
Diversity of Mixotrophic Neutrophilic Thiosulfate- and Iron-Oxidizing Bacteria from Deep-Sea Hydrothermal Vents

Key Laboratory of Marine Biogenetic Resources, Third Institute of Oceanography, Ministry of Natural Resources, Xiamen 361005, China
*
Authors to whom correspondence should be addressed.
Microorganisms 2023, 11(1), 100; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11010100
Submission received: 11 November 2022
/
Revised: 22 December 2022
/
Accepted: 22 December 2022
/
Published: 30 December 2022
(This article belongs to the Special Issue Diversity of Extremophiles in Hydrothermal Environments)
Abstract
:At deep-sea hydrothermal vents, sulfur oxidation and iron oxidation are of the highest importance to microbial metabolisms, which are thought to contribute mainly in chemolithoautotrophic groups. In this study, 17 mixotrophic neutrophilic thiosulfate- and iron-oxidizing bacteria were isolated from hydrothermal fields on the Carlsberg Ridge in the Indian Ocean, nine to the γ-proteobacteria (Halomonas (4), Pseudomonas (2), Marinobacter (2), and Rheinheimera (1)), seven to the α-proteobacteria (Thalassospira, Qipengyuania, Salipiger, Seohaeicola, Martelella, Citromicrobium, and Aurantimonas), and one to the Actinobacteria (Agromyces), as determined by their 16S rRNA and genome sequences. The physiological characterization of these isolates revealed wide versatility in electron donors (Fe(II) and Mn(II), or thiosulfate) and a variety of lifestyles as lithotrophic or heterotrophic, microaerobic, or anaerobic. As a representative strain, Pseudomonas sp. IOP_13 showed its autotrophic gowth from 105 cells/ml to 107 cells/ml;carbon dioxide fixation capacity with the δ13CVPDB in the biomass increased from −27.42‰ to 3460.06‰; the thiosulfate-oxidizing ability with produced SO42− increased from 60 mg/L to 287 mg/L; and the iron-oxidizing ability with Fe(II) decreased from 10 mM to 5.2 mM. In addition, iron-oxide crust formed outside the cells. Gene coding for energy metabolism involved in possible iron, manganese, and sulfur oxidation, and denitrification was identified by their genome analysis. This study sheds light on the function of the mixotrophic microbial community in the iron/manganese/sulfur cycles and the carbon fixation of the hydrothermal fields.
1. Introduction
Iron and sulfur are elements widely present in the earth. The sources of iron and sulfur in the ocean include dust, coastal and shallow sediments, sea ice, and hydrothermal fluids [1]. Due to the perennial lack of light, the hydrothermal fluid ejected from the deep-sea hydrothermal vent contains a large amount of low-valent iron, manganese, reducing sulfide, methane, hydrogen, and other reducing compounds, which can be used as an electron donor for chemoautotrophic bacteria [2]. Additionally, heterotrophic γ-proteobacteria and α-proteobacteria were occasionally reported to have lithotrophic iron-, manganese-, and sulfur-oxidation capacities in some environments. Edwards et al. first reported that strains belonging to γ-proteobacteria (Alcanivorax sp., Halomonas sp., Marinobacter sp., and Pseudomonas sp.), α-proteobacteria (Aurantimonas sp., Nitratireductor sp. Stappia sp., and Hyphomonas sp.), and Actinobacteria (Microbacterium sp.) isolated from the eastern flank of Juan de Fuca Ridge off the coast of the Pacific Northwest were identified to have iron-oxidation and nitrate-reduction functions [3,4]. Representatives of six genera (Metallogenium, Leptothrix, Siderocapsa, Naumaniella, Bacillus, and Pseudomonas) in the phyla of Proteobacteria and firmicutes were also proven to participate in the oxidation of Fe(II) and Mn(II) in the bottom sediments of Lake Baikal in Russia [5]. Elemental iron and sulfur and amorphous iron sulfide enrichments derived from electrode biomass demonstrated products that were indicative of sulfur or iron oxidation by enrichment cultures belonging the genera Halomonas, Idiomarina, Marinobacter, and Pseudomonas of the γ-proteobacteria and Thalassospira and Thioclava from the α-proteobacteria [6]. Heterotrophic γ-proteobacteria (e.g., Pseudoalteromonas sp. and Pseudomonas sp.), isolated from a volcanic seamount located in the eastern-most island of American Samoa, were also reported to have the ability to catalyze ferrous iron oxidation under microaerobic conditions [7]. Despite the reports on the lithotrophic iron/manganese/sulfur-oxidizing ability of those heterotrophic Proteobacteria and firmicutes, little is known about these processes in hydrothermal vents. In this study, we isolated several neutrophilic thiosulfate- and iron-oxidizing bacteria, mainly thought as heterotrophic Proteobacteria, from deep-sea hydrothermal sulfides in Carlsberg Ridge in the Indian Ocean and further characterized their properties and possible molecular mechanisms.
2. Material and Methods
2.1. Sampling, Enrichment and Isolation
The samples were collected in the Wocan and Daxi hydrothermal vents, on the slowly expanding Carlsberg Ridge in the northwestern Indian Ocean on the DY38 cruise by the R/V Xiang Yang Hong 09, from 4 March to 23 March 2017. The crewed submersible Jiao Long collected samples of mussels, sulfides, hydrothermal sediments, and hydrothermal plumes around the active hydrothermal vents through five dives at several sites (6.4° N, 60.5° E; 6.8° N, 60.2° E; 3.7° N, 60.8° E), at a depth of 2928–3454 m (Table 1). Samples were stored in hermetically sealed sterile vials on board and transported at 4 °C to the laboratory. Oxygen- and Fe(II)-opposing gradient tubes were used with zero-valent iron and the released Fe(II) as the sole electron donor. The zero-valent iron plug at the bottom of the gradient tube provided the source of Fe(II). Sodium bicarbonate was used as the sole carbon source in the medium. Agarose-stabilized gradient tube cultures were set up with a zero-valent iron (ZVI) plug (1% w/v high-melt agarose) and semisolid medium (0.15% w/v low-melt agarose) [8]. Per 1000 mL of distilled water, the medium contained 27.50 g NaCl, 0.72 g KCl, 1.40 g CaCl2·2H2O, 5.38 g MgCl2, 6.78 g MgSO4·7H2O, 1.00 g NH4Cl, 10 mM NaHCO3, and 10 mM MES. After autoclaving, the medium was supplemented by a 1 mL/L trace mineral solution (https://www.atcc.org/products/md-tms, accessed on 25 December 2020) and a 1 mL/L vitamin solution (https://www.atcc.org/products/md-vs; www.atcc.org/products/md-tms, accessed on 25 December 2020) and was adjusted to pH 6.2 with CO2. The culture was enriched using agarose-stabilized gradient tubes at 28 °C [8,9]. When a yellow band appeared in the upper layer, a 200 μL medium was collected from the yellow band and then plated on a marine agar 2216 medium (BD Difco, San diego, CA, USA) at 28 °C for further isolation. After repeated plate streaking, the pure culture was obtained. These isolates were deposited in the Marine Culture Collection of China, MCCC (https://mccc.org.cn/, accessed on 25 December 2020) (Table 1).
2.2. Determination of Iron-Oxidation Capacity
The growths of isolated strains with a solid electron donor in the gradient tubes were tested. The varieties of electron donors include zero-valent iron, FeS, FeCO3, basalt, and iron-manganese nodules. FeCO3 and FeS were prepared as described by Hallbeck et al. [10] and Hanert et al. [11]. All reagents were chemically pure (CP) grade unless otherwise indicated. The basalt, pyrite, and iron-manganese nodules used are natural minerals from deep-sea environments, which were ground and sterilized for later use. The semisolid media with different electron donors were prepared separately. The strains were inoculated into the medium and cultured at 28 °C for up to 2 months.
Aerobic and anaerobic growths were also tested in aqueous media with oxygen or nitrate as electron acceptors. The preparation of stock solutions of FeCl2 was prepared as described by Emerson et al. [8]. The artificial seawater (ASW) in the iron oxidation-nitrate-reduction medium included 28.13 g NaCl, 0.77 g KCl, 1.60 g CaCl2·2H2O, 4.80 g MgCl2·6H2O, 0.11 g NaHCO3, 3.50 g MgSO4·7H2O, 0.001 g resazurin, and 1000 mL distilled water. The medium included 5 mM nitrate as the electron acceptor and was supplemented by 1/10,000 of yeast extract (Oxoid, Basingstoke, UK). Serum bottles with 50 mL volume were filled anoxically with 20 mL medium and were sealed with butyl rubber stoppers. The liquid in each serum bottle is gassed with a filter-sterilized N2:CO2 (80:20 v/v) gas mix for a minimum of 3 min. After autoclaving, the medium was supplemented by 10 mM NaHCO3, 1 mL/L Wolfe’s trace mineral solution, 1 mL/L vitamin solution, and 10 mM FeCl2·4H2O. The supplemented solution was sterilized by filtration through 0.22 μm Millipore filters. The anaerobic iron-oxidizing medium contained nitrate as the electron acceptor and FeCl2·4H2O as the energy source and reducing agents. The concentrations of nitrite produced divalent iron, and the total iron in the supernatant and that in the precipitate in the medium were measured.
2.3. Determination of Manganese-Oxidation Capacity
The growth in the K medium with 1 mM MnCl2 was added to test the ability of manganese oxidation. Per 1000 mL of distilled water, the medium contained 27.5 g NaCl, 0.05 g K2HPO4, 1.40 g CaCl2·2H2O, 1.00 g NH4Cl, 5.38 g MgCl2, 0.72 g KCl, 6.78 g MgSO4·7H2O, 2.0 g Peptone, 0.5 g yeast extract, and 4.766 g HEPES, and it was adjusted to pH 7.6 with NaOH. After autoclaving, the medium was supplemented by 1 mL sterilized MnCl2 (1 M) [12]. The strains were inoculated into K agar medium and cultivated for up to 20 days at 28 °C. The presence of Mn oxides around colonies was confirmed by using the leucoberbelin blue (LBB) assay [13].
2.4. Determination of Thiosulfate-Oxidizing Capacity
Growth in the autotrophic sulfur-oxidizing medium with sodium thiosulfate as the sole electron donor and sodium bicarbonate as the sole carbon source was observed, which was modified according to Lyu et al. [14]. The artificial seawater (ASW) in the SOB medium included 30.00 g NaCl, 0.14 g K2HPO4, 0.14 g CaCl2·2H2O, 0.25 g NH4Cl, 4.18 g MgCl2·6H2O, 0.33 g KCl, 0.005 g NiCl2·6H2O, and 1000 mL distilled water. After autoclaving, the medium was supplemented by sterilized 10 mL trace mineral solution, 10 mL/L vitamin solution, 12.5 mL NaHCO3 (8%), and 10 mL Na2S2O3 (1 M). Before inoculation, the cells were washed three times with sterilized seawater to exclude interference from organic carbon sources and then transferred to the autotrophic media three times. The strains were inoculated into the medium and cultured at 28 °C for 10 days, and the pH, thiosulfate, and sulfate concentrations were measured.
2.5. Determination of Carbon-Fixation Capacity
Carbon-fixation experiments were performed using autotrophic sulfur-oxidation medium with 10 mM NaH13CO3 as sole carbon source and sodium thiosulfate as sole energy source. Before inoculation, the organisms were washed three times with sterilized seawater to exclude interference from organic carbon sources and transferred three times. The strains were inoculated into the medium and cultured at 28 °C for 11 days. The pellets were lyophilized for 24 h and determined by Stable Isotope Ratio Mass Spectrometer (IRMS) (measuring accuracy = ±0.2‰; DeltaVAdvantage; Bremen, Germany). The measured method for δ13C‰ was referred to in Orcutt et al. [15]. The growth curve was determined by measuring the OD value of the culture medium, and the absorbance was tested at 600 nm by using a spectrophotometer (Varioskan LUX, Thermo Scientific, Waltham, MA, USA).
2.6. Measurements of Fe(II), Fe(III), Sulfate, and Nitrate
2.7. Fluorescence Microscopy and Scanning Electron Microscopy/Energy Dispersive X-ray Spectroscopy (SEM/EDS)
The cells were observed using the LIVE/DEAD BacLight Bacterial Viability Kit (Invitrogen, Carlsbad, CA, USA), which contained SYTO 9 and propidium iodide [20], whose excitation/emission wavelengths were respectively set at 480/500 nm and 490/635 nm to perform dual-channel imaging for green and red fluorescence and further counted using a fluorescence microscope (Eclipse 80i, Nikon, Tokyo, Japan). The pellets were gently mounted on a 0.2 µm pore-size polycarbonate filter and air-dried to further examine them by using a FEI/Quorum PP3000T field-emission instrument (Quorum, Laughton, UK) and to analyze the elements by energy dispersive spectroscopy (EDS).
2.8. Phylogenetic Analysis of the Isolated Strains
The genomic DNA of isolated strains was extracted by the bacterial genomic extraction kit (Saibaisheng Co., Beijing, China). The complete 16S rRNA gene was amplified by PCR using bacterial universal primers: 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-ACGGCTACCTTGTTACGACT-3′) [21], and Sanger sequencing was performed. The multiple-sequence alignments were performed by ClustalW (Kyoto university, Kyoto, Japon) [22].The maximum-likelihood tree, based on the small-subunit (16S) rRNA gene, was constructed by IQtree v.1.6.12 (https://www.iqtree.org, accessed on 1 December 2022), with 1000 bootstraps using the “TIM3+F+I+G4” model chosen according to BIC [23]. Sequences of related taxa were obtained from the GenBank database (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genbank/, accessed on 1 August 2022 and EzBioCloud (https://www.ezbiocloud.net/, accessed on 1 August 2022) [24].
2.9. Genomic Analysis of the Isolated Strains
The genome sequences of isolated strains were determined by Shanghai Majorbio Bio-pharm Technology Co., Ltd. (Shanghai, China), using Solexa paired-end (500 bp library) sequencing technology. A total of 1 Gbp clean data with 300 × coverage was generated using the Hiseq 2000 platform (Illumina, San Diego, CA, USA). The clean data were assembled by SPAdes v.3.8.1 (http://cab.spbu.ru/software/spades/, (accessed on 20 January 2021) with the default settings [25]. The contigs longer than 1 kb and similar read coverage were kept for further analysis. SeqPrep (https://github.com/jstjohn/SeqPrep, accessed on 22 January 2021) and Trimmomatic [26] were used to check the Illumina library quality. Check M was used to check the genome quality [27]. Genome annotation was performed using the Rapid Annotation Subsystems Technology (RAST) server [28]. A whole genome BLASTX search was performed against the Kyoto Encyclopedia of Genes and Genomes (KEGG) (https://www.genome.jp/kegg/, accessed on 22 January 2021) [29], Clusters of Orthologous Groups (COG) (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/research/cog/api/cog/, accessed on 22 January 2021) [30], Gene Ontology (GO) (http://geneontology.org/, accessed on 22 January 2021) [31] and Swiss-Prot databases (https://www.uniprot.org/, accessed on 22 January 2021) [32]. Further, rRNAs and tRNAs were predicted using barrnap (https://github.com/tseemann/barrnap, accessed on 10 February 2021) and tRNAscan-SE (http://trna.ucsc.edu/tRNAscan-SE/, accessed on 10 February 2021) [33], respectively. The average nucleotide identity (ANI) was calculated using the ANI calculator tool from EzBioCloud (https://www.ezbiocloud.net/tools/ani, accessed on 22 November 2022) [24]. The amino acid identity (AAI) values between the genomes of isolates and the reference strains of close genera were calculated using a web-based calculator available from (http://enve-omics.ce.gatech.edu/aai/, accessed on 22 November 2022) [34].
2.10. Nucleotide Sequence Accession Numbers for Strains
GenBank accession numbers of the 16S rRNA gene and genome for strains were deposited. They are shown in Table 1.
3. Results
3.1. Phylogenetic Affiliations
Previous studies showed that most Fe-oxidizing bacteria (FeOB) belong to the bacterial phylum Proteobacteria, including the α-, β-, γ-, δ-, and ζ- classes [35]. Figure 1 shows the phylogenetic relationships among 17 strains in this study relative to other known representative FeOB. All the isolates in this study fell mainly within the α- and γ- classes of the Proteobacteria, except for strain IOP_2, belonging to the phylum Actinobacteria. Strains IOP_29 and IOP_41 were affiliated with Marinobacter (M. adhaerens HP15 and M. shengliensis). Some Marinobacter strains were reported to have neutrophilic iron-oxidizing capabilities from the in situ and lab enrichments of basalts, olivine minerals, etc. near hydrothermal vents, the subseafloor, and iron mine [4,6,36,37,38]; sulfur-oxidizing capabilities from marine sediments [37,39]; and Mn(II)-oxidizing capabilities from submarine basalts at Loihi seamount off the southeast coast of the island of Hawaii [40]. Four strains, namely IOP_6, IOP_14, IOP_19, and IOP_31, belong to the genus Halomonas by 16s rRNA gene analysis but with low ANI and AAI values (Supplementary Table S1). Halomonas spp. have been frequently identified as Mn(II)-oxidizing and Fe(II)-oxidizing bacteria from basalt in the Juan de Fuca Ridge flank, the volcanic Loihi seamount [36,40,41], the hydrothermal fields in Juan de Fuca Ridge flank, and [36,42] the acid mine drainage (AMD) environments in Southwest China [43]. Some Halomonas strains isolated from hydrothermal fluids and the sediment of hydrothermal field in the East Pacific Rise also showed sulfur-oxidizing capacity [44]. Strains IOP_13 and IOP_25 belong to genus Pseudomonas, which also has been frequently reported as iron-oxidizing bacteria. As described in Sudek et al. [7], Pseudomonas spp. had iron-oxidizing and Mn(II)-oxidizing capacities under microaerobic conditions from a volcanic seamount in the Juan de Fuca Ridge flank. Pseudomonas sp. FE13-26 extracted from sludge was also reported to efficiently oxide Fe(II) with extracellular enzyme ferroxidase [45]. Pseudomonas sp. LOB-2 was reported to have Mn(II)-oxidizing capacity isolated from submarine basalts at the Loihi seamount [40].
Additionally, seven isolates were affiliated to genera Thalassospira, Qipengyuania, Salipiger, Seohaeicola, Martelella, Citromicrobium, and Aurantimonas in the class α-proteobacteria (Figure 1, Table 2). Most of the iron-oxidizing α-proteobacteria were phototrophs, with the exception of Paracoccus ferrooxidans [46,47], which was a nitrate-dependent bacterium. Moreover, α-proteobacteria (Aurantimonas sp., Nitratireductor sp., Stappia sp., and Hyphomonas sp.) isolated from the eastern flank of Juan de Fuca Ridge were identified to have iron-oxidation and nitrate-reduction functions [4]. The genera Thalassospira and Martelella isolated from hydrothermal sulfides of the South Atlantic were previously also characterized to exhibit a sulfur-oxidizing ability [48]. Those results and our data all proved that some strains in α-proteobacteria had the ability of iron and/or sulfur oxidation to adapt to environmental conditions in deep-sea hydrothermal vents, which had not been notified before.
Strain IOP_2 was closest to Agromyces soli MJ21, with a 94.19% similarity in 16S rRNA genes. The iron-oxidizing ability was reported in organisms of the phylum Actinobacteria: Microbacterium sp. [3] and Ferrimicrobium acidiphilum [49]. The result of this study also showed the possible iron-oxidizing ability of other strains in phylum Actinobacteria.
3.2. Growth Test with Different Electron Donors and Acceptors
The growth tests with different electron donors and acceptors in gradient tubes and liquid medium under the aerobic, microaerobic, and anaerobic conditions for all strains are summarized in Table 2.
3.2.1. Iron-Oxidizing Capacity
All the 17 isolates could oxidize FeS to form an iron-oxide layer in NCMA medium gradient tube (Supplementary Figure S1). Live/dead staining results showed that most of the cells of strain IOP_13, the same as other isolates, were alive in the iron-oxide layer (Supplementary Figure S2). Most isolates (13/17) could oxidize zero-valent iron (Table 2). Ten isolates, including all the Halomonas sp., Pseudomonas sp., Marinobacter sp., Thalassospira sp. IOP_1, and Aurantimonas sp. IOP_38, could oxidize FeCO3. Eight isolates, namely IOP_2, IOP_12, IOP_13, IOP_23, IOP_24, IOP_25, IOP_29, and IOP_38, could oxidize pyrite to iron oxides after 1 month. Notably, Agromyces sp. IOP_2 and Halomonas sp. IOP_6 could grow with basalt as the electron donor in gradient tubes.
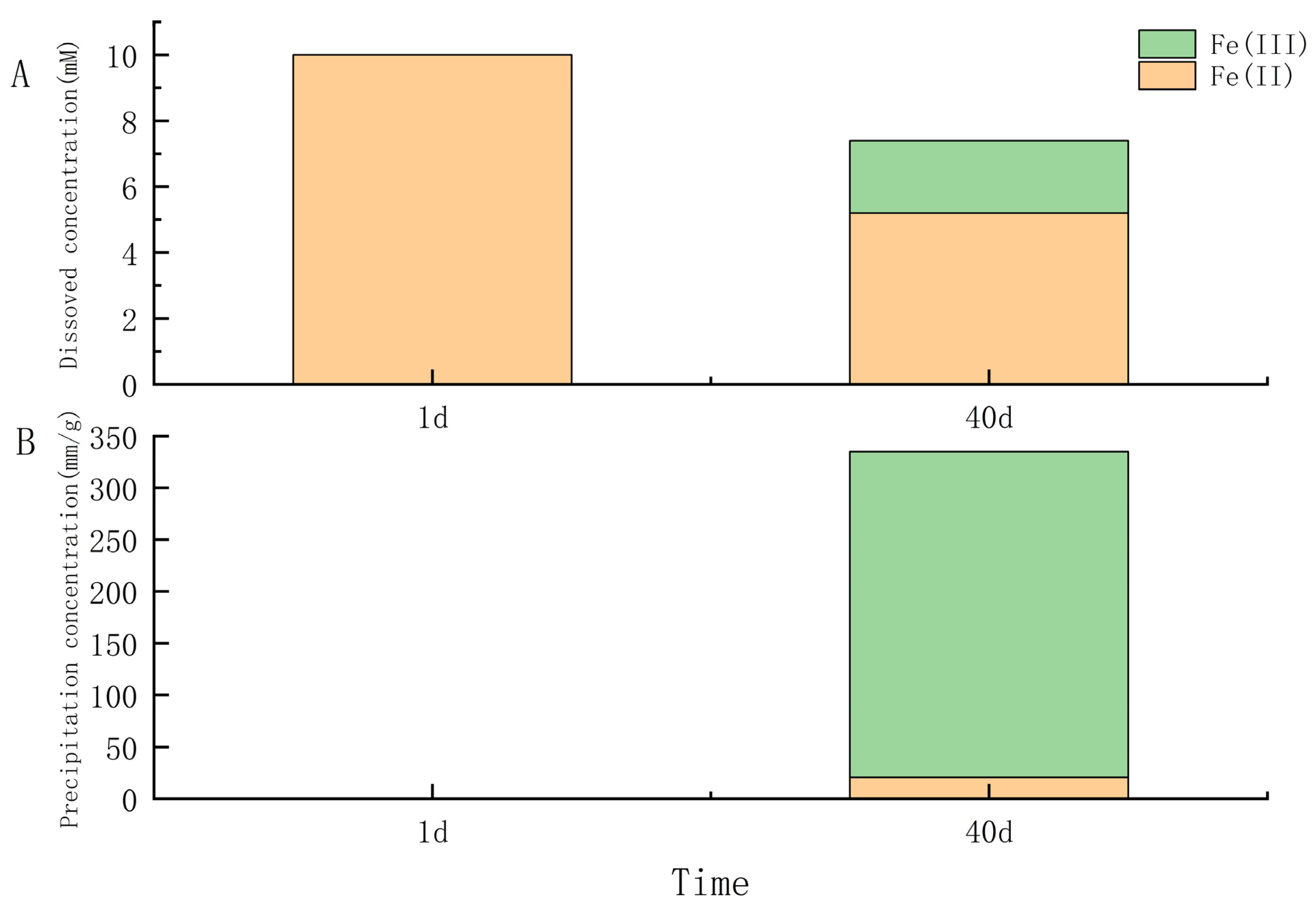
Under anaerobic conditions, all nine γ-proteobacteria and four of seven strains in α-proteobacteria (Salipiger, Seohaeicola, Martelella, and Citromicrobium) could anaerobically grow with oxidizing FeCl2 and reducing nitrate. The growth of the cells, nitrite accumulation, and yellow-green precipitate after 40 days of all strains in anaerobic iron-oxidation media were examined. The results showed that under light and fluorescent microscopy, all the cells were observed to be encrusted with Fe(III) minerals, and some cells became dead with red fluorescent (Figure 2). Some isolates (IOP_6, IOP_14, IOP_19, IOP_31 in genus Halomonas; IOP_13 and IOP_25 in genus Pseudomonas; IOP_29, IOP_41 in genus Marinobacter; IOP_21 in genus Rheinheimera; IOP_16 in genus Salipiger; IOP_23 in genus Seohaeicola; IOP_24 in genus Martelella; and IOP_28 in genus Citromicrobium) produced yellow trivalent iron-oxide precipitates in the bottles during the process of cultivation. Nitrite is usually produced an intermediate product produced by bacteria using nitrate as an electron acceptor for anaerobic respiration. The accumulation of nitrite in those cultures was determined to be significantly higher (12.15 to 82.03 μM) than that in the negative control. This demonstrated that these strains use nitrate as an electron acceptor for anaerobic respiration. The nitrate reduction, nitrite formation, and Fe(II) oxidation were not observed in the noninoculated control medium. Strain Pseudomonas sp. IOP_13 produced significantly more nitrite than other strains and produced an orange trivalent iron-oxide precipitate after 10 days of growth (Supplementary Figure S3). The microscopy and EDS analysis of the strain Pseudomonas sp. IOP_13 showed that the cells were encrusted with minerals containing iron (Figure 2). The aqueous Fe(II) concentration in the culture decreased from 10 mM to 5.20 mM with dissolved Fe(III), which ranged from 0 mM to 2.2 mM after 40 days of growth (Figure 3A). The orange iron-oxide precipitate contained absorbed 20.63 mM/g Fe(II) and 314.58 mM/g Fe(III) (Figure 3B).
3.2.2. Manganese-Oxidizing Capacity
Manganese-oxidizing bacteria can oxidize manganese chloride to manganese dioxide and produce dark-brown colonies on manganese-oxidizing plates. All isolates were cultured in a manganese-oxidation medium, and LBB assay was used on colonies cultured for 20 days. The redox stain (LBB) turned blue in the presence of the accumulation and conversion of manganese by strains Pseudomonas spp. IOP_13, IOP_25, and Salipiger sp. IOP_16.
Strain IOP_16 showed the capacity for Mn(II) oxidation; the closest organism to this strain was the manganese-oxidizing bacterium Salipiger manganoxidans VSW210, isolated from a shallow-water hydrothermal vent in Espalamaca (Azores) [50]. Two strains in the genus Pseudomonas (IOP_13 and IOP_25) that were capable of oxidizing Mn(II) were also identified. Supplementary Figure S4B showed the formation of a blue color on the colonies of strain IOP_13 after LBB staining. Orange halos around the colonies showed its siderphore-producing ability (Supplementary Figure S4A). Some Pseudomonas spp. were reported to have a potential of Mn(II)-oxidizing capacity. Kepkay and Nealson (1987) reported the occurrence of the growth of marine bacteria Pseudomonas sp. S-36, both mixotrophically on succinate or bicarbonate with Mn(II), in Mn-limited chemostats [51]. Mn(II)-oxidizing bacterium Pseudomonas putida GB-1 from a freshwater environment was also identified [52,53,54,55].
3.2.3. Thiosulfate-Oxidizing Capacity
The growth and SO42− production of all strains in a thiosulfate oxidation medium, which contained an inorganic carbon source and sodium thiosulfate as an energy source, were examined. Halomonas spp. IOP_6, IOP_14, IOP_19, and IOP_31; Pseudomonas spp. IOP_13 and IOP_25; Marinobacter spp. IOP_29 and IOP_41 in γ-proteobacteria and Thalassospira sp. IOP_1; Citromirobium sp. IOP_28; and Aurantimonas sp. IOP_38 in α-proteobacteria showed an autotrophic sulfur-oxidizing capacity with monomeric sulfur and/or sulfate produced.
The growth curves and the concentrations of electron donor S2O32− and product SO42− of strains Pseudomonas sp. IOP_13 and Halomonas sp. IOP_14 were further measured, as shown in Figure 4A,B. The results showed that the cell concentrations increased from the initial 105 cells/mL to the final 107 cells/mL after 8 days. The electron donor S2O32− was consistently consumed by strains IOP_13 and IOP_14 after inoculation and produced SO42− (287 mg/L, 394 mg/L) without SO32− detected. The pH value of the culture increased during the first 3 days (0.2 units and 0.5 units) and then stabilized (Figure 4C). This indicated that these strains produced alkaline substances in the process of sulfur oxidation.
3.3. Carbon Dioxide–Fixation Capacity
All the strains were cultured in a sulfur-containing medium that used NaHCO3 as the sole carbon source. The results showed that 12/17 strains exhibited autotrophic growth. Strain IOP_13 was chosen to culture with NaH13CO3 as the sole carbon source to test its autotrophic capacity. The growth curve of strain IOP_13 and the value of δ13C in its biomass are shown in Figure 5. The results showed that bacterial cells increased from 105 to 107 cells/mL and that the δ13CVPDB value of the bacterium increased from −27.42‰ to 3460.06‰ within 11 days, which indicated that strain IOP_13 utilized inorganic carbon as a carbon source.
3.4. Genome Characteristics of Isolated Strains
The 17 isolates were sequenced with 300 × coverage, and the draft genomes were produced with 98.69%–100.00% completeness (Supplementary Table S2). The genome similarities between isolates and closest relatives were also analyzed, with 76.50%–100.00% ANI values and 65.88%–99.95% AAI values (Supplementary Table S1).
An analysis of the metabolic reconstruction using the KEGG database suggested that most of them have complete glycolysis, Entner–Doudoroff, pentosephosphate, and tricarboxylic acid cycle pathways, which showed their heterotrophic traits. The results for the functional gene annotation of strains are shown in Figure 6. Five strains, including Halomonas spp. IOP_6, IOP_14, and IOP_19 and Pseudomonas spp. IOP_13 and IOP_25, had high similarity with the sulfur-oxidation gene TsdA (Supplementary Figure S5). Tetrathionate-forming thiosulfate dehydrogenase (TsdA), identified by [56] from the purple sulfur bacterium Allochromatium vinosum, and this protein play the role of sulfur-based energy metabolism through the oxidation of thiosulfate. These isolates with the TsdA gene also showed a sulfur-oxidation ability, which could be carried out by the tetrasulphate pathway. Aurantimonas sp. IOP_38 and Seohaeicola sp. IOP_23 were annotated with the complete Sox sulfur-oxidation gene (SoxABCDHSFRSWXYZ). Strain IOP_38 could grow in an autotrophic sulfur-containing medium, but no sulfate or monosulfate production was detected. Seohaeicola sp. IOP_23 could grow only in a hetertrophic sulfur-containing medium.
The genes-encoding multicopper oxidases CumA [57] (Supplementary Figure S6) with the possible involvement in Mn(II) oxidation were found in strains IOP_13, IOP_16 and IOP_25, which have the capacity for Mn(II) oxidation.
Proteins thought to be involved in Fe(II) oxidation, namely Cyc1 [58,59], Cyc2 [58,59,60], FoxABC [61], FoxEYZ [62], Sulfocyanin [63], MtoAB [64], and PioABC [65], were queried against the genomes. A Fegenie analysis [66] showed that there were no known iron-oxidizing genes found in those isolates, in this study. The genomes were further searched for proteins containing the characteristic c-type cytochrome binding CXXCH motif [67]. Cytochromes c require the covalent attachment of heme via two thioether bonds at conserved CXXCH motifs, a process accomplished in prokaryotes by eight integral membrane proteins (CcmABCDEFGH). Heme is trafficked from inside the cell to outside (via CcmABCD) and chaperoned (holoCcmE) to the cytochrome c synthetase (CcmF/H) [68]. The results showed that 15 strains have possible c-type cytochromes maturation in systems I, except Halomonas sp. IOP_31 and Agromyces sp. IOP_2 (Figure 6). Strain IOP_2 contained CcsA, which belongs to the systems II pathway [67].
Some strains can couple iron oxidation with nitrate reduction. Denitrification-related genes were analyzed. Genes encoding the NarKGHJI, NirSKV, NorDQBCFE, and NosXLYFDZR proteins were annotated in 10 strains grown with iron oxidation and nitrate reduction. Enzymes NarGHI (nitrate reductase A) identified from E. coli [69] play the role of denitrification and support anaerobic growth on nitrate with a nonfermentable carbon source condition, and the genes encoding NarGHI were found in strains IOP_6, IOP_13, IOP_14, IOP_19, IOP_21, IOP_24, IOP_25, IOP_29, IOP_31, and IOP_41 (Supplementary Figure S7), but not IOP_38.
4. Discussion
4.1. Widespread of Mixotrophic Bacterial Strains in the Hydrothermal Vents
There is rich heterotrophic microbial life in hydrothermal vents. Versatile heterotrophic α- and γ-proteobacteria have been found in different venting areas of the Menez Gwen hydrothermal field in the Mid-Atlantic Ridge from the diffuse fluid discharge points through the mixing gradients to the plumes and the surrounding seawater [70]. Generalist species belonging to the genera Marinobacter, Vibrio, Pseudoalteromonas, Halomonas, Pseudomonas, and Alcanivorax, among others, have been repeatedly isolated from hydrothermal vent samples in the Pacific [4,71,72,73,74], and their abundance in vent fluids collected from the Pacific Ocean was estimated to be up to 28% of the total micro-organisms [75]. Phylotypes closely related to cultured species, e.g., Alteromonas, Halomonas, and Marinobacter, were relatively abundant in some crustal fluid samples in the Suiyo seamount off the eastern coast of Japan [76]. In this study, those heterotrophic α- and γ-proteobacteria were isolated from mussels, sulfides, hydrothermal sediments, and hydrothermal plumes around active hydrothermal vents (Table 1), which also indicated that they are widespread in these types of environments.
Compared with other Proteobacterial classes, ζ-proteobacterial populations as iron oxidizers have a narrow growth range and spread only in oxic-anoxic transition zones near shore environments [77] and iron-rich hydrothermal systems, such as iron-oxide material, hydrothermal sediment, etc. [78,79]. Until now, only seven species in two genera, Mariprofundus [80] and Ghiorsea [81], had been isolated, which have a narrow growth range of oxygen concentration in 0.07–2.0 μM [77]. Heterotrophic α- and γ-proteobacteria shown across broad environmental gradients and dominate in the hydrothermal plumes, sulfides, and sediments might have important roles in the element cycle of iron and sulfur, which have been under estimation before.
4.2. Diverse Metabolism of Mixotrophic Bacterial Strains in the Hydrothermal Vents
Theoretical studies demonstrate that mixotrophy is advantageous in oligotrophic or fluctuating environments [82]. Micro-organisms survive using an adaptation ability for effective competition in the hydrothermal ecosystem with shifting biogeochemical conditions [83,84]. For example, archaea Pyrobaculum islandicum from hydrothermal vents can live as heterotrophs or autotrophs strictly under piezophilic conditions. The SUP05 clade of gammaproteobacteria (Thioglobaceae) could act as abundant autotrophs in the hydrothermal fluids and in association with eukaryotes at hydrothermal vents or as heterotrophes throughout the ocean [85]. The heterotrophic genera Pseudomonas, Halomonas, and Bacillus were all reported to possess the tetrathionate-forming ability using thiosulfate as a supplemental inorganic energy source [86,87]. Many strains in the Pseudomonas genus, such as strains of Pseudomonas stutzeri, were shown to oxidize sodium sulfide, change thiosulfate into tetrathionate, or grow on a variety of substrates, such as FeCl2, FeCO3 and FeSO4, as their sole energy source under anaerobically in the presence of nitrate [88,89,90]. We further predicted the sulfur-oxidizing and denitrification genes in 317 genomes belonging Pseudomonas in UniProt (http://sparql.uniprot.org/ accessed on 10 November 2022). It showed that 10 species, including P. stutzeri strain A1501, P. aeruginosa, P. veronii, P. wadenswilerensis, P. reidholzensis, P. fluorescens, P. extremaustralis, P. marincola, Pseudomonas sp. IsoF, and Pseudomonas sp. 9Ag-encoded TsdAB genes and NarI, were detected in 115 Pseudomonas genomes. It indicated that some Pseudomonas spp. have thiosulfate-oxidizing and nitrate-reducing potential for mixotrophic life. Some micro-organisms have been characterized as heterotrophes, but their autotrophic ability was not shown before. This study revealed their versatile metabolism as autotrophes and heterotrophes with sulfur and iron oxidization in aerobic-microaerobic-anaerobic conditions, and it further predicted possible mechanisms of these processes based on the annotation of their genomes. Sulfur oxidation with the tetrasulphate pathway by thiosulfate dehydrogenase was found common in Halomonas and Pseudomonas spp. of γ-proteobacteria, in this study. The thiosulfate-oxidizing enzyme (Sox) was found only in two isolates (Seohaeicola sp. IOP_23 and Aurantimonas sp. IOP_38) of α-proteobacteria. Almost all of them had genes for a complete pathway of denitrification. A total of 16 strains had c-type cytochromes maturation systems and c-type cytochromes with one or more heme-binding motifs, which may have a function in Fe redox reactions, as reported for iron oxidizers (Rhodobacter and Sideroxydans spp.) [62,64]. It is also noteworthy that Agromyces sp. IOP_2 in the phylum Actinobacteria with iron- and sulfur-oxidizing abilities lack all known iron- and sulfur-oxidizing genes, which needs to be further investigated.
However, it remains unknown how mixotrophic micro-organisms respond to different conditions across broad environmental gradients. The characterization of mixtrophs and their metabolisms deserve further attention.
5. Conclusions
With a rich variety of chemical energy sources and steep physical and chemical gradients, hydrothermal vent systems offer a range of habitats to support microbial life.
In this study, we isolated iron-oxidizing and thiosulfate-oxidizing bacteria from deep-sea hydrothermal vents and shed light on the potential diverse functions of these heterotrophic bacteria in γ- and α-proteobacteria and Actinobacteria. Based on their broad growth ranges and versatile metabolisms, we predicted that heterotrophic bacteria which have capacities of sulfur, iron, and manganese oxidation may play important roles in carbon, sulfur, and metal cycling in hydrothermal vents.
Supplementary Materials
The following supporting information can be downloaded at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/microorganisms11010100/s1, Figure S1: Gradient tube (ZVI-based) growth of FeOB; Figure S2: Microscopic photo of bacterial cells from the iron-oxides layer in the gradient tube; Figure S3: Orange trivalent iron precipitate after 10 days growth for strain Pseudomonas sp. IOP_13 in nitrate-reduction and iron-oxidation medium; Figure S4: Colonies of strain Pseudomonas sp. IOP_13 growing on solid test media; Figure S5: Sequence alignment of TsdA proteins; Figure S6: Sequence alignment of CumA proteins in the isolates and CumA identified from Pseudomonas putida GB-1; Figure S7: Sequence alignment of Nar I proteins in the isolates and Nar I (respiratory nitrate reductase 1 gamma chain) identified from E. coli; Table S1: Average nucleotide identity (ANI) and average amino acid identity (AAI) between the genomes of isolates and their relatives; Table S2: Genomic characteristics of isolated strains.
Author Contributions
X.Z. and Z.S. designed the research outline. X.Z. and Y.H. drafted the manuscript. Y.H., X.Z. and F.X. performed the experiments. Z.S. collected the sample from the Indian Ocean. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Key R&D Program of China (No. 2021YFF0501304), the National Natural Science Foundation of China (No.91951201 and No.42030412), the Scientific Research Foundation of Third Institute of Oceanography, MNR (2019021), the China Ocean Mineral Resources R&D Association (COMRA) program (No. DY135-B2-01) and the National Microbial Resource Center (No. NMRC-2022-9).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to the R/V Xiang Yang Hong 09 and HOV Jiao Long operation teams for helping us to collect the samples from the deep-sea hydrothermal vent field.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boyd, P.W.; Ellwood, M.J. The biogeochemical cycle of iron in the ocean. Nat. Geosci. 2010, 3, 675. [Google Scholar] [CrossRef]
- Jannasch, H.W. The Chemosynthetic support of life and the microbial diversity at Deep-Sea hydrothermal vents. Proc. R. Soc. Lond. Ser. B 1985, 225, 277. [Google Scholar]
- Edwards, K.J.; Bach, W.; McCollom, T.M.; Rogers, D.R. Neutrophilic iron-oxidizing bacteria in the ocean: Their habitats, diversity, and roles in mineral deposition, rock alteration, and biomass production in the deep-sea. Geomicrobiol. J. 2004, 21, 393–404. [Google Scholar] [CrossRef]
- Edwards, K.J.; Rogers, D.R.; Wirsen, C.O.; McCollom, T. Isolation and characterization of novel psychrophilic, neutrophilic, Fe-oxidizing, chemolithoautotrophic α-and γ-Proteobacteria from the deep sea. Appl. Environ. Microbiol. 2003, 69, 2906–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharova, Y.R.; Parfenova, V.V.; Granina, L.Z.; Kravchenko, O.S.; Zemskaya, T.I. Distribution of iron- and manganese-oxidizing bacteria in the bottom sediments of Lake Baikal. Inland Water Biol. 2010, 3, 313–321. [Google Scholar] [CrossRef]
- Rowe, A.R.; Chellamuthu, P.; Lam, B.; Okamoto, A.; Nealson, K.H. Marine sediments microbes capable of electrode oxidation as a surrogate for lithotrophic insoluble substrate metabolism. Front. Microbiol. 2015, 5, 784. [Google Scholar] [CrossRef] [Green Version]
- Sudek, L.A.; Templeton, A.S.; Tebo, B.M.; Staudigel, H. Microbial ecology of Fe (hydr) oxide mats and basaltic rock from Vailulu’u Seamount, American Samoa. Geomicrobiol. J. 2009, 26, 581–596. [Google Scholar] [CrossRef]
- Emerson, D.; Floyd, M.M. Enrichment and isolation of iron-oxidizing bacteria at neutral pH. Methods Enzymol. 2005, 397, 112–123. [Google Scholar]
- Emerson, D.; Rentz, J.A.; Lilburn, G.T.; Davis, R.E.; Aldrich, H.; Chan, C.; Moyer, C.L. A novel lineage of Proteobacteria involved in formation of marine fe-oxidizing microbial mat communities. PLoS ONE 2007, 2, e667. [Google Scholar] [CrossRef] [Green Version]
- Hallbe, L.; Stahl, F.; Pedersen, K. Phylogeny and phenotypic characterization of the stalk-forming and iron-oxidizing bacterium Gallionella ferruginea. J. Gen. Microbiol. 1993, 139, 1531–1539. [Google Scholar] [CrossRef]
- Hanert, H.H. The Genus Gallionella. In The Prokaryotes: Volume 7: Proteobacteria: Delta, Epsilon Subclass; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 990–995. [Google Scholar]
- Woods, D.; Sokol, P. The Prokaryotes, Vol. 5. Proteobacteria: Alpha and Beta Subclasses; Springer: New York, NY, USA, 2006. [Google Scholar]
- Krumbein, W.; Altmann, H. A new method for the detection and enumeration of manganese oxidizing and reducing microorganisms. Helgoländer Wiss. Meeresunters. 1973, 25, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Lyu, J.; Shao, Z. Sulfur metabolism of Hydrogenovibrio thermophilus strain S5 and its adaptations to deep-sea hydrothermal vent environment. Front. Microbiol. 2017, 8, 2513. [Google Scholar] [CrossRef] [PubMed]
- Orcutt, B.N.; Sylvan, J.B.; Rogers, D.R.; Delaney, J.; Lee, R.W.; Girguis, P.R. Carbon fixation by basalt-hosted microbial communities. Front. Microbiol. 2015, 6, 904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stookey, L.L. Ferrozine—A new spectrophotometric reagent for Iron. Anal. Chem. 1970, 42, 779–781. [Google Scholar] [CrossRef] [Green Version]
- Josephson, B. A volumetric method for the determination of sulphur and sulphate ion. Analyst 1939, 64, 181–185. [Google Scholar] [CrossRef]
- Small, H.; Stevens, T.S.; Bauman, W.C. Novel ion exchange chromatographic method using conductimetric detection. Anal. Chem. 1975, 47, 1801–1809. [Google Scholar] [CrossRef]
- Cox, R.D. Determination of nitrate and nitrite at the parts per billion level by chemiluminescence. Anal. Chem. 1980, 52, 332–335. [Google Scholar] [CrossRef]
- Berney, M.; Hammes, F.; Bosshard, F.; Weilenmann, H.-U.; Egli, T. Assessment and interpretation of bacterial viability by using the LIVE/DEAD BacLight Kit in combination with flow cytometry. Appl. Environ. Microbiol. 2007, 73, 3283–3290. [Google Scholar] [CrossRef] [Green Version]
- Hiraishi, A. Direct automated sequencing of 16s rDNA amplified by polymerase chain reaction from bacterial cultures without DNA purification. Lett. Appl. Microbiol. 1992, 15, 210–213. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki Kinoshita, K.F.; Itoh, M.; Kawashima, S.; Katayama, T.; Araki, M.; Hirakawa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, 354–357. [Google Scholar] [CrossRef]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Krylov, D.M.; Makarova, K.S.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; Rao, B.S.; et al. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome Biol. 2004, 5, R7. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Bairoch, A.; Apweiler, R. The SWISS-PROT protein sequence database and its supplement TrEMBL in 2000. Nucleic Acids Res. 2000, 28, 45–48. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA Genes in Genomic Sequences. In Methods in Molecular Biology; Humana: New York, NY, USA, 2019; Volume 1962, pp. 1–14. [Google Scholar]
- Rodriguez, R.L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ Prepr. 2016, 4, e1900v1. [Google Scholar]
- Makita, H. Iron-oxidizing bacteria in marine environments: Recent progresses and future directions. World J. Microbiol. Biotechnol. 2018, 34, 110. [Google Scholar] [CrossRef]
- Smith, A.; Popa, R.; Fisk, M.; Nielsen, M.; Wheat, C.G.; Jannasch, H.W.; Fisher, A.T.; Becker, K.; Sievert, S.M.; Flores, G. In situ enrichment of ocean crust microbes on igneous minerals and glasses using an osmotic flow-through device. Geochem. Geophys. Geosyst. 2011, 12, Q06007. [Google Scholar] [CrossRef]
- Zhang, X.; Fang, J.; Bach, W.; Edwards, K.J.; Orcutt, B.N.; Wang, F. Nitrogen stimulates the growth of subsurface basalt-associated microorganisms at the western flank of the Mid-Atlantic Ridge. Front. Microbiol. 2016, 7, 633. [Google Scholar] [CrossRef] [Green Version]
- Bonis, B.M.; Gralnick, J.A. Marinobacter subterrani, a genetically tractable neutrophilic Fe (II)-oxidizing strain isolated from the Soudan Iron Mine. Front. Microbiol. 2015, 6, 719. [Google Scholar] [CrossRef]
- Choi, B.-R.; Pham, V.H.; Park, S.-J.; Kim, S.-J.; Roh, D.-H.; Rhee, S.-K. Characterization of facultative sulfur-oxidizing Marinobacter sp. BR13 isolated from marine sediment of Yellow Sea, Korea. J. Korean Soc. Appl. Biol. Chem. 2009, 52, 309–314. [Google Scholar] [CrossRef]
- Templeton, A.S.; Staudigel, H.; Tebo, B.M. Diverse Mn (II)-oxidizing bacteria isolated from submarine basalts at Loihi Seamount. Geomicrobiol. J. 2005, 22, 127–139. [Google Scholar] [CrossRef]
- Homann, V.V.; Sandy, M.; Tincu, J.A.; Templeton, A.S.; Tebo, B.M.; Butler, A. Loihichelins A-F, a suite of amphiphilic siderophores produced by the marine bacterium Halomonas LOB-5. J. Nat. Prod. 2009, 72, 884–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaye, J.Z.; Sylvan, J.B.; Edwards, K.J.; Baross, J.A. Halomonas and Marinobacter ecotypes from hydrothermal vent, subseafloor and deep-sea environments. FEMS Microbiol. Ecol. 2011, 75, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Xiao, E.; Krumins, V.; Dong, Y.; Xiao, T.; Ning, Z.; Chen, H.; Xiao, Q. Characterization of the microbial community composition and the distribution of Fe-metabolizing bacteria in a creek contaminated by acid mine drainage. Appl. Microbiol. Biotechnol. 2016, 100, 8523–8535. [Google Scholar] [CrossRef]
- Podgorsek, L.; Petri, R.; Imhoff, J.F. Cultured and genetic diversity, and activities of sulfur-oxidizing bacteria in low-temperature hydrothermal fluids of the North Fiji Basin. Marine Ecol. Prog. Ser. 2004, 266, 65–76. [Google Scholar] [CrossRef]
- Xu, X.; Yang, G.; Wang, F.; Li, M. Ferro-oxidase producing conditions of Pseudomonas sp. J. Microbiol. 2009, 29, 16–19. [Google Scholar]
- Hedrich, S.; Schlömann, M.; Johnson, D.B. The iron-oxidizing proteobacteria. Microbiology 2011, 157, 1551–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blöthe, M.; Roden, E.E. Composition and activity of an autotrophic Fe (II)-oxidizing, nitrate-reducing enrichment culture. Appl. Environ. Microbiol. 2009, 75, 6937–6940. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Jiang, L.; Li, S.; Zhong, T.; Lai, Q.; Shao, Z. Diversity of culturable sulfur-oxidizing bacteria in deep-sea hydrothermal vent environments of the South Atlantic. Wei Sheng Wu Xue Bao Acta Microbiol. Sin. 2016, 56, 88–100. [Google Scholar]
- Johnson, D.B.; Bacelar-Nicolau, P.; Okibe, N.; Thomas, A.; Hallberg, K.B. Ferrimicrobium acidiphilum gen. nov., sp. nov. and Ferrithrix thermotolerans gen. nov., sp. nov.: Heterotrophic, iron-oxidizing, extremely acidophilic actinobacteria. Int. J. Syst. Evol. Microbiol. 2009, 59, 1082–1089. [Google Scholar] [CrossRef]
- Rajasabapathy, R.; Mohandass, C.; Dastager, S.G.; Liu, Q.; Li, W.-J.; Colaço, A. Citreicella manganoxidans sp. nov., a novel manganese oxidizing bacterium isolated from a shallow water hydrothermal vent in Espalamaca (Azores). Antonie Leeuwenhoek 2015, 108, 1433–1439. [Google Scholar] [CrossRef]
- Kepkay, P.; Nealson, K. Growth of a manganese oxidizing Pseudomonas sp. in continuous culture. Arch. Microbiol. 1987, 148, 63–67. [Google Scholar] [CrossRef]
- Okazaki, M.; Sugita, T.; Shimizu, M.; Ohode, Y.; Iwamoto, K.; de Vrind-de Jong, E.W.; de Vrind, J.P.; Corstjens, P.L. Partial purification and characterization of manganese-oxidizing factors of Pseudomonas fluorescens GB-1. Appl. Environ. Microbiol. 1997, 63, 4793–4799. [Google Scholar] [CrossRef] [Green Version]
- Brouwers, G.-J.; de Vrind, J.P.; Corstjens, P.L.; Cornelis, P.; Baysse, C.; de Vrind-de Jong, E.W. cumA, a gene encoding a multicopper oxidase, is involved in Mn2+ oxidation in Pseudomonas putida GB-1. Appl. Environ. Microbiol. 1999, 65, 1762–1768. [Google Scholar] [CrossRef] [Green Version]
- Murray, K.J.; Mozafarzadeh, M.L.; Tebo, B.M. Cr (III) oxidation and Cr toxicity in cultures of the manganese (II)-oxidizing Pseudomonas putida strain GB-1. Geomicrobiol. J. 2005, 22, 151–159. [Google Scholar] [CrossRef]
- Toner, B.; Fakra, S.; Villalobos, M.; Warwick, T.; Sposito, G. Spatially resolved characterization of biogenic manganese oxide production within a bacterial biofilm. Appl. Environ. Microbiol. 2005, 71, 1300–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denkmann, K.; Grein, F.; Zigann, R.; Siemen, A.; Bergmann, J.; van Helmont, S.; Nicolai, A.; Pereira, I.A.; Dahl, C. Thiosulfate dehydrogenase: A widespread unusual acidophilic c-type cytochrome. Environ. Microbiol. 2012, 14, 2673–2688. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Long, Y.; Yan, H.; Cai, H.; Li, Y.; Wang, X. Gene cloning, identification, and characterization of the multicopper oxidase CumA from Pseudomonas sp. 593. Biotechnol. Appl. Biochem. 2017, 64, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Castelle, C.; Guiral, M.; Malarte, G.; Ledgham, F.; Leroy, G.; Brugna, M.; Giudici-Orticoni, M.-T. A new iron-oxidizing/O2-reducing supercomplex spanning both inner and outer membranes, isolated from the extreme acidophile Acidithiobacillus ferrooxidans. J. Biol. Chem. 2008, 283, 25803–25811. [Google Scholar] [CrossRef]
- Barco, R.A.; Emerson, D.; Sylvan, J.B.; Orcutt, B.N.; Jacobson Meyers, M.E.; Ramírez, G.A.; Zhong, J.D.; Edwards, K.J. New insight into microbial iron oxidation as revealed by the proteomic profile of an obligate iron-oxidizing chemolithoautotroph. Appl. Environ. Microbiol. 2015, 81, 5927–5937. [Google Scholar] [CrossRef] [Green Version]
- McAllister, S.M.; Polson, S.W.; Butterfield, D.A.; Glazer, B.T.; Sylvan, J.B.; Chan, C.S. Validating the Cyc2 neutrophilic Fe oxidation pathway using meta-omics of Zetaproteobacteria iron mats at marine hydrothermal vents. bioRxiv 2019. bioRxiv:722066. [Google Scholar]
- Bathe, S.; Norris, P.R. Ferrous iron-and sulfur-induced genes in Sulfolobus metallicus. Appl. Environ. Microbiol. 2007, 73, 2491–2497. [Google Scholar] [CrossRef] [Green Version]
- Croal, L.R.; Jiao, Y.; Newman, D.K. The fox operon from Rhodobacter strain SW2 promotes phototrophic Fe (II) oxidation in Rhodobacter capsulatus SB1003. J. Bacteriol. 2007, 189, 1774–1782. [Google Scholar] [CrossRef] [Green Version]
- Ilbert, M.; Bonnefoy, V. Insight into the evolution of the iron oxidation pathways. Biochim. Biophys. Acta (BBA)-Bioenerg. 2013, 1827, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, Z.; Belchik, S.M.; Edwards, M.J.; Liu, C.; Kennedy, D.W.; Merkley, E.D.; Lipton, M.S.; Butt, J.N.; Richardson, D.J. Identification and characterization of MtoA: A decaheme c-type cytochrome of the neutrophilic Fe (II)-oxidizing bacterium Sideroxydans lithotrophicus ES-1. Front. Microbiol. 2012, 3, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, Y.; Newman, D.K. The pio operon is essential for phototrophic Fe (II) oxidation in Rhodopseudomonas palustris TIE-1. J. Bacteriol. 2007, 189, 1765–1773. [Google Scholar] [CrossRef] [Green Version]
- Garber, A.I.; Nealson, K.H.; Okamoto, A.; McAllister, S.M.; Chan, C.S.; Barco, R.A.; Merino, N. FeGenie: A comprehensive tool for the identification of iron genes and iron gene neighborhoods in genome and metagenome assemblies. Front. Microbiol. 2020, 11, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kranz, R.G.; Richard-Fogal, C.; Taylor, J.-S.; Frawley, E.R. Cytochrome c biogenesis: Mechanisms for covalent modifications and trafficking of heme and for heme-iron redox control. Microbiol. Mol. Biol. Rev. 2009, 73, 510–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, M.C.; Rankin, J.A.; Kranz, R.G. Heme trafficking and modifications during system I cytochrome c biogenesis: Insights from heme redox potentials of Ccm proteins. Biochemistry 2016, 55, 3150–3156. [Google Scholar] [CrossRef] [Green Version]
- Enoch, H.G.; Lester, R.L. The role of a novel cytochrome b-containing nitrate reductase and quinone in the in vitro reconstruction of formate-nitrate reductase activity of E. coli. Biochem. Biophys. Res. Commun. 1974, 61, 1234–1241. [Google Scholar] [CrossRef]
- Meier, D.V.; Bach, W.; Girguis, P.R.; Gruber-Vodicka, H.R.; Reeves, E.P.; Richter, M.; Vidoudez, C.; Amann, R.; Meyerdierks, A. Heterotrophic Proteobacteria in the vicinity of diffuse hydrothermal venting. Environ. Microbiol. 2016, 18, 4348–4368. [Google Scholar] [CrossRef]
- Kaye, J.Z.; Marquez, M.C.; Ventosa, A.; Baross, J.A. Halomonas neptunia sp. nov., Halomonas sulfidaeris sp. nov., Halomonas axialensis sp. nov. and Halomonas hydrothermalis sp. nov.: Halophilic bacteria isolated from deep-sea hydrothermal-vent environments. Int. J. Syst. Evolut. Microbiol. 2004, 54, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Raguénès, G.; Christen, R.; Guezennec, J.; Pignet, P.; Barbier, G. Vibrio diabolicus sp. nov., a new polysaccharide-secreting organism isolated from a deep-sea hydrothermal vent polychaete annelid, Alvinella pompejana. Int. J. Syst. Evolut. Microbiol. 1997, 47, 989–995. [Google Scholar]
- Takai, K.; Sugai, A.; Itoh, T.; Horikoshi, K. Palaeococcus ferrophilus gen. nov., sp. nov., a barophilic, hyperthermophilic archaeon from a deep-sea hydrothermal vent chimney. Int. J. Syst. Evolut. Microbiol. 2000, 50, 489–500. [Google Scholar] [CrossRef]
- Vetriani, C.; Chew, Y.S.; Miller, S.M.; Yagi, J.; Coombs, J.; Lutz, R.A.; Barkay, T. Mercury adaptation among bacteria from a deep-sea hydrothermal vent. Appl. Environ. Microbiol. 2005, 71, 220–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaye, J.Z.; Baross, J.A. High incidence of halotolerant bacteria in Pacific hydrothermal-vent and pelagic environments. FEMS Microbiol. Ecol. 2000, 32, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Nakawake, M.; Kita, J.; Yamanaka, T.; Utsumi, M.; Okamura, K.; Ishibashi, J.-i.; Ohkuma, M.; Yamagishi, A. Characteristics of microbial communities in crustal fluids in a deep-sea hydrothermal field of the Suiyo Seamount. Front. Microbiol. 2013, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Chiu, B.K.; Kato, S.; McAllister, S.M.; Field, E.K.; Chan, C.S. Novel pelagic iron-oxidizing Zetaproteobacteria from the Chesapeake Bay oxic–anoxic transition zone. Front. Microbiol. 2017, 8, 1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAllister, S.M.; Davis, R.E.; McBeth, J.M.; Tebo, B.M.; Emerson, D.; Moyer, C.L. Biodiversity and emerging biogeography of the neutrophilic iron-oxidizing Zetaproteobacteria. Appl. Environ. Microbiol. 2011, 77, 5445–5457. [Google Scholar] [CrossRef] [Green Version]
- Emerson, D.; Moyer, C.L. Microbiology of seamounts: Common patterns observed in community structure. Oceanography 2010, 23, 148–163. [Google Scholar] [CrossRef]
- Emerson, D.; Moyer, C.L. Neutrophilic Fe-oxidizing bacteria are abundant at the Loihi Seamount hydrothermal vents and play a major role in Fe oxide deposition. Appl. Eenviron. Microbiol. 2002, 68, 3085–3093. [Google Scholar] [CrossRef]
- Mori, J.F.; Scott, J.J.; Hager, K.W.; Moyer, C.L.; Küsel, K.; Emerson, D. Physiological and ecological implications of an iron-or hydrogen-oxidizing member of the Zetaproteobacteria, Ghiorsea bivora, gen. nov., sp. nov. ISME J. 2017, 11, 2624–2636. [Google Scholar] [CrossRef] [Green Version]
- Litchman, E. Resource competition and the ecological success of phytoplankton. In Evolution of Primary Producers in the Sea; Paul, G., Falkowski, A., Knoll, H., Eds.; Academic Press: New York, NY, USA, 2007; pp. 351–375. [Google Scholar]
- Segerer, A.H.; Burggraf, S.; Fiala, G.; Huber, G.; Huber, R.; Pley, U.; Stetter, K.O. Life in hot springs and hydrothermal vents. Orig. Life Evol. Biosph. 1993, 23, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Dick, G.J. The microbiomes of deep-sea hydrothermal vents: Distributed globally, shaped locally. Nat. Rev. Microbiol. 2019, 17, 271–283. [Google Scholar] [CrossRef]
- Morris, R.M.; Spietz, R.L. The Physiology and Biogeochemistry of SUP05. Annu. Rev. Mar. Sci. 2022, 14, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y. Oxidation of inorganic sulfur compounds by obligately organotrophic bacteria. Microbiology 2003, 72, 641–653. [Google Scholar] [CrossRef]
- Du, R.; Gao, D.; Wang, Y.; Liu, L.; Cheng, J.; Liu, J.; Zhang, X.H.; Yu, M. Heterotrophic sulfur oxidation of Halomonas titanicae SOB56 and its habitat adaptation to the hydrothermal environment. Front. Microbiol. 2022, 13, 888833. [Google Scholar] [CrossRef] [PubMed]
- Straub, K.L.; Benz, M.; Schink, B.; Widdel, F. Anaerobic, nitrate-dependent microbial oxidation of ferrous iron. Appl. Environ. Microbiol. 1996, 62, 1458–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Shao, S.; Huang, T.; Ma, F.; Yang, S.; Zhou, Z.; Zheng, S. Anaerobic nitrate-dependent iron (II) oxidation by a novel autotrophic bacterium, Pseudomonas sp. SZF15. J. Environ. Chem. Eng. 2015, 3, 2187–2193. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Teske, A.; Robertson, L.A.; Kuenen, J.G. Anaerobic oxidation of thiosulfate to tetrathionate by obligately heterotrophic bacteria, belonging to the Pseudomonas stutzeri group. FEMS Microbiol. Ecol. 1999, 30, 113–123. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic relationships between cultured strains in this study (blue), reported autotrophic Fe oxidizers (red) and close relatives (black). The tree is a maximum-likelihood tree, based on the small-subunit (16S) rRNA gene (723 bp), showing bootstrap values (out of 1000 replicates).
Figure 1.
Phylogenetic relationships between cultured strains in this study (blue), reported autotrophic Fe oxidizers (red) and close relatives (black). The tree is a maximum-likelihood tree, based on the small-subunit (16S) rRNA gene (723 bp), showing bootstrap values (out of 1000 replicates).

Figure 2.
Light (A), fluorescence (B), and electron (C) microscopic images of strain Pseudomonas sp. IOP_13. Cells were grown anaerobically with ferrous chloride (FeCl2) and nitrate (NO3−).
Figure 2.
Light (A), fluorescence (B), and electron (C) microscopic images of strain Pseudomonas sp. IOP_13. Cells were grown anaerobically with ferrous chloride (FeCl2) and nitrate (NO3−).

Figure 3.
Fe(II) and nitrate consumption of Pseudomonas sp. IOP 13 in the presence of nitrate (10 mM) and ferrous iron over a period of 40 days. (A) Fe concentration in solution, (B) Fe concentration in precipitates.
Figure 3.
Fe(II) and nitrate consumption of Pseudomonas sp. IOP 13 in the presence of nitrate (10 mM) and ferrous iron over a period of 40 days. (A) Fe concentration in solution, (B) Fe concentration in precipitates.

Figure 4.
The growth of isolates in autotrophic sulfur-containing media. (A) the growth curve of Pseudomonas sp. IOP 13, (B) the growth curve of Halomonas sp. IOP_14, and (C) the pH change in cultures of strain IOP_13 and strain IOP_14.
Figure 4.
The growth of isolates in autotrophic sulfur-containing media. (A) the growth curve of Pseudomonas sp. IOP 13, (B) the growth curve of Halomonas sp. IOP_14, and (C) the pH change in cultures of strain IOP_13 and strain IOP_14.

Figure 5.
Growth curve and carbon-fixation capacity of Pseudomonas sp. IOP_13 grown in an autotrophic thiosulfate–containing medium.
Figure 5.
Growth curve and carbon-fixation capacity of Pseudomonas sp. IOP_13 grown in an autotrophic thiosulfate–containing medium.

Figure 6.
Functional gene annotations of the isolates in this study. Ccm, c-type cytochrome membrane proteins; CumA, multicopper oxidases; TsdA, tetrathionate-forming thiosulfate dehydrogenase; Sox, sulfur-oxidation gene (SoxABCDHSFRSWXYZ); Nar, nitrate reductase; Nor, nitric oxide reductase; Nir, nitrite reductase.
Figure 6.
Functional gene annotations of the isolates in this study. Ccm, c-type cytochrome membrane proteins; CumA, multicopper oxidases; TsdA, tetrathionate-forming thiosulfate dehydrogenase; Sox, sulfur-oxidation gene (SoxABCDHSFRSWXYZ); Nar, nitrate reductase; Nor, nitric oxide reductase; Nir, nitrite reductase.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
GenBank accession numbers of 16S rRNA genes and genomes for isolates in this study.
| MCCC Deposition No. | Strains | Source | Closest Species | 16S rDNA Identity (%) | 16S rDNA GenBank Accession | Genome GenBank Accession |
|---|---|---|---|---|---|---|
| M28193 | IOP_1 | mussels | Thalassospira xiamenensis M-5(T) | 98.79 | MW800032 | JAINWB000000000 |
| MCCC 1A14002 | IOP_2 | mussels | Agromyces soli MJ21(T) | 94.19 | MW805711 | JAINWC000000000 |
| MCCC 1A14012 | IOP_6 | sulfides | Halomonas zincidurans B6 (T) | 98.64 | MW805712 | JAIRBO000000000 |
| MCCC 1A13998 | IOP_12 | hydrothermal sediments | Qipengyuania citrea RE35F/1(T) | 98.78 | MW805714 | JAINWE000000000 |
| M28194 | IOP_13 | hydrothermal sediments | Pseudomonas kunmingensis HL22-2(T) | 98.61 | MW805738 | JAINWF000000000 |
| M28195 | IOP_14 | plume | Halomonas titanicae BH1(T) | 98.48 | MW805742 | JAINWD000000000 |
| MCCC 1A13999 | IOP_16 | plume | Salipiger manganoxidans VSW210(T) | 99.48 | MW805743 | JAINWH000000000 |
| MCCC 1A14001 | IOP_19 | sulfides | Halomonas meridiana DSM 5425 (T) | 98.82 | MW805750 | JAINWP000000000 |
| MCCC 1A14003 | IOP_21 | sulfides | Rheinheimera pleomorphica PKS7 (T) | 99.22 | MW805752 | JAINWG000000000 |
| MCCC 1A14004 | IOP_23 | sulfides | Seohaeicola saemankumensis SD-15 (T) | 99.04 | MW805753 | JAINWI000000000 |
| MCCC 1A14005 | IOP_24 | sulfides | Martelella mediterranea DSM 17316 (T) | 98.42 | MW805754 | JAINWJ000000000 |
| M28196 | IOP_25 | sulfides | Pseudomonas stutzeri ATCC 17588(T) | 98.55 | MW805758 | JAINWK000000000 |
| MCCC 1A14006 | IOP_28 | sulfides | Citromicrobium bathyomarinum JF-1(T) | 99.93 | MZ048019 | JAINWL000000000 |
| MCCC 1A14007 | IOP_29 | sulfides | Marinobacter adhaerens HP15(T) | 100.00 | MW805759 | JAINWM000000000 |
| MCCC 1A14008 | IOP_31 | sulfides | Halomonas hydrothermalis Slthf2(T) | 99.64 | MW805761 | JAIRBP000000000 |
| MCCC 1A14010 | IOP_38 | sulfides | Aurantimonas coralicida DSM 14790(T) | 99.93 | MW805764 | JAINWN000000000 |
| MCCC 1A14013 | IOP_41 | sulfides | Marinobacter shengliensis LZ-6(T) | 99.86 | MW805765 | JAINWO000000000 |
Table 2.
Growth substrate tests of isolates in this study.
| Strains | Most Similar Type Species | Iron Oxidiation | Manganese Oxidation | Sulfur Oxidation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Autotrophic-Microaerobic | Hetertrophic-Anaerobic | Heterotrophic | Autotrophic | Heterotrophic | ||||||
| Fe0 | FeS | FeCO3 | Pyrite | Basalt | FeCl2 | MnCl2 | S2O32− | S2O32− | ||
| Gammaproteobacteria | ||||||||||
| IOP_6 | Halomonas zincidurans B6 (T) | + | + | + | − | + | + | − | + | − |
| IOP_14 | Halomonas titanicae BH1(T) | + | + | + | − | − | + | − | + | + |
| IOP_19 | Halomonas meridiana DSM 5425 (T) | + | + | + | − | − | + | − | + | + |
| IOP_31 | Halomonas hydrothermalis Slthf2(T) | + | + | + | − | − | + | − | + | − |
| IOP_13 | Pseudomonas kunmingensis HL22-2(T) | + | + | + | + | − | + | + | + | − |
| IOP_25 | Pseudomonas stutzeri ATCC 17588(T) | + | + | + | + | − | + | + | + | − |
| IOP_29 | Marinobacter adhaerens HP15(T) | + | + | + | + | − | + | − | + | + |
| IOP_41 | Marinobacter shengliensis LZ-6(T) | + | + | + | − | − | + | − | + | − |
| IOP_21 | Rheinheimera pleomorphica PKS7 (T) | − | + | − | − | − | + | − | − | + |
| Alphaproteobacteria | ||||||||||
| IOP_1 | Thalassospira xiamenensis M-5(T) | + | + | + | − | − | − | − | + | + |
| IOP_12 | Qipengyuania citrea RE35F/1(T) | − | + | − | + | − | − | − | − | + |
| IOP_16 | Salipiger manganoxidans VSW210(T) | + | + | − | − | − | + | + | − | + |
| IOP_23 | Seohaeicola saemankumensis SD-15 (T) | + | + | − | + | − | + | − | − | + |
| IOP_24 | Martelella mediterranea DSM 17316 (T) | + | + | − | + | − | + | − | − | + |
| IOP_28 | Citromicrobium bathyomarinum JF-1(T) | − | + | − | − | − | + | − | + | − |
| IOP_38 | Aurantimonas coralicida DSM 14790(T) | + | + | + | + | − | − | − | + | + |
| Actinobacteria | ||||||||||
| IOP_2 | Agromyces soli MJ21(T) | − | + | − | + | + | − | − | + | − |
| NC1 | − | − | − | − | − | − | − | − | − | |
| NC2 | − | − | − | − | − | − | − | − | − | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
He, Y.; Zeng, X.; Xu, F.; Shao, Z. Diversity of Mixotrophic Neutrophilic Thiosulfate- and Iron-Oxidizing Bacteria from Deep-Sea Hydrothermal Vents. Microorganisms 2023, 11, 100. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11010100
AMA Style
He Y, Zeng X, Xu F, Shao Z. Diversity of Mixotrophic Neutrophilic Thiosulfate- and Iron-Oxidizing Bacteria from Deep-Sea Hydrothermal Vents. Microorganisms. 2023; 11(1):100. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11010100
Chicago/Turabian StyleHe, Yang, Xiang Zeng, Fei Xu, and Zongze Shao. 2023. "Diversity of Mixotrophic Neutrophilic Thiosulfate- and Iron-Oxidizing Bacteria from Deep-Sea Hydrothermal Vents" Microorganisms 11, no. 1: 100. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms11010100
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.