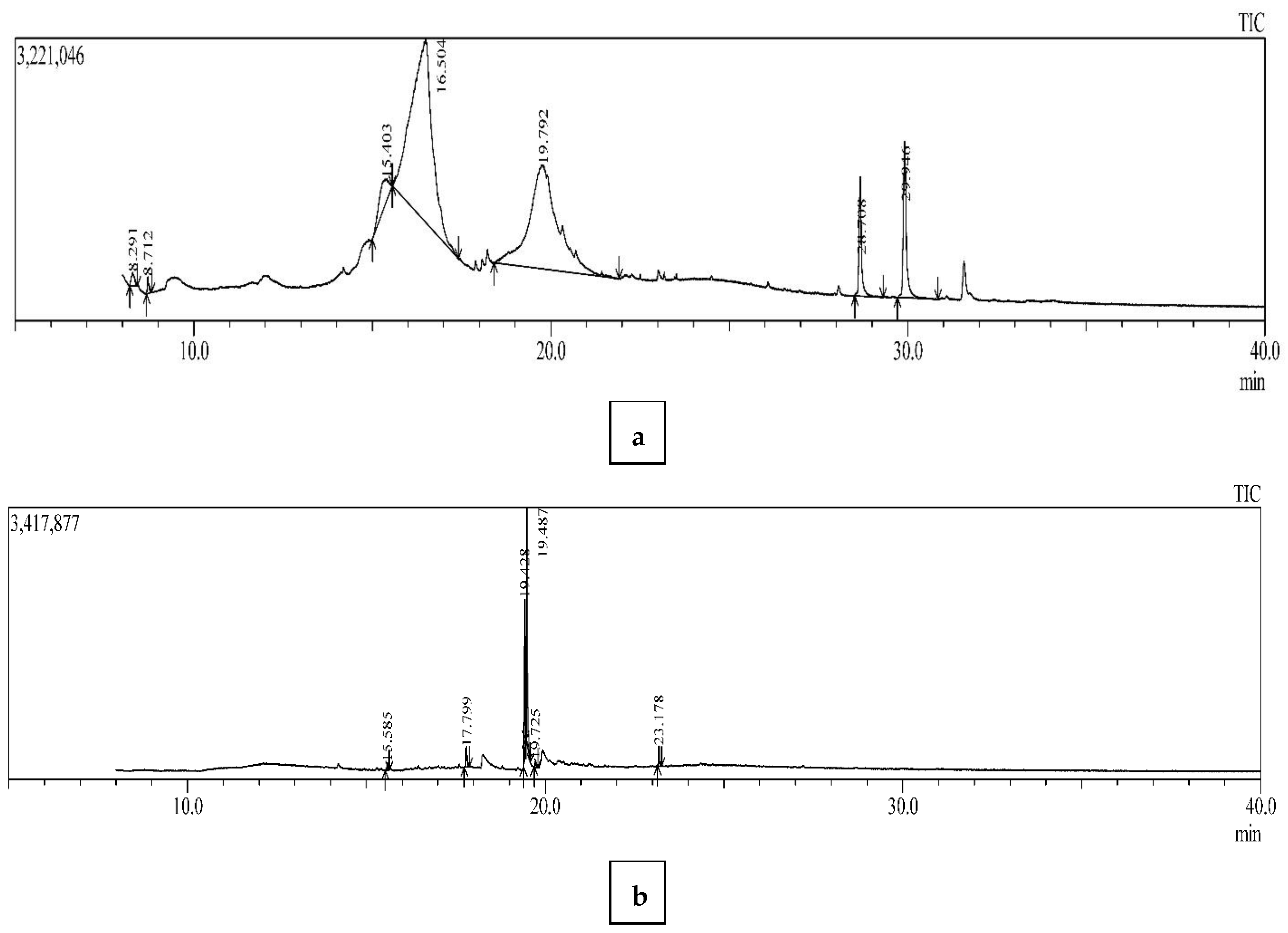
3.3. GC-MS Analysis for Bioactive Compounds
3.4. SEM of the Treated Juveniles (TJ) and Untreated Juveniles (UTJ)
3.5. Effect of Root Dip Treatment with CFs of G. lucidum on Multiplication of M. incognita Parasitized Eggplant (in Planta Study)
3.6. Effects of Root Dip Treatment with CFs of G. lucidum on Eggplant Growth
3.7. Effect of CFs of G. lucidum on the Physiology of Eggplant
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kandoliya, U.K.; Bajaniya, V.K.; Bhadja, N.K.; Bodar, N.P.; Golakiya, B.A. Antioxidant and nutritional components of eggplant (Solanum melongena L.) fruit grown in Saurastra region. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 806–813. [Google Scholar]
- FAOSTAT. Production of Lentils, “Crops/World Regions/Production Quantity, from Picklists”. 2020. Available online: http://faostat3.fao.org/browse/Q/QC/E (accessed on 22 January 2022).
- Abd-Elgawad, M.M. Biological control of nematodes infecting eggplant in Egypt. Bull. Nat. Res. Cent. 2021, 45, 6. [Google Scholar] [CrossRef]
- Peng, D.; Chen, Q. Nematode chitin and application. Adv. Exp. Med. Biol. 2019, 1142, 209–219. [Google Scholar]
- Kumar, A.; Patil, J.A.; Yadav, S.; Ram, S. Screening, confirmation and field evaluation of promising resistant germplasm of different pulses against root knot nematode, Meloidogyne javanica. J. Environ. Biol. 2020, 41, 1594–1598. [Google Scholar] [CrossRef]
- Chitwood, D.J. Phytochemical based strategies for nematode control. Ann. Rev. Phytopathol. 2002, 40, 221–249. [Google Scholar] [CrossRef] [Green Version]
- Xiang, N.; Lawrence, K.S.; Donald, P.A. Biological control potential of plant growth-promoting rhizobacteria suppression of Meloidogyne incognita on cotton and Heterodera glycines on soybean: A review. J. Phytopathol. 2018, 166, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Wasser, S.P. Current findings, future trends, and unsolved problems in studies of medicinal mushrooms. App. Microbiol. Biotech. 2011, 89, 1323–1332. [Google Scholar] [CrossRef]
- Li, G.H.; Yu, Z.F.; Li, X.; Wang, X.B.; Zheng, L.J.; Zhang, K.Q. Nematicidal metabolites produced by the endophytic fungus Geotrichum sp. AL4. Chem. Biodiv. 2007, 4, 1520–1524. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; David, J.C.; Stalpers, J.A. Ainsworth and Bisby’s Dictionary of the Fungi, 10th ed.; CABI Publishing: Wallingford, UK, 2008; p. 771. [Google Scholar]
- Ahmad, M.F. Ganoderma lucidum: Persuasive biologically active constituents and their health endorsement. Biomed. Pharmacol. 2018, 107, 507–519. [Google Scholar] [CrossRef]
- Sanodiya, B.S.; Thakur, G.S.; Baghel, R.K.; Prasad, G.B.K.S.; Bisen, P.S. Ganoderma lucidum: A potent pharmacological macrofungus. Curr. Pharma. Biotechnol. 2009, 10, 717–742. [Google Scholar] [CrossRef]
- Smith, J.; Rowan, N.; Sullivan, R. Medicinal Mushrooms: Their Therapeutic Properties and Current Medical Usage with Special Emphasis on Cancer Treatment; Special Report Commissioned by Cancer Research UK, The University of Strathclyde in Glasgow; Johns Hopkins University Press: Baltimore, MD, USA, 2002; p. 256. [Google Scholar]
- Gao, Y.; Zhou, S.H.; Huang, M.; Xu, A. Antibacterial and antiviral value of the genus Ganoderma P. Karst. species (Aphyllophoromycetideae): A review. Int. J. Med. Mushrooms 2003, 5, 235–246. [Google Scholar] [CrossRef]
- Wang, H.; Ng, T.B. Ganodermin, an antifungal protein from fruiting bodies of the medicinal mushroom Ganoderma lucidum. Peptides 2006, 27, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.S.; Kao, C.H.; Xu, Y.; Glucina, M.P.; Paterson, R.R.M.; Ferguson, L.R. From 2000 years of Ganoderma lucidum to recent developments in nutraceuticals. Phytochem 2015, 114, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Fan, J.; Liu, Y.; Guo, W.; Cao, H.; Xiao, J.; Liu, B. Hepatoprotective activity of Ganoderma lucidum triterpenoids in alcohol-induced liver injury in mice, an iTRAQ-based proteomic analysis. Food Chem. 2019, 271, 148–156. [Google Scholar] [CrossRef]
- Ahmad, M.F. Ganoderma lucidum: A rational pharmacological approach to surmount cancer. J. Ethnopharmacol. 2020, 260, 113047. [Google Scholar] [CrossRef]
- Ren, L.; Zhang, J.; Zhang, T. Immunomodulatory activities of polysaccharides from Ganoderma on immune effector cells. Food Chem. 2021, 340, 127933. [Google Scholar] [CrossRef]
- Akshaya, S.B.; Krishnamoorthy, A.S.; Nakkeeran, S.; Poornima, K.; Sivakumar, U. Inhibitory potential of ethyl acetate extract from mushrooms against root-knot nematode (Meloidogyne incognita). J. Entomol. Zool. Stud. 2021, 9, 528–534. [Google Scholar]
- Themuhi, M.; Shanthi, A.; Krishnamoorthy, A.S.; Swarnakumari, N.; Amirtham, D. In vitro screening of macro basidiomycetous fungi against root-knot nematode, Meloidogyne incognita. J. Entomol. Zool. Stud. 2020, 8, 641–644. [Google Scholar] [CrossRef]
- Wang, X.C.; Xi, R.J.; Li, Y.; Wang, D.M.; Yao, Y.J. The species identity of the widely cultivated Ganoderma lucidum (Ling-zhi), in China. PLoS ONE 2012, 7, 40857. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.B.; Yao, Y.J. Boletus reticuloceps, a new combination for Aureoboletus reticuloceps. Sydowia-Horn 2005, 57, 131–136. [Google Scholar]
- Singh, S.; Kuca, K.; Kalia, A. Alterations in Growth and Morphology of Ganoderma lucidum and Volvariella volvaceae in Response to Nanoparticle Supplementation. Mycobiology 2020, 48, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Satria, D.; Amen, Y.; Niwa, Y.; Ashour, A.; Allam, A.E.; Shimizu, K. Lucidumol D, a new lanostane-type triterpene from fruiting bodies of Reishi (Ganoderma lingzhi). Nat. Prod. Res. 2019, 33, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.K.; Adnan, M.; Cho, D.H. A network pharmacology analysis on drug-like compounds from Ganoderma lucidum for alleviation of atherosclerosis. J. Food Biochem. 2021, 45, e13906. [Google Scholar] [CrossRef] [PubMed]
- Eisenback, J.D. Detailed morphology and anatomy of second-stage juveniles, males, and females of the genus Meloidogyne (root-knot nematodes). In An Advanced Treatise on Meloidogyne; Sasser, J.N., Carter, C.C., Eds.; Biology and Control, North Carolina State University Graphics: Raleigh, NC, USA, 1985; Volume 1, pp. 47–77. [Google Scholar]
- El-Rokiek, K.G.; El-Nagdi, W.M. Dual effects of leaf extracts of Eucalyptus citriodora on controlling purslane and root-knot nematode in sunflower. J. Plant Prot. Res. 2011, 51, 121–129. [Google Scholar] [CrossRef]
- Kiontke, K.; Manegold, A.; Sudhaus, W. Redescription of Diplogasteroides nasuensis Takaki, 1941 and D. magnus Volk, 1950 (Nematoda: Diplogastrina) associated with Scarabaeidae (Coleoptera). Nematology 2001, 3, 817–832. [Google Scholar]
- Daykin, M.E.; Hussey, R.S. Staining and Histopathological techniques in nematology. In An Advanced Treatise on Meloidogyne, Methodology, Raleigh; Baker, K.R., Carter, C.C., Sasser, J.N., Eds.; North Carolina State University Graphics: Elizabeth City, NC, USA, 1985; Volume II, pp. 39–48. [Google Scholar]
- Cobb, N.A. Estimating the Nema Populations of the Soil. Agriculture Technical Circulation Bulletin of Plant Industries; US Department of Agriculture: Washington, DC, USA, 1918; p. 48.
- Southey, J.F. Laboratory Methods for Work with Plant and Soil Nematodes; HMSO: London, UK, 1986.
- MacKinney, G. Absorption of light by chlorophyll solutions. J. Biol. Chem. 1941, 140, 315–322. [Google Scholar] [CrossRef]
- Smania, J.R.A.; Monache, F.D.; Smania, E.D.F.A.; Cuneo, R.S. Antibacterial activity of steroidal compounds isolated from Ganoderma applanatum (Pers.) Pat. (Aphyllophoromycetideae) fruit body. Int. J. Med. Mush. 1999, 1, 325–330. [Google Scholar] [CrossRef]
- Boh, B.; Berovic, M.; Zhang, J.; Zhi-Bin, L. Ganoderma lucidum and its pharmaceutically active compounds. Biotech. Ann. Rev. 2007, 13, 265–301. [Google Scholar]
- Zhou, X.; Lin, J.; Yin, Y.; Zhao, J.; Sun, X.; Tang, K. Ganodermataceae: Natural products and their related pharmacological functions. Amer. J. Chin. Med. 2007, 35, 559–574. [Google Scholar] [CrossRef]
- Bua-art, S.; Saksirirat, W.; Kanokmedhakul, S.; Hiransalee, A.; Lekphrom, R. Extraction of bioactive compounds from luminescent mushroom (Neonothopanus nambi) and its effect on Root-Knot Nematode (Meloidogyne incognita). Asia. Pac. J. Sci. Tech. 2010, 15, 726–737. [Google Scholar]
- Meyer, S.; Huettel, R.; Liu, X.Z.; Humber, R.; Juba, J.; Nitao, J. Activity of fungal culture filtrates against soybean cyst nematode and root-knot nematode egg hatch and juvenile motility. Nematology 2004, 6, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Akram, M.; Khan, M.R. Interaction of Meloidogyne incognita and Fusarium oxysporum f. sp. lycopersici on tomato. Ann. Plant Prot. Sci. 2006, 14, 448–451. [Google Scholar]
- Khalil, M.S.; Kenawy, A.; Gohrab, M.A.; Mohammed, E.E. Impact of microbial agents on Meloidogyne incognita management and morphogenesis of tomato. J. Biopest. 2012, 5, 28. [Google Scholar]
- Arya, R. Effect of culture filtrates of Epicoccum purpurascens and Penicillium vermiculatum grown on different carbon sources on hatching and mortality of larvae of root knot nematode (Meloidogyne incognita). Indian J. Pure Appl. Biosci. 2011, 26, 9–14. [Google Scholar]
- Zhao, D.; Zhu, X.; Chen, L.; Liu, W.; Chen, J.; Wang, S.; Liu, X. Toxicity of a secondary metabolite produced by Simplicillium chinense Snef5 against the root-knot nematode Meloidogyne incognita. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2020, 70, 550–555. [Google Scholar] [CrossRef]
- Innocenti, M.; Tenca, P.; Frittoli, E.; Faretta, M.; Tocchetti, A.; Di Fiore, P.P.; Scita, G. Mechanisms through which Sos-1 coordinates the activation of Ras and Rac. J. Cell Biol. 2002, 156, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badalyan, S.M.; Garibyan, N.G.; Innocenti, G. Antagonistic activity of xylotrophic mushrooms against pathogenic fungi of cereals in dual culture. Antagonistic activity of xylotrophic mushrooms against pathogenic fungi of cereals in dual culture. Phytopathol. Med. 2002, 41, 220–225. [Google Scholar]
- Sridhar, S.; Sivaprakasam, E.; Balakumar, R.; Kavitha, D. Evaluation of antibacterial and antifungal activity of Ganoderma lucidum (Curtis) P. Karst fruit bodies extracts. World J. Sci. Tech. 2011, 1, 8–11. [Google Scholar]
- Liu, X.; Xiang, M.; Che, Y. The living strategy of nematophagous fungi. Mycoscience 2009, 50, 20–25. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age of Fungus | Concentration (%) | Inhibition (%) |
|---|---|---|
| Two week | 100 | 58 (89.2) a |
| 50 | 146 (72.8) b | |
| 10 | 253 (53.0) c | |
| 1 | 409 (24.0) f | |
| Four week | 100 | 123 (77.1) b |
| 50 | 264 (51.0) c | |
| 10 | 360 (33.1) e | |
| 1 | 437(18.7) g | |
| Eight week | 100 | 237 (55.9) c |
| 50 | 328 (39.0) d | |
| 10 | 377 (29.9) e | |
| 1 | 462 (14.0) h | |
| Control | - | 538 (0.0) i |
| Age of Fungus | Concentration (%) | Inhibition (%) |
|---|---|---|
| Two week | 100 | 102 (81.0) a |
| 50 | 169 (68.5) b | |
| 10 | 312 (42.3) c | |
| 1 | 430 (20.0) f | |
| Four week | 100 | 166 (69.1) b |
| 50 | 355 (34.0) d | |
| 10 | 403 (25.1) e | |
| 1 | 458 (14.9) g | |
| Eight week | 100 | 296 (45.0) c |
| 50 | 371 (31.0) d | |
| 10 | 418 (22.3) f | |
| 1 | 497 (7.6) h | |
| Control | - | 538 (0.0) i |
| Age of Fungus | Exposure Period (h) | Concentration (%) | Juvenile Mortality (%) |
|---|---|---|---|
| Two week | 12 | 100 | 25.9 j |
| 50 | 18.9 n | ||
| 10 | 12.2 | ||
| 1 | 0.0 w | ||
| 24 | 100 | 40.5 f | |
| 50 | 28.6 i | ||
| 10 | 19.3 mn | ||
| 1 | 8.4 stu | ||
| 48 | 100 | 58.6 c | |
| 50 | 36.1 g | ||
| 10 | 23.6 kl | ||
| 1 | 8.1 tu | ||
| 72 | 100 | 83.8 a | |
| 50 | 54.3 d | ||
| 10 | 36.3 g | ||
| 1 | 20.9 mn | ||
| Four week | 12 | 100 | 19.5 mn |
| 50 | 11.4 qr | ||
| 10 | 0.0 w | ||
| 1 | 0.0 w | ||
| 24 | 100 | 24.4 kl | |
| 50 | 16.5 op | ||
| 10 | 7.0 u | ||
| 1 | 0.0 w | ||
| 48 | 100 | 39.9 d | |
| 50 | 27.7 ij | ||
| 10 | 15.9 p | ||
| 1 | 3.9 v | ||
| 72 | 100 | 67.3 b | |
| 50 | 39.0 f | ||
| 10 | 29.0 i | ||
| 1 | 15.0 p | ||
| Eight week | 12 | 100 | 0.0 w |
| 50 | 0.0 w | ||
| 10 | 0.0 w | ||
| 1 | 0.0 w | ||
| 24 | 100 | 16.7 op | |
| 50 | 9.1 rst | ||
| 10 | 2.1 vw | ||
| 1 | 0.0 w | ||
| 48 | 100 | 29.2 i | |
| 50 | 21.9 lm | ||
| 10 | 10.0 qrst | ||
| 1 | 0.0 w | ||
| 72 | 100 | 47.0 e | |
| 50 | 31.9 h | ||
| 10 | 19.6 mn | ||
| 1 | 11.0 qrs | ||
| Control | - | - | 0.0 w |
| Age of Fungus | Exposure Period (h) | Concentration (%) | Juvenile Mortality (%) |
|---|---|---|---|
| Two week | 12 | 100 | 23.8 j |
| 50 | 16.4 n | ||
| 10 | 9.1 qr | ||
| 1 | 0.0 x | ||
| 24 | 100 | 32.4 ef | |
| 50 | 19.5 lm | ||
| 10 | 13.0 o | ||
| 1 | 5.3 uv | ||
| 48 | 100 | 47.8 c | |
| 50 | 28.8 gh | ||
| 10 | 17.5 mn | ||
| 1 | 6.7 tu | ||
| 72 | 100 | 73.8 a | |
| 50 | 46.6 c | ||
| 10 | 30.9 fg | ||
| 1 | 16.9 n | ||
| Four week | 12 | 100 | 10.2 pqrs |
| 50 | 3.4 vw | ||
| 10 | 0.0 x | ||
| 1 | 0.0 x | ||
| 24 | 100 | 18.7 mn | |
| 50 | 10.4 pqr | ||
| 10 | 4.0 vw | ||
| 1 | 2.3 wx | ||
| 48 | 100 | 33.6 e | |
| 50 | 25.3 ij | ||
| 10 | 11.5 opq | ||
| 1 | 4.1 vw | ||
| 72 | 100 | 61.6 b | |
| 50 | 33.7 e | ||
| 10 | 21.3 kl | ||
| 1 | 13.0 o | ||
| Eight week | 12 | 100 | 0.0 x |
| 50 | 0.0 x | ||
| 10 | 0.0 x | ||
| 1 | 0.0 x | ||
| 24 | 100 | 12.2 op | |
| 50 | 7.1 tu | ||
| 10 | 1.9 wx | ||
| 1 | 0.0 x | ||
| 48 | 100 | 27.4 hi | |
| 50 | 18.4 mn | ||
| 10 | 7.8 st | ||
| 1 | 0.0 x | ||
| 72 | 100 | 38.9 d | |
| 50 | 23.3 jk | ||
| 10 | 11.6 op | ||
| 1 | 8.0 rst | ||
| Control | - | - | 0.0 x |
| Fungus Part | Peak Number | Retention Time (Min) | Area | Compound | Molecular Formula | Chemical Functional Group |
|---|---|---|---|---|---|---|
| Pileus | 1 | 8.291 | 911,259 | 2,3-Dihydro- 3,5-Dihydroxy- 6-Methyl -(4H)-pyran-4-one | C6H8O4 | Pyrones |
| 2 | 8.712 | 574,471 | 2-Hexene-3,4,4-Trimethyl | C9H18 | Alkenes | |
| 3 | 15.403 | 5,934,223 | Beta-D-Glucopyranoside, methyl | C7H14O6 | Glucosides | |
| 4 | 16.504 | 83,965,777 | DL-Arabinitol | C5H12O5 | Sugar Alcohol | |
| 5 | 19.792 | 66,792,284 | D-Mannitol | C6H14O6 | Sugar Alcohol | |
| 6 | 28.708 | 6,731,054 | Silane, dimethyl (3-fluorophenoxy) tetradecyloxy | C22H39FO2Si | Silanes | |
| 7 | 29.946 | 10,642,382 | Cholesta-8,14-dien-3-ol, 3 beta, 5-alpha | C27H44O | Steroids | |
| Stipe | 1 | 15.585 | 169,985 | (3aR,4R,7R)-1,4,9,9-Tetramethyl-3,4,5,6,7,8-hexahydro-2H-3a,7-methanoazulen-2-one | C5H22O | Cyclic Ketones |
| 2 | 17.799 | 531,800 | Hexadecanoic acid, methyl ester | C17H34O2 | Palmitates | |
| 3 | 19.428 | 3,143,212 | 9,12-Octadecadienoic acid (Z,Z)-, methyl ester | C19H34O2 | Linoleic Acids | |
| 4 | 19.487 | 5,395,870 | 9-Octadecenoic acid, methyl ester, (E) | C19H36O2 | Oleic Acids | |
| 5 | 19.725 | 173,129 | Methyl stearate | C19H38O2 | Ester | |
| 6 | 23.178 | 454,082 | Bis(2-ethylhexyl) phthalate | C24H38O4 | Phthalic Acid |
| Treatments | Juveniles/kg Soil | Females/Root System | Number of Galls/Root System | Number of Egg Masses/Root System | Number of Eggs/Egg Mass |
|---|---|---|---|---|---|
| 100% | 5293 e | 123 e | 68 e | 97 de | 129 f |
| 50% | 6342 d | 153 d | 83 d | 103 d | 165 e |
| 10% | 11,942 c | 259 bc | 126 bc | 131 bc | 330 bcd |
| 1% | 12,349 b | 278 b | 132 ab | 137 ab | 342 bc |
| UIC | 12,844 a | 327 a | 139 a | 146 a | 355 a |
| UUC | - | - | - | - | - |
| Treatment | Plant Length (cm) | Plant Fresh Weight (g) | Plant Dry Weight (g) | Physiological Parameters | Yield/Plant (g) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | Shoot | Root | Total Chlorophyll (mg/g Fresh Leaf) | Carotenoid (mg/g Fresh Leaf) | ||
| 100% | 48.39 b | 18.86 b | 220 b | 92.0 b | 34.20 b | 10.45 b | 1.93 b | 0.81 b | 605 b |
| 50% | 46.98 bc | 18.52 b | 192 c | 79.5 c | 28.25 c | 9.10 c | 1.67 c | 0.69 c | 530 c |
| 10% | 45.32 bc | 17.25 bc | 147 d | 61.0 d | 23.50 d | 7.40 d | 1.50 d | 0.60 cd | 432 d |
| 1% | 43.56 cd | 16.99 cd | 132 e | 48.0 e | 22.10 de | 6.40 de | 1.49 de | 0.58 cd | 376 e |
| UIC | 25.45 e | 13.3 e | 110 f | 40.0 f | 22.63 de | 6.45 de | 1.45 d | 0.54 de | 347 ef |
| UUC | 54.26 a | 21.57 a | 267 a | 106.0 a | 46.33 a | 16.55 a | 2.72 a | 0.96 a | 876 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatima, S.; Khan, F.; Asif, M.; Alotaibi, S.S.; Islam, K.; Shariq, M.; Khan, A.; Ikram, M.; Ahmad, F.; Khan, T.A.; et al. Root-Knot Disease Suppression in Eggplant Based on Three Growth Ages of Ganoderma lucidum. Microorganisms 2022, 10, 1068. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051068
Fatima S, Khan F, Asif M, Alotaibi SS, Islam K, Shariq M, Khan A, Ikram M, Ahmad F, Khan TA, et al. Root-Knot Disease Suppression in Eggplant Based on Three Growth Ages of Ganoderma lucidum. Microorganisms. 2022; 10(5):1068. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051068
Chicago/Turabian StyleFatima, Saba, Faryad Khan, Mohd Asif, Saqer S. Alotaibi, Khushbu Islam, Mohammad Shariq, Arshad Khan, Mohd Ikram, Faheem Ahmad, Tabreiz Ahmad Khan, and et al. 2022. "Root-Knot Disease Suppression in Eggplant Based on Three Growth Ages of Ganoderma lucidum" Microorganisms 10, no. 5: 1068. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10051068