

Effect of Biogenic Silver Nanoparticles on the Quorum-Sensing System of Pseudomonas aeruginosa PAO1 and PA14
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Bio-AgNPs
2.3. Minimum Inhibitory Concentration (MIC)
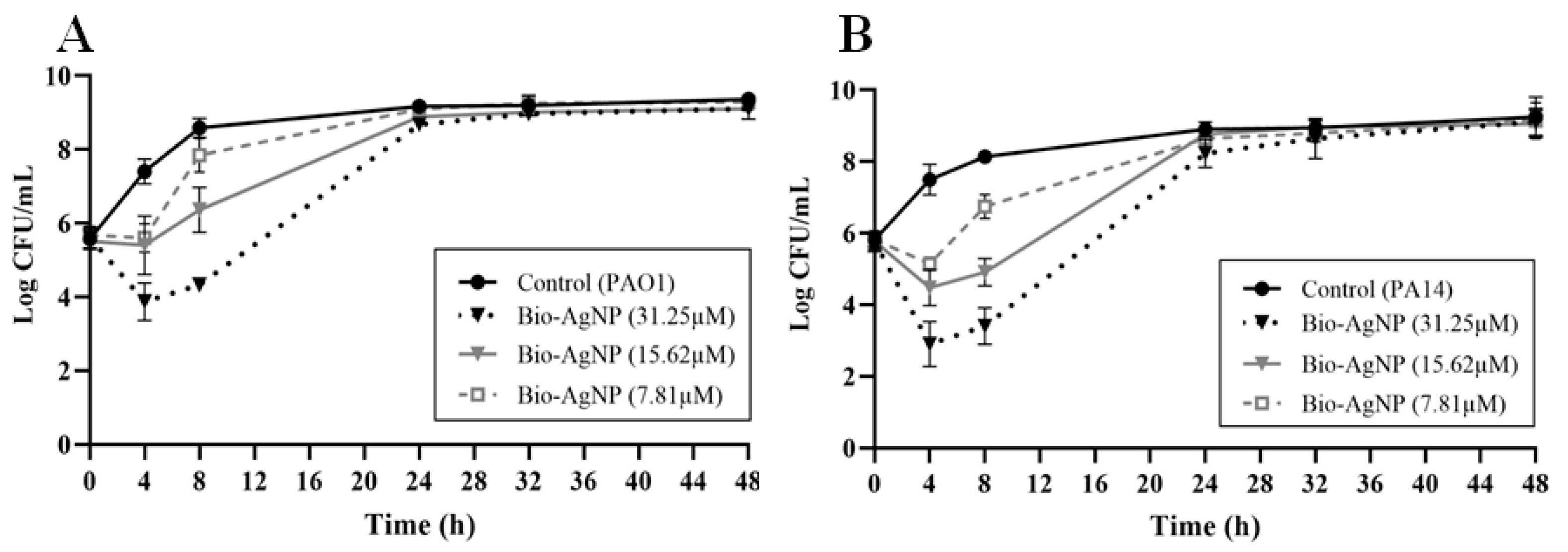
2.4. Growth Curve Analysis
2.5. Assessment of Virulence Factors
2.5.1. Biofilm Formation
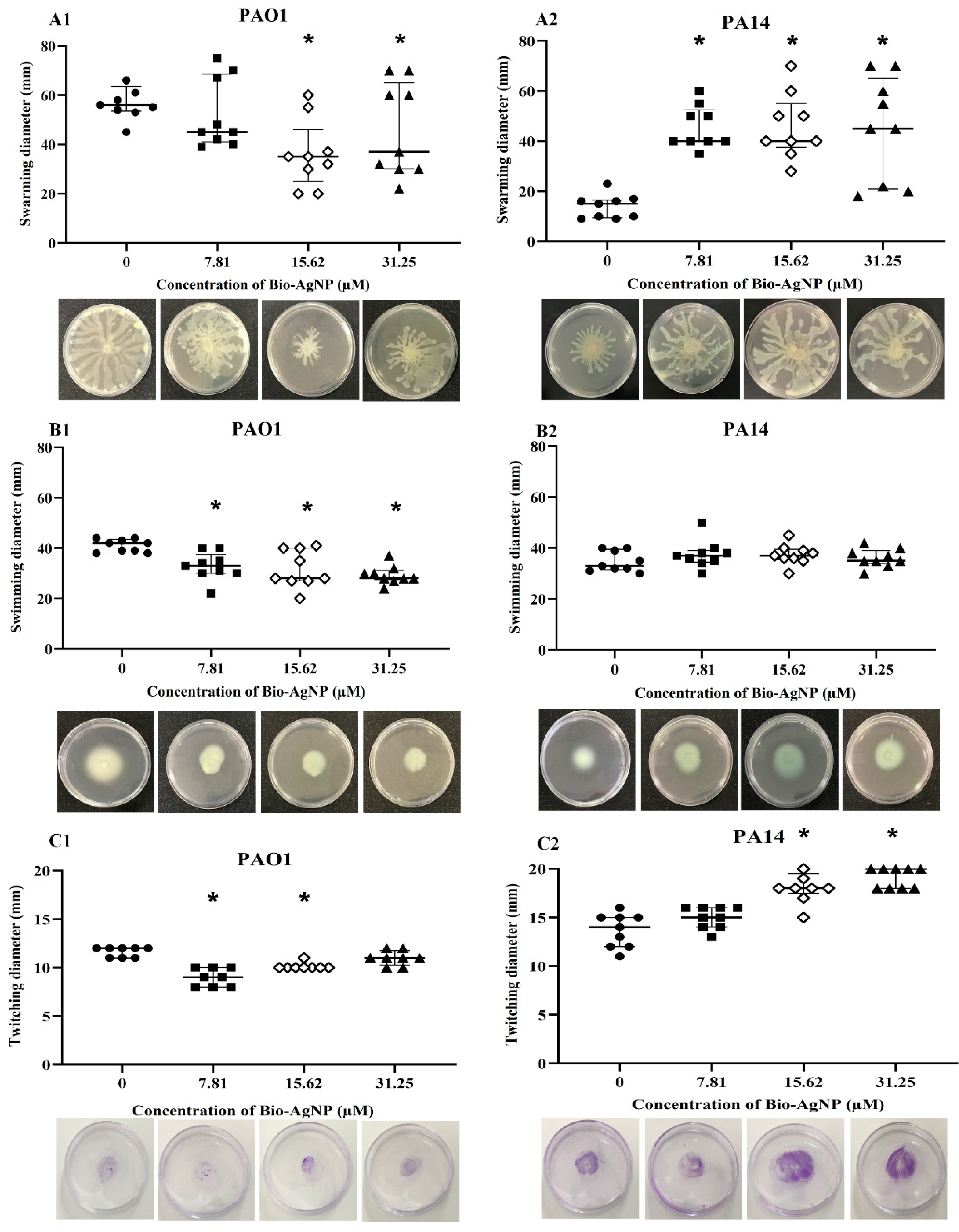
2.5.2. Swarming Motility
2.5.3. Swimming Motility
2.5.4. Twitching Motility
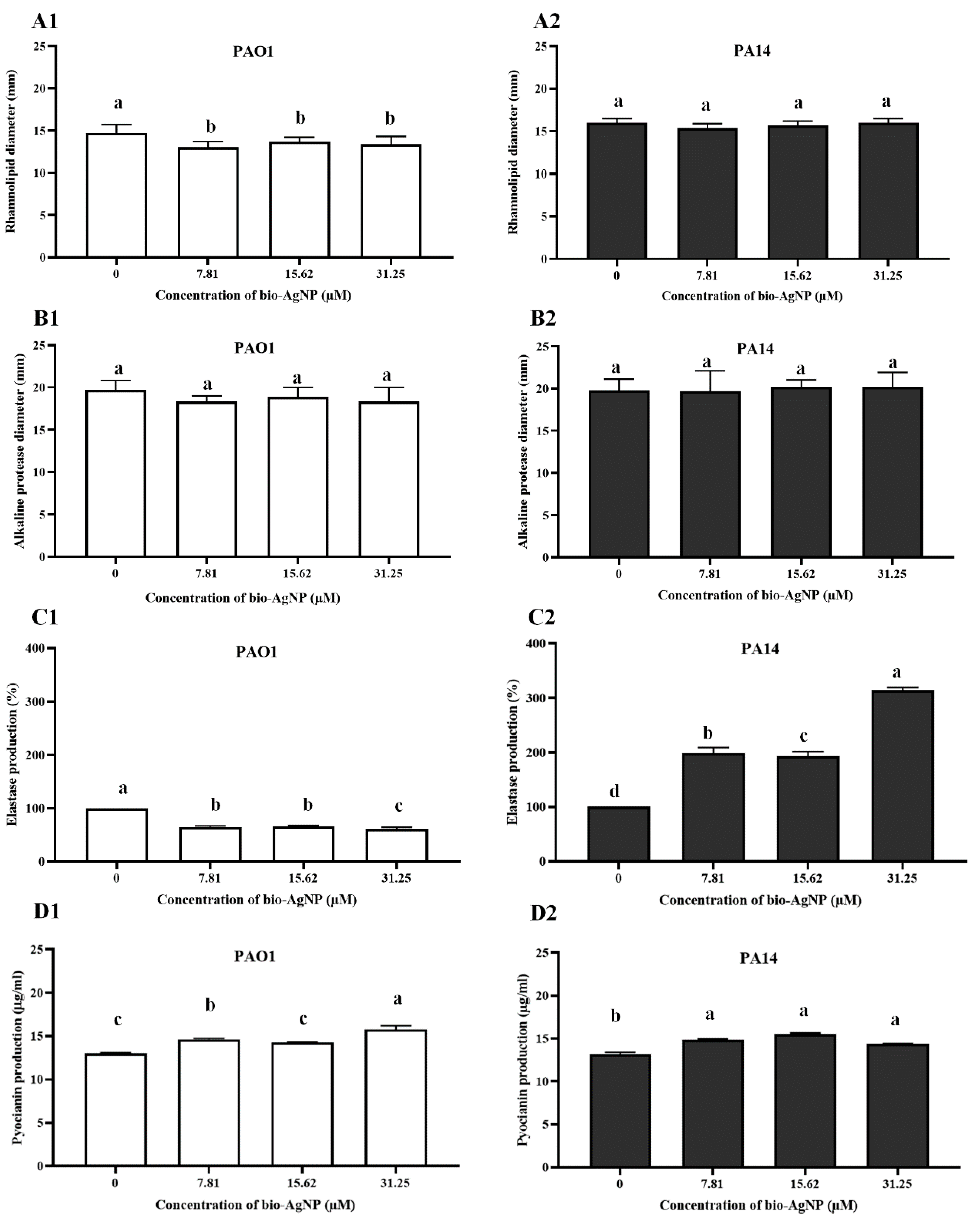
2.5.5. Rhamnolipids
2.5.6. Alkaline Protease
2.5.7. Elastase B (LasB)
2.5.8. Pyocyanin
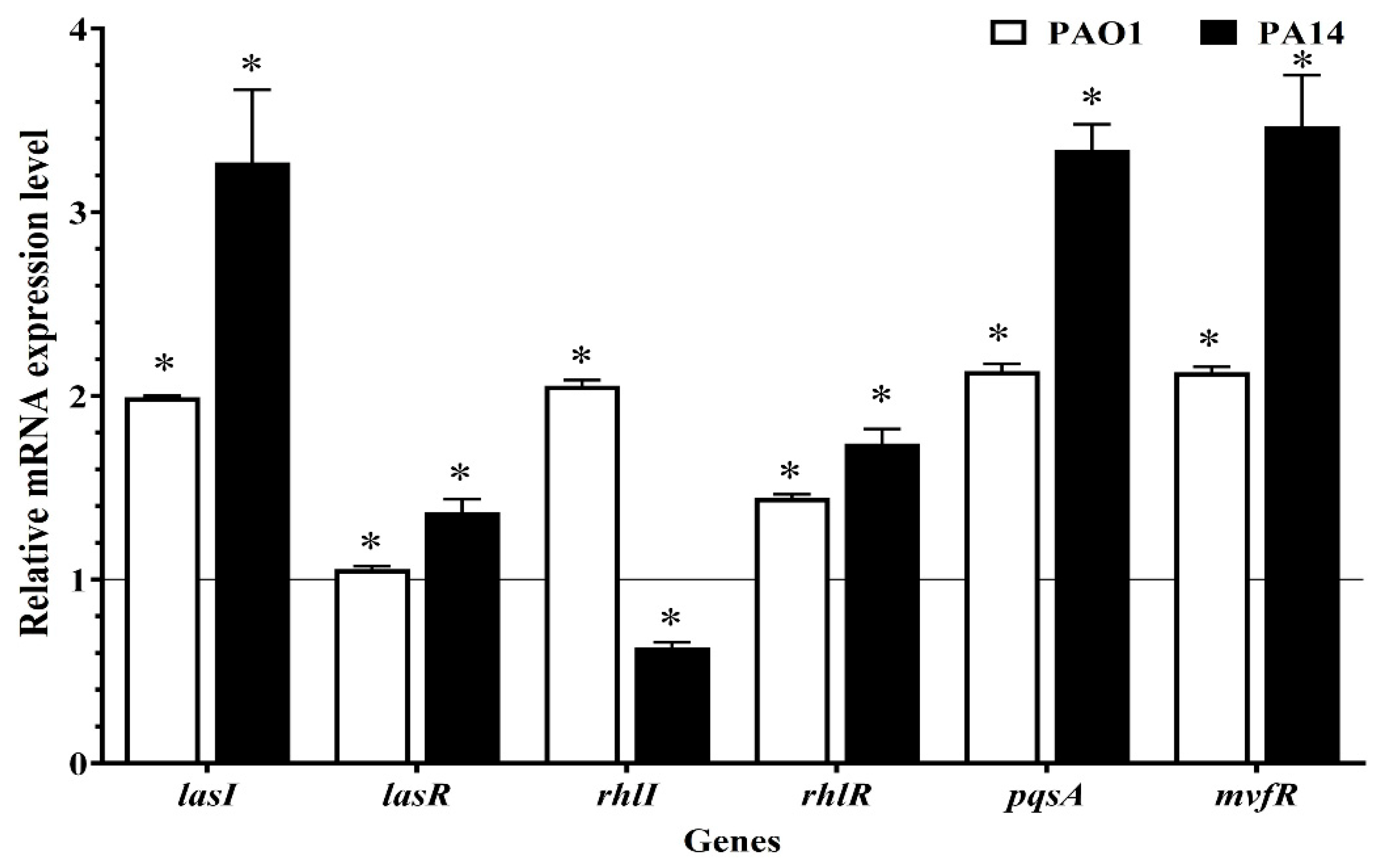
2.6. RNA Extraction and Real-Time Polymerase Chain Reaction (RT-qPCR)
2.7. Statistical Analysis
3. Results
3.1. MIC of bio-AgNPs against P. aeruginosa
3.2. Effect of bio-AgNPs on P. aeruginosa Virulence Factors
3.2.1. Biofilm Formation
3.2.2. Effect on Swarming, Swimming, and Twitching Motilities
3.2.3. Rhamnolipids
3.2.4. Alkaline Protease
3.2.5. LasB
3.2.6. Pyocyanin
3.3. Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tuon, F.F.; Dantas, L.R.; Suss, P.H.; Tasca Ribeiro, V.S. Pathogenesis of the Pseudomonas aeruginosa Biofilm: A Review. Pathogens 2022, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Chadha, J.; Harjai, K.; Chhibber, S. Revisiting the virulence hallmarks of Pseudomonas aeruginosa: A chronicle through the perspective of quorum sensing. Environ. Microbiol. 2022, 24, 2630–2656. [Google Scholar] [CrossRef] [PubMed]
- Cillóniz, C.; Dominedò, C.; Torres, A. Multidrug Resistant Gram-Negative Bacteria in Community-Acquired Pneumonia. Crit. Care 2019, 23, 79. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Fleitas Martínez, O.; Cardoso, M.H.; Ribeiro, S.M.; Franco, O.L. Recent Advances in Anti-virulence Therapeutic Strategies with a Focus on Dismantling Bacterial Membrane Microdomains, Toxin Neutralization, Quorum-Sensing Interference and Biofilm Inhibition. Front. Cell. Infect. Microbiol. 2019, 2, 74. [Google Scholar] [CrossRef]
- Piewngam, P.; Chiou, J.; Chatterjee, P.; Otto, M. Alternative approaches to treat bacterial infections: Targeting quorum-sensing. Expert. Rev. Anti. Infect. Ther. 2020, 18, 499–510. [Google Scholar] [CrossRef]
- Li, Q.; Mao, S.; Wang, H.; Ye, X. The Molecular Architecture of Pseudomonas aeruginosa Quorum-Sensing Inhibitors. Mar. Drugs 2022, 20, 488. [Google Scholar] [CrossRef]
- García-Contreras, R. Is Quorum Sensing interference a viable alternative to treat Pseudomonas aeruginosa infections? Front. Microbiol. 2016, 14, 1454. [Google Scholar] [CrossRef]
- Defoirdt, T. Quorum-Sensing Systems as Targets for Antivirulence Therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef]
- Castillo-Juárez, I.; Maeda, T.; Mandujano-Tinoco, E.A.; Tomás, M.; Pérez-Eretza, B.; García-Contreras, S.J.; Wood, T.K.; García-Contreras, R. Role of quorum sensing in bacterial infections. World J. Clin. Cases 2015, 16, 575–598. [Google Scholar] [CrossRef]
- Goswami, J. Quorum Sensing by Super Bugs and their Resistance to Antibiotics: A Short Review. Glob. J. Pharmaceu. Sci. 2017, 3, 555614. [Google Scholar]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 11, 576–588. [Google Scholar] [CrossRef]
- Lee, J.; Wu, J.; Deng, Y.Y.; Wang, J.; Wang, C.; Wang, J.; Chang, C.; Dong, Y.; Williams, P.; Zhang, L.-H. A cell-cell communication signal integrates quorum sensing and stress response. Nat. Chem. Biol. 2013, 9, 339–343. [Google Scholar] [CrossRef]
- Lee, J.; Zhang, L. The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell. 2015, 6, 26–41. [Google Scholar] [PubMed]
- Cornelis, P. Putting an end to the Pseudomonas aeruginosa IQS controversy. Microbiologyopen 2020, 9, e962. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Pattnaik, S.; Khan, M.B.; Ampasala, D.R.; Busi, S.; Sarma, V.V. Inhibition of quorum sensing-associated virulence factors and biofilm formation in Pseudomonas aeruginosa PAO1 by Mycoleptodiscus indicus PUTY1. Braz. J. Microbiol. 2020, 51, 467–487. [Google Scholar] [CrossRef] [PubMed]
- Durán, N.; Marcato, P.D.; Alves, O.L.; Souza, G.I.H.; Esposito, E. Mechanistic aspects of biosynthesis of silver nanoparticles by several Fusarium oxysporum strains. J. Nanobiotechnol. 2005, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Scandorieiro, S.; Rodrigues, B.C.D.; Nishio, E.K.; Panagio, L.A.; de Oliveira, A.G.; Durán, N.; Nakazato, G.; Kobayashi, R.K.T. Biogenic Silver Nanoparticles Strategically Combined With Origanum vulgare Derivatives: Antibacterial Mechanism of Action and Effect on Multidrug-Resistant Strains. Front. Microbiol. 2022, 6, 842600. [Google Scholar] [CrossRef]
- CLSI-Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. In Performance Standards for Antimicrobial Susceptibility Testing, 10th ed.; Approved Standard; M07-A9; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Jorgensen, J.H. Antimicrobial susceptibility testing of bacteria that grow aerobically. Infect. Dis. Clin. N. Am. 1993, 7, 393–409. [Google Scholar] [CrossRef]
- Ramos-Vivas, J.; Chapartegui-González, I.; Fernández-Martínez, M.; González-Rico, C.; Fortún, J.; Escudero, R.; Marco, F.; Linares, L.; Montejo, M.; Aranzamendi, M.; et al. Biofilm formation by multidrug resistant Enterobacteriaceae strains isolated from solid organ transplant recipients. Sci. Rep. 2019, 9, 8928. [Google Scholar] [CrossRef]
- Norizan, S.N.; Yin, W.F.; Chan, K.G. Caffeine as a potential quorum sensing inhibitor. Sensors 2013, 13, 5117–5129. [Google Scholar] [PubMed]
- Inoue, T.; Shingaki, R.; Fukui, K. Inhibition of swarming motility of Pseudomonas aeruginosa by branched-chain fatty acids. FEMS Microbiol. Lett. 2008, 281, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Otton, L.M.; Campos, M.S.; Meneghetti, K.L.; Corção, G. Influence of twitching and swarming motilities on biofilm formation in Pseudomonas strains. Arch. Microbiol. 2017, 199, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Siegmund, I.; Wagner, F. New method for detecting rhamnolipids excreted by Pseudomonas species during growth on mineral agar. Biotechnol. Tech. 1991, 4, 265–268. [Google Scholar] [CrossRef]
- Aybey, A.; Demirkan, E. Inhibition of quorum sensing-controlled virulence factors in Pseudomonas aeruginosa by human serum paraoxonase. J. Med. Microbiol. 2016, 65, 106–113. [Google Scholar] [CrossRef]
- Banerjee, M.; Moulick, S.; Bhattacharya, K.K.; Parai, D.; Chattopadhyay, S.; Mukherjee, S.K. Attenuation of Pseudomonas aeruginosa quorum sensing, virulence and biofilm formation by extracts of Andrographis paniculata. Microb. Pathog. 2017, 113, 85–93. [Google Scholar] [CrossRef]
- El-Mowafy, S.A.; El Galil, K.H.A.; Habib, E.E.; Shaaban, M.I. Quorum sensing inhibitory activity of sub-inhibitory concentrations of β-lactams. Afr. Health. Sci. 2017, 17, 199–207. [Google Scholar] [CrossRef]
- Li, N.; Wang, L.; Yan, H.; Wang, M.; Shen, D.; Yin, J.; Shentu, J. Effects of low-level engineered nanoparticles on the quorum sensing of Pseudomonas aeruginosa PAO1. Environ. Sci. Pollut. Res. Int. 2018, 25, 7049–7058. [Google Scholar] [CrossRef]
- Ouyang, K.; Mortimer, M.; Holden, P.A.; Cai, P.; Wu, Y.; Gao, C.; Huang, Q. Towards a better understanding of Pseudomonas putida biofilm formation in the presence of ZnO nanoparticles (NPs): Role of NP concentration. Environ. Int. 2020, 137, 105485. [Google Scholar] [CrossRef]
- Scandorieiro, S.; de Camargo, L.C.; Lancheros, C.A.; Yamada-Ogatta, S.F.; Nakamura, C.V.; de Oliveira, A.G.; Andrade, C.J.T.J.; Duran, N.; Nakazato, G.; Kobayashi, R.K.T. Synergistic and Additive Effect of Oregano Essential Oil and Biological Silver Nanoparticles against Multidrug-Resistant Bacterial Strains. Front. Microbiol. 2016, 7, 760. [Google Scholar] [CrossRef]
- Harrison, E.M.; Carter, M.E.; Luck, S. Pathogenicity islands PAPI-1 and PAPI-2 contribute individually and synergistically to the virulence of Pseudomonas aeruginosa strain PA14. Infect. Immun. 2010, 78, 1437–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikkelsen, H.; McMullan, R.; Filloux, A. The Pseudomonas aeruginosa reference strain PA14 displays increased virulence due to a mutation in ladS. PLoS ONE 2011, 6, e29113. [Google Scholar] [CrossRef] [PubMed]
- Sawa, T.; Momiyama, K.; Mihara, T.; Kainuma, A.; Kinoshita, M.; Moriyama, K. Molecular epidemiology of clinically high-risk Pseudomonas aeruginosa strains: Practical overview. Microbiol. Immunol. 2020, 64, 331–344. [Google Scholar] [CrossRef]
- Lee, D.G.; Urbach, J.M.; Wu, G.; Liberati, N.T.; Feinbaum, R.L.; Miyata, S.; Diggins, L.T.; He, J.; Saucier, M.; Déziel, E.; et al. Genomic analysis reveals that Pseudomonas aeruginosa virulence is combinatorial. Genome Biol. 2006, 7, R90. [Google Scholar] [CrossRef]
- Kasetty, S.; Katharios-Lanwermeyer, S.; O’Toole, G.A.; Nadell, C.D. Differential surface competition and biofilm invasion strategies of Pseudomonas aeruginosa PA14 and PA01. J. Bacteriol. 2021, 203, e0026521. [Google Scholar] [PubMed]
- Colvin, K.M.; Gordon, V.D.; Murakami, K.; Borlee, B.R.; Wozniak, D.J.; Wong, G.C.; Parsek, M.R. The pel polysaccharide can serve a structural and protective role in the biofilm matrix of Pseudomonas aeruginosa. PLoS Pathog. 2011, 7, e1001264. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.K.; Vachier, J.; De Anda, J.; Zhao, K.; Baker, A.E.; Bennett, R.R.; Armbruster, C.R.; Lewis, K.A.; Tarnpol, R.L.; Lomba, C.J.; et al. Social cooperativity during surface attachment in early biofilms: A quantitative comparison between reversible attachment in P. aeruginosa PA14 and PAO1. mBio 2020, 11, e02644-19. [Google Scholar] [CrossRef]
- Skariyachan, S.; Sridhar, V.S.; Packirisamy, S.; Kumargowda, S.T.; Challapilli, S.B. Recent perspectives on the molecular basis of biofilm formation by Pseudomonas aeruginosa and approaches for treatment and biofilm dispersal. Folia Microbiol. 2018, 63, 413–432. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef]
- Brindhadevi, K.; Lewisoscar, F.; Mylonakis, E.; Shanmugam, S.; Verma, T.N.; Pugazhendhi, A. Biofilm and Quorum sensing mediated pathogenicity in Pseudomonas aeruginosa. Process. Biochem. 2020, 96, 49–57. [Google Scholar] [CrossRef]
- Rasamiravaka, T.; Labtani, Q.; Duez, P.; El Jaziri, M. The Formation of Biofilms by Pseudomonas aeruginosa: A Review of the Natural and Synthetic Compounds Interfering with Control Mechanisms. BioMed Res. Int. 2015, 2015, 759348. [Google Scholar] [PubMed] [Green Version]
- Das, T.; Kutty, S.K.; Tavallaie, R.; Ibugo, A.I.; Panchompoo, J.; Sehar, S.; Aldous, L.; Yeung, A.W.S.; Thomas, S.R.; Kumar, N.; et al. Phenazine virulence factor binding to extracellular DNA is important for Pseudomonas aeruginosa biofilm formation. Sci. Rep. 2015, 5, 8398. [Google Scholar] [PubMed]
- Newman, J.W.; Floyd, R.V.; Fothergill, J.L. The contribution of Pseudomonas aeruginosa virulence factors and host factors in the establishment of urinary tract infections. FEMS Microbiol. Lett. 2017, 364, fnx124. [Google Scholar]
- Rossi, E.; Paroni, M.; Landini, P. Biofilm and motility in response to environmental and host-related signals in Gram negative opportunistic pathogens. J. Appl. Microbiol. 2018, 125, 1587–1602. [Google Scholar]
- Persat, A.; Inclan, Y.F.; Engel, J.N.; Stone, H.A.; Gitai, Z. Type IV pili mechanochemically regulate virulence factors in Pseudomonas aeruginosa. PNAS 2015, 112, 7563–7568. [Google Scholar]
- Garuglieri, E.; Cattò, C.; Villa, F.; Zanchi, R.; Cappitelli, F. Effects of sublethal concentrations of silver nanoparticles on Escherichia coli and Bacillus subtilis under aerobic and anaerobic conditions. Biointerphases 2016, 11, 04B308. [Google Scholar]
- Saeki, E.K.; Yamada, A.Y.; de Araujo, L.A.; Anversa, L.; Garcia, D.O.; Souza, R.L.B.; Martins, H.M.; Kobayashi, R.K.T.; Nakazato, G. Subinhibitory concentrations of biogenic silver nanoparticles affect motility and biofilm formation in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2021, 11, 656984. [Google Scholar]
- Villa, F.; Remelli, W.; Forlani, F.; Gambino, M.; Landini, P.; Cappitelli, F. Effects of chronic sub-lethal oxidative stress on biofilm formation by Azotobacter vinelandii. Biofouling 2012, 28, 823–833. [Google Scholar]
- Galdino, A.C.M.; Branquinha, M.H.; Santos, A.L.S.; Viganor, L. Pseudomonas aeruginosa and Its Arsenal of Proteases: Weapons to Battle the Host. In Pathophysiological Aspects of Proteases; Chakraborti, S., Dhalla, N., Eds.; Springer: Singapore, 2017. [Google Scholar]
- Yu, H.; He, X.; Xie, W.; Xiong, J.; Sheng, H.; Guo, S.; Huang, C.; Zhang, D.; Zhang, K. Elastase LasB of Pseudomonas aeruginosa promotes biofilm formation partly through rhamnolipid-mediated regulation. Can. J. Microbiol. 2014, 60, 227–235. [Google Scholar]
- Hoge, R.; Pelzer, A.; Rosenau, F.; Wilhelm, S.; Duesseldorf, H. Weapons of a pathogen: Proteases and their role in virulence of Pseudomonas aeruginosa. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 2, 383–395. [Google Scholar]
- Song, Y.; Cai, Z.H.; Lao, Y.M.; Jin, H.; Ying, K.-Z.; Lin, G.-H.; Zhou, J. Antibiofilm activity substances derived from coral symbiotic bacterial extract inhibit biofouling by the model strain Pseudomonas aeruginosa PAO1. Microb. Biotechnol. 2018, 11, 1090–1105. [Google Scholar] [PubMed]
- Rathinam, P.; Vijay Kumar, H.S.; Viswanathan, P. Eugenol exhibits anti-virulence properties by competitively binding to quorum sensing receptors. Biofouling 2017, 33, 624–639. [Google Scholar]
- Liao, S.; Zhang, Y.; Pan, X.; Zhu, F.; Jiang, C.; Liu, Q.; Cheng, Z.; Dai, G.; Wu, G.; Wang, L.; et al. Antibacterial activity and mechanism of silver nanoparticles against multidrug-resistant Pseudomonas aeruginosa. J. Nanomed. 2019, 14, 1469–1487. [Google Scholar]
- Cugini, C.; Morales, D.K.; Hogan, D.A. Candida albicans-produced farnesol stimulates Pseudomonas quinolone signal production in LasR-defective Pseudomonas aeruginosa strains. Microbiology 2010, 156, 3096–3107. [Google Scholar] [PubMed]
- Chuang, S.K.; Vrla, G.D.; Fröhlich, K.S.; Gitai, Z. Surface association sensitizes Pseudomonas aeruginosa to quorum sensing. Nat. Commun. 2019, 10, 4118. [Google Scholar]
- Yang, Y.; Alvarez, P.J.J. Sublethal Concentrations of Silver Nanoparticles Stimulate Biofilm Development. Environ. Sci. Technol. Lett. 2015, 2, 221–226. [Google Scholar]
- Singh, B.R.; Singh, B.N.; Singh, A.; Khan, W.; Naqvi, A.H.; Sigh, H.B. Mycofabricated biosilver nanoparticles interrupt Pseudomonas aeruginosa quorum sensing systems. Sci. Rep. 2015, 5, 13719. [Google Scholar] [PubMed] [Green Version]
- Liu, L.; Li, J.; Zi, S.; Liu, F.; Deng, C.; Ao, X.; Zhang, P. AgNP combined with quorum sensing inhibitor increased the antibiofilm effect on Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2019, 103, 6195–6204. [Google Scholar]
- Mohanty, A.; Tan, C.H.; Cao, B. Impacts of nanomaterials on bacterial quorum sensing: Differential effects on different signals. Environ. Sci. Nano 2016, 3, 351–356. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | Sequence | Amplicon Size (bp) |
|---|---|---|---|
| proC | F | 5′-GAG CAA CTG ATC GTC TCC ATC-3′ | 100 |
| R | 5′-GGG TGT TGG GCA TGC AG-3′ | ||
| lasI | F | 5′-GTA GGC GTG GAG AAG ATG ATG-3′ | 122 |
| R | 5′-ATC TGG GTC TTG GCA TTG AG-3′ | ||
| lasR | F | 5′-CTG TGG ATG CTC AAG GAC TAC-3′ | 111 |
| R | 5′-CCA CTG CAA CAC TTC CTT CT-3′ | ||
| rhlI | F | 5′-GTC GGT CTG GGA GCT TTC-3′ | 100 |
| R | 5′-CAG GTA CCA GGC GCA TT-3′ | ||
| rhlR | F | 5′-CTG TGG TGG GAC GGT TTG-3′ | 139 |
| R | 5′-GGG TGA AGG GAA TCG TGT G-3′ | ||
| pqsA | F | 5′-GCT GTA TTC GAT TCC CAA GAT G-3′ | 100 |
| R | 5′-CCA GGT ATC GTC GAG CAG-3′ | ||
| mvfR | F | 5′-GCT TCG CCT GAT CCC TTA C-3′ | 104 |
| R | 5′-GCA GCA CCC GGA GAT TG-3′ |
| Strain | Compound | Concentration | Biofilm Formation 1 | Inhibition Rate (%) 2 | Increase Rate (%) 2 |
|---|---|---|---|---|---|
| PAO1 | Controle | 0 µM | 0.260 ± 0.040 | - | - |
| PAO1 | bio-AgNP | 7.81 µM | 0.257 ± 0.021 | 1.31% | - |
| PAO1 | bio-AgNP | 15.62 µM | 0.256 ± 0.051 | 1.73% | - |
| PAO1 | bio-AgNP | 31.25 µM | 0.238 ± 0.068 | 8.74% | - |
| PA14 | Controle | 0 µM | 0.052 ± 0.007 c | - | - |
| PA14 | bio-AgNP | 7.81 µM | 0.118 ± 0.015 a | - | 127.86% |
| PA14 | bio-AgNP | 15.62 µM | 0.085 ± 0.020 b | - | 64.09% |
| PA14 | bio-AgNP | 31.25 µM | 0.073 ± 0.013 b | - | 41.22% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeki, E.K.; Martins, H.M.; Camargo, L.C.d.; Anversa, L.; Tavares, E.R.; Yamada-Ogatta, S.F.; Lioni, L.M.Y.; Kobayashi, R.K.T.; Nakazato, G. Effect of Biogenic Silver Nanoparticles on the Quorum-Sensing System of Pseudomonas aeruginosa PAO1 and PA14. Microorganisms 2022, 10, 1755. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091755
Saeki EK, Martins HM, Camargo LCd, Anversa L, Tavares ER, Yamada-Ogatta SF, Lioni LMY, Kobayashi RKT, Nakazato G. Effect of Biogenic Silver Nanoparticles on the Quorum-Sensing System of Pseudomonas aeruginosa PAO1 and PA14. Microorganisms. 2022; 10(9):1755. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091755
Chicago/Turabian StyleSaeki, Erika Kushikawa, Heloísa Moreira Martins, Larissa Ciappina de Camargo, Laís Anversa, Eliandro Reis Tavares, Sueli Fumie Yamada-Ogatta, Lucy Megumi Yamauchi Lioni, Renata Katsuko Takayama Kobayashi, and Gerson Nakazato. 2022. "Effect of Biogenic Silver Nanoparticles on the Quorum-Sensing System of Pseudomonas aeruginosa PAO1 and PA14" Microorganisms 10, no. 9: 1755. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091755