
Characterization and Assessment of 2, 4-Diacetylphloroglucinol (DAPG)-Producing Pseudomonas fluorescens VSMKU3054 for the Management of Tomato Bacterial Wilt Caused by Ralstonia solanacearum

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods and Materials
2.1. Dual Culture Method
2.2. Assessing Carbohydrate Utilization and Hydrolytic Enzyme Production
2.3. Production of Antimicrobial Metabolites by VSMKU3054
2.4. Detection of 2,4-DAPG Antimicrobial Encoding Genes in P. fluorescens by PCR
2.5. Optimizing Growth Conditions for the Production of Antimicrobial Metabolites by P. fluorescens VSMKU3054
2.6. Mass Production and Extraction of DAPG
2.7. Purification and Characterization of Metabolites
2.8. Assay of Minimum Inhibitory Concentrations (MIC) of DAPG and Its Effect on Cell Viability
2.9. Determination of Reactive Oxygen Species (ROS)
2.10. DNA Damage of R. solanacearum
2.11. Efficiency of P. fluorescens on the Suppression of R. solanacearum under Greenhouse Experiment
2.11.1. Preparation of P. fluorescens and R. solanacearum
2.11.2. Preparation of Tomato Seedlings
2.11.3. Soil Application (SA)
2.11.4. Root Dipping (RD)
2.11.5. Seed Treatments (ST)
2.11.6. Foliar Spray (FS)
2.12. Statistical Analysis
3. Results
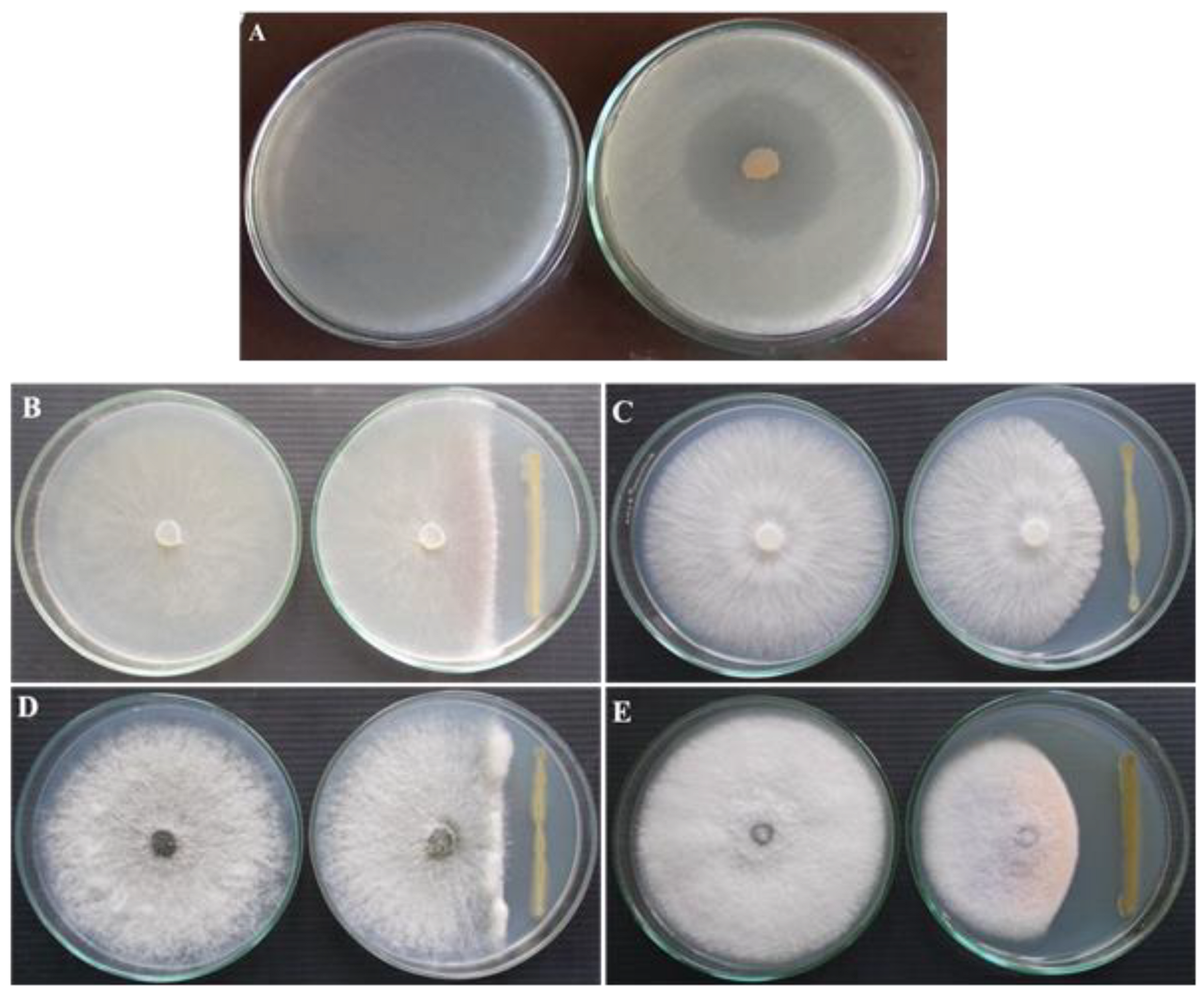
3.1. Antagonistic Activity of P. fluorescens VSMKU3054 against Plant Pathogens
3.2. Phenotypic and Biochemical Characterization
3.3. Secondary Metabolites Produced from P. fluorescens VSMKU3054
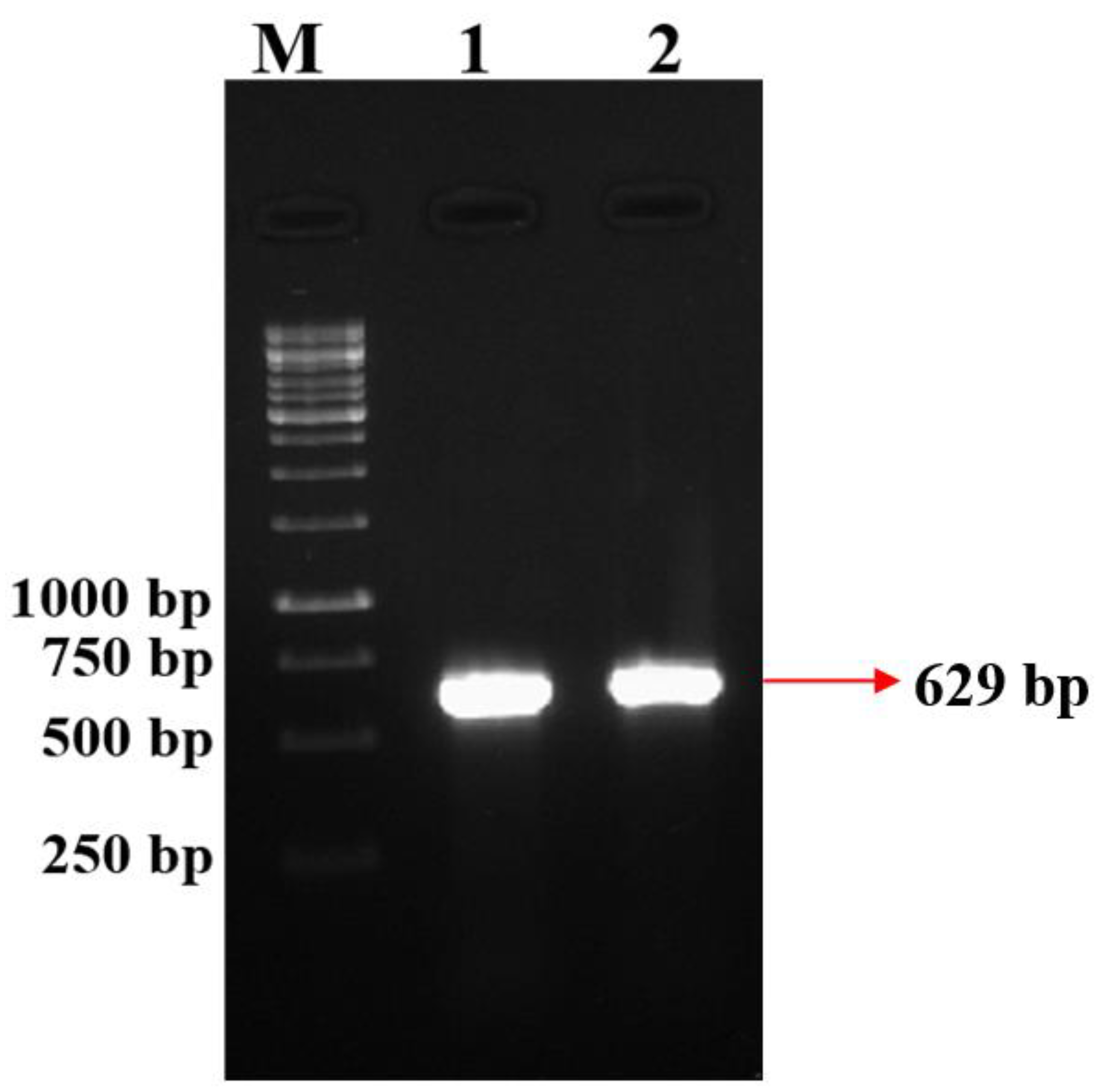
3.4. Detection of DAPG Antibiotic Gene Amplification by PCR
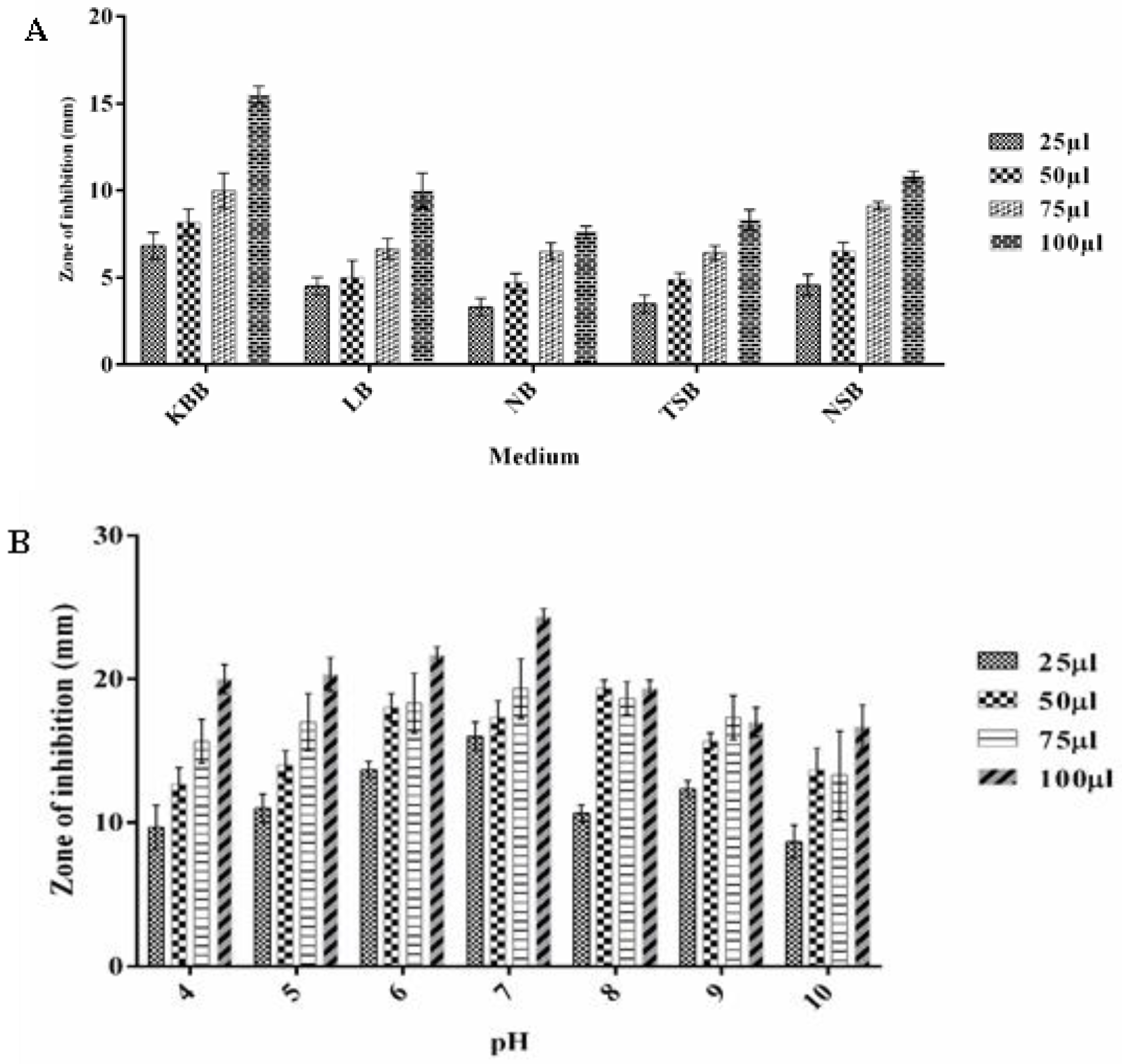
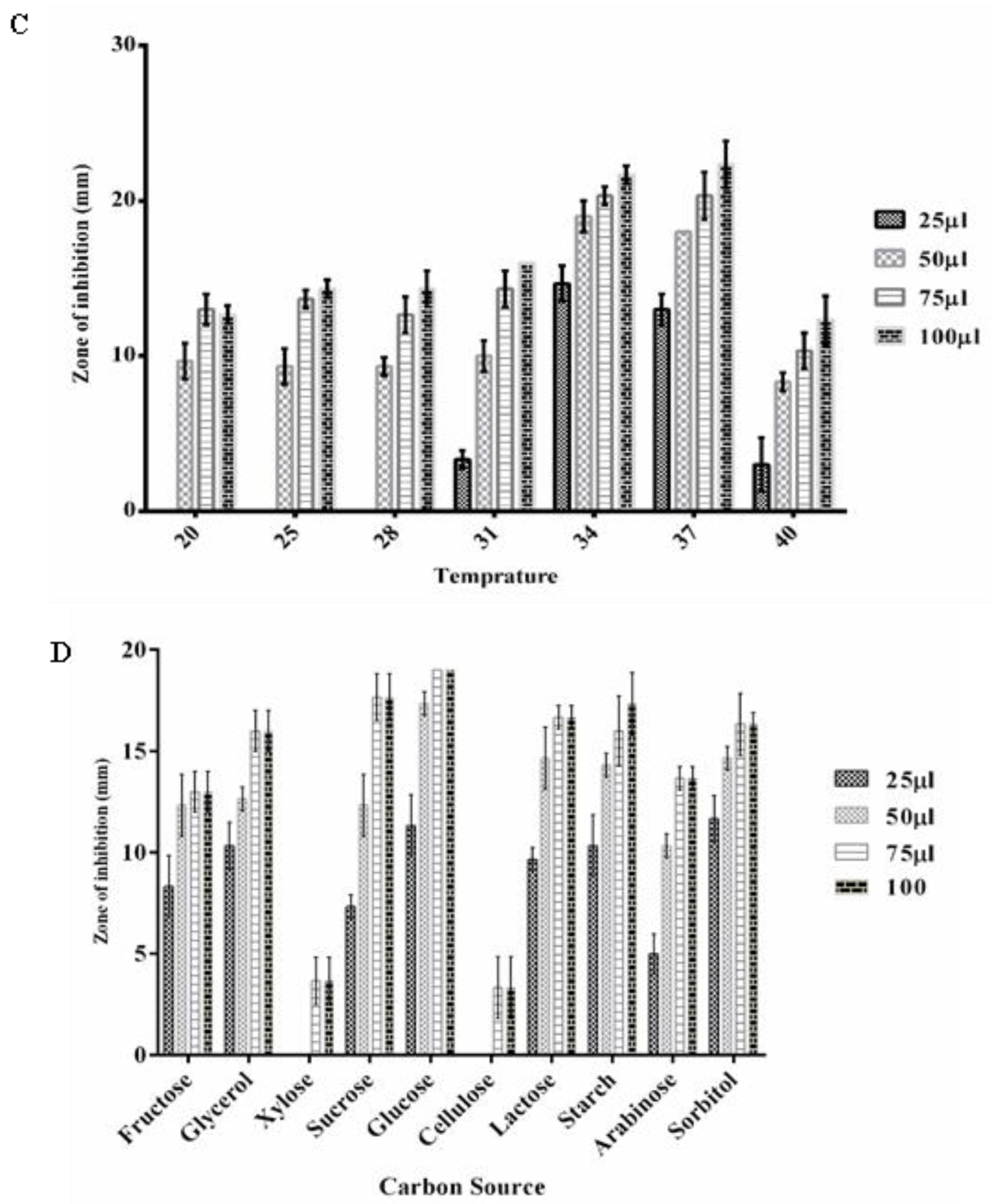
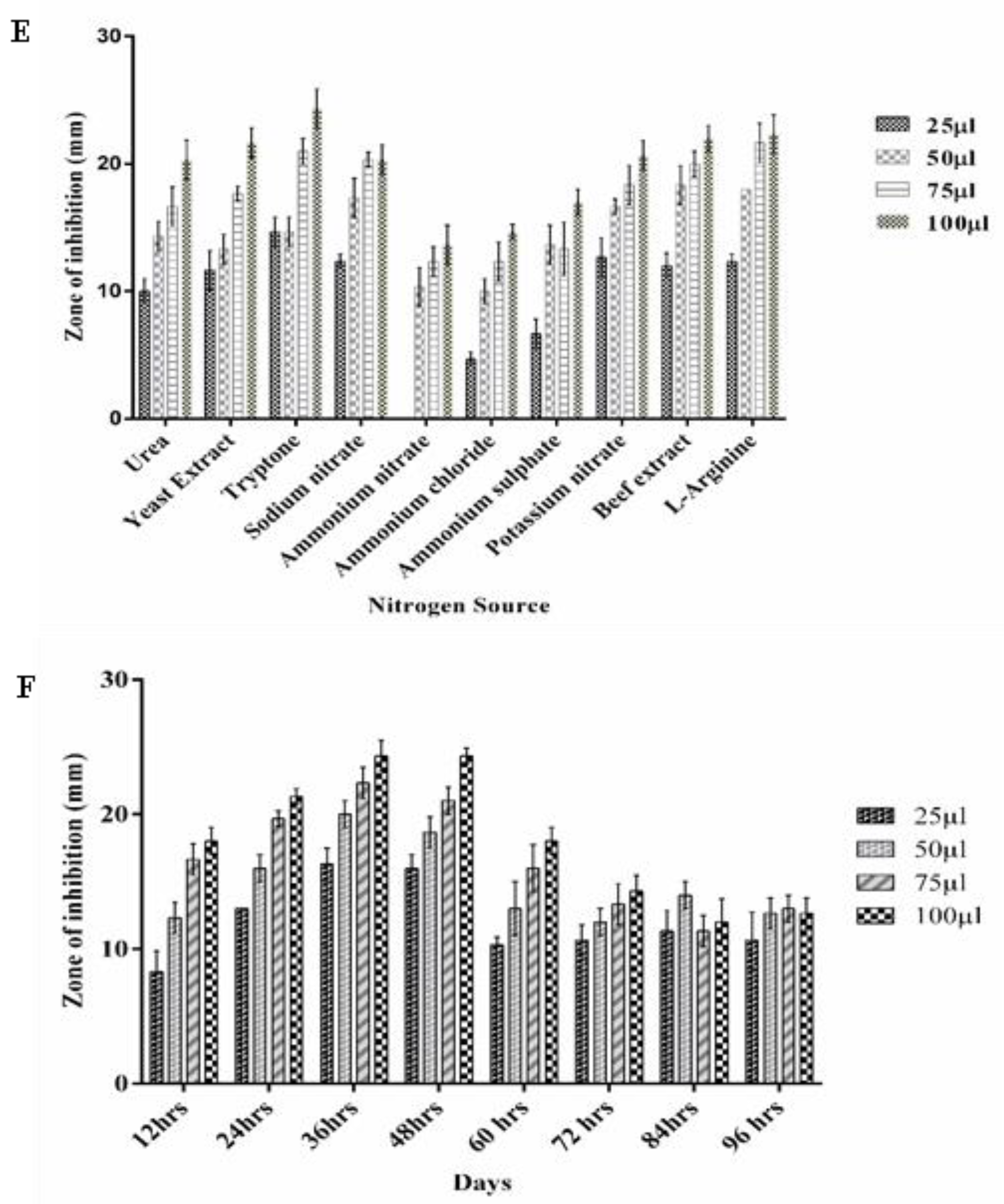
3.5. Optimizing the Growth Conditions for Antimicrobial Metabolite Production by P. fluorescens



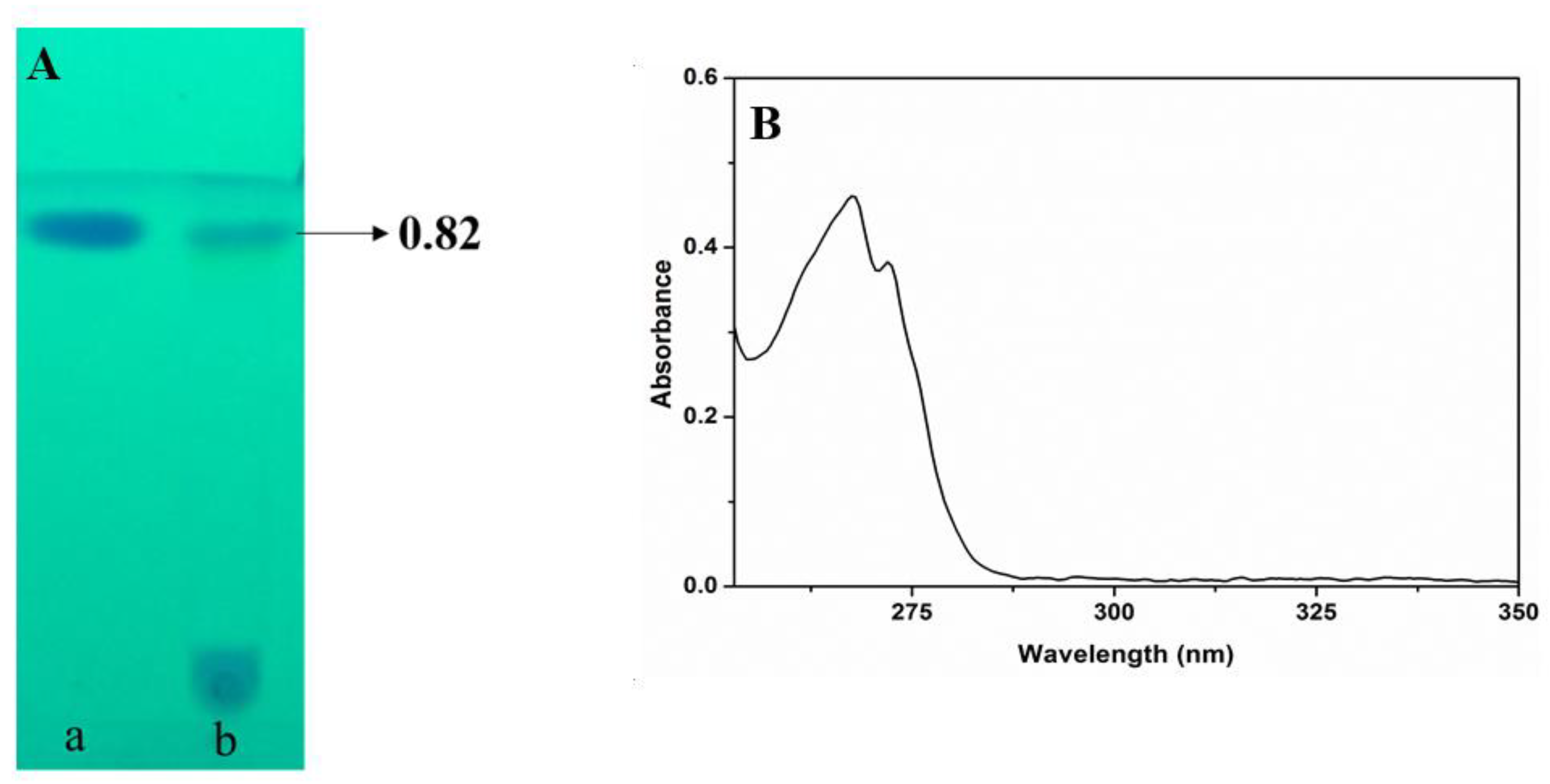
3.6. Extraction and Purification of Secondary Metabolites
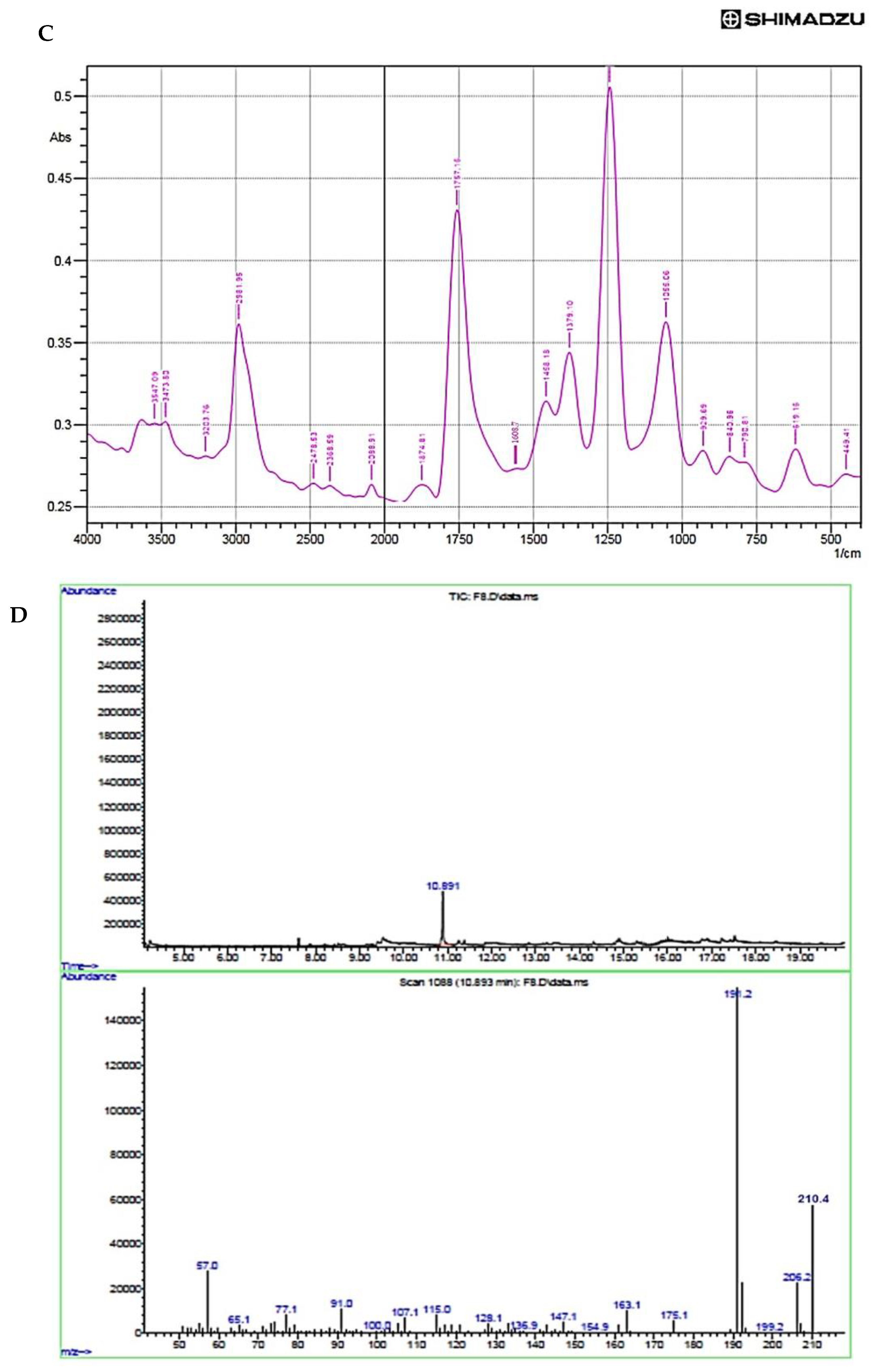
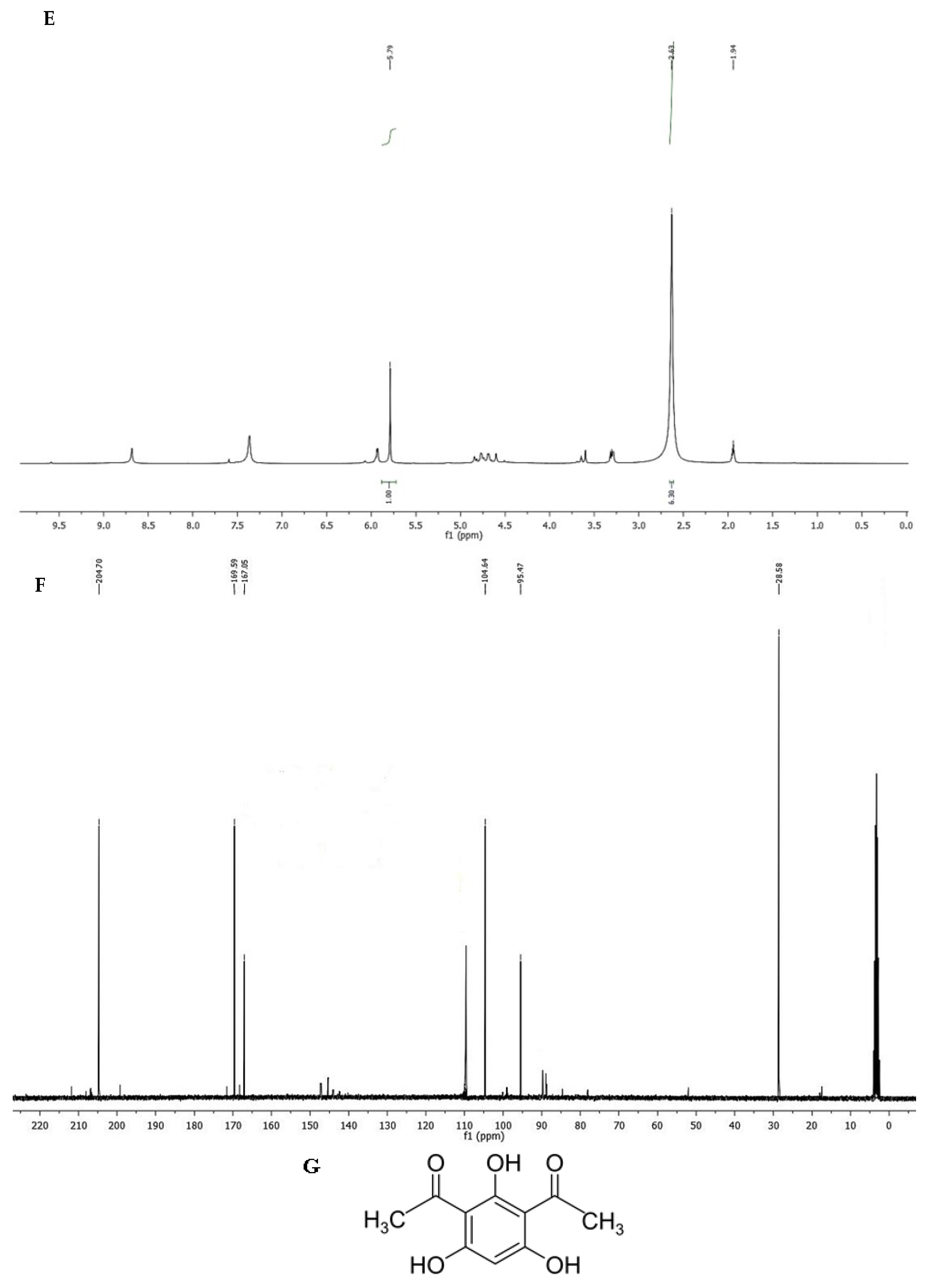
3.7. Characterization of Purified Compound



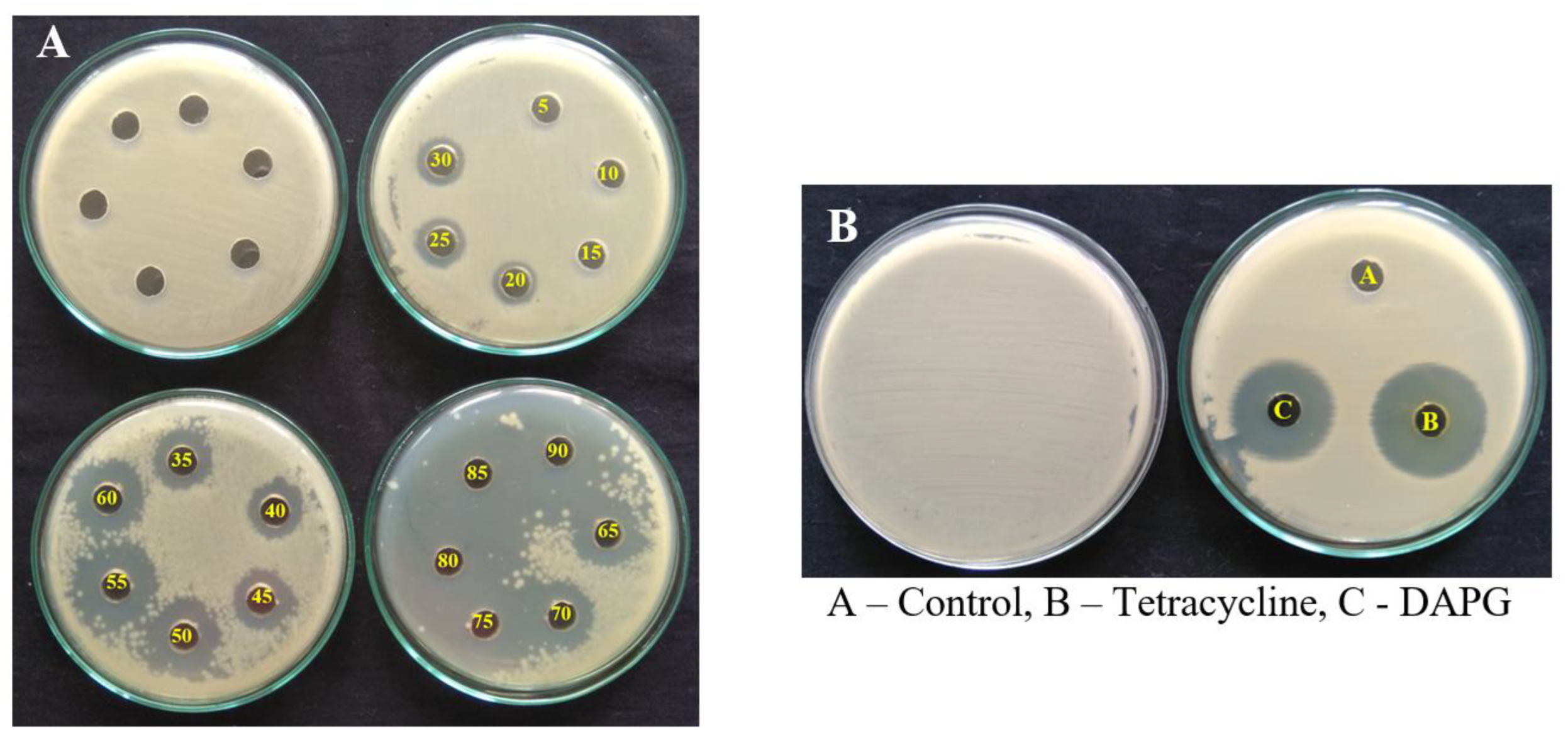
3.8. Bioactive Assay of DAPG against R. solanacearum
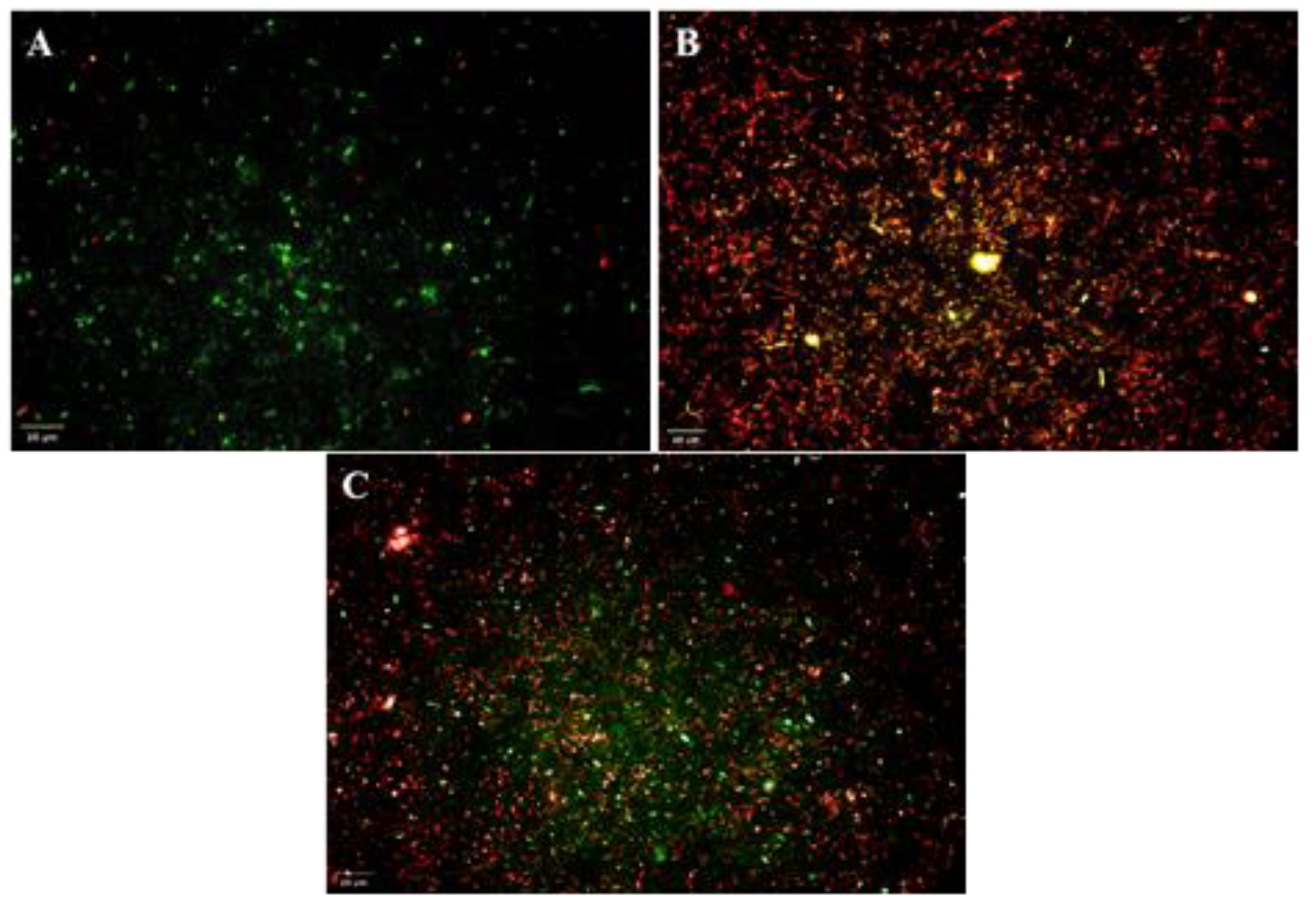
3.9. Live/Dead Cell Assay of R. solanacearum
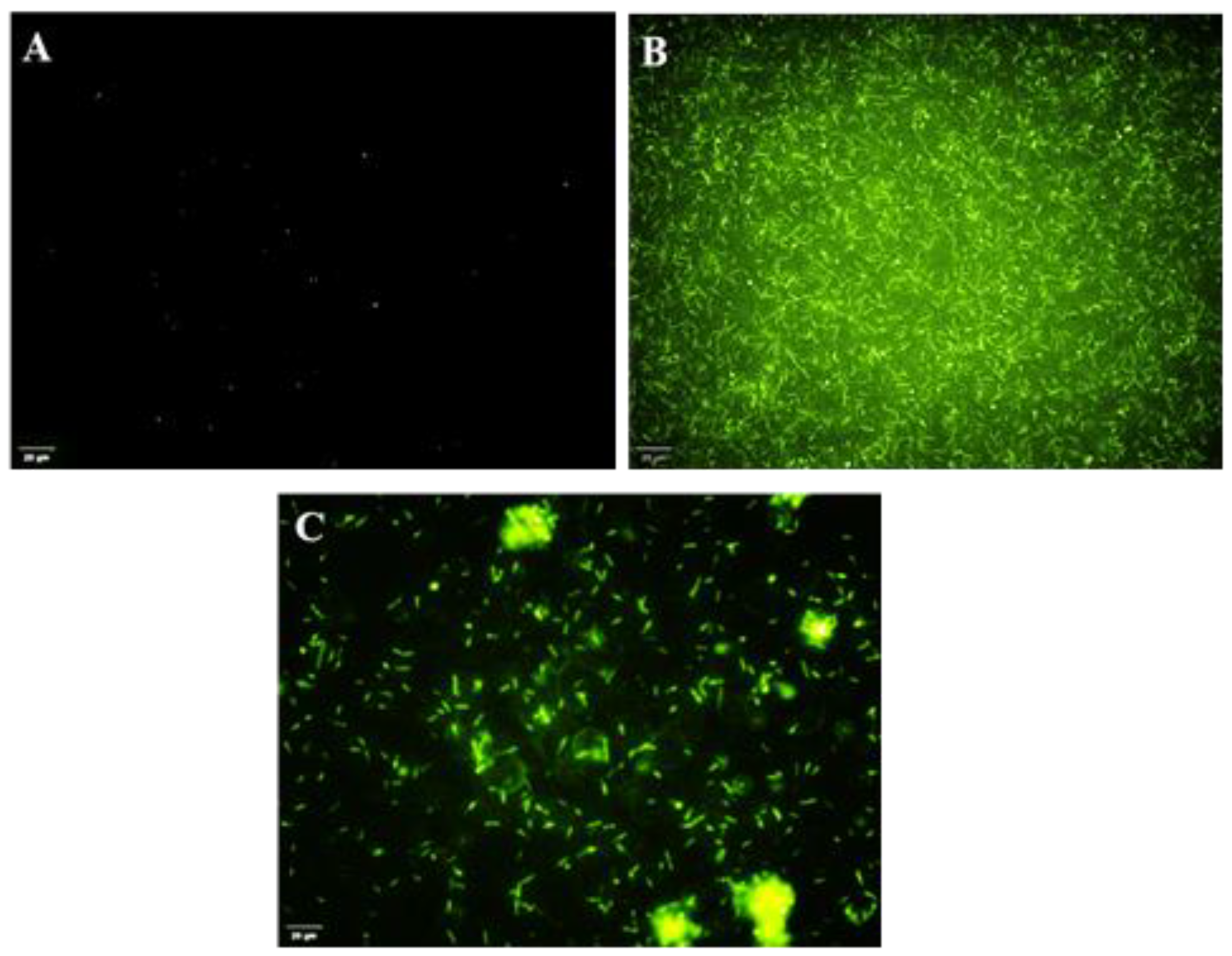
3.10. Reactive Oxygen Species Assay—DAPG
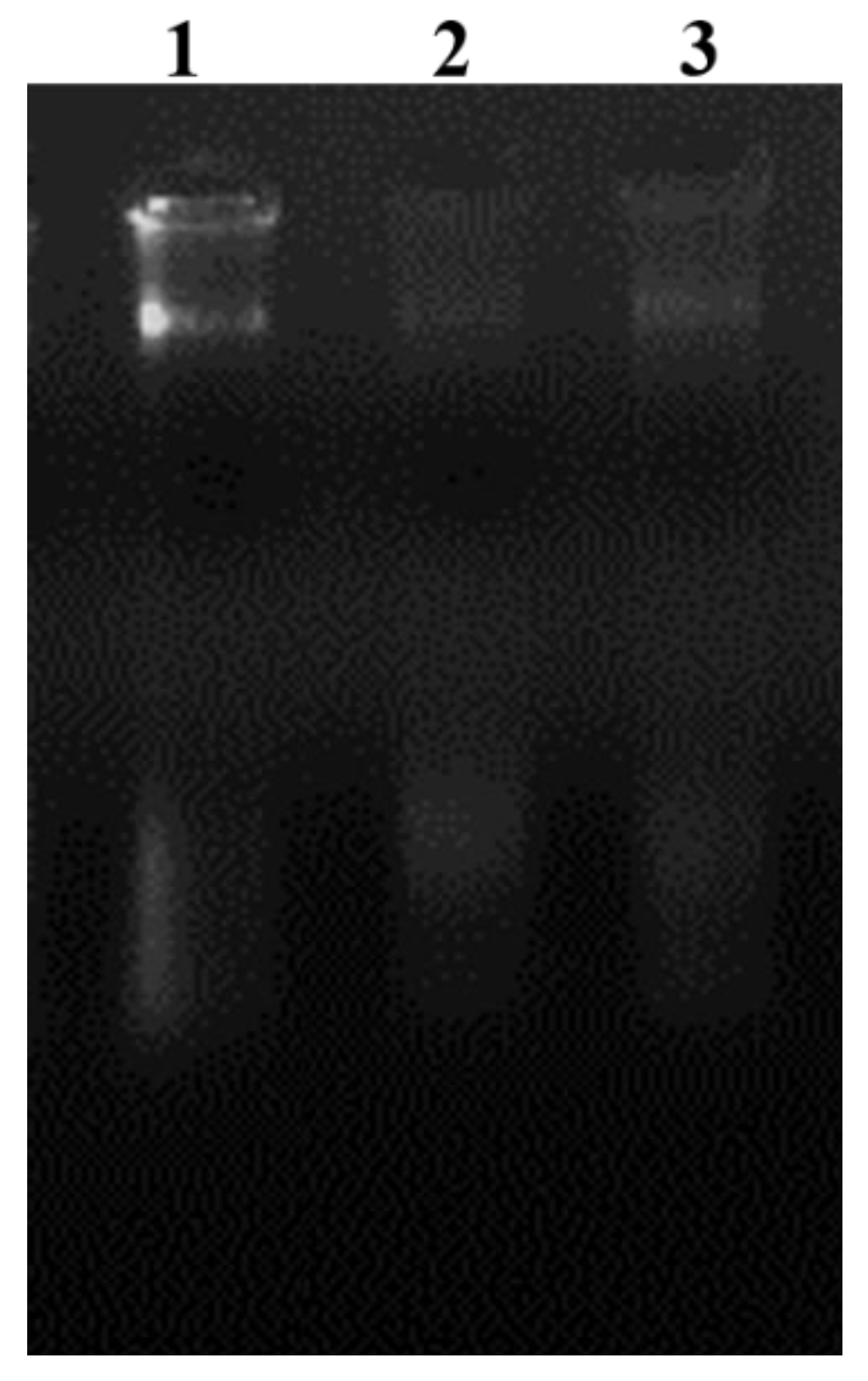
3.11. DNA Damage Assay
3.12. Biocontrol Efficiency of P. fluorescens VSMKU3054 and 2,4-DAPG against Tomato Bacterial Wilt Disease under Greenhouse Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacobs, J.M.; Babujee, L.; Meng, F.; Milling, A.; Allen, C. The in planta transcriptome of Ralstonia solanacearum: Conserved physiological and virulence strategies during bacterial wilt of tomato. mBio 2012, 3, e00114-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popoola, A.; Ganiyu, S.; Enikuomehin, O.; Bodunde, J.; Adedibu, O.; Durosomo, H.; Karunwi, O. Isolation and Characterization of Ralstonia solanacearum Causing Bacterial Wilt of Tomato in Nigeria. Niger. J. Biotechnol. 2015, 29, 1. [Google Scholar] [CrossRef]
- Mohammed, A.F.; Oloyede, A.R.; Odeseye, A.O. Biological control of bacterial wilt of tomato caused by Ralstonia solanacearum using Pseudomonas species isolated from the rhizosphere of tomato plants. Arch. Phytopathol. Plant Prot. 2020, 53, 1–16. [Google Scholar] [CrossRef]
- Denny, T.P. Plant pathogenic Ralstonia species. In Plant-Associated Bacteria; Gnanamanickam, S.S., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 573–644. [Google Scholar] [CrossRef]
- Cho, H.; Song, E.S.; Lee, Y.K.; Lee, S.; Lee, S.W.; Jo, A.; Lee, B.M.; Kim, J.G.; Hwang, I. Analysis of genetic and pathogenic diversity of Ralstonia solanacearum causing potato bacterial wilt in Korea. Plant Pathol. J. 2018, 34, 23–34. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Ranamukhaarachchi, S.L. Soil-borne antagonists for biological control of bacterial wilt disease caused by Ralstonia solanacearum in tomato and pepper. J. Plant Pathol. 2010, 92, 395–406. [Google Scholar] [CrossRef]
- Aino, M. Studies on biological control of bacterial wilt caused by Ralstonia solanacearum using endophytic bacteria. J. Gen. Plant Pathol. 2016, 82, 323–325. [Google Scholar] [CrossRef]
- Alit-Susanta, W.G.N.; Takikawa, Y. Phenotypic characterization of Pseudomonas fluorescens PfG32R and its spontaneous gacS mutants and biocontrol activity against bacterial wilt disease of tomato. J. Gen. Plant Pathol. 2006, 72, 168–175. [Google Scholar] [CrossRef]
- Garrido-Sanz, D.; Arrebola, E.; Martínez-Granero, F.; García-Méndez, S.; Muriel, C.; Blanco-Romero, E.; Martín, M.; Rivilla, R.; Redondo-Nieto, M. Classification of isolates from the Pseudomonas fluorescens complex into phylogenomic groups based in group-specific markers. Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ramamoorthy, V.; Viswanathan, R.; Raguchander, T.; Prakasam, V.; Samiyappan, R. Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases. Crop Prot. 2001, 20, 1–11. [Google Scholar] [CrossRef]
- Ramamoorthy, V.; Raguchander, T.; Samiyappan, R. Induction of defense-related proteins in tomato roots treated with Pseudomonas fluorescens Pf1 and Fusarium oxysporum f. sp. lycopersici. Plant Soil 2002, 239, 55–68. [Google Scholar] [CrossRef]
- Suresh, P.; Shanmugaiah, V.; Rajakrishnan, R.; Muthusamy, K.; Ramamoorthy, V. Pseudomonas fluorescens VSMKU3054 mediated induced systemic resistance in tomato against Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2022, 119, 101836. [Google Scholar] [CrossRef]
- Marimuthu, S.; Subbian, P.; Ramamoorthy, V.; Samiyappan, R. Synergistic effect of combined application of Azospirillum and Pseudomonas fluorescens with inorganic fertilizers on root rot incidence and yield of cotton. J. Plant Dis. Prot. 2002, 109, 569–577. [Google Scholar]
- Ramamoorthy, V.; Govindaraj, L.; Dhanasekaran, M.; Vetrivel, S.; Kumar, K.K.; Ebenezar, E. Combination of driselase and lysing enzyme in one molar potassium chloride is effective for the production of protoplasts from germinated conidia of Fusarium verticillioides. J. Microbiol. Methods 2015, 111, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, E.; Tienda, S.; Vida, C.; De Vicente, A.; Cazorla, F.M. Fitness features involved in the biocontrol interaction of Pseudomonas chlororaphis with host plants: The case study of PcPCL1606. Front. Microbiol. 2019, 10, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, D.; Keel, C.; Laville, J.; Maurhofer, M.; Oberhänsli, T.; Schnider, U.; Voisard, C.; Wüthrich, B.; Defago, G. Secondary metabolites of Pseudomonas fluorescens strain CHA0 involved in the suppression of root diseases. In Advances in Molecular Genetics of Plant-Microbe Interactions; Springer: Dordrecht, The Netherlands, 1991; Volume 1, pp. 450–456. [Google Scholar] [CrossRef]
- Scales, B.S.; Dickson, R.P.; Lipuma, J.J.; Huffnagle, G.B. Microbiology, genomics, and clinical significance of the Pseudomonas fluorescens species complex, an unappreciated colonizer of humans. Clin. Microbiol. Rev. 2014, 27, 927–948. [Google Scholar] [CrossRef] [Green Version]
- Moin, S.; Ali, S.A.; Hasan, K.A.; Tariq, A.; Sultana, V.; Ara, J.; Ehteshamul-Haque, S. Managing the root rot disease of sunflower with endophytic fluorescent Pseudomonas associated with healthy plants. Crop Prot. 2020, 130, 105066. [Google Scholar] [CrossRef]
- Keel, C.; Schnider, U.; Maurhofer, M.; Voisard, C.; Laville, J.; Burger, U.; Wirthner, P.J.; Haas, D.; Défago, G. Suppression of Root Diseases by Pseudomonas fluorescens CHA0: Importance of the Bacterial Secondary Metabolite 2,4-Diacetylphloroglucinol. Mol. Plant-Microbe Interact. 1992, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Almario, J.; Moënne-loccoz, Y.; Muller, D. Soil Biology & Biochemistry Monitoring of the relation between 2, 4-diacetylphloroglucinol-producing Pseudomonas and Thielaviopsis basicola populations by real-time PCR in tobacco black root-rot suppressive and conducive soils. Soil Biol. Biochem. 2013, 57, 144–155. [Google Scholar] [CrossRef]
- Zhou, T.T.; Li, C.Y.; Chen, D.; Wu, K.; Shen, Q.R.; Shen, B. PhlF- mutant of Pseudomonas fluorescens J2 improved 2,4-DAPG biosynthesis and biocontrol efficacy against tomato bacterial wilt. Biol. Control 2014, 78, 1–8. [Google Scholar] [CrossRef]
- Tada, M.; Takakuwa, T.; Nagai, M.; Yoshii, T. Antiviral and antimicrobial activity of 2, 4-Diacylphloroglucinols, 2-Acylcyclohexane-1, 3-diones and 2-Carboxamidocyclo-hexane-1, 3-diones. Agr. Biol. Chem. 1990, 54, 3061–3063. [Google Scholar] [CrossRef]
- Zhou, T.; Chen, D.; Li, C.; Sun, Q.; Li, L.; Liu, F.; Shen, Q.; Shen, B. Isolation and characterization of Pseudomonas brassicacearum J12 as an antagonist against Ralstonia solanacearum and identification of its antimicrobial components. Microbiol. Res. 2012, 167, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Veena, V.K.; Kennedy, K.; Lakshmi, P.; Krishna, R.; Sakthivel, N. Anti-leukemic, anti-lung, and anti-breast cancer potential of the microbial polyketide 2, 4-diacetylphloroglucinol (DAPG) and its interaction with the metastatic proteins than the antiapoptotic Bcl-2 proteins. Mol. Cell. Biochem. 2016, 414, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, R.; Joshi, A.A.; Ghanekar, M.P. Pseudomonads: Major antagonistic endophytic bacteria to suppress bacterial wilt pathogen, Ralstonia solanacearum in the eggplant (Solanum melongena L.). World J. Microbiol. Biotechnol. 2009, 25, 47–55. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Subramanian, D.; Yoon, E.; Kwon, T.; Chun, S.C. Meta-analysis reveals that the genus Pseudomonas can be a better choice of biological control agent against bacterial wilt disease caused by Ralstonia solanacearum. Plant Pathol. J. 2016, 32, 216–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsayed, T.R.; Jacquiod, S.; Nour, E.H.; Sørensen, S.J.; Smalla, K. Biocontrol of Bacterial Wilt Disease Through Complex Interaction Between Tomato Plant, Antagonists, the Indigenous Rhizosphere Microbiota, and Ralstonia solanacearum. Front. Microbiol. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Kwak, Y.S.; Han, S.; Thomashow, L.S.; Rice, J.T.; Paulitz, T.C.; Kim, D.; Weller, D.M. Saccharomyces cerevisiae genome-wide mutant screen for sensitivity to 2, 4-diacetylphloroglucinol, an antibiotic produced by Pseudomonas fluorescens. Appl. Environ. Microbiol. 2011, 77, 1770–1776. [Google Scholar] [CrossRef] [Green Version]
- Suresh, P.; Varathraju, G.; Shanmugaiah, V.; Almaary, K.S.; Elbadawi, Y.B.; Mubarak, A. Partial purification and characterization of 2, 4-diacetylphloroglucinol producing Pseudomonas fluorescens VSMKU3054 against bacterial wilt disease of tomato. Saudi J. Biol. Sci. 2021, 28, 2155–2167. [Google Scholar] [CrossRef]
- Suresh, P.; Shanmugaiah, V.; Almaary, K.S.; Dawoud, T.M.; Elbadawi, Y.B. Fluorescent pseudomonads (FPs) as a potential biocontrol and plant growth promoting agent associated with tomato rhizosphere. J. King Saud Univ.-Sci. 2021, 33, 101423. [Google Scholar] [CrossRef]
- Naik, P.R.; Raman, G.; Narayanan, K.B.; Sakthivel, N. Assessment of genetic and functional diversity of phosphate solubilizing fluorescent pseudomonads isolated from rhizospheric soil. BMC Microbiol. 2008, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shanmugaiah, V.; Mathivanan, N.; Balasubramanian, N.; Manoharan, P.T. Optimization of cultural conditions for production of chitinase by Bacillus laterosporous MML2270 isolated from rice rhizosphere soil. Afr. J. Biotechnol. 2008, 7, 2562–2568. [Google Scholar] [CrossRef]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Bakker, A.W.; Schippers, B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas SPP-mediated plant growth-stimulation. Soil Biol. Biochem. 1987, 19, 451–457. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya. 1948, 17, 362–370. [Google Scholar]
- Harikrishnan, H.; Shanmugaiah, V.; Balasubramanian, N.; Sharma, M.P.; Kotchoni, S.O. Antagonistic potential of native strain Streptomyces aurantiogriseus VSMGT1014 against sheath blight of rice disease. World J. Microbiol. Biotechnol. 2014, 30, 3149–3161. [Google Scholar] [CrossRef] [PubMed]
- Immanuel, J.E.; Hemamalini, R.; Gnanamanickam, S.S. Genetic Diversity of Biocontrol Strains of Pseudomonas fluorescens Producing 2, 4-diacetylphloroglucinol from Southern India. J. Crop Improv. 2012, 26, 228–243. [Google Scholar] [CrossRef]
- Biniarz, P.; Coutte, F.; Gancel, F.; Łukaszewicz, M. High-throughput optimization of medium components and culture conditions for the efficient production of a lipopeptide pseudofactin by Pseudomonas fluorescens BD5. Microb. Cell Fact. 2018, 17, 121. [Google Scholar] [CrossRef]
- Paret, M.L.; Cabos, R.; Kratky, B.A.; Alvarez, A.M. Effect of plant essential oils on Ralstonia solanacearum race 4 and bacterial wilt of edible ginger. Plant Dis. 2010, 94, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wu, L.; Yao, X.; Zhao, S.; Wang, J.; Li, S.; Ding, W. Hydroxycoumarins: New, effective plant-derived compounds reduce Ralstonia pseudosolanacearum populations and control tobacco bacterial wilt. Microbiol. Res. 2018, 215, 15–21. [Google Scholar] [CrossRef]
- Ramalingam, V.; Revathidevi, S.; Shanmuganayagam, T.; Muthulakshmi, L.; Rajaram, R. Biogenic gold nanoparticles induce cell cycle arrest through oxidative stress and sensitize mitochondrial membranes in A549 lung cancer cells. RSC Adv. 2016, 6, 20598–20608. [Google Scholar] [CrossRef]
- Cai, L.; Chen, J.; Liu, Z.; Wang, H.; Yang, H.; Ding, W. Magnesium oxide nanoparticles: Effective agricultural antibacterial agent against Ralstonia solanacearum. Front. Microbiol. 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Xue, Q.Y.; Chen, Y.; Li, S.M.; Chen, L.F.; Ding, G.C.; Guo, D.W.; Guo, J.H. Evaluation of the strains of Acinetobacter and Enterobacter as potential biocontrol agents against Ralstonia wilt of tomato. Biol. Control 2009, 48, 252–258. [Google Scholar] [CrossRef]
- Amaresan, N.; Jayakumar, V.; Kumar, K.; Thajuddin, N. Biocontrol and plant growth-promoting ability of plant-associated bacteria from tomato (Lycopersicum esculentum) under field condition. Microb. Pathog. 2019, 136, 103713. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Dong, Y.; Liao, H.; Huang, J.; Song, S.; Xu, Y.; Shen, Q. Antagonistic bacterium Bacillus amyloliquefaciens induces resistance and controls the bacterial wilt of tomato. Pest Manag. Sci. 2013, 69, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Abo-Elyousr, K.A.M.; Khalil Bagy, H.M.M.; Hashem, M.; Alamri, S.A.M.; Mostafa, Y.S. Biological control of the tomato wilt caused by Clavibacter michiganensis subsp. michiganensis using formulated plant growth-promoting bacteria. Egypt. J. Biol. Pest Control 2019, 29. [Google Scholar] [CrossRef]
- Jayalakshmi, R.; Oviya, R.; Premalatha, K.; Mehetre, S.T.; Paramasivam, M.; Kannan, R.; Theradimani, M.; Pallavi, M.S.; Mukherjee, P.K.; Ramamoorthy, V. Production, stability and degradation of Trichoderma gliotoxin in growth medium, irrigation water and agricultural soil. Sci. Rep. 2021, 11, 16536. [Google Scholar] [CrossRef]
- Vanitha, S.C.; Niranjana, S.R.; Mortensen, C.N.; Umesha, S. Bacterial wilt of tomato in Karnataka and its management by Pseudomonas fluorescens. BioControl 2009, 54, 685–695. [Google Scholar] [CrossRef]
- Xue, Q.Y.; Ding, G.C.; Li, S.M.; Yang, Y.; Lan, C.Z.; Guo, J.H.; Smalla, K. Rhizocompetence and antagonistic activity towards genetically diverse Ralstonia solanacearum strains—An improved strategy for selecting biocontrol agents. Appl. Microbiol. Biotechnol. 2013, 97, 1361–1371. [Google Scholar] [CrossRef]
- Vidhyasekaran, P.; Kamala, N.; Ramanathan, A.; Rajappan, K.; Paranidharan, V.; Velazhahan, R. Induction of systemic resistance by Pseudomonas fluorescens Pf1 against Xanthomonas oryzae pv. Oryzae in rice leaves. Phytoparasitica 2001, 29, 155. [Google Scholar] [CrossRef]
- Cantore, P.L.O.; Shanmugaiah, V.; Nicola, S.I. Antibacterial activity of essential oil components and their potential use in seed disinfection. J. Agric. Food Chem. 2009, 57, 9454–9461. [Google Scholar] [CrossRef]
- Cabanás, C.G.L.; Legarda, G.; Ruano-Rosa, D.; Pizarro-Tobías, P.; Valverde-Corredor, A.; Niqui, J.L.; Triviño, J.C.; Roca, A.; Mercado-Blanco, J. Indigenous Pseudomonas spp. Strains from the Olive (Olea europaea L.) rhizosphere as effective biocontrol agents against Verticillium dahliae: From the host roots to the bacterial genomes. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Chenniappan, C.; Narayanasamy, M.; Daniel, G.M.; Ramaraj, G.B.; Ponnusamy, P.; Sekar, J.; Vaiyapuri Ramalingam, P. Biocontrol efficiency of native plant growth promoting rhizobacteria against rhizome rot disease of turmeric. Biol. Control 2019, 129, 55–64. [Google Scholar] [CrossRef]
- Costa, R.; Gomes, N.C.; Peixoto, R.S.; Rumjanek, N.; Berg, G.; Mendonça-Hagler, L.C.; Smalla, K. Diversity and antagonistic potential of Pseudomonas spp. associated to the rhizosphere of maize grown in a subtropical organic farm. Soil Biol. Biochem. 2006, 38, 2434–2447. [Google Scholar] [CrossRef]
- Alsohim, A.S.; Taylor, T.B.; Barrett, G.A.; Gallie, J.; Zhang, X.X.; Altamirano-Junqueira, A.E.; Johnson, L.J.; Rainey, P.B.; Jackson, R.W. The biosurfactant viscosin produced by Pseudomonas fluorescens SBW 25 aids spreading motility and plant growth promotion. Environ. Microbiol. 2014, 16, 2267–2281. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.; Srinivasamurthy, R.; Dash, P.K.; Gupta, P. Isolation, characterization and evaluation of the biocontrol potential of Pseudomonas protegens RS-9 against Ralstonia solanacearum in Tomato. Indian J. Exp. Biol. 2017, 55, 595–603. [Google Scholar]
- Vinay, J.U.; Naik, M.K.; Rangeshwaran, R.; Chennappa, G.; Shaikh, S.S.; Sayyed, R.Z. Detection of antimicrobial traits in fluorescent pseudomonads and molecular characterization of an antibiotic pyoluteorin. 3 Biotech 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Durairaj, K.; Velmurugan, P.; Park, J.H.; Chang, W.S.; Park, Y.J.; Senthilkumar, P.; Choi, K.M.; Lee, J.H.; Oh, B.T. Potential for plant biocontrol activity of isolated Pseudomonas aeruginosa and Bacillus stratosphericus strains against bacterial pathogens acting through both induced plant resistance and direct antagonism. FEMS Microbiol. Lett. 2017, 364, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shanahan, P.; O’Sullivan, D.J.; Simpson, P.; Glennon, J.D.; O’Gara, F. Isolation of 2,4-diacetylphloroglucinol from a fluorescent pseudomonad and investigation of physiological parameters influencing its production. Appl. Environ. Microbiol. 1992, 58, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Harikrishnan, H.; Shanmugaiah, V.; Nithya, K.; Balasubramanian, N.; Sharma, M.P.; Gachomo, E.W.; Kotchoni, S.O. Enhanced production of phenazine-like metabolite produced by Streptomyces aurantiogriseus VSMGT1014 against rice pathogen, Rhizoctonia solani. J. Basic Microbiol. 2016, 56, 153–161. [Google Scholar] [CrossRef]
- Schmidt, N.G.; Żądło-Dobrowolska, A.; Ruppert, V.; Höflehner, C.; Wiltschi, B.; Kroutil, W. Molecular cloning, expression, and characterization of acyltransferase from Pseudomonas protegens. Appl. Microbiol. Biotechnol. 2018, 102, 6057–6068. [Google Scholar] [CrossRef] [Green Version]
- Saber, F.M.A.; Abdelhafez, A.A.; Hassan, E.A.; Ramadan, E.M. Characterization of fluorescent pseudomonads isolates and their efficiency on the growth promotion of tomato plant. Ann. Agric. Sci. 2015, 60, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Sekar, J.; Prabavathy, V.R. Novel Phl-producing genotypes of finger millet rhizosphere associated pseudomonads and assessment of their functional and genetic diversity. FEMS Microbiol. Ecol. 2014, 89, 32–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayyadurai, N.; Ravindra Naik, P.; Sreehari Rao, M.; Sunish Kumar, R.; Samrat, S.K.; Manohar, M.; Sakthivel, N. Isolation and characterization of a novel banana rhizosphere bacterium as fungal antagonist and microbial adjuvant in micropropagation of banana. J. Appl. Microbiol. 2006, 100, 926–937. [Google Scholar] [CrossRef] [PubMed]
- Brucker, R.M.; Baylor, C.M.; Walters, R.L.; Lauer, A.; Harris, R.N.; Minbiole, K.P.C. The identification of 2,4-diacetylphloroglucinol as an antifungal metabolite produced by cutaneous bacteria of the salamander plethodon cinereus. J. Chem. Ecol. 2008, 34, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Marchand, P.A.; Weller, D.M.; Bonsall, R.F. Convenient Synthesis of 2, 4-Diacetylphloroglucinol, a Natural Antibiotic Involved in the Control of Take-All Disease of Wheat. J. Agric. Food Chem. 2000, 48, 1882–1887. [Google Scholar] [CrossRef]
- Gutiérrez-García, K.; Neira-González, A.; Pérez-Gutiérrez, R.M.; Granados-Ramírez, G.; Zarraga, R.; Wrobel, K.; Barona-Gómez, F.; Flores-Cotera, L.B. Phylogenomics of 2,4-Diacetylphloroglucinol-Producing Pseudomonas and Novel Antiglycation Endophytes from Piper auritum. J. Nat. Prod. 2017, 80, 1955–1963. [Google Scholar] [CrossRef]
- Kang, B.R. Biocontrol of tomato Fusarium wilt by a novel genotype of 2,4-diacetylphloroglucinol- producing Pseudomonas sp. NJ134. Plant Pathol. J. 2012, 28, 93–100. [Google Scholar] [CrossRef]
- Nagendran, S.; Vaikuntavasan, P.; Muthusamy, K.; Dananjeyan, B.; Venkatasamy, B. Chromatographic Detection of Antibacterial Antibiotics from Pseudomonas fluorescens (TNAU PF1) and Their Effect on Bacterial Wilt Pathogen Ralstonia solanacearum (KK2). Int. Res. J. Pure Appl. Chem. 2019, 20, 1–10. [Google Scholar] [CrossRef]
- Applerot, G.; Lellouche, J.; Lipovsky, A.; Nitzan, Y.; Lubart, R.; Gedanken, A.; Banin, E. Understanding the antibacterial mechanism of CuO nanoparticles: Revealing the route of induced oxidative stress. Small 2012, 8, 3326–3337. [Google Scholar] [CrossRef]
- Le Ouay, B.; Stellacci, F. Antibacterial activity of silver nanoparticles: A surface science insight. Nano Today 2015, 10, 339–354. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Bulut, O.; Some, S.; Mandal, A.K.; Yilmaz, M.D. Green synthesis of silver nanoparticles: Biomolecule-nanoparticle organizations targeting antimicrobial activity. RSC Adv. 2019, 9, 2673–2702. [Google Scholar] [CrossRef] [Green Version]
- Qessaoui, R.; Bouharroud, R.; Furze, J.N.; El Aalaoui, M.; Akroud, H.; Amarraque, A.; Vaerenbergh, J.V.; Tahzima, R.; Mayad, E.H.; Chebli, B. Applications of New Rhizobacteria Pseudomonas Isolates in Agroecology via Fundamental Processes Complementing Plant Growth. Sci. Rep. 2019, 9, 12832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.; Wei, W.; Li, G.; Sun, M.; Li, H.; Wu, J.; Hu, F. Isolated Pseudomonas aeruginosa strain VIH2 and antagonistic properties against Ralstonia solanacearum. Microb. Pathog. 2017, 111, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Shanmugaiah, V.; Ramesh, S.; Jayaprakashvel, M.; Mathivanan, N. Biocontrol and plant growth promoting potential of a Pseudomonas sp. MML2212 from the rice rhizosphere. Mitt Biol. Bundesanst Land Forstwirtsch 2006, 408, 320–324. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Plant Pathogens | Zone of Inhibition (mm) |
|---|---|---|
| 1. | R. solanacearum | 28.2 ± 0.6 |
| 2. | R. solani | 24.57 ± 0.86 |
| 3. | S. rolfsii | 21.5 ± 0.89 |
| 4. | M. phaseolina | 23.77 ± 0.38 |
| 5. | F. oxysporum | 27.13 ± 0.76 |
| S. No. | Wavelength (cm−1) | Functional Groups |
|---|---|---|
| 1. | 3473.8 | Phenolic OH group |
| 2. | 2981.95 | Methyl group |
| 3. | 1757.15 | Aromatic ring |
| 4. | 1608.7 | C–H groups in C–CH3 compound |
| 5. | 1458.18 | Aryl carbonyl compounds |
| 6. | 1244.09 | C–OH in alcohols |
| 7. | 1055.06 | Ethers |
| 8. | 929.69 | Acid esters |
| S. No. | Concentration (µg/mL) | Zone of Inhibition (mm) | |
|---|---|---|---|
| DAPG | Tetracycline | ||
| 1. | 5 | 0 ± 0 | 0 ± 0 |
| 2. | 10 | 0 ± 0 | 0 ± 0 |
| 3. | 15 | 0 ± 0 | 9.83 ± 1.04 |
| 4. | 20 | 1.83 ± 0.76 | 11.63 ± 0.35 |
| 5. | 25 | 2.33 ± 0.57 | 13.37 ± 0.71 |
| 6. | 30 | 3.33 ± 0.57 | 15.27 ± 0.7 |
| 7. | 35 | 5.67 ± 1.04 | 13.6 ± 0.4 |
| 8. | 40 | 7.67 ± 0.57 | 14.7 ± 0.31 |
| 9. | 45 | 7.83 ± 0.49 | 14.9 ± 0.11 |
| 10. | 50 | 8.4 ± 0.5 | 15.3 ± 0.6 |
| 11. | 55 | 10 ± 0.4 | 15.67 ± 0.58 |
| 12. | 60 | 10.9 ± 0.35 | 16.13 ± 0.23 |
| 13. | 65 | 13.4 ± 0.61 | 16.73 ± 0.25 |
| 14. | 70 | 15.53 ± 0.41 | 17.13 ± 0.23 |
| 15. | 75 | 16.67 ± 1.15 | 18.83 ± 0.29 |
| 16. | 80 | 17.57 ± 0.4 | 19. 67 ± 1.15 |
| 17. | 85 | 18.43 ± 0.55 | 20.5 ± 0.5 |
| 18. | 90 | 19.03 ± 0.35 | 21 ± 1 |
| 19. | 95 | 20.17 ± 1.04 | 22.83 ± 1.04 |
| 20. | 100 | 20.67 ± 0.58 | 23 ± 1 |
| 21. | 110 | 21.5 ± 0.5 | 24.33 ± 0.28 |
| 22. | 115 | 22.33 ± 0.57 | 24 ± 1 |
| 23. | 120 | 22.5 ± 0.86 | 25.5 ± 0.5 |
| S. No. | Treatments | Percentage of Live Cells | No. of Fields |
|---|---|---|---|
| 1. | R. solanacearum | 89 | 9 |
| 2. | R. solanacearum + Tetracycline | 29 | 9 |
| 3. | R. solanacearum + DAPG | 36 | 9 |
| Treatments | Shoot Length (cm) | Root Length (cm) | No. of Branches | Fresh Weight (g) | Dry Weight (g) | Disease Incidence (%) | Biocontrol Efficacy (%) | Biomass Increase (%) |
|---|---|---|---|---|---|---|---|---|
| T1 (Control) | 93.33 cd ± 1.52 | 25.67 f ± 2.08 | 12.0 ef ± 1 | 20.23 d ± 1.06 | 3.36 cde ± 0.45 | 0 ± 0 | 0 ± 0 | 35.45 ef ± 1.11 |
| T2 (R. solanacearum) | 79.0 f ± 1 | 15.0 g ± 1.73 | 11.67 f ± 1.52 | 15.08 e ± 0.68 | 2.31 f ± 0.18 | 94.97 ± 1.87 | 0 ± 0 | 0.0 h ± 0.00 |
| T3 (Antibiotic + R.S) | 96.67 c ± 1.15 | 31.33 e ± 2.51 | 14.33 cde ± 1.15 | 24.7 b ± 1.35 | 4.32 b ± 0.27 | 23.70 ± 0.45 | 75.56 ± 0.88 | 63.76 b ± 3.61 |
| T4 (P. fluorescens culture) (SA) | 112.67 a ± 2.08 | 53.0 a ± 2.64 | 22.33 a ± 1.52 | 27.72 a ± 1.58 | 5.40 a ± 0.21 | 0 ± 0 | 0 ± 0 | 80.65 a ± 7.87 |
| T5 (P. fluorescens culture filtrate) (SA) | 104 b ± 1 | 34.0 cde ± 3.60 | 12.0 ef ± 1 | 22.56 bcd ± 1.3 | 4.34 b ± 0.26 | 0 ± 0 | 0 ± 0 | 49.56 c ± 2.19 |
| T6 (Compound + R.S) (FS) | 104.33 b ± 2.51 | 22.0 f ± 2.64 | 15.67 bc±1.52 | 21.77d ± 2.12 | 4.23 b ± 0.1 | 47.61 ± 1.03 | 49.86 ± 0.23 | 47.54 cd ± 3.13 |
| Soil Drenching | ||||||||
| T7 (Culture + R.S) | 106.0 b ± 2 | 31.67 de±2.51 | 17.0 b±1.73 | 22.1 cd ± 1.53 | 4.43 b ± 0.12 | 46.66 ± 1.44 | 50.85±1.48 | 46.61 cd ±9.28 |
| T8 (Culture filtrate + R.S) | 86.33 e ± 1.15 | 23.33 f ± 2.08 | 16.33bc ± 0.57 | 20.06 d ± 1.07 | 3.22 de ± 0.19 | 70.06 ± 2.08 | 26.21 ± 2.12 | 29.99 f ± 6.18 |
| Root Dipping | ||||||||
| T9 (Culture + R.S) | 92.33 d ± 1.15 | 40.0 b ± 1.00 | 21.67 a ± 1.52 | 24.31 bc ± 1.24 | 3.28 de ± 0.38 | 69.73 ± 2.54 | 26.56 ± 2.71 | 61.27 b ± 5.24 |
| T10 (Culture filtrate + R.S) | 72.0 g ± 3 | 25.0 c ± 2.00 | 13.0 def ± 1 | 15.83 e ± 1.08 | 2.98 e ± 0.19 | 71.01 ± 1.34 | 25.18 ± 0.32 | 16.38 g ± 1.65 |
| Seed Treatment | ||||||||
| T11 (Culture + R.S) | 83.67 e ± 1.52 | 24.33 f ± 1.52 | 14.33 cde ± 0.57 | 21.46 d ± 0.52 | 3.26 de ± 0.18 | 46.75 ± 1.32 | 50.76 ± 1.32 | 42.42cde ± 3.41 |
| T12 (Culture filtrate + R.S) | 104.33 b ± 3.21 | 35.67 cd ± 1.1 | 15.33 bcd ± 2.08 | 22.42 bcd ± 2.1 | 3.79 c ± 0.29 | 70.83 ± 1.36 | 25.41 ± 0.72 | 48.50 cd ± 7.91 |
| Foliar Spray | ||||||||
| T13 (Culture + R.S) | 95.33 cd ± 1.52 | 31.67 de ± 2.08 | 12.33 ef ± 0.57 | 21.05 d ± 1.22 | 3.70 cd ± 0.24 | 70.1 ± 2.02 | 26.17 ± 2.05 | 39.60 de ± 5.58 |
| T14 (Culture filtrate + R.S) | 83.67 e ± 2.51 | 36.0 c ± 2.04 | 17.67 b ± 1.52 | 21.47 d ± 0.85 | 3.53 cd ± 0.21 | 48.74 ± 2.63 | 48.68 ± 2.28 | 41.54 cde ± 4.38 |
| LSD (0.05) | 3.41 | 3.77 | 2.20 | 2.25 | 0.43 | - | - | 8.60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suresh, P.; Rekha, M.; Gomathinayagam, S.; Ramamoorthy, V.; Sharma, M.P.; Sakthivel, P.; Sekar, K.; Valan Arasu, M.; Shanmugaiah, V. Characterization and Assessment of 2, 4-Diacetylphloroglucinol (DAPG)-Producing Pseudomonas fluorescens VSMKU3054 for the Management of Tomato Bacterial Wilt Caused by Ralstonia solanacearum. Microorganisms 2022, 10, 1508. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081508
Suresh P, Rekha M, Gomathinayagam S, Ramamoorthy V, Sharma MP, Sakthivel P, Sekar K, Valan Arasu M, Shanmugaiah V. Characterization and Assessment of 2, 4-Diacetylphloroglucinol (DAPG)-Producing Pseudomonas fluorescens VSMKU3054 for the Management of Tomato Bacterial Wilt Caused by Ralstonia solanacearum. Microorganisms. 2022; 10(8):1508. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081508
Chicago/Turabian StyleSuresh, Perumal, Murukesan Rekha, Subramanian Gomathinayagam, Vellaisamy Ramamoorthy, Mahaveer P. Sharma, Perumal Sakthivel, Karuppannan Sekar, Mariadhas Valan Arasu, and Vellasamy Shanmugaiah. 2022. "Characterization and Assessment of 2, 4-Diacetylphloroglucinol (DAPG)-Producing Pseudomonas fluorescens VSMKU3054 for the Management of Tomato Bacterial Wilt Caused by Ralstonia solanacearum" Microorganisms 10, no. 8: 1508. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10081508