
Prevalence, Subtype Distribution and Zoonotic Significance of Blastocystis sp. Isolates from Poultry, Cattle and Pets in Northern Egypt
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
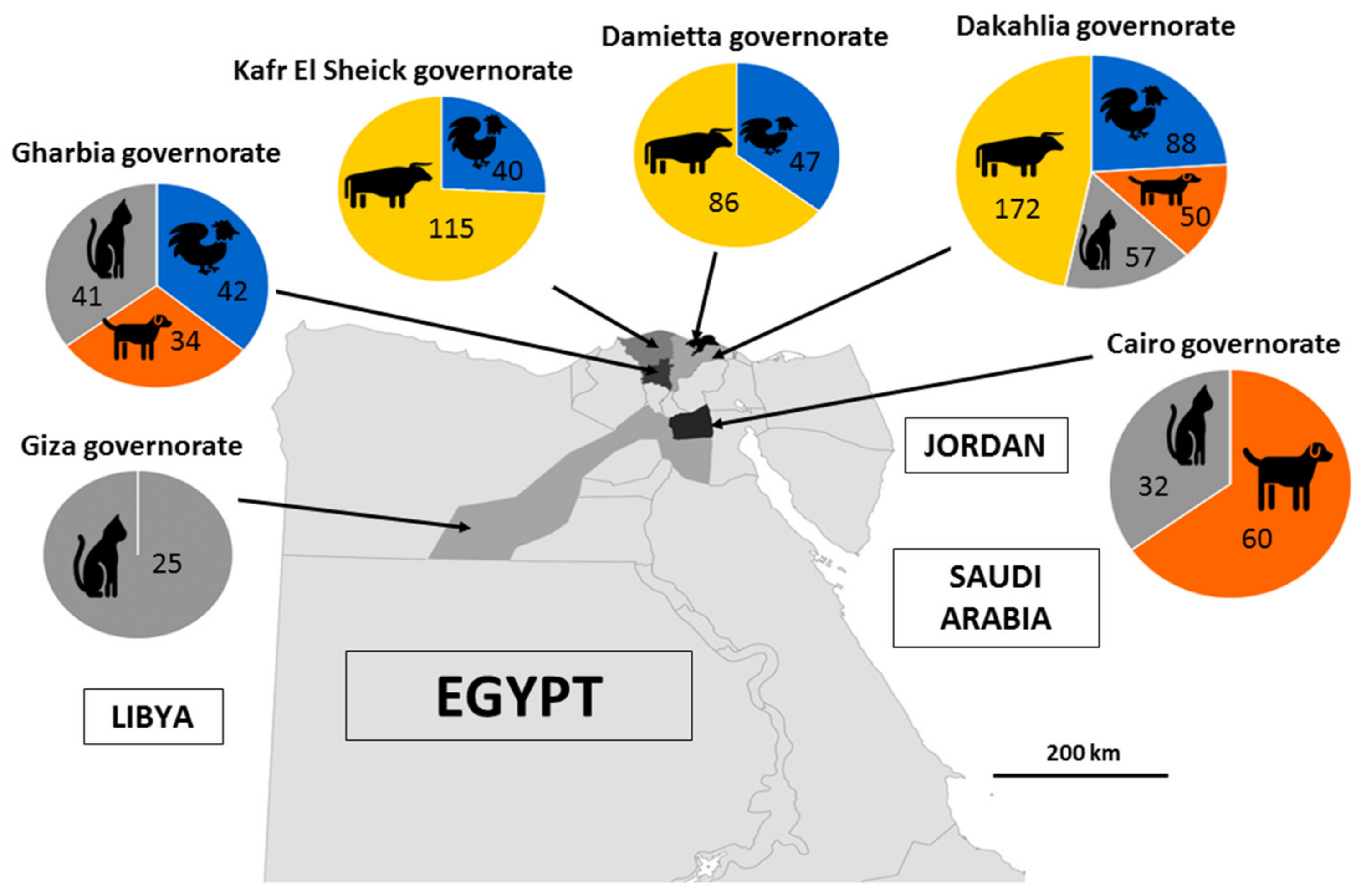
2.2. Specimen Collection
2.3. DNA Extraction
2.4. Blastocystis sp. Detection and Subtyping of Isolates
2.5. Statistical Analysis
3. Results and Discussion

{kind=link}
| Number of Subtyped Isolates | Subtyping Method | Blastocystis sp. STs | MI a | Not Typed | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| ST1 | ST3 | ST4 | ST5 | ST10 | ST14 | |||||
| 7 | Sequencing | 0 | 0 | 1 | 0 | 1 | 5 | 0 | 0 | [37] |
| 13 | PCR-STS b | 2 | 3 | 0 | 2 | 0 | 0 | 0 | 6 | [36] |
| 41 | Sequencing | 0 | 1 | 1 | 0 | 13 | 10 | 16 | 0 | Present study |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hublin, J.S.Y.; Maloney, J.G.; Santin, M. Blastocystis in domesticated and wild mammals and birds. Res. Vet. Sci. 2021, 135, 260–282. [Google Scholar] [CrossRef]
- Nemati, S.; Reza Zali, M.; Johnson, P.; Mirjalali, H.; Karanis, P. Molecular prevalence and subtype distribution of Blastocystis sp. in Asia and Australia. J. Water Health 2021, 19, 687. [Google Scholar] [CrossRef] [PubMed]
- Rauff-Adetotun, A.A.; Meor Termizi, F.H.; Shaari, N.; Lee, I.L. The coexistence of Blastocystis spp. in humans, animals and environmental sources from 2010–2021 in Asia. Biology 2021, 10, 990. [Google Scholar] [CrossRef] [PubMed]
- El Safadi, D.; Cian, A.; Nourrisson, C.; Pereira, B.; Morelle, C.; Bastien, P.; Bellanger, A.P.; Botterel, F.; Candolfi, E.; Desoubeaux, G.; et al. Prevalence, risk factors for infection and subtype distribution of the intestinal parasite Blastocystis sp. from a large-scale multi-center study in France. BMC Infect. Dis. 2016, 16, 451. [Google Scholar] [CrossRef] [Green Version]
- Lhotska, Z.; Jirku, M.; Hlozkova, O.; Brozova, K.; Jirsova, D.; Stensvold, C.R.; Kolisko, M.; Pomajbikova, K.J. A study on the prevalence and subtype diversity of the intestinal protist Blastocystis sp. in a gut-healthy human population in the Czech Republic. Front. Cell. Infect. Microbiol. 2020, 10, 544335. [Google Scholar] [CrossRef]
- Muadica, A.S.; Köster, P.C.; Dashti, A.; Bailo, B.; Hernandez-de-Mingo, M.; Reh, L.; Balasegaram, S.; Verlander, N.Q.; Chercoles, E.R.; Carmena, D. Molecular diversity of Giardia duodenalis, Cryptosporidium spp. and Blastocystis sp. in asymptomatic school children in Leganés, Madrid (Spain). Microorganisms 2020, 8, 466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Safadi, D.; Gaayeb, L.; Meloni, D.; Cian, A.; Poirier, P.; Wawrzyniak, I.; Delbac, F.; Dabboussi, F.; Delhaes, L.; Seck, M.; et al. Children of Senegal River Basin show the highest prevalence of Blastocystis sp. ever observed worldwide. BMC Infect. Dis. 2014, 14, 164. [Google Scholar] [CrossRef]
- Lokmer, A.; Cian, A.; Froment, A.; Gantois, N.; Viscogliosi, E.; Chabé, M.; Ségurel, L. Use of shotgun metagenomics for the identification of protozoa in the gut microbiota of healthy individuals from worldwide populations with various industrialization levels. PLoS ONE 2019, 14, e0211139. [Google Scholar] [CrossRef]
- Khaled, S.; Gantois, N.; Tidjani Ly, A.; Senghor, S.; Even, G.; Dautel, E.; Dejager, R.; Sawant, M.; Baydoun, M.; Benamrouz-Vanneste, S.; et al. Prevalence and subtype distribution of Blastocystis sp. in Senegalese school children. Microorganisms 2020, 8, 1408. [Google Scholar] [CrossRef]
- Tan, K.S.W. New insights on classification, identification, and clinical relevance of Blastocystis spp. Clin. Microbiol. Rev. 2008, 21, 639–665. [Google Scholar] [CrossRef]
- Clark, C.G.; van der Giezen, M.; Alfellani, M.A.; Stensvold, C.R. Recent developments in Blastocystis research. Adv. Parasitol. 2013, 82, 1–32. [Google Scholar] [PubMed] [Green Version]
- Stensvold, C.R.; Clark, C.G. Current status of Blastocystis: A personal view. Parasitol. Int. 2016, 65, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Ajjampur, S.S.; Tan, K.S.W. Pathogenic mechanisms in Blastocystis spp.—Interpreting results from in vitro and in vivo studies. Parasitol. Int. 2016, 65, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Wojciech, L.; Gascoigne, N.R.J.; Peng, G.; Tan, K.S.W. New insights into the interactions between Blastocystis, the gut microbiota, and host immunity. PLoS Pathog. 2021, 17, e1009253. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Lewis, H.C.; Hammerun, A.M.; Porsbo, L.J.; Nielsen, S.S.; Olsen, K.E.; Arendrup, M.C.; Nielsen, H.V.; Molbak, K. Blastocystis: Unravelling potential risk factors and clinical significance of a common but neglected parasite. Epidemiol. Infect. 2009, 137, 1655–1663. [Google Scholar] [CrossRef] [Green Version]
- Fréalle, E.; El Safadi, D.; Cian, A.; Aubry, E.; Certad, G.; Osman, M.; Wacrenier, A.; Dutoit, E.; Creusy, C.; Dubos, F.; et al. Acute Blastocystis-associated appendicular peritonitis in a child, Casablanca, Morocco. Emerg. Infect. Dis. 2015, 21, 91–94. [Google Scholar] [CrossRef]
- Aykur, M.; Camyar, A.; Türk, B.G.; Sin, A.Z.; Dagci, H. Evaluation of association with subtypes and alleles of Blastocystis with chronic spontaneous urticarial. Acta Trop. 2022, 231, 106455. [Google Scholar] [CrossRef]
- Audebert, C.; Even, G.; Cian, A.; Blastocystis Investigation Group; Loywick, A.; Merlin, S.; Viscogliosi, E.; Chabé, M. Colonization with the enteric protozoa Blastocystis is associated with increased diversity of human gut bacterial microbiota. Sci. Rep. 2016, 6, 25255. [Google Scholar] [CrossRef]
- Tito, R.Y.; Chaffron, S.; Caenepeel, C.; Lima-Mendez, G.; Wang, J.; Vieira-Silva, S.; Falony, G.; Hildebrand, F.; Darzi, Y.; Rymenans, L.; et al. Population-level analysis of Blastocystis subtype prevalence and variation in the human gut microbiota. Gut 2019, 68, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Even, G.; Lokmer, A.; Rodrigues, J.; Audebert, C.; Viscogliosi, E.; Ségurel, L.; Chabé, M. Changes in the human gut microbiota associated with colonization by Blastocystis sp. and Entamoeba spp. in non-industrialized populations. Front. Cell. Infect. Microbiol. 2021, 11, 533528. [Google Scholar] [CrossRef]
- Yason, J.A.; Liang, Y.R.; Png, C.W.; Zhang, Y.; Tan, K.S.W. Interactions between a pathogenic Blastocystis subtype and gut microbiota: In vitro and in vivo studies. Microbiome 2019, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfellani, M.A.; Taner-Mulla, D.; Jacob, A.S.; Imeede, C.A.; Yoshikawa, H.; Stensvold, C.R.; Clark, C.G. Genetic diversity of Blastocystis in livestock and zoo animals. Protist 2013, 164, 497–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloney, J.G.; Jang, Y.; Molokin, A.; George, N.S.; Santin, M. Wide genetic diversity of Blastocystis in white-tailed deer (Odocoileus virginianus) from Maryland, USA. Microorganisms 2021, 9, 1343. [Google Scholar] [CrossRef] [PubMed]
- Higuera, A.; Herrera, G.; Jimenez, P.; Garcia-Corredor, D.; Pulido-Medellin, M.; Bulla-Castaneda, D.M.; Pinilla, J.C.; Moreno-Perez, D.A.; Maloney, J.G.; Santin, M.; et al. Identification of multiple Blastocystis subtypes in domestic animals from Colombia using amplicon-based next generation sequencing. Front. Vet. Sci. 2021, 8, 732129. [Google Scholar] [CrossRef]
- Baek, S.; Maloney, J.G.; Molokin, A.; George, N.S.; Vecino, J.A.C.; Santin, M. Diversity of Blastocystis subtypes in horses in Colombia and identification of two new subtypes. Microorganisms 2022, 10, 1693. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Clark, C.G. Pre-empting Pandora’s box: Blastocystis subtypes revisited. Trends Parasitol. 2020, 36, 229–232. [Google Scholar] [CrossRef]
- Alfellani, M.A.; Stensvold, C.R.; Vidal-Lapiedra, A.; Onuoha, E.S.; Fagbenro-Beyioku, A.F.; Clark, C.G. Variable geographic distribution of Blastocystis subtypes and its potential implications. Acta Trop. 2013, 126, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, J.D.; Sánchez, A.; Hernández, C.; Florez, C.; Bernal, M.C.; Giraldo, J.C.; Reyes, P.; Lopez, M.C.; Garcia, L.; Cooper, P.J.; et al. Geographic distribution of human Blastocystis subtypes in South America. Infect. Genet. Evol. 2016, 41, 32–35. [Google Scholar] [CrossRef]
- Khaled, S.; Gantois, N.; Ayoubi, A.; Even, G.; Sawant, M.; El Houmayraa, J.; Nabot, M.; Benamrouz-Vanneste, S.; Chabé, M.; Certad, G.; et al. Blastocystis sp. prevalence and subtypes distribution amongst Syrian refugee communities living in North Lebanon. Microorganisms 2021, 9, 184. [Google Scholar] [CrossRef]
- Osario-Pulgarin, M.I.; Higuera, A.; Beltran-Alzate, J.C.; Sanchez-Jimenez, M.; Ramirez, J.D. Epidemiological and molecular characterization of Blastocystis infection in children attending daycare centers in Medellin, Colombia. Biology 2021, 10, 669. [Google Scholar] [CrossRef]
- Jinatham, V.; Maxamhud, S.; Popluechai, S.; Tsaousis, A.D.; Gentekaki, E. Blastocystis One Health approach in a rural community of Northern Thailand: Prevalence, subtypes and novel transmission routes. Front. Microbiol. 2021, 12, 746340. [Google Scholar] [CrossRef] [PubMed]
- Guilavogui, T.; Gantois, N.; Even, G.; Desramaut, J.; Dautel, E.; Denoyelle, C.; Cissé, F.I.; Touré, S.C.; Kourouma, B.L.; Sawant, M.; et al. Detection, molecular identification and transmission of the intestinal protozoa Blastocystis sp. in Guinea from a large-scale epidemiological study conducted in the Conakry area. Microorganisms 2022, 10, 446. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Owen, H.; Traub, R.J.; Cuttell, L.; Inpankaew, T.; Bielefeldt-Ohmann, H. Molecular epidemiology of Blastocystis in pigs and their in-contact humans in Southeast Queensland, Australia, and Cambodia. Vet. Parasitol. 2014, 203, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Greige, S.; El Safadi, D.; Bécu, N.; Gantois, N.; Pereira, B.; Chabé, M.; Benamrouz-Vanneste, S.; Certad, G.; El Hage, R.; Chemaly, M.; et al. Prevalence and subtype distribution of Blastocystis sp. isolates from poultry in Lebanon and evidence of zoonotic potential. Parasit. Vectors 2018, 11, 389. [Google Scholar] [CrossRef] [Green Version]
- Awadallah, M.A.I.; Salem, L.M.A. Zoonotic enteric parasites transmitted from dogs in Egypt with special concern to Toxocara canis infection. Vet. World 2015, 8, 946–957. [Google Scholar] [CrossRef] [Green Version]
- Mokhtar, A.; Youssef, A. Subtype analysis of Blastocystis spp. isolated from domestic mammals and poultry and its relation to transmission to their in-contact humans in Ismailia Governorate, Egypt. Parasitol. United J. 2018, 11, 90–98. [Google Scholar] [CrossRef]
- Abdo, S.M.; El-Adawy, H.; Farag, H.F.; El-Taweel, H.A.; Elhadad, H.; El-Badry, A.A.M. Detection and molecular identification of Blastocystis isolates from humans and cattle in Northern Egypt. J. Parasit. Dis. 2021, 45, 738–745. [Google Scholar] [CrossRef]
- Sayed, F.G.; Galal, L.A.; Elossily, N.A.; Ahmad, R.A.N. Prevalence of Blastocystis spp. in house and farm raised chicken in Assiut. Al-Azhar Assiut Med. J. 2015, 13, 442–449. [Google Scholar]
- Stensvold, C.R.; Arendrup, M.C.; Jespersgaard, C.; Molbak, K.; Nielsen, H.V. Detecting Blastocystis using parasitologic and DNA-based methods: A comparative study. Diagn. Microbiol. Infect. Dis. 2007, 59, 303–307. [Google Scholar] [CrossRef]
- Poirier, P.; Wawrzyniak, I.; Albert, A.; El Alaoui, H.; Delbac, F.; Livrelli, V. Development and evaluation of a real-time PCR assay for detection and quantification of Blastocystis parasites in human stool samples: Prospective study of patients with hematological malignancies. J. Clin. Microbiol. 2011, 49, 975–983. [Google Scholar] [CrossRef] [Green Version]
- Shams, M.; Shamsi, L.; Sadrebazzaz, A.; Asghari, A.; Badali, R.; Omidian, M.; Hassanipour, S. A systematic review and meta-analysis on the global prevalence and subtypes distribution of Blastocystis sp. in infection in cattle: A zoonotic concern. Comp. Immunol. Microbiol. Infect. Dis. 2021, 76, 101650. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gong, B.; Yang, F.; Zhang, W.; Zheng, Y.; Liu, A. Subtype distribution and genetic characterization of Blastocystis in pigs, cattle, sheep and goat in northeastern China’s Heilongjiang province. Infect. Genet. Evol. 2018, 57, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Hastutiek, P.; Yuniarti, W.M.; Djaeri, M.; Lastuti, N.D.R.; Suprihati, E.; Suwanti, L.T. Prevalence and diversity of gastrointestinal protozoa in Madura cattle at Bangkalan Regency, East Java, Indonesia. Vet. World 2019, 12, 198. [Google Scholar] [CrossRef]
- Aynur, Z.E.; Güçlü, Ö.; Yildiz, Ï.; Aynur, H.; Ertabaklar, H.; Bozdogan, B.; Ertug, S. Molecular characterization of Blastocystis in cattle in Turkey. Parasitol. Res. 2019, 118, 1055–1059. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Tao, W.; Gong, B.; Yang, H.; Li, Y.; Song, M.; Lu, Y.; Li, W. First report of Blastocystis infections in cattle in China. Vet. Parasitol. 2017, 246, 38–42. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.H.; Seo, M.G.; Kim, H.Y.; Kim, J.W.; Lee, Y.R.; Kim, J.H.; Kwon, O.D.; Kwak, D. Occurrence and genetic diversity of Blastocystis in Korean cattle. Vet. Parasitol. 2018, 258, 70–73. [Google Scholar] [CrossRef]
- Maloney, J.G.; Lombard, J.E.; Urie, N.J.; Shivley, C.B.; Santin, M. Zoonotic and genetically diverse subtypes of Blastocystis in US pre-weaned dairy heifer calves. Parasitol. Res. 2019, 118, 575–582. [Google Scholar] [CrossRef]
- Fayer, R.; Santin, M.; Macarisin, D. Detection of concurrent infection of dairy cattle with Blastocystis, Cryptosporidium, Giardia and Enterocytozoon by molecular and microscopic methods. Parasitol. Res. 2012, 111, 1349–1355. [Google Scholar] [CrossRef]
- Greige, S.; El Safadi, D.; Khaled, S.; Gantois, N.; Baydoun, M.; Chemaly, M.; Benamrouz-Vanneste, S.; Chabé, M.; Osman, M.; Certad, G.; et al. First report on the prevalence and subtype distribution of Blastocystis sp. in dairy cattle in Lebanon and assessment of zoonotic transmission. Acta Trop. 2019, 194, 23–29. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Tokoro, M.; Nagamoto, T.; Arayama, S.; Asih, P.B.S.; Rozi, I.E.; Syafruddin, D. Molecular survey of Blastocystis sp. from humans and associated animals in an Indonesian community with poor hygiene. Parasitol. Int. 2016, 65, 780–784. [Google Scholar] [CrossRef]
- D’Alfonso, R.; Santoro, M.; Essi, D.; Monsia, A.; Kaboré, Y.; Glé, C.; di Cave, D.; Sorge, R.P.; di Cristianzano, V.; Berrilli, F. Blastocystis in Côte d’Ivoire: Molecular identification and epidemiological data. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2243–2250. [Google Scholar] [CrossRef] [PubMed]
- Shams, M.; Shamsi, L.; Yousefi, A.; Sadrebazzaz, A.; Asghari, A.; Mohammadi-Ghalehbir, B.; Shahabi, S.; Hatam, G. Current global status, subtype distribution and zoonotic significance of Blastocystis in dogs and cats: A systematic review and meta-analysis. Parasit. Vectors 2022, 15, 225. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, X.; Gu, Y.; Liu, J.; Luo, J. Prevalence of Cryptosporidium, Giardia, Blastocystis, and trichomonads in domestic cats in east China. J. Vet. Med. Sci. 2019, 81, 890–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, D.; Seo, M.G. Genetic analysis of zoonotic gastrointestinal protozoa and microsporidia in shelter cats in South Korea. Pathogens 2020, 9, 894. [Google Scholar] [CrossRef] [PubMed]
- Can, H.; Köseoğlu, A.E.; Erkunt Alak, S.; Güvendi, M.; Ün, C.; Karakavuk, M.; Değirmenci Döşkaya, A.; Aykur, M.; Aksoy Gökmen, A.; Gürüz, A.Y.; et al. Molecular prevalence and subtyping of Blastocystis sp. isolates in stray cats of Izmir, Turkey: First report of “ST4 allele 42” in cats. Pol. J. Vet. Sci. 2021, 24, 217. [Google Scholar]
- Ruaux, C.G.; Stang, B.V. Prevalence of Blastocystis in shelter-resident and client-owned companion animals in the US Pacific Northwest. PLoS ONE 2014, 9, e107496. [Google Scholar] [CrossRef]
- Mohammadpour, I.; Bozorg-Ghalati, F.; Gazzonis, A.L.; Manfredi, M.T.; Motazedian, M.H.; Mohammadpour, N. First molecular subtyping and phylogeny of Blastocystis sp. isolated from domestic and synanthropic animals (dogs, cats and brown rats) in southern Iran. Parasit. Vectors 2020, 13, 365. [Google Scholar] [CrossRef]
- Paulos, S.; Köster, P.C.; de Lucio, A.; Hernandez-de-Mingo, M.; Cardona, G.A.; Fernandez-Crespo, J.C.; Stensvold, C.R.; Carmena, D. Occurrence and subtype distribution of Blastocystis sp. in humans, dogs and cats sharing household in northern Spain and assessment of zoonotic transmission risk. Zoonoses Public Health 2018, 65, 993–1002. [Google Scholar] [CrossRef]
- Rudzinska, M.; Kowalewska, B.; Kurpas, M.; Szostakowska, B. Rare occurrence of Blastocystis in pet animals and their owners in the Pomeranian Voivodeship in Poland in the light of literature data. J. Clin. Med. 2022, 11, 2975. [Google Scholar] [CrossRef]
- Spanakos, G.; Papadogiannakis, E.; Kontos, V.; Menounos, P.; Velonakis, E.; Koutis, C.; Vakalis, N. Molecular screening for Blastocystis sp. in canine faecal samples in Greece. J. Hell. Vet. Med. Soc. 2011, 62, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Osman, M.; Bories, J.; El Safadi, D.; Poirel, M.T.; Gantois, N.; Benamrouz-Vanneste, S.; Delhaes, L.; Hugonnard, M.; Certad, G.; Zenner, L.; et al. Prevalence and genetic diversity of the intestinal parasites Blastocystis sp. and Cryptosporidium spp. in household dogs in France and evaluation of zoonotic transmission risk. Vet. Parasitol. 2015, 214, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cuttell, L.; Bielefeldt-Ohmann, H.; Ipankaew, T.; Owen, H.; Traub, R.J. Diversity of Blastocystis subtypes in dogs in different geographical settings. Parasit. Vectors 2013, 6, 215. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Lin, X.; Sun, Y.; Qi, N.; Lv, M.; Wu, C.; Li, J.; Hu, J.; Yu, L.; Cai, H.; et al. Occurrence and genotypes of Cryptosporidium spp., Giardia duodenalis, and Blastocystis sp. in household, shelter, breeding, and pet market dogs in Guangzhou, southern China. Sci. Rep. 2020, 10, 17736. [Google Scholar] [CrossRef] [PubMed]
- Gazzonis, A.L.; Marangi, M.; Zanzani, S.A.; Villa, L.; Giangaspero, A.; Manfredi, M.T. Molecular epidemiology of Blastocystis sp. in dogs housed in Italian rescue shelters. Parasitol. Res. 2019, 118, 3011–3017. [Google Scholar] [CrossRef] [PubMed]

| Animal Group | Governorate | No. Examined | No. Positive (%) | STs (No.) |
|---|---|---|---|---|
| Cattle | Dakahlia | 172 | 27 (15.7) | ST10 (9); ST14 (7); MI a (11) |
| Gharbia | 0 | - | - | |
| Damietta | 86 | 4 (4.7) | ST10 (2); ST14 (1); MI a (1) | |
| Kafr El Sheikh | 115 | 10 (8.7) | ST3 (1); ST4 (1); ST10 (2); ST14 (2); MI a (4) | |
| Cairo | 0 | 0 | - | |
| Giza | 0 | - | - | |
| Subtotal | 373 | 41 (11.0) | ST3 (1); ST4 (1); ST10 (13); ST14 (10); MI b (16) | |
| Chickens | Dakahlia | 88 | 10 (11.4) | ST7 (10) |
| Gharbia | 42 | 10 (23.8) | ST7 (10) | |
| Damietta | 47 | 11 (23.4) | ST7 (10); ST14 (1) | |
| Kafr El Sheikh | 40 | 6 (15.0) | ST7 (6) | |
| Cairo | 0 | - | - | |
| Giza | 0 | - | - | |
| Subtotal | 217 | 37 (17.0) | ST7 (36); ST14 (1) | |
| Cats | Dakahlia | 57 | 1 (1.7) | ST14 (1) |
| Gharbia | 41 | 1 (2.4) | ST14 (1) | |
| Damietta | 0 | - | - | |
| Kafr El Sheikh | 0 | - | - | |
| Cairo | 32 | 0 (0) | ||
| Giza | 25 | 2 (8.0) | ST3 (1); ST14 (1) | |
| Subtotal | 155 | 4 (2.6) | ST3 (1); ST14 (3) | |
| Dogs | Dakahlia | 50 | 0 (0) | - |
| Gharbia | 34 | 0 (0) | - | |
| Damietta | 0 | - | - | |
| Kafr El Sheikh | 0 | - | - | |
| Cairo | 60 | 0 (0) | - | |
| Giza | 0 | - | - | |
| Subtotal | 144 | 0 (0) | - | |
| Total | 889 | 82 (9.2) | ST3 (2); ST4 (1); ST7 (36); ST10 (13); ST14 (14); MI (16) |
| Governorate | Sex | Age in Months | Life Style | Breed | ST |
|---|---|---|---|---|---|
| Gharbia | F | 18 | Veterinary clinic | Persian | ST14 |
| Dakahlia | F | 2 | Shelter | Nile Valley Egyptian | ST14 |
| Giza | M | 9 | Pet shop | Persian | ST3 |
| Giza | F | 12 | Veterinary clinic | Shirazi | ST14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naguib, D.; Gantois, N.; Desramaut, J.; Arafat, N.; Even, G.; Certad, G.; Chabé, M.; Viscogliosi, E. Prevalence, Subtype Distribution and Zoonotic Significance of Blastocystis sp. Isolates from Poultry, Cattle and Pets in Northern Egypt. Microorganisms 2022, 10, 2259. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10112259
Naguib D, Gantois N, Desramaut J, Arafat N, Even G, Certad G, Chabé M, Viscogliosi E. Prevalence, Subtype Distribution and Zoonotic Significance of Blastocystis sp. Isolates from Poultry, Cattle and Pets in Northern Egypt. Microorganisms. 2022; 10(11):2259. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10112259
Chicago/Turabian StyleNaguib, Doaa, Nausicaa Gantois, Jeremy Desramaut, Nagah Arafat, Gaël Even, Gabriela Certad, Magali Chabé, and Eric Viscogliosi. 2022. "Prevalence, Subtype Distribution and Zoonotic Significance of Blastocystis sp. Isolates from Poultry, Cattle and Pets in Northern Egypt" Microorganisms 10, no. 11: 2259. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10112259