1. Introduction
The human small intestine plays a significant role in nutrient absorption and homeostasis maintenance via host–microbe interactions. Due to the difficulty of accessing the human small intestine for monitoring dietary effects on gut homeostasis and nutrient absorption, in vitro models can be used to gain this knowledge. The intestinal environment is rich in bacterial populations ranging from 10
1–10
3 bacteria per gram of contents in the duodenum to 10
4–10
7 bacteria per gram of contents in the jejunum and ileum and 10
11–10
12 bacteria per gram of contents in the colon [
1]. The environment of the small intestine is primarily composed of Gram-positive bacterial species such as
Lactocaseibacillus and
Enterococcus in the jejunum,
Streptococcus in the duodenum and jejunum, and
Bifidum bacilli [
2] and Gram-negative
Escherichia coli in the distal ileum [
3]. The microbiota resides in the body as a community, competing for nutrients and metabolites, and are present in the mucus layer of the intestinal barrier facing the lumen. Under stressful conditions, disruption in the intestinal barrier and microbiota (dysbiosis) causes an altered permeability and disturbed intestinal function [
4,
5]. Intestinal alkaline phosphatase (IAP), a brush border membrane enzyme, which interacts with the microbiome to regulate the intestinal epithelial barrier, and its activity, together with intestinal permeability are often used as indicators of the integrity of the intestinal barrier.
The human small intestine, responsible for nutrient absorption and transport, is equipped with multiple transport proteins for nutrients such as glucose, proteins, and fatty acids [
6]. In a healthy individual, 85% of carbohydrates, 66–95% of proteins, and all fats are absorbed in the small intestine before moving to the large intestine [
7]. The gut microbiota draws its nutrition from the dietary components, and a change in diet can lead to changes in the microbial composition. For example, a change from a low-fat to a high-fat diet in mice led to a shift in their microbiota structure [
8].
Nanoparticles (NPs) are used in the food industry to enhance the taste, aesthetics, or shelf life of food [
9,
10]. Food-grade titanium dioxide (TiO
2), referred to as E171 in the European Union (EU), contains about 36% of particles in the nanoscale [
11] and is widely used in the food industry to improve the color of foods such as candies, chewing gums, pastries, and sauces [
10,
11,
12]. Regular consumption of TiO
2-coated dietary products can be significant. Recently, the European Union issued a ban on the use of E171 food-grade TiO
2 in Europe due to genotoxicity concerns [
13], which is expected to impact the trade of the commodity, but in the absence of potential alternatives to the additive, its use is currently inevitable. As part of the dietary intake, these nanoparticles pass through the human gastrointestinal (GI) tract to enter the intestinal environment. TiO
2 NPs can exhibit toxic effects through alteration of the gut microbiota and interference with nutrient transporters [
11]. Some studies using TiO
2 NPs show its involvement in increased inflammatory cytokine production, disruption of the structure of the intestinal epithelium and mucus layer, and gut homeostasis both in vitro [
14] and in vivo [
15,
16]. Whereas other studies using rats show little or no effect of TiO
2 ingestion but the elimination of it through the excretory system [
17]. Such contradictory results call for the need to establish a standard study design to get more comparable and easily reproducible results.
This study uses a semi-synthetic food matrix supplemented with E171 food-grade TiO2 NPs to form a physiologically relevant food model for assessing the role of NPs on GI function. An in vitro model of the intestinal barrier was created using Caco-2 and HT29-MTX-E12 cell lines. The small intestinal microbiome is represented by adding planktonic cultures of individuals or a mix of bacterial species (forming a bacterial mock community), including Gram-negative Escherichia coli, and Gram-positive Lacticaseibacillus rhamnosus, Streptococcus salivarius, Bifidobacterium bifidum, and Enterococcus faecalis. The intestinal monolayer with bacteria was exposed to the food matrix, with or without NP, following an in vitro digestion to assess the effects of NPs and bacterial species on intestinal permeability, IAP activity, and nutrient transport. This study presents a novel in vitro intestinal model with a multispecies community of bacteria representing the microbiome, and a standard food matrix to mimic the small intestinal environment that can be used to assess how food additives affect the GI environment.
2. Materials and Methods
2.1. Intestinal Co-Culture
The human colon carcinoma Caco-2 cell line was purchased from the American Type Culture Collection (Manassas, VA, USA) at passage 17 and used for experimentation at passage 45–55. The HT29-MTX-E12 cell line was purchased from Sigma-Aldrich, St. Louis, MO, USA, and used in experiments at passage 50–60. These cells were grown in Dulbecco’s Modified Eagle Medium (DMEM, Gibco® Thermo Fisher Scientific, Waltham, MA, USA), containing 4.5 g/L glucose and 10% (v/v) heat-inactivated fetal bovine serum (HI-FBS, Gibco® Thermo Fisher Scientific, Waltham, MA, USA). The cells were maintained in 5% CO2 at 37 °C, and the media was changed every 48 h. Once the cells reached 80% confluency, they were passaged and seeded onto polycarbonate, 0.4 µm pore size, 0.33 cm2 membrane, 24-well Transwell® inserts for permeability, nutrient transport, and bacterial viability studies, or onto polycarbonate 96-well culture plates for brush border membrane enzyme activity assessment. Prior to seeding, the wells/inserts were coated with rat tail Type I collagen (BD Biosciences, San Jose, CA, USA) at a concentration of 8 μg/cm2 for 1 h at room temperature. Both types of cells were stained with trypan blue, counted using a hemocytometer, and seeded at a density of 105 cells/cm2 and a ratio of 75:25 (Caco-2:HT29). The co-culture of cells was grown for 14 days before experiments were conducted.
2.2. Bacterial Cell Culture
Escherichia coli (ATCC 53103), Lacticaseibacillus rhamnosus (ATCC 11775), Streptococcus salivarius SS2, Bifidobacterium bifidum VPI 1124, and Enterococcus faecalis NCTC 775 were purchased from the American Type Culture Collection. The bacterial species were cultured in brain heart infusion (BHI) broth medium supplemented with 0.5% glucose, 0.05% cysteine, and 0.1% agar. E. coli and L. rhamnosus cultures were grown at 37 °C, 5% CO2 for 24 and 48 h, respectively. B. bifidum was cultured in an anaerobic GasPak at 37 °C for 48 h. S. salivarius cultures were incubated under normal atmospheric conditions at 37 °C for 24 h. E. faecalis was cultured for 12 h at 37 °C, 5% CO2 condition. The bacterial stocks were diluted in 0.9% saline solution, plated on BHI agar medium supplemented with 0.5% glucose, 0.05% cysteine, and 1.5% agar, and grown in their respective culture conditions. The optical density (OD) of each dilution was read at a wavelength of 600 nm. The colonies formed on agar plates were quantified and associated with the OD600 measured for each dilution to prepare a standard curve. Prior to each experiment, bacterial concentrations were estimated using OD600. Planktonic culture of the five individual bacterial species was added to the apical chamber of the Transwell plates or in the 96-well plate at a concentration of 103 CFU/mL in combination with the control or test conditions. Bacteria mix (BM) of the five species was prepared at a ratio of 30% L. rhamnosus, 30% B. bifidum, 20% S. salivarius, 15% E. faecalis, and 5% E. coli. Each bacterial species was diluted to 103 CFU/mL before combining in the said ratio to form the BM. The BM was combined with the control or test condition before experiments.
2.3. Food Model Preparation
The food model (FM) was based on the previously developed food matrix described by Zhang et al. [
18]. The FM consists of the nutrients from a standard US diet. Sodium caseinate (Sigma, 3.44%
w/
w), a protein source, was dissolved in 10 mL of sterile 10 mM phosphate buffer solution, pH 7. The solution was homogenized using a homogenizer at medium speed for 45 min. The solution was substituted with 3.42% (
w/
w) corn oil (fat source) and another 10 mL of sterile 10 mM phosphate buffer solution, followed by homogenization at high speed for 15 min to obtain a fine emulsion. Finally, 0.7% (
w/
w) pectin (dietary fiber source), 5.15% (
w/
w) starch, 4.57% (
w/
w) sucrose (sugar source), and 0.534% sodium chloride (mineral source) were added to the emulsion with continuous stirring in between. The prepared nutrient emulsion was frozen at −80 °C overnight and freeze-dried using a freeze dryer for a week. The freeze-dried product was referred to as the food model and stored at −20 °C until needed. The FM was diluted and streaked on BHI agar plates to check for sterility. For experiments, 1 g of food model was subjected to in vitro digestion and added to the seeded assay plates.
2.4. Nanoparticle Preparation
E171 food-grade titanium dioxide nanoparticles (TiO
2 NPs) were purchased from Fiori Colori (Aromata Group). The TiO
2 NPs have been previously characterized [
19]. TiO
2 NPs were sonicated at a concentration of 1 mg/mL (prepared using sterile 18 MΩ water) for 2 min at 10% amplitude on ice. The sonicated solution was diluted 100 times in sterile 18 MΩ water, and 5.92 µL of the diluted NP solution was used for experiments to achieve a final dose of 1.25 × 10
−6 mg/cm
2 or 1.25 µg/cm
2. The calculations are based on the surface area of the small intestine and average human intake of TiO
2 [
5,
20]. TiO
2 was combined with 1 g of food model to represent the FM + TiO
2 condition.
2.5. In Vitro Digestion
The control and test conditions were digested in vitro in a sterile environment prior to experimental exposure. A serum-free, 5 mM glucose DMEM solution served as a control condition, while test conditions included FM, TiO
2, and FM + TiO
2. The in vitro digestion protocol has been previously described by Glahn et al. [
21]. The weighed samples were dissolved in 10 mL of sterile 140 mM NaCl + 5 mL KCl with a pH of 2. The pH was then re-adjusted to 2 with sterile 0.1 M HCl. Next, 0.5 mL of a porcine pepsin solution (25 mg/mL) prepared in 0.1 M HCl was added to each sample. The samples were then incubated for 1 h at 37 °C, 5% CO
2 on a rocker (55 oscillations/min). This constituted the “gastric” phase of digestion. This was followed by “intestinal” digestion, where the pH of the samples was increased to 5.5–6 using sterilized 0.1 M NaHCO
3 or 1 M NaHCO
3. A pancreatin bile solution was prepared using 2 mg/mL pancreatin (a mixture of trypsin, amylase, lipase, ribonuclease, and protease activities) and 11 mg/mL bile extract (a mixture of glycine and taurine conjugates of hyodeoxycholic and other bile acids) and filter sterilized with a 0.45 μm filter. Next, 2.5 mL of the prepared pancreatin-bile solution was added to each sample, and the pH was brought up to 6.9–7 using sterile 0.1 M NaHCO
3 or 1 M NaHCO
3. The volume of each sample tube was made up to 15 mL by weight using a sterile 140 mM NaCl + 5 mM KCl solution, pH 6.7. The samples were then referred to as digests and used for exposure studies.
2.6. Permeability Assay
The permeability assay was used to quantify the changes in barrier integrity using Lucifer yellow (LY, Life Technologies L453), a fluorescent paracellular permeability marker. LY stock of 1 mg/mL was prepared aseptically in 18 MΩ water and stored at 4 °C until needed. A standard curve of LY was plotted using 0, 10, 20, 30, 40, and 50 μM LY concentrations and the fluorescence values for each concentration. A 100 μL of digested sample was added to the apical chamber of Transwell plates with 100 μL of 50 μM LY. Samples (100 μL) were then collected from the basolateral chamber every 15 min for the first hour and every 30 min for the next three hours and transferred into a 96-well opaque black bottom plate. The removed 100 μL volume, when sampling, was substituted by adding 100 μL DMEM to the basolateral chamber of the Transwell plate. At the end of the exposure experiment, the 96-well plate was read at 485/560 nm using a Synergy 2 plate reader, controlled by Biotek’s Gen5™ Reader Control and Data Analysis Software (V1.11.5). The resulting fluorescence values of the samples and the standards were used to calculate the permeability (cm/s) across the intestinal monolayer.
2.7. Intestinal Alkaline Phosphatase (IAP) and Bradford Assays
IAP activity was determined using IAP and Bradford assay. The sample digests were added to the seeded 96-well plates and allowed a 4 h exposure at 37 °C, 5% CO2. At the end of the 4 h exposure to digested control or test conditions, the wells were washed with 1× PBS, followed by the addition of 200 μL of 18 MΩ water. The plate was then subjected to sonication at 4 °C for 15 min. The samples were scraped off the bottom of the wells and subsequently used for the assay. A standard curve was prepared with dilutions of a standard solution of 0.25 mg/mL p-nitrophenol, prepared in p-nitrophenyl phosphate (pNPP). The assay was carried out by adding 25 μL of sample or standard solution in 85 μL of pNPP solution to a 96-well clear bottom plate, which was then incubated for 1 h at room temperature. For the Bradford assay, a standard curve was prepared using a 1 mg/mL bovine serum albumin (BSA, Rockland, NY, USA) solution prepared in 18 MΩ water. The experimental setup included a 5 μL sample or standard solution and 250 μL of Bradford reagent (Sigma) in a 96-well clear bottom plate. The plate was incubated at room temperature for 15–30 min in the dark. The IAP assay and Bradford assay plates were read at 405 nm and 595 nm, respectively, using the Synergy 2 plate reader, controlled by Biotek’s Gen5™ Reader Control and Data Analysis Software.
2.8. Nutrient Transport Assay
The Caco-2 and HT29-MTX E12 cell monolayer was grown on Transwell membranes for 14 days before performing nutrient transport assays. During the 14-day period, tight junction integrity is established, the Caco-2 cells mature into enterocytes, and a mucus layer is formed by HT29-MTX E12 cells [
22]. On day 15, the monolayers were exposed to digested FM and/or TiO
2 samples or control solutions, in combination with 10
3 CFU/well bacteria for 4 h. The DMEM medium used for the assays was serum free with 5 mM glucose. This ensured that the resulting protein and glucose content detected after the assays was a result of the nutrients in the food model. After the exposure period, samples were collected from the basolateral chamber for nutrient transport assays.
2.8.1. Glucose Assay
The basolateral samples were subjected to charcoal treatment due to low glucose levels [
23]. Activated charcoal (2%
w/
w) was added to the samples and mixed on a vortex for 30 s. The samples were then centrifuged for 30 s, and the supernatant was used for the assay. Prepared samples were added to glucose hexokinase liquid stable reagent (Infinity
TM) at a ratio of 1:150 in a 24-well clear bottom plate. Samples were incubated for 3 min at 37 °C, and the absorbance was read at 340 nm using a Synergy 2 plate reader, controlled by Biotek’s Gen5™ Reader Control and Data Analysis Software. The absorbance values along with the glucose standard (1 mg/mL in 0.1% benzoic acid) were used to calculate the amount of glucose (μg) per mL of each sample.
2.8.2. Protein Assay
The basolateral samples were subjected to charcoal treatment, as mentioned in
Section 2.8.1. The protein content was assessed using a Bradford reagent. A 5 μL supernatant sample was added to a 96-well clear bottom plate with 250 μL of Bradford reagent. The protein assay was carried out as described in
Section 2.6.
2.8.3. Triglyceride Assay
The fatty acid content in the basolateral samples was detected using a triglyceride assay kit (Sigma, MAK266). A triglyceride standard curve was prepared using dilutions of a 0.2 mM standard solution. A 50 μL sample or standard solution was added to a 96-well clear bottom plate, along with a 2 μL lipase solution. Samples were incubated for 20 min at room temperature. A master mix was prepared using 46 μL TG assay buffer, 2 μL TG probe, and 2 μL TG enzyme mix per well. A 50 μL of the prepared master mix was added to the plate at the end of incubation. Samples were then incubated for 30–60 min at room temperature in the dark. The absorbance values were read at 570 nm using a Synergy 2 plate reader, controlled by Biotek’s Gen5™ Reader Control and Data Analysis Software. Using the standard curve, the triglyceride content (μg) per μL sample was calculated.
2.9. Bacterial Viability Quantification
The 4 h exposure to independent bacterial species and digested samples was followed by bacterial quantification. Bacterial viability was quantified using viable counts and quantitative PCR.
2.9.1. Viable Counts
A 100 μL sample was collected from the apical and basolateral chambers of the Transwell plates. The samples were serially diluted with sterile 0.9% saline solution and plated on BHI agar plates using the drop-plate technique. The bacteria that adhered to the Caco-2/HT29-MTX E12 monolayer were harvested from the Transwell inserts. The wells were washed with 1× PBS, followed by cutting of the Transwell membrane. The membrane was then transferred to a 400 μL sterile saline solution and homogenized for 30 s per sample. The homogenized samples were diluted and plated on BHI agar plates. To allow selective growth of each bacterial species in the bacteria mix, the samples were plated on MRS agar for L. rhamnosus, MacConkey agar for E. coli, ESM agar for E. faecalis, using the drop plate technique, blood agar for S. salivarius, and BSM agar for B. bifidum using the spread plate technique. Plates were incubated at their respective optimum growth conditions for 24–48 h, based on the bacterial species, and bacterial colonies were quantified to determine the bacterial viability (total log CFU).
2.9.2. qPCR
Samples for qPCR bacterial quantification were harvested by scraping into sterile saline, and cells were collected by centrifugation (1 min at 18,000× g,), the supernatant discarded, and the cell pellet stored at −80 °C. Genomic DNA (gDNA) was extracted according to manufacturer instructions (ZymoBIOMICS DNA Miniprep Kit), and samples were stored at −80 °C. qPCR was performed with the PerfeCTa® SYBR® Green FastMix® (Quantabio), the primers for each bacterial species, and an Eppendorf Mastercycler Ep Realplex instrument. Primers were used to quantify B. bifidum (16s rRNA, FW ACGCGAAGAACCTTACCTGG, RV 5′-ATCTCACGACACGAGCTGAC-3′), E. faecalis (16s rRNA, FW 5′-TACGGCGAACATACAAAGCG-3′, RV 5′-ATTTTGAACACGCGACTGG-3′), L. rhamnosus (16s rRNA, FW 5′-AGACACGGCCCAAACTCTAC-3′, RV 5′-CGTTGCTCCATCAGACTTGC-3′), S. salivarius (Hypothetical Protein, FW 5′-TGCCTGGAACTAACGGTAGC-3′, RV 5′-CAACGACTTCAGAGCCTCCC-3′), and E. coli (tnaA, FW 5′-GACTGGACCATCGAGCAGAT-3′, RV 5′-CACGCAAAGGGTTCTGCACTC-3′). Standard curves were previously generated by relating CT values to viable cell counts and allowing experimental CT values to be interpolated into equivalent cell counts. In brief, cultures of target species were grown to stationary phase and quantified by drop plating. Genomic DNA was extracted from 1 mL of stationary-phase cultures and purified. DNA was serially diluted to produce standards of decreasing DNA concentration. Quantitative PCR was then performed on these standards, and the resulting CT values were plotted against viable cell counts. Standard curves were calculated to interpolate cell counts from experimental CT values.
2.10. Statistical Analysis
The data sets were tested for normality using the Shapiro–Wilk normality test and transformed using the Box–Cox transformation with appropriate conditions to achieve a normal distribution. The LY permeability results were analyzed using a one-way ANOVA. The IAP assay, bacterial viability, and nutrient transport assay results were compared using a two-way ANOVA followed by false discovery rate (FDR) correction using GraphPad Prism 8.3.1. FDR correction was performed using the Benjamini and Hochberg procedure [
24] implemented in the R [
25] package multi-test [
26].
4. Discussion
Food-grade TiO
2 was approved by the Food and Drug Administration (FDA) in 1966 as INS171 and by the European Union in 2006 as E171. However, there have been growing health concerns with the consumption of TiO
2 NPs due to their potential impact on the intestinal tract, the gut microbiota, and their potential to cross the gut and cause an immune reaction in the body. The host GI tract provides a constant nutrient supply and stable growth environment to the microorganisms while gaining protection from the pathogens due to their competitive exclusion by the commensal bacteria, thus forming a symbiotic relationship [
27].
The negative effects of TiO
2 NPs on the GI are widely published [
11,
14,
28]; however, the ingestion of such nanoparticles as part of a diet and their interaction with the gut microbiota have yet to be fully established. The properties of the NPs can change when added to food [
29] and after being absorbed by the GI, resulting in varied effects [
30,
31]. In this study, a normal US diet was created in the lab using all the essential nutrients, as previously described by Zhang et al. [
18]. This semi-synthetic meal, referred to as the food model (FM), represented a US diet with an additional intake of products rich in TiO
2 NPs (E171 grade) at a realistic dose of 1.25 μg/cm
2, calculated based on the average human intake of TiO
2 NPs and the surface area of the small intestine [
20,
32]. The food model and TiO
2 were subjected to a two-step in vitro digestion to create a physiologically relevant diet, which was then introduced into a Caco-2/HT29-MTX E12 intestinal model. Prior to exposure to the intestinal monolayer, the sterility of the food model was confirmed using the drop-plate culture technique. The food model or TiO
2 did not have any significant impact on the permeability of the barrier compared to the control condition, which showed a permeability of 10
−5 cm/s. However, when TiO
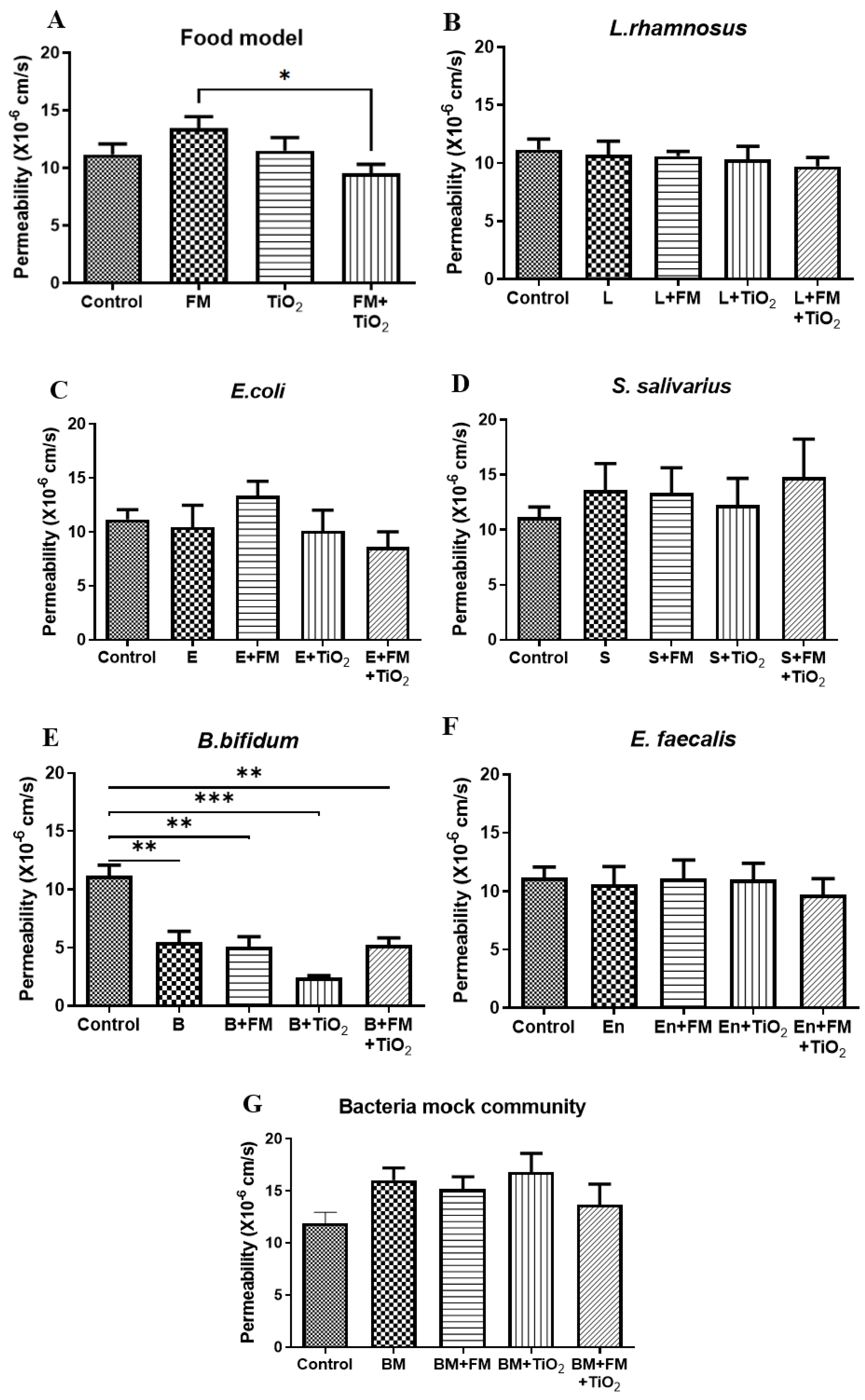
2 was incorporated into the food model, the permeability decreased by 29% compared to the food model alone (
Figure 1A).
The upper small intestine is predominantly composed of Gram-positive aerobes, and the terminal ileum has an equal number of aerobes and anaerobes such as
Bifidum bacilli [
2]. The Gram-positive lactobacilli comprise the majority of the normal flora in the jejunum. Lactobacilli, along with staphylococci and streptococci, are also present in the intestine distally [
33].
L. rhamnosus [
34,
35] and
Bifidobacterium spp. [
36] are well-known probiotic species with positive effects on intestinal health.
E. faecalis is a common commensal bacterium and is one of the first colonizers in an infant [
37], while
E. coli is a Gram-negative bacterium more prevalent in the colon and can also be found in high quantities in the small intestine [
3]. The bacteria mix in this study was designed to include a variety of bacterial species ranging from aerobes (
Enterococcus and
Escherichia) to facultative anaerobes (
Staphylococcus, Lactobacillus) and anaerobes (
Bifidobacterium). The broad selection of bacterial species enabled the investigation of the host–diet–microbiota interaction as close to the in vivo scenario as feasible. In this study, the presence of
B. bifidum resulted in a significant decrease in permeability across the in vitro intestinal barrier (
Figure 1E), while the other bacteria did not impact the permeability, indicating a quick response of this probiotic species towards regulating intestinal health. IAP is expressed by the enterocytes primarily in the duodenum and is heavily involved in the regulation of gut homeostasis [
38]. IAP can potentially restore the commensal gut bacteria through its ability to control the pH. IAP is a crucial contributor to the maintenance of gut homeostasis and health through interactions with the intestinal microbiota and diet. While the benefits of probiotic bacterial species such as
L. rhamnosus and
B. bifidum are widely established, the positive influence of other commensal species such as
E. coli is not as well known. The positive impact of
E. coli on IAP is suggestive of such benefits. The interaction between IAP levels and
E. coli has also been published in an in vivo model wherein wild-type mice showed higher levels of
Lactobacillaceae and commensal
E. coli populations compared to IAP knock-out mice. Upon oral administration of IAP, the colonization of
E. coli increased in the IAP knock-out population [
39]. The food model and TiO
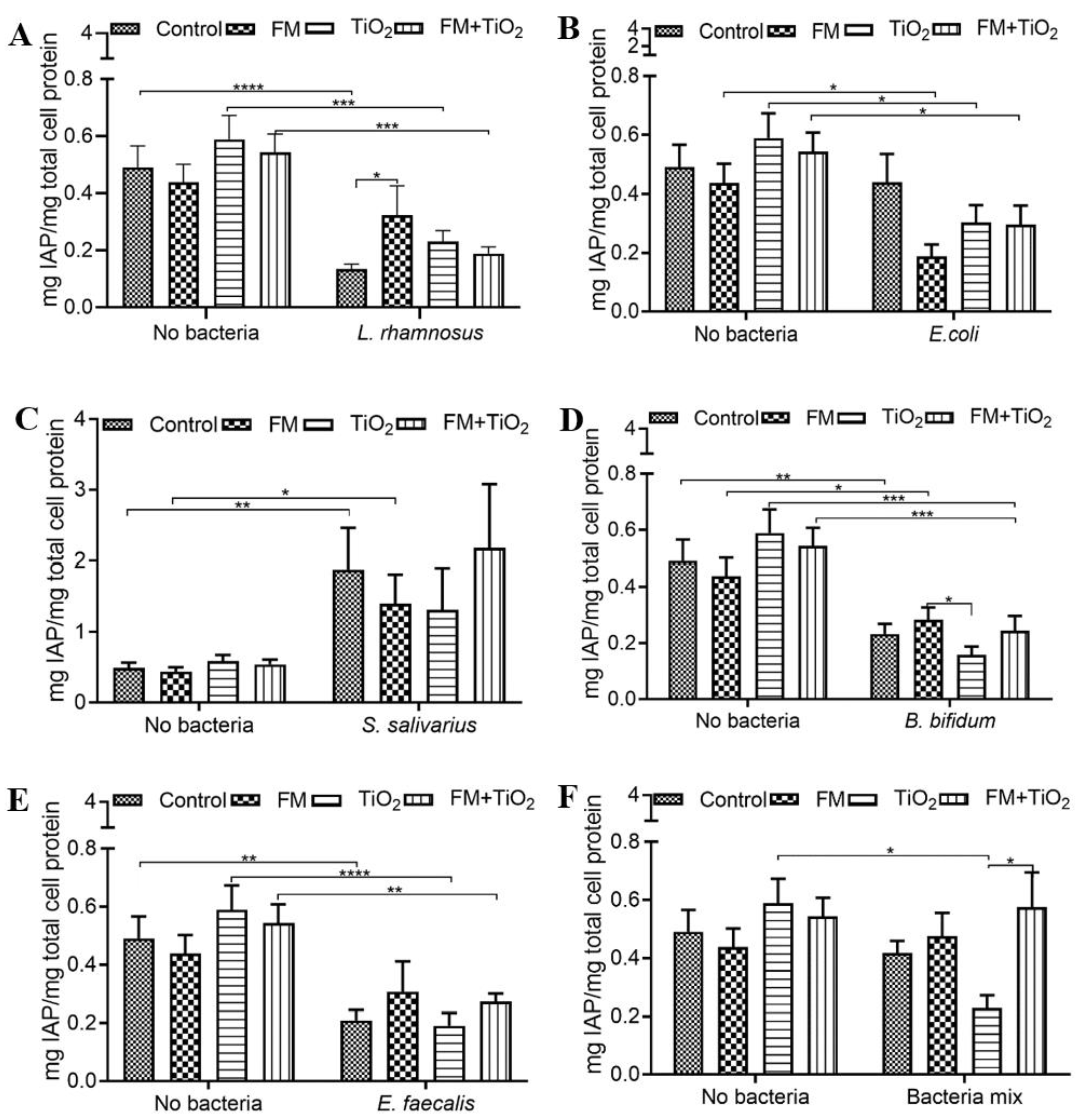
2 did not cause any changes in endogenous IAP activity compared to the control condition. However, as previously reported [
32], IAP was significantly reduced with the addition of
L. rhamnosus,
E. coli,
B. bifidum, and
E. faecalis (
Figure 2), indicative of the changing gut environment with the addition of bacteria. The five bacterial species used in this study were combined to form a bacterial mock community (BM) at a physiologically relevant ratio of the bacterial population [
40,
41]. The co-culture of mammalian and bacterial cells was allowed for a short duration (4 h), during which the experiments were carried out. The BM did not significantly alter the human cell monolayer permeability (
Figure 1G). As opposed to the change in IAP due to individual bacterial species, the bacterial mock community maintained the IAP at the same levels as no bacteria control in most cases (
Figure 2F), suggesting that as a community, the bacteria did not negatively impact the gut environment.
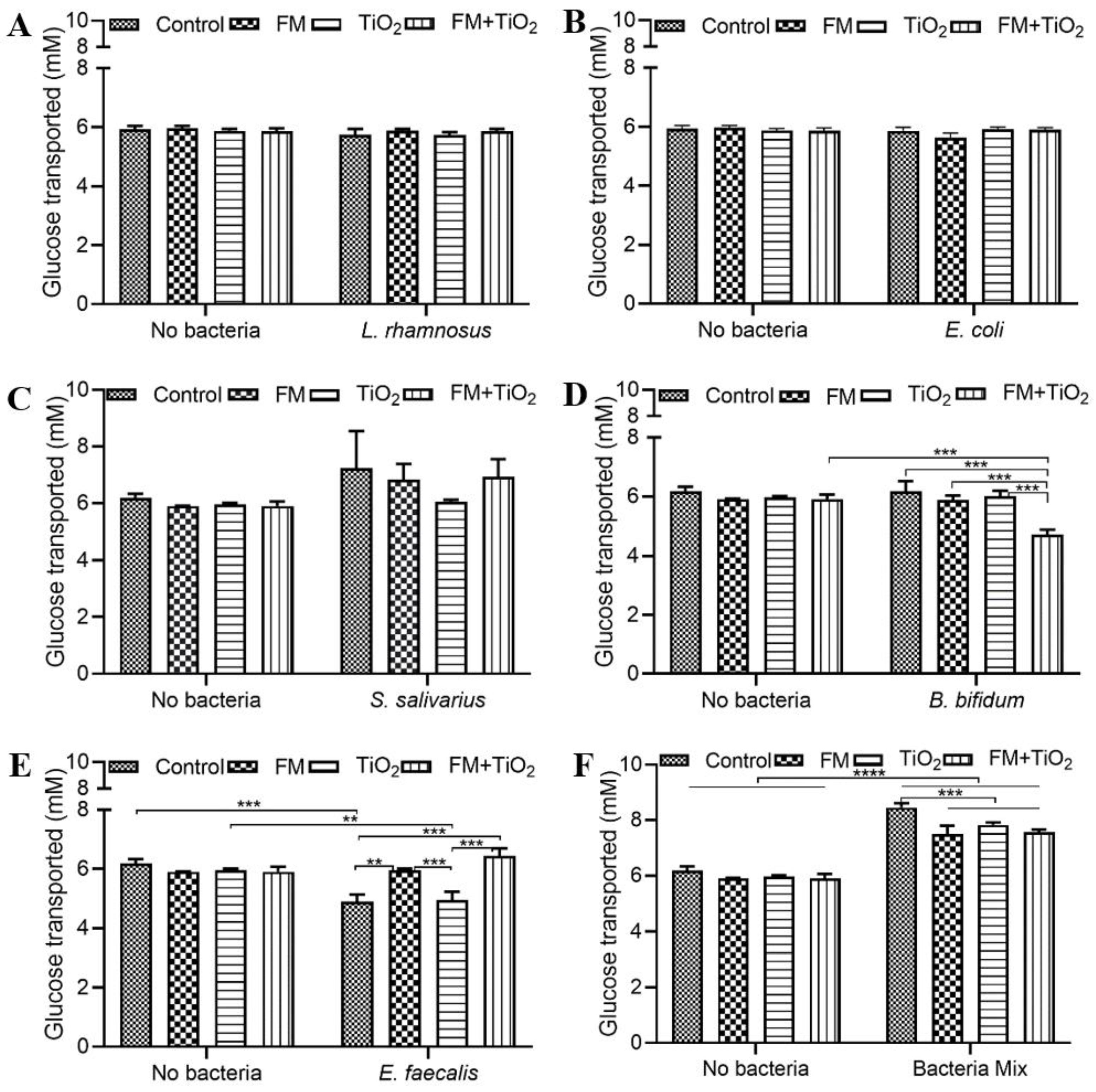
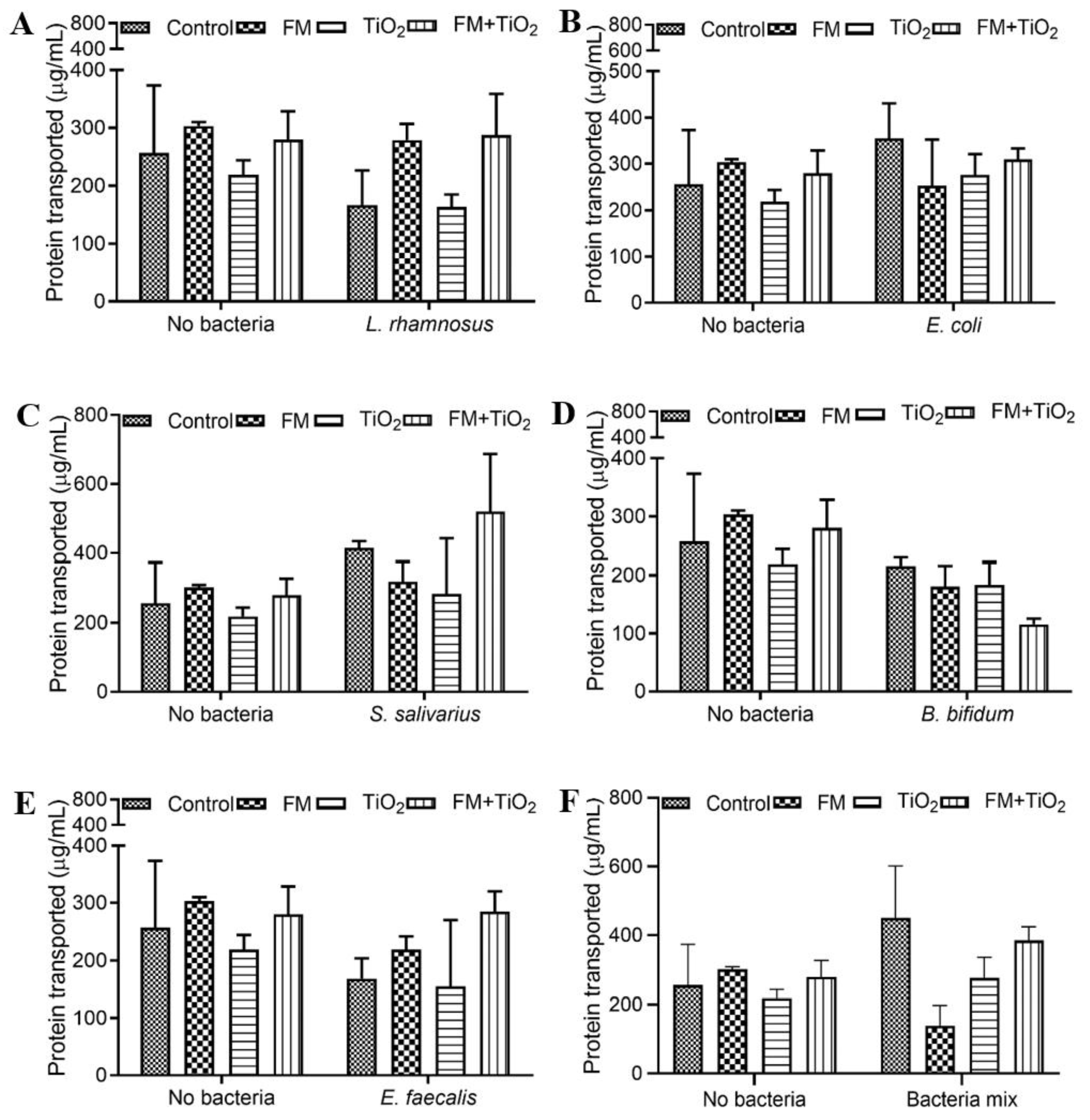
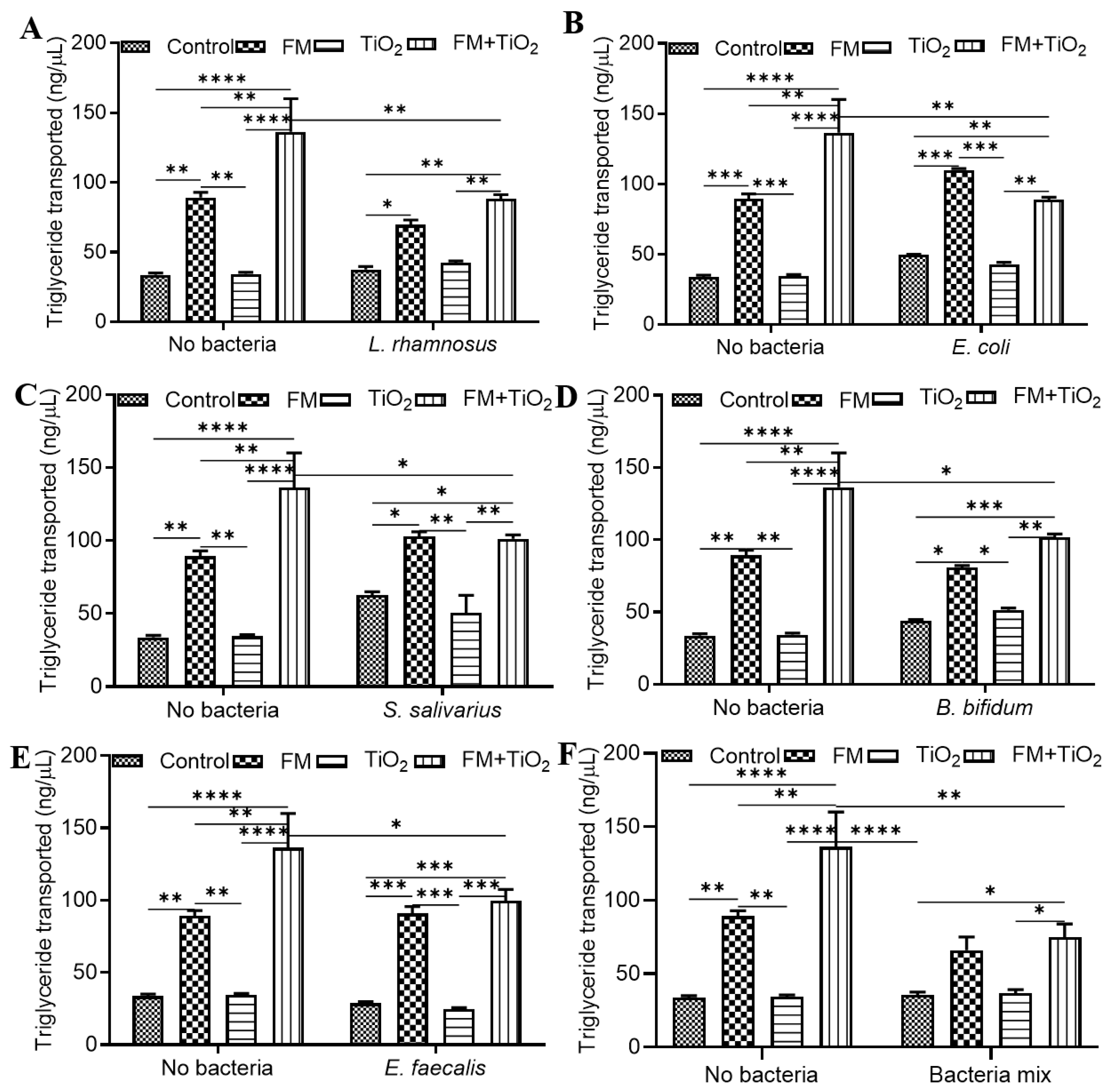
The human small intestine, as the primary site of nutrient absorption, can lead to nutritional deficiencies and malnutrition due to any disruption in the intestinal barrier. Therefore, factors that influence intestinal health may also impact the body’s nutritional availability. The intestinal microbiome plays a significant role in the de novo biosynthesis and bioavailability of several nutrients [
42]. In this study, three nutrients were selected to determine the relationship between NPs, diet, and gut microbiota. Glucose transport across the barrier was significantly increased with BM, while protein transport did not show any changes across conditions. Triglycerides do not traverse the cellular barrier due to their hydrophobicity. Instead, they are absorbed through hydrolysis into fatty acids and monoacylglycerol in the intestinal lumen. Intestinal epithelial cells, primarily in the upper villi [
43], take up the lipids and resynthesize them to triglycerides, which are secreted out in the form of chylomicrons. One of the components of the food model was fatty acids, which cause higher triglyceride transport in the presence of the FM, with a further increase with TiO
2 in the food matrix. Previous studies show a decrease in fatty acid absorption due to TiO
2 NPs [
19,
44]; in contrast, our results show that when NPs are part of a food matrix, their effect on TG transport changes. This suggests that while TiO
2 on its own may have a more adverse effect on intestinal permeability in this model, when consumed as part of a regular diet at a controlled level, it does not pose the same risk. However, further studies are needed to study not only TiO
2 absorption but also its accumulation in the body to assess the overall risk factors. The change in transport of each of these nutrients was different in response to the different bacteria. As part of a common community, the bacteria showed positive or no significant changes in protein and TG transport, except for the FM + TiO
2 condition. Glucose transport was enhanced with the bacterial mock community compared to the single bacteria condition (
Figure 3). Microbe–microbe interactions can have combined effects on intestinal health. Literature suggests improved blood glucose levels in the presence of beneficial bacteria upon supplementation with pre- and probiotic cultures [
45], which is seen in these results as well. Further supporting investigation with respect to insulin sensitivity and immune response would be needed, however, to determine this effect conclusively. Overall, the results confirm the successful incorporation of the bacterial mock community for acute exposure.
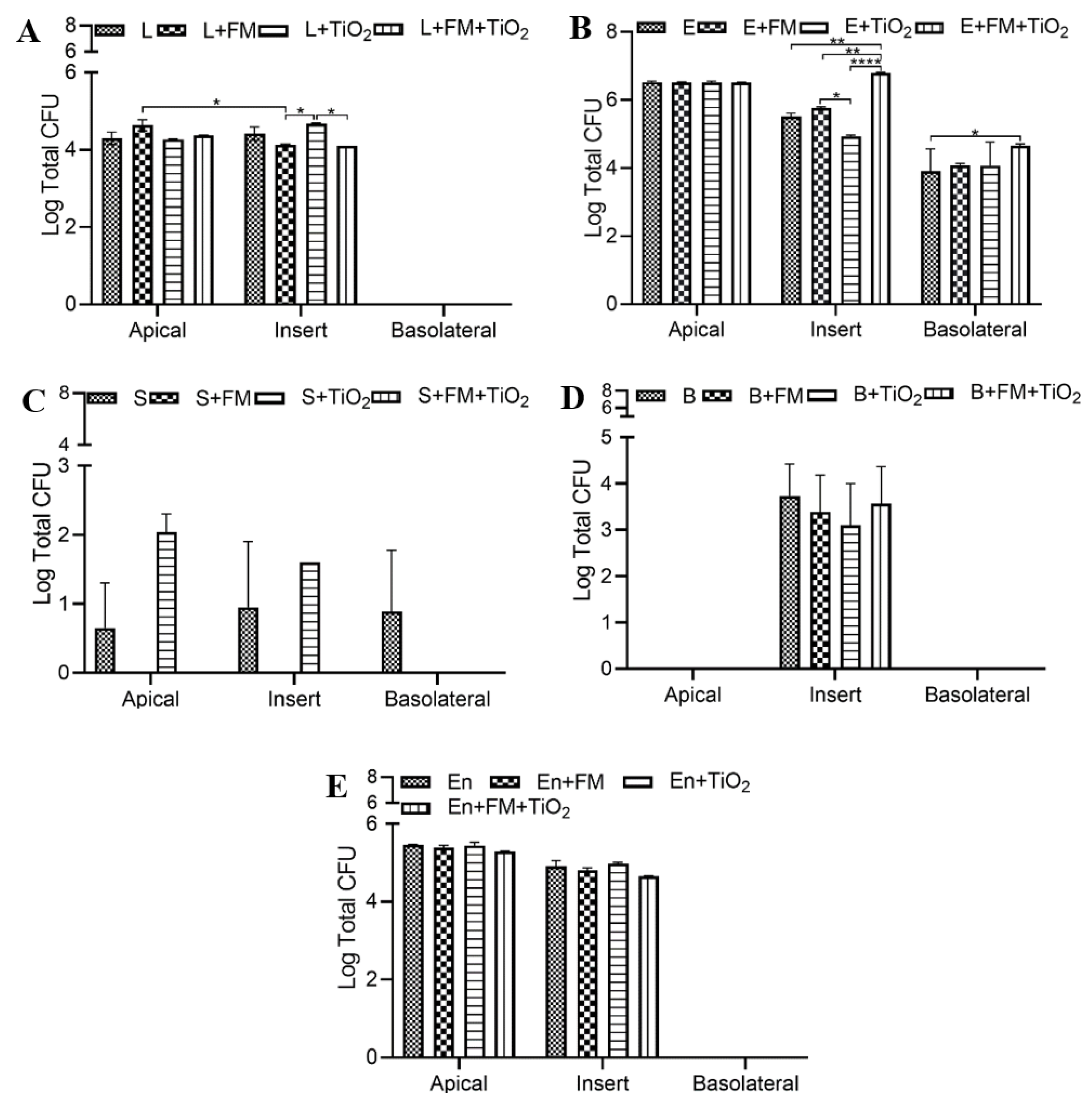
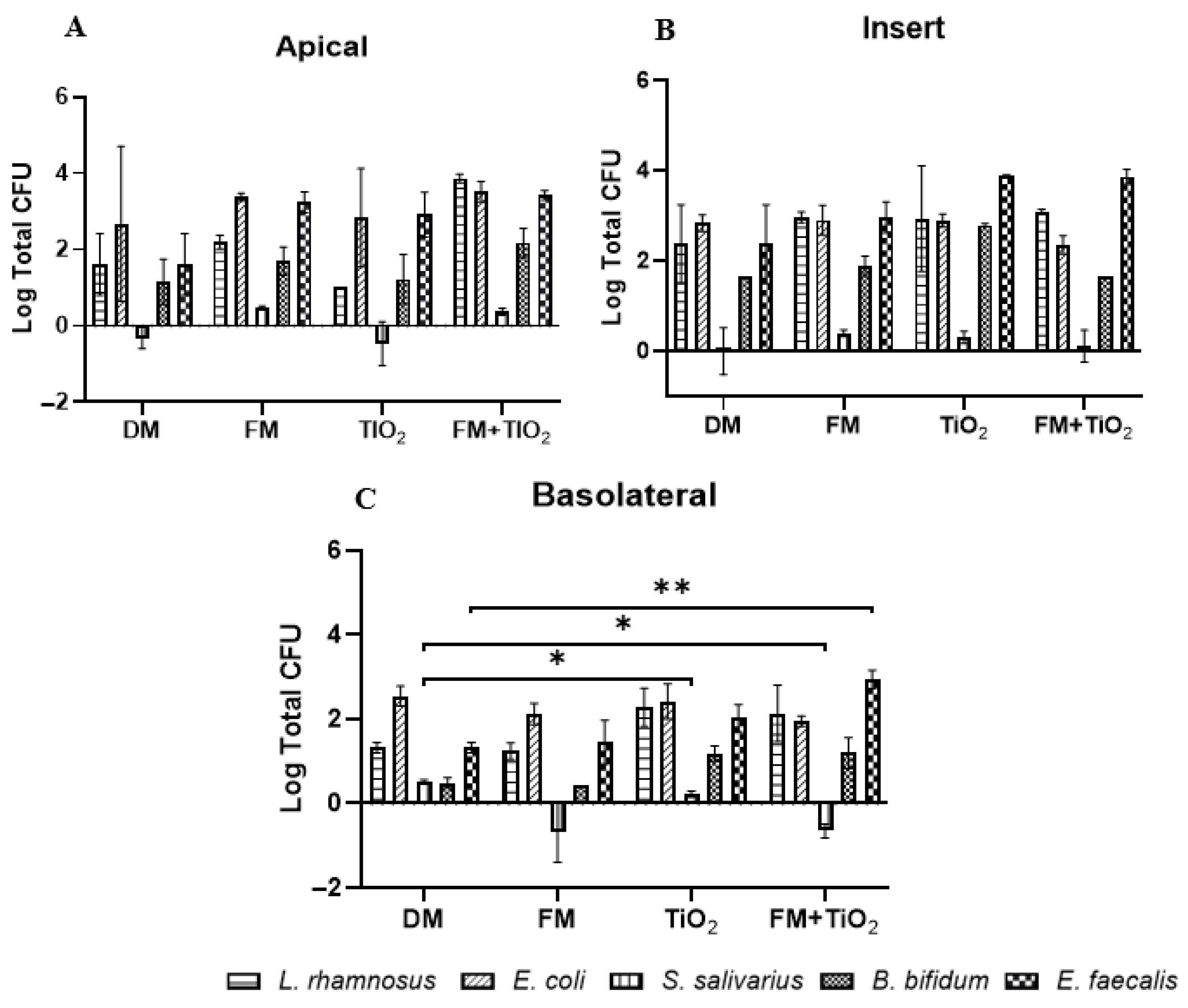
The duplication rate and the optimum growth conditions of bacteria differ from one another, but they still co-exist in the human gut. In this model, the individual bacterial populations do not cross the intestinal barrier, except for
E. coli and
S. salivarius (
Figure 6); however, when in the bacterial mock community, all bacterial species were present in the basolateral chamber (
Figure 7). Previously, a study published by Pinget et al. showed that TiO
2 causes a decrease in colonic mucin 2 expression, which causes a reduced mucus layer leading to inflammatory reactions and gut dysbiosis [
5,
46]. This is also observed in the current study, where
E. coli attachment on the insert, which can be interpreted as bacterial entrapment in the mucus layer in vivo, was reduced in the presence of TiO
2 (
Figure 6B). The acute exposure of the intestinal monolayer to both TiO
2 and a bacterial community could be a contributing factor to the increased presence of bacteria across the barrier.
S. salivarius is a facultative anaerobe whose presence was undetectable when cultured planktonically in the model due to its unique growth conditions. Since the Caco-2/HT29 experiments were conducted in a 5% CO
2 incubator at 37 °C, the growth conditions were not optimal for
S. salivarius. When cultured as part of the bacterial mock community, the
S. salivarius species was able to survive, potentially due to microbe–microbe or microbe–mucosal interactions within the model [
47].
S. salivarius migrated across the barrier when the monolayer was exposed to TiO
2, but this occurrence was not observed when TiO
2 was used as part of the food model, confirming the varying nature of NPs as a part of the food matrix. As a single species,
B. bifidum, a strict anaerobe with a slow duplication rate, was detected only on the insert and not in the apical or basolateral chamber. Its presence on the inserts confirms that the effects on permeability and transport we see in this study are in fact due to its ability to attach to the monolayer. As part of the mock community,
B. bifidum survival was enhanced, confirming the importance of using a bacterial mock community to get a more physiological representation. The attachment of all the bacterial species to the insert and their presence in the apical chamber confirm the successful incorporation of the bacterial community into the model.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






