
Dynamic In Vitro Models of the Human Gastrointestinal Tract as Relevant Tools to Assess the Survival of Probiotic Strains and Their Interactions with Gut Microbiota

Abstract
:1. Introduction
2. Experimental Section
2.1. Yeast Strain
2.2. Simulated Human Digestive Conditions
2.2.1. In Vitro Digestion in the TIM-1 System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| In Vitro Models | Main Parameters | |
|---|---|---|
| TIM-1 (gastric and smallintestinal model) |  |
|
| ARCOL (large intestinal model) |  |
|
2.2.2. In Vitro Fermentation in the ARCOL Model
| In Vitro Digestion Parameters | Fasted – “Glass of Water” Protocol | Fed – “Solid Meal” Protocol |
|---|---|---|
| pH |  | |
| Transit time |  | |
| Gastric compartment | ||
| 130 U/min of pepsin | 520 U/min of pepsin | |
| 5 U/min of lipase | 20 U/min of lipase | |
| 0.25 mL/min of HCl 0.3 M if necessary | 0.25 mL/min of HCl 1.5 M if necessary | |
| and min | and min | |
| Digestive secretions | Duodenal compartment | |
| 20 mg/min of bile extract during 30 min of digestion then 10 mg/min | ||
| 0.25 mg/min of intestinal electrolyte solution | ||
| 20 mg/min of pancreatin (4 USP) | 80 mg/min of pancreatin (4 USP) | |
| 0.25 mL/min of NaHCO3 0.5 M if necessary | 0.25 mL/min of NaHCO3 1 M if necessary | |
| Jejunal compartment | ||
| 0.25 mL/min of NaHCO3 0.5 M if necessary | 0.25 mL/min of NaHCO3 1 M if necessary | |
| Ileal compartment | ||
| 0.25 mL/min of NaHCO3 0.5 M if necessary | 0.25 mL/min of NaHCO3 1 M if necessary | |
| and min | and min | |
| Dialysis (Jejunum and ileum) | 10 mL/min | |
2.3. Yeast Survival in the in Vitro Gastrointestinal Tract
2.4. Composition and Metabolic Activity of Human Colonic Microbiota
| Name | Sequence 5’–3’ | Target | Annealing | References |
|---|---|---|---|---|
| Temperature (°C) | ||||
| SYBR green | ||||
| BAC338F | ACTCCTACGGGAGGCAG | Total bacteria | 58 | [19] |
| BAC338F | GTATTACCGCGGCTGCTG | |||
| 789cfbF | CRAACAGGATTAGATACCCT | Bacteroidetes | 61 | [20] |
| cfb967R | GGTAAGGTTCCTCGCGTAT | |||
| Act920F3 | TACGGCCGCAAGGCTA | Actinobacteria | 61 | [20] |
| Act1200R | TCRTCCCCACCTTCCTCCG | |||
| 928F-Firm | TGAAACTYAAAGGAATTGACG | Firmicutes | 61 | [20] |
| 1040FirmR | ACCATGCACCACCTGTC | |||
| Eco1457F | CATTGACGTTACCCGCAGAAGAAGC | Enterobacteriaceae | 63 | [21] |
| Eco1652R | CTCTACGAGACTCAAGCTTGC | |||
| F_Lacto05 | AGCAGTAGGGAATCTTCCA | Lactobacillus/Pediococcus/Leuconostoc | 60 | [22] |
| R_Lacto04 | CGCCACTGGTGTCTYTCCATATA | |||
| F_Fpra 428 | TGTAAACTCCTGTTGTTGAGGAAGATAA | Faecalibacterium prausnitzii | 60 | [23] |
| R_Fpra 583 | GCGCTCCCTTTACACCCA | |||
| TaqMan | ||||
| F_Bact 1369 | CGGTGAATACGTTCCCGG | Total bacteria | 60 | [22] |
| P_TM1389F | FAM-CTTGTACACACCGCCCGTC-TAMRA | |||
| R_Prok1492R | TACGGCTACCTTGTTACGACTT | |||
| E. coli-F | CATGCCGCGTGTATGAAGAA | Escherichia coli | 60 | [24] |
| E. coli-P | FAM-TATTAACTTTACTCCCTTCCTCCCCGCTGAA-TAMRA | |||
| E. coli-R | CGGGTAACGTCAATGAGCAAA | |||
| F_Bifid 09c | CGGGTGAGTAATGCGTGACC | Bifidobacteria | 60 | [22] |
| P_Bifid | FAM-CTCCTGGAAACGGGTG-TAMRA | |||
| R_Bifid 06 | TGATAGGACGCGACCCCA | |||
| F_Bacter 11 | CCTWCGATGGATAGGGGTT | Bacteroides/Prevotella | 60 | [22] |
| P_Bac303 | YY-AAGGTCCCCCACATTG-TAMRA | |||
| R_Bacter 08 | CACGCTACTTGGCTGGTTCAG | |||
2.5. Statistical Analysis
3. Results
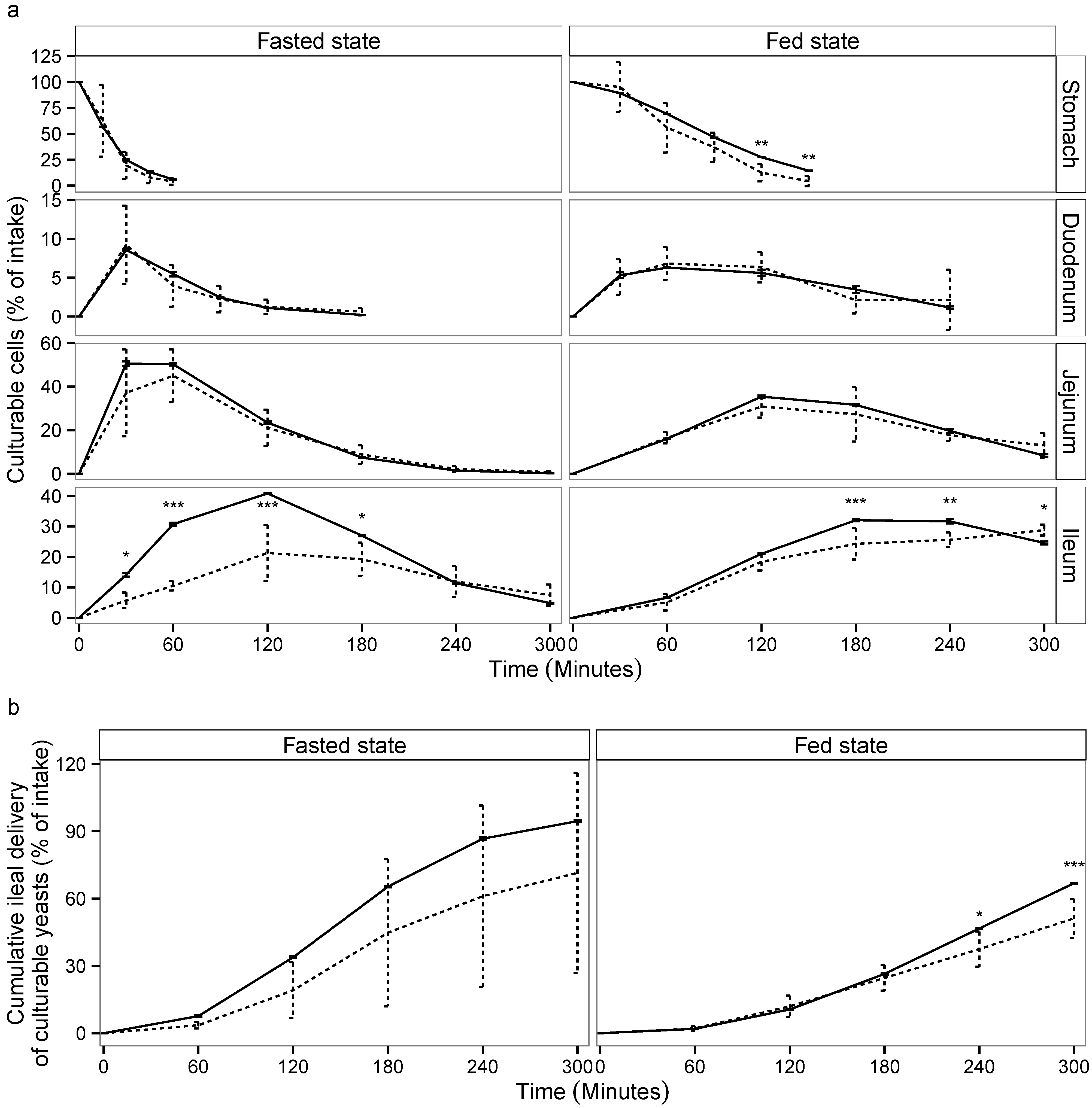
3.1. Yeast Viability in the Upper Gastro-Intestinal Tract


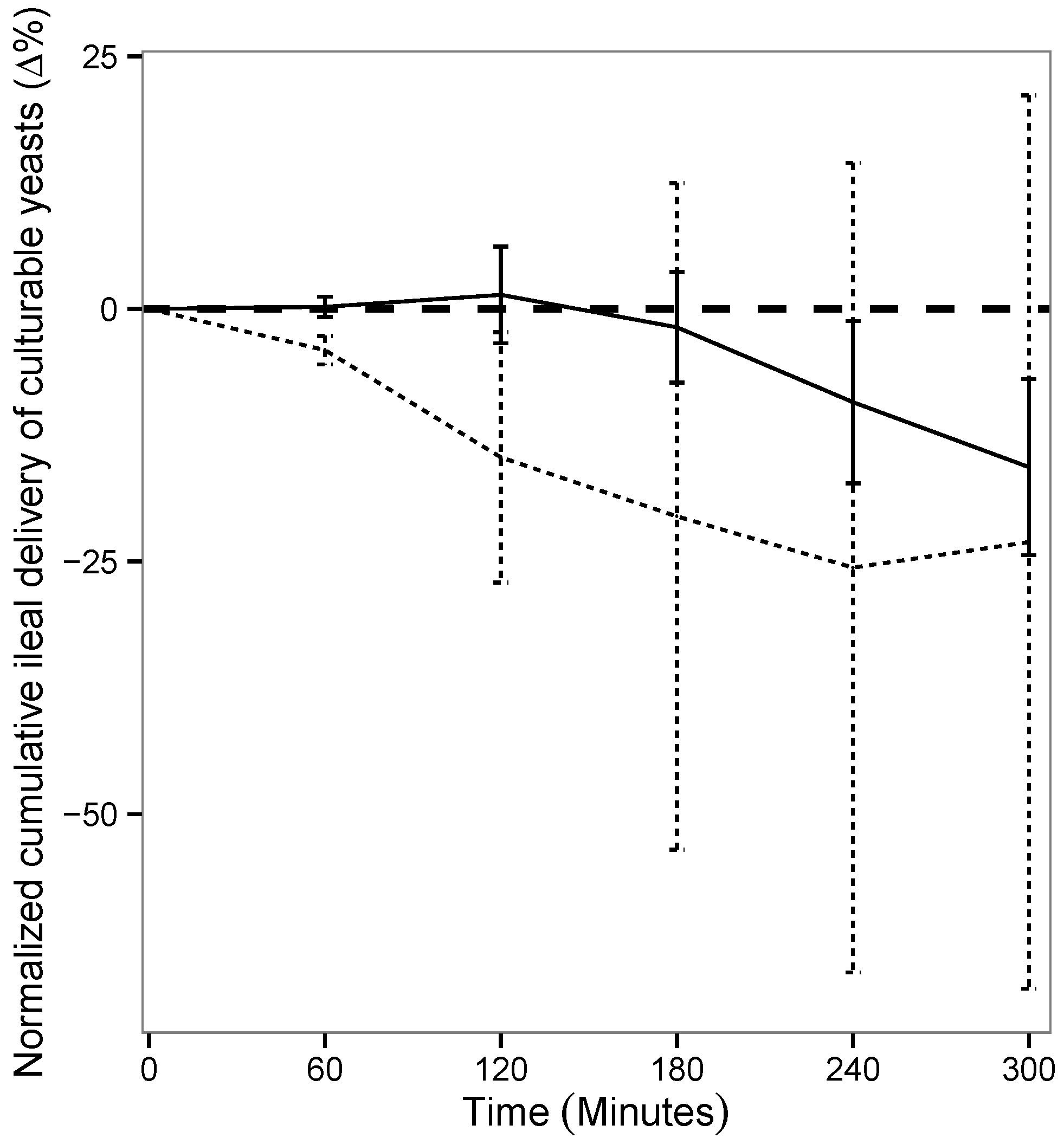
3.2. Yeast Viability in the Lower Gastro-Intestinal Tract

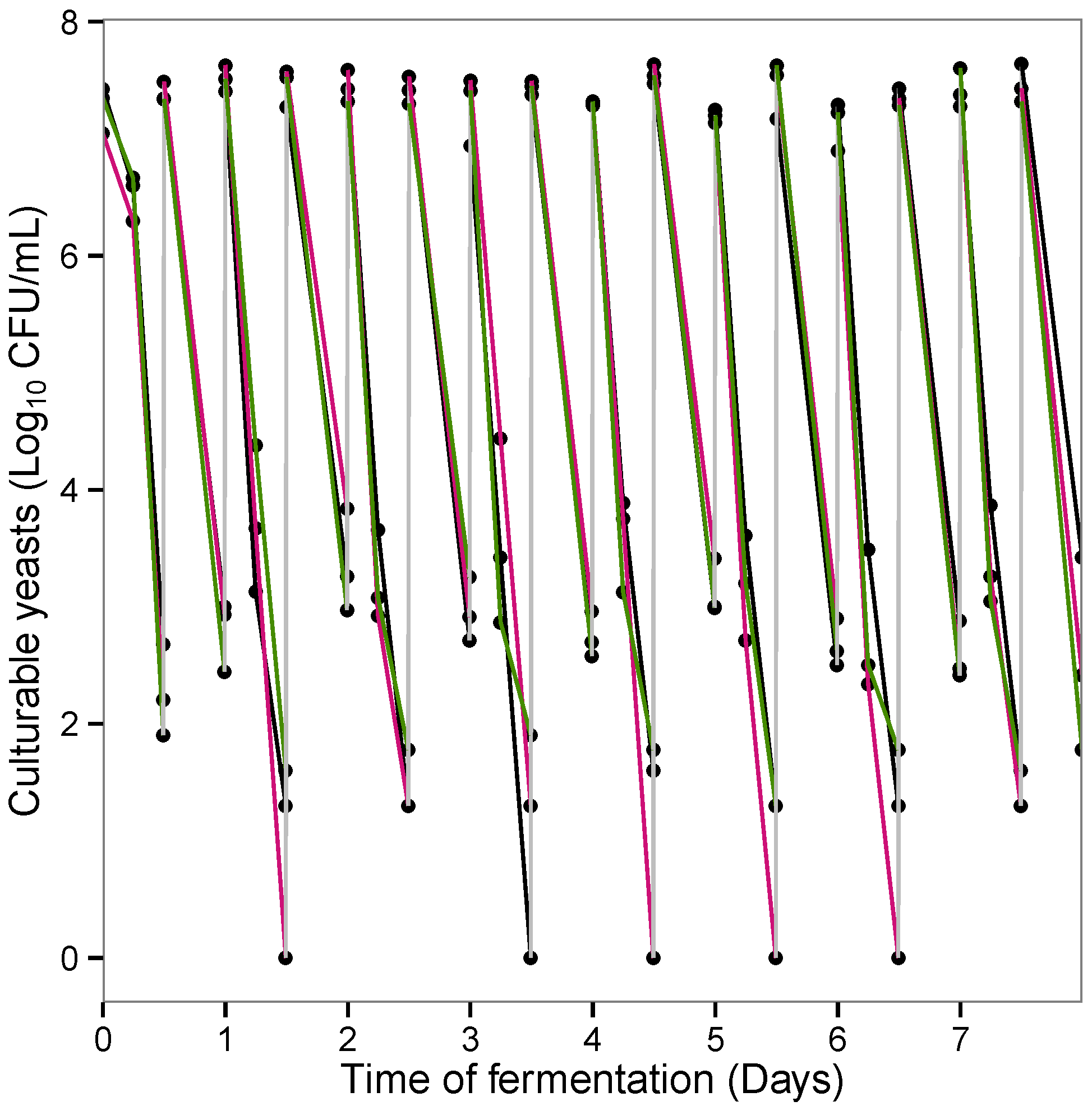
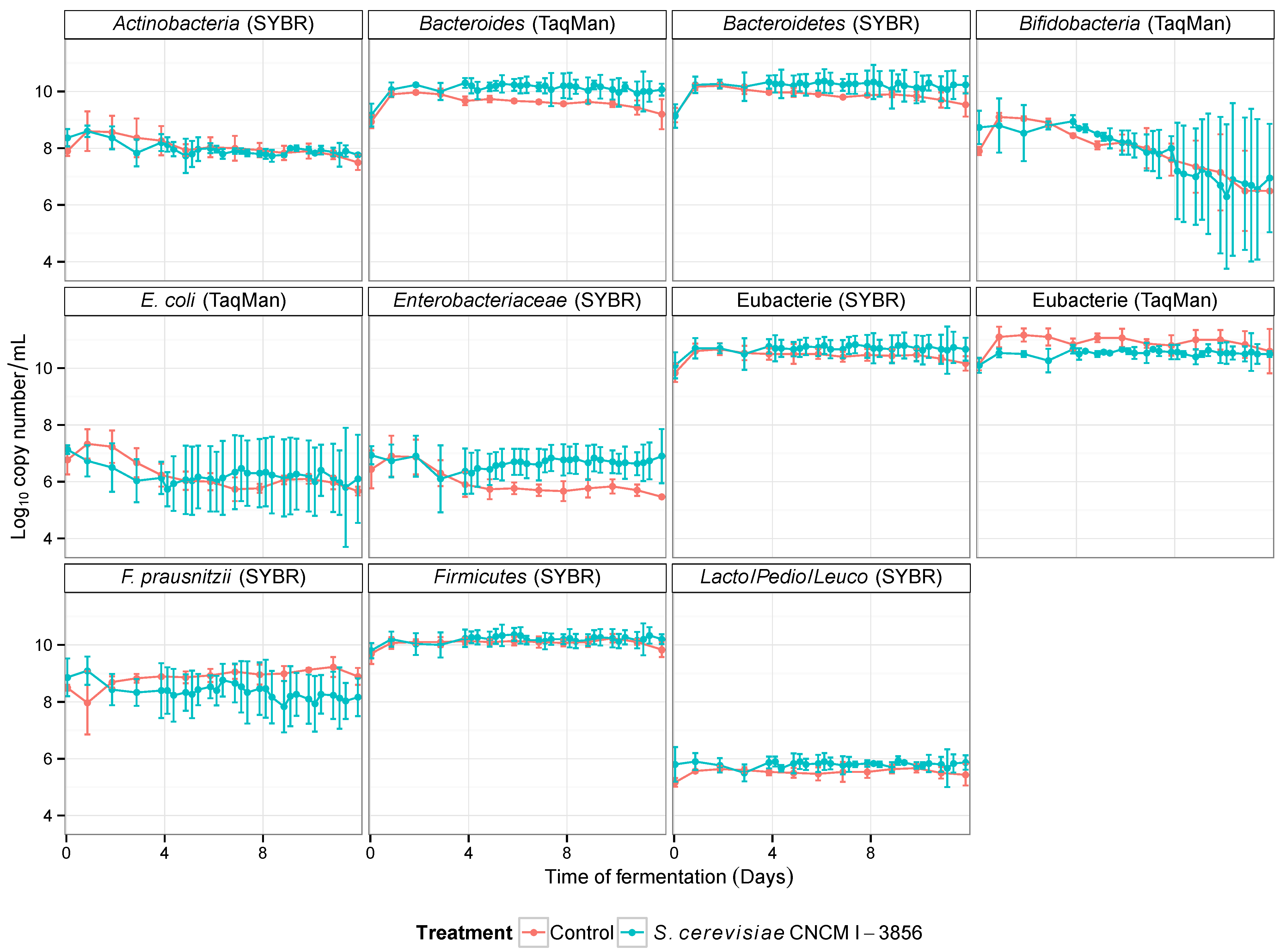
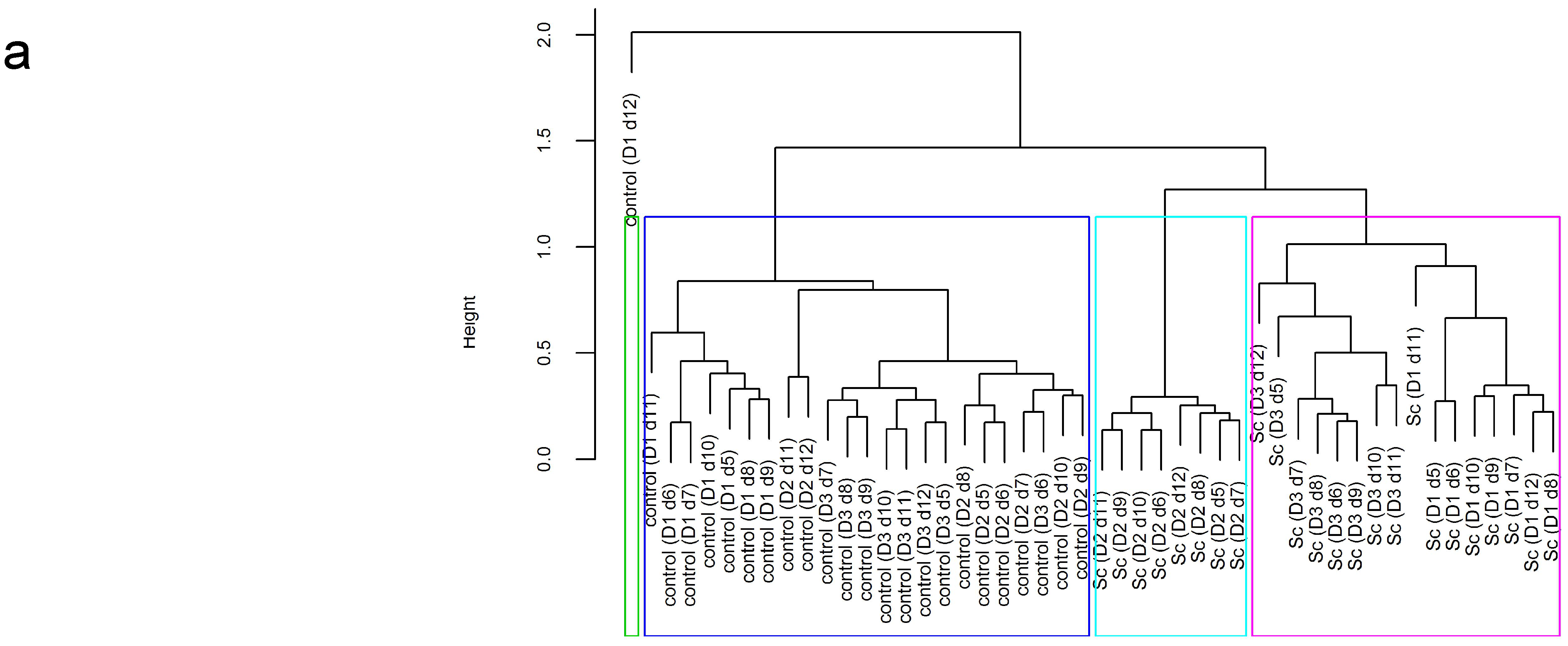
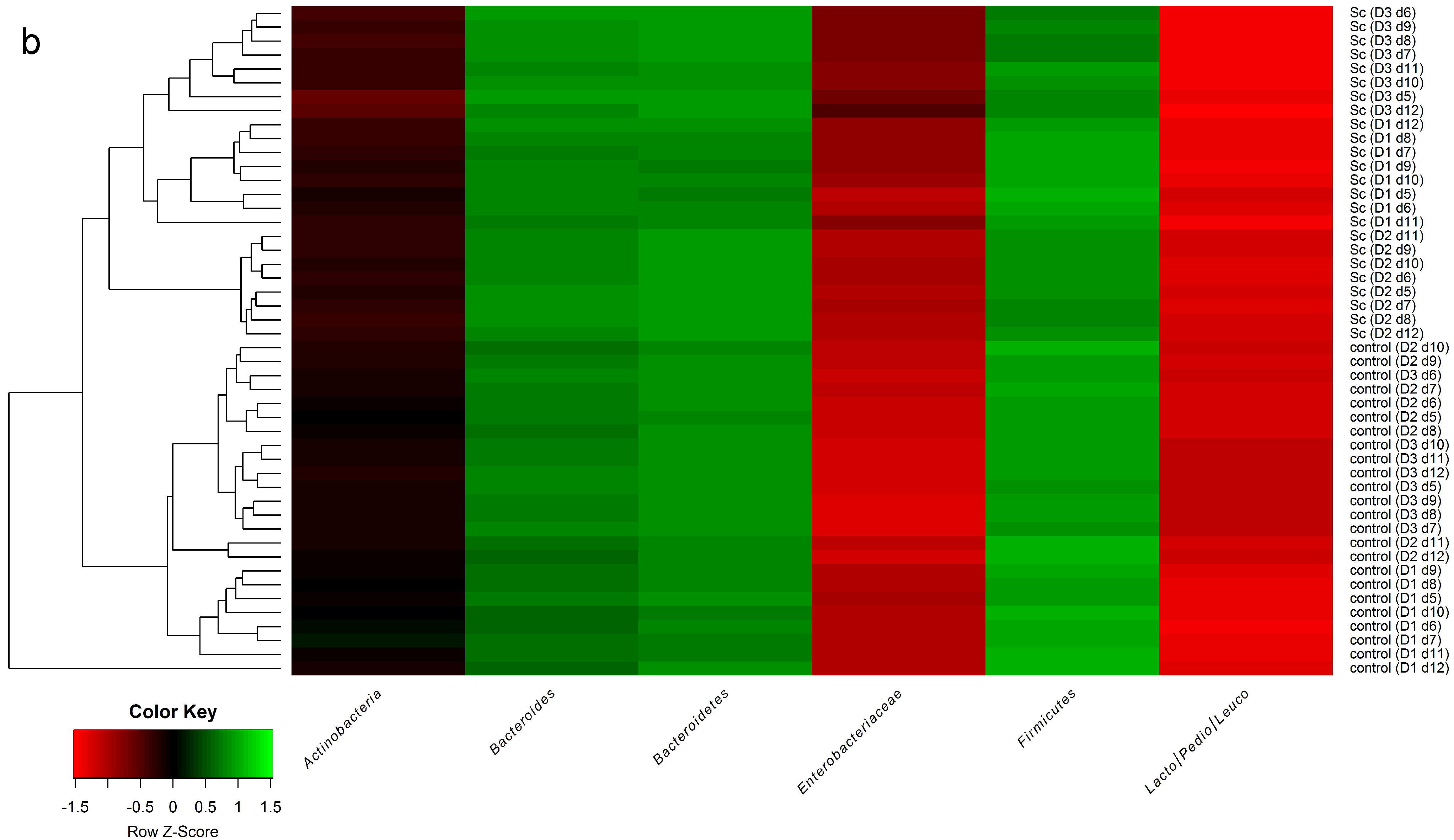
3.3. Influence of Probiotic Yeast on Human Colonic Microbiota Composition and Metabolic Activity



| SCFA | Control | S. cerevisiae CNCM I-3856 |
|---|---|---|
| Acetate | 67.2 ± 3.6 | 66.9 ± 3.0 |
| Propionate | 18.5 ± 2.6 | 18.7 ± 2.2 |
| Butyrate | 14.4 ± 2.0 | 14.4 ± 1.9 |
| iso-Butyrate | 1.9 ± 0.3 | 2.1 ± 0.2 |
| Valerate | 4.1 ± 1.0 | 4.8 ± 0.7 |
| iso-Valerate | 2.7 ± 0.4 | 3.0 ± 0.3 |
| Hexanoic acid | 2.5 ± 1.2 | 2.8 ± 1.7 |
| Heptanoic acid | 0.3 ± 0.3 | 0.6 ± 0.6 |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [PubMed]
- Hatoum, R.; Labrie, S.; Fliss, I. Antimicrobial and probiotic properties of yeasts: From fundamental to novel applications. Front. Microbiol. 2012, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, T.; Pothoulakis, C. Efficacy and safety of the probiotic Saccharomyces boulardii for the prevention and therapy of gastrointestinal disorders. Therap. Adv. Gastroenterol. 2012, 5, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Im, E.; Pothoulakis, C. Recent advances in Saccharomyces boulardii research. Gastroenterol. Clin. Biol. 2010, 34, S62–S70. [Google Scholar] [CrossRef]
- McFarland, L.V. Systematic review and meta-analysis of Saccharomyces boulardii in adult patients. World J. Gastroenterol. 2010, 16, 2202–2222. [Google Scholar] [CrossRef] [PubMed]
- Foligné, B.; Dewulf, J.; Vandekerckove, P.; Pignède, G.; Pot, B. Probiotic yeasts: Anti-inflammatory potential of various non-pathogenic strains in experimental colitis in mice. World J. Gastroenterol. 2010, 16, 2134–2145. [Google Scholar] [CrossRef] [PubMed]
- Sivignon, A.; de Vallée, A.; Barnich, N.; Denizot, J.; Darcha, C.; Pignède, G.; Vandekerckove, P.; Darfeuille-Michaud, A. Saccharomyces cerevisiae CNCM I-3856 prevents colitis induces by AIEC bacteria in transgenic mouse model mimicking Crohn’s disease. Inflamm. Bowel Dis. 2015, 21, 276–286. [Google Scholar] [CrossRef] [PubMed]
- De Chambrun, G.P.; Neut, C.; Chau, A.; Cazaubiel, M.; Pelerin, F.; Justen, P.; Desreumaux, P. A randomized clinical trial of Saccharomyces cerevisiae versus placebo in the irritable bowel syndrome. Dig. Liver Dis. 2015, 47, 119–1. [Google Scholar] [CrossRef] [PubMed]
- Zanello, G.; Berri, M.; Dupont, J.; Sizaret, P.Y.; D’Inca, R.; Salmon, H.; Meurens, F. Saccharomyces cerevisiae modulates immune gene expressions and inhibits ETEC-mediated ERK1/2 and p38 signaling pathways in intestinal epithelial cells. PLoS ONE 2011. [Google Scholar] [CrossRef] [PubMed]
- Etienne-Mesmin, L.; Livrelli, V.; Privat, M.; Denis, S.; Cardot, J.M.; Alric, M.; Blanquet-Diot, S. Effect of a new probiotic Saccharomyces cerevisiae strain on survival of Escherichia coli O157:H7 in a dynamic gastrointestinal model. Appl. Environ. Microb. 2011, 77, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Thévenot, J.; Cordonnier, C.; Rougeron, A.; Le Goff, O.; Nguyen, H.T.T.; Denis, S.; Alric, M.; Livrelli, V.; Blanquet-Diot, S. Enterohemorrhagic Escherichia coli infection has donor-dependent effect on human gut microbiota and may be antagonized by probiotic yeast during interaction with Peyer’s patches. Appl. Microbiol. Biotechnol. 2015, 99, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Thévenot, J.; Etienne-Mesmin, L.; Denis, S.; Chalancon, S.; Alric, M.; Livrelli, V.; Blanquet-Diot, S. Enterohemorrhagic Escherichia coli O157:H7 survival in an in vitro model of the human large intestine and interactions with probiotic yeasts and resident microbiota. Appl. Environ. Microbiol. 2013, 79, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Blanquet-Diot, S.; Denis, S.; Chalancon, S.; Chaira, F.; Cardot, J.M.; Alric, M. Use of artificial digestive systems to investigate the biopharmaceutical factors influencing the survival of probiotic yeast during gastrointestinal transit in humans. Pharm. Res. 2012, 29, 1444–1453. [Google Scholar] [CrossRef] [PubMed]
- Elashoff, J.D.; Reedy, T.J.; Meyer, J.H. Analysis of gastric emptying data. Gastroenterology 1982, 83, 1306–1312. [Google Scholar] [PubMed]
- Minekus, M. The TNO Gastro-Intestinal Model (TIM). In The Impact of Food Bioactives on Health; Springer: Doetinchem, The Netherlands, 2015; pp. 37–46. [Google Scholar]
- Minekus, M.; Marteau, P.; Havenaar, R.; Huis in’t Veld, J.H. A multi-compartmental dynamic computer-controlled model simulating the stomach and small intestine. ATLA 1995, 23, 197–209. [Google Scholar]
- Macfarlane, G.; Macfarlane, S.; Gibson, G. Validation of a three-stage compound continuous culture system for investigating the effect of retention time on the ecology and metabolism of bacteria in the human colon. Microb. Ecol. 1998, 35, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–813. [Google Scholar] [PubMed]
- Yu, Y.; Lee, C.; Kim, J.; Hwang, S. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol. Bioeng. 2005, 89, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Bacchetti De Gregoris, T.; Aldred, N.; Clare, A.S.; Burgess, J.G. Improvement of phylum-and class-specific primers for real-time PCR quantification of bacterial taxa. J. Microbiol. Meth. 2011, 86, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Bartosch, S.; Fite, A.; Macfarlane, G.T.; McMurdo, M.E. Characterization of bacterial communities in feces from healthy elderly volunteers and hospitalized elderly patients by using real-time PCR and effects of antibiotic treatment on the fecal microbiota. Appl. Environ. Microb. 2004, 70, 3575–3581. [Google Scholar] [CrossRef] [PubMed]
- Furet, J.P.; Firmesse, O.; Gourmelon, M.; Bridonneau, C.; Tap, J.; Mondot, S.; Doré, J.; Corthier, G. Comparative assessment of human and farm animal faecal microbiota using real-time quantitative PCR. FEMS Microbiol. Ecol. 2009, 68, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Medina, M.; Denizot, J.; Dreux, N.; Robin, F.; Billard, E.; Bonnet, R.; Darfeuille-Michaud, A.; Barnich, N. Western diet induces dysbiosis with increased E. coli in CEABAC10 mice, alters host barrier function favouring AIEC colonisation. Gut 2014, 63, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Huijsdens, X.W.; Linskens, R.K.; Mak, M.; Meuwissen, S.G.; Vandenbroucke-Grauls, C.M.; Savelkoul, P.H. Quantification of bacteria adherent to gastrointestinal mucosa by real-time PCR. J. Clin. Microbiol. 2002, 40, 4423–4427. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, K.; Gel, Y.R.; Brunner, E.; Konietschke, F. nparLD: An R software package for the nonparametric analysis of longitudinal data in factorial experiments. J. Stat. Softw. 2012, 50, 1–23. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Konietschke, F.; Placzek, M.; Schaarschmidt, F.; Hothorn, L.A. nparcomp: An R software package for nonparametric multiple comparisons and simultaneous confidence intervals. J. Stat. Softw. 2015, 64, 1–17. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Bruun Brockhoff, P.; Haubo Bojesen Christensen, R. lmerTest: Tests in Linear Mixed Effects Models, 2014; R package version 2.0-20. Avaliable online: https://cran.r-project.org/web/packages/lmerTest/index.html (accessed on 25 September 2015).
- Guerra, A.; Etienne-Mesmin, L.; Livrelli, V.; Denis, S.; Blanquet-Diot, S.; Alric, M. Relevance and challenges in modeling human gastric and small intestinal digestion. Trends Biotechnol. 2012, 30, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.N.; Zihler, A.; Chassard, C.; Lacroix, C. Advances and perspectives in in vitro human gut fermentation modeling. Trends Biotechnol. 2012, 30, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Do Espirito Santo, A.P.; Perego, P.; Converti, A.; Oliveira, M.N. Influence of food matrices on probiotic viability—A review focusing on the fruity bases. Trends Food Sci. Technol. 2011, 22, 377–385. [Google Scholar] [CrossRef]
- Possemiers, S.; Marzorati, M.; Verstraete, W.; van de Wiele, T. Bacteria and chocolate: a successful combination for probiotic delivery. Int. J. Food Microbiol. 2010, 141, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Graff, S.; Chaumeil, J.C.; Boy, P.; Lai-Kuen, R.; Charrueau, C. Formulations for protecting the probiotic Saccharomyces boulardii from degradation in acidic condition. Biol. Pharm. Bull. 2008, 31, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Ghelardi, E.; Celandroni, F.; Salvetti, S.; Gueye, S.; Lupetti, A.; Senesi, S. Survival and persistence of Bacillus clausii in the human gastrointestinal tract following oral administration as spore-based probiotic formulation. J. Appl. Microbiol. 2015, 119, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Cheow, W.S.; Kiew, T.Y.; Hadinoto, K. Controlled release of Lactobacillus rhamnosus biofilm probiotics from alginate-locust bean gum microcapsules. Carbohydr. Polym. 2014, 103, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Koziolek, M.; Grimm, M.; Becker, D.; Iordanov, V.; Zou, H.; Shimizu, J.; Wanke, C.; Garbacz, G.; Weitschies, W. Investigation of pH and temperature profiles in the GI tract of fasted human subjects using the Intellicap® system. J. Pharm. Sci. 2015, 104, 2855–2863. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.; Eddy, D.E.; Bartoli, W.P.; Paul, G.L. Gastric emptying of water and isocaloric carbohydrate solutions consumed at rest. Med. Sci. Sports Exerc. 1994, 26, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Worsøe, J.; Fynne, L.; Gregersen, T.; Schlageter, V.; Christensen, L.A.; Dahlerup, J.F.; Rijkhoff, N.J.; Laurberg, S.; Krogh, K. Gastric transit and small intestinal transit time and motility assessed by a magnet tracking system. BMC Gastroenterol. 2011, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, G. Pancreatic secretory response to ordinary meals: Studies with pure pancreatic juice. Gastroenterology 1988, 94, 428–433. [Google Scholar]
- Northfield, T.; McColl, I. Postprandial concentrations of free and conjugated bile acids down the length of the normal human small intestine. Gut 1973, 14, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Vertzoni, M.; Archontaki, H.; Reppas, C. Determination of intralumenal individual bile acids by HPLC with charged aerosol detection. J. Lipid Res. 2008, 49, 2690–2695. [Google Scholar] [CrossRef] [PubMed]
- Elmer, G.; McFarland, L.; Surawicz, C.; Danko, L.; Greenberg, R. Behaviour of Saccharomyces boulardii in recurrent Clostridium difficile disease patients. Aliment. Pharmacol. Ther. 1999, 13, 1663–1668. [Google Scholar] [CrossRef] [PubMed]
- Minelli, E.B.; Benini, A. Relationship between number of bacteria and their probiotic effects. Microb. Ecol. Health Dis. 2008, 20, 180–183. [Google Scholar] [CrossRef]
- Klein, S.M.; Elmer, G.W.; McFarland, L.V.; Surawicz, C.M.; Levy, R.H. Recovery and elimination of the biotherapeutic agent, Saccharomyces boulardii, in healthy human volunteers. Pharm. Res. 1993, 10, 1615–1619. [Google Scholar] [CrossRef] [PubMed]
- Pecquet, S.; Guillaumin, D.; Tancrede, C.; Andremont, A. Kinetics of Saccharomyces cerevisiae elimination from the intestines of human volunteers and effect of this yeast on resistance to microbial colonization in gnotobiotic mice. Appl. Environ. Microbiol. 1991, 57, 3049–3051. [Google Scholar] [PubMed]
- Fietto, J.L.; Araújo, R.S.; Valadão, F.N.; Fietto, L.G.; Brandão, R.L.; Neves, M.J.; Gomes, F.C.; Nicoli, J.R.; Castro, I.M. Molecular and physiological comparisons between Saccharomyces cerevisiae and Saccharomyces boulardii. Can. J. Microbiol. 2004, 50, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Graff, S.; Chaumeil, J.C.; Boy, P.; Lai-Kuen, R.; Charrueau, C. Influence of pH conditions on the viability of Saccharomyces boulardii yeast. J. Gen. Appl. Microbiol. 2008, 54, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Loening-Baucke, V.; Kirsch, S.; Doerffel, Y. Functional biostructure of colonic microbiota (central fermenting area, germinal stock area and separating mucus layer) in healthy subjects and patients with diarrhea treated with Saccharomyces boulardii. Gastroenterol. Clin. Biol. 2010, 34, S79–S92. [Google Scholar] [CrossRef]
- Vanhoutte, T.; de Preter, V.; de Brandt, E.; Verbeke, K.; Swings, J.; Huys, G. Molecular monitoring of the fecal microbiota of healthy human subjects during administration of lactulose and Saccharomyces boulardii. Appl. Environ. Microbiol. 2006, 72, 5990–5997. [Google Scholar] [CrossRef] [PubMed]
- Bolnick, D.I.; Snowberg, L.K.; Hirsch, P.E.; Lauber, C.L.; Parks, B.; Lusis, A.J.; Knight, R.; Caporaso, J.G.; Svanbäck, R. Individual diet has sex-dependent effects on vertebrate gut microbiota. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.M.; Girard-Pipau, F.; Filippi, J.; Hébuterne, X.; Moyse, D.; Hinojosa, G.C.; Pompei, A.; Rampal, P. Effects of Saccharomyces boulardii on fecal short-chain fatty acids and microflora in patients on long-term total enteral nutrition. World J. Gastroenterol. 2005, 11, 6165–6169. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Matamoros, S.; Geurts, L.; Delzenne, N.M.; Cani, P.D. Saccharomyces boulardii administration changes gut microbiota and reduces hepatic steatosis, low-grade inflammation, and fat mass in obese and type 2 diabetic db/db mice. MBio 2014, 5, e01011–e01014. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordonnier, C.; Thévenot, J.; Etienne-Mesmin, L.; Denis, S.; Alric, M.; Livrelli, V.; Blanquet-Diot, S. Dynamic In Vitro Models of the Human Gastrointestinal Tract as Relevant Tools to Assess the Survival of Probiotic Strains and Their Interactions with Gut Microbiota. Microorganisms 2015, 3, 725-745. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3040725
Cordonnier C, Thévenot J, Etienne-Mesmin L, Denis S, Alric M, Livrelli V, Blanquet-Diot S. Dynamic In Vitro Models of the Human Gastrointestinal Tract as Relevant Tools to Assess the Survival of Probiotic Strains and Their Interactions with Gut Microbiota. Microorganisms. 2015; 3(4):725-745. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3040725
Chicago/Turabian StyleCordonnier, Charlotte, Jonathan Thévenot, Lucie Etienne-Mesmin, Sylvain Denis, Monique Alric, Valérie Livrelli, and Stéphanie Blanquet-Diot. 2015. "Dynamic In Vitro Models of the Human Gastrointestinal Tract as Relevant Tools to Assess the Survival of Probiotic Strains and Their Interactions with Gut Microbiota" Microorganisms 3, no. 4: 725-745. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3040725