Biosynthesis of Polyketides in Streptomyces

1
Microbial Strain Collection (MISG), Helmholtz Centre for Infection Research (HZI), 38124 Braunschweig, Germany
2
Research Unit for Clean Technology, Indonesian Institute of Sciences (LIPI), Bandung 40135, Indonesia
3
Research Center for Chemistry, Indonesian Institute of Sciences (LIPI), Serpong 15314, Indonesia
*
Authors to whom correspondence should be addressed.
Microorganisms 2019, 7(5), 124; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7050124
Submission received: 18 March 2019
/
Revised: 24 April 2019
/
Accepted: 27 April 2019
/
Published: 6 May 2019
(This article belongs to the Special Issue Actinobacteria and Myxobacteria—Important Resources for Novel Antibiotics)
Abstract
:Polyketides are a large group of secondary metabolites that have notable variety in their structure and function. Polyketides exhibit a wide range of bioactivities such as antibacterial, antifungal, anticancer, antiviral, immune-suppressing, anti-cholesterol, and anti-inflammatory activity. Naturally, they are found in bacteria, fungi, plants, protists, insects, mollusks, and sponges. Streptomyces is a genus of Gram-positive bacteria that has a filamentous form like fungi. This genus is best known as one of the polyketides producers. Some examples of polyketides produced by Streptomyces are rapamycin, oleandomycin, actinorhodin, daunorubicin, and caprazamycin. Biosynthesis of polyketides involves a group of enzyme activities called polyketide synthases (PKSs). There are three types of PKSs (type I, type II, and type III) in Streptomyces responsible for producing polyketides. This paper focuses on the biosynthesis of polyketides in Streptomyces with three structurally-different types of PKSs.
1. Introduction
Polyketides, a large group of secondary metabolites, are known to possess remarkable variety, not only in their structure, and but also in their function [1,2]. Polyketides exhibit a wide range of bioactivities such as antibacterial (e.g., tetracycline), antifungal (e.g., amphotericin B), anticancer (e.g., doxorubicin), antiviral (e.g., balticolid), immune-suppressing (e.g., rapamycin), anti-cholesterol (e.g., lovastatin), and anti-inflammatory activity (e.g., flavonoids) [3,4,5,6,7,8,9]. Some organisms can produce polyketides such as bacteria (e.g., tetracycline from Streptomyces aureofaciens) [10], fungi (e.g., lovastatin from Phomopsis vexans) [11], plants (e.g., emodin from Rheum palmatum) [12], protists (e.g., maitotoxin-1 from Gambierdiscus australes) [13], insects (e.g., stegobinone from Stegobium paniceum) [14], and mollusks (e.g., elysione from Elysia viridis) [15]. These organisms could use the polyketides they produce as protective compounds and for pheromonal communication in the case for insects.
Since the beginning of the 1940s, the history of antibiotics has greatly related to microorganisms. One of the groups of bacteria that produce many important antibiotics is Actinobacteria. Actinobacteria are Gram-positive, have high GC content, and comprise various genera known for their secondary metabolite production, such as Streptomyces, Micromonospora, Kitasatospora, Nocardiopsis, Pseudonocardia, Nocardia, Actinoplanes, Saccharopolyspora, and Amycolatopsis [16,17]. Their most important genus is Streptomyces, which has a filamentous form like fungi and has become a source of around two-thirds of all known natural antibiotics [18]. Among the antibiotics produced by Streptomyces, polyketides are one group of the very important compounds. Some examples of polyketides produced by Streptomyces are rapamycin (produced by Streptomyces hygroscopicus), oleandomycin (produced by Streptomyces antibioticus), actinorhodin (produced by Streptomyces coelicolorA3(2)), daunorubicin (produced by Streptomyces peucetius) and caprazamycin (produced by Streptomyces sp. MK730-62F2) [19,20,21,22,23].
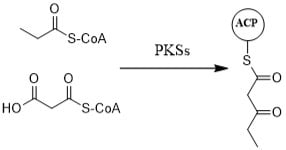
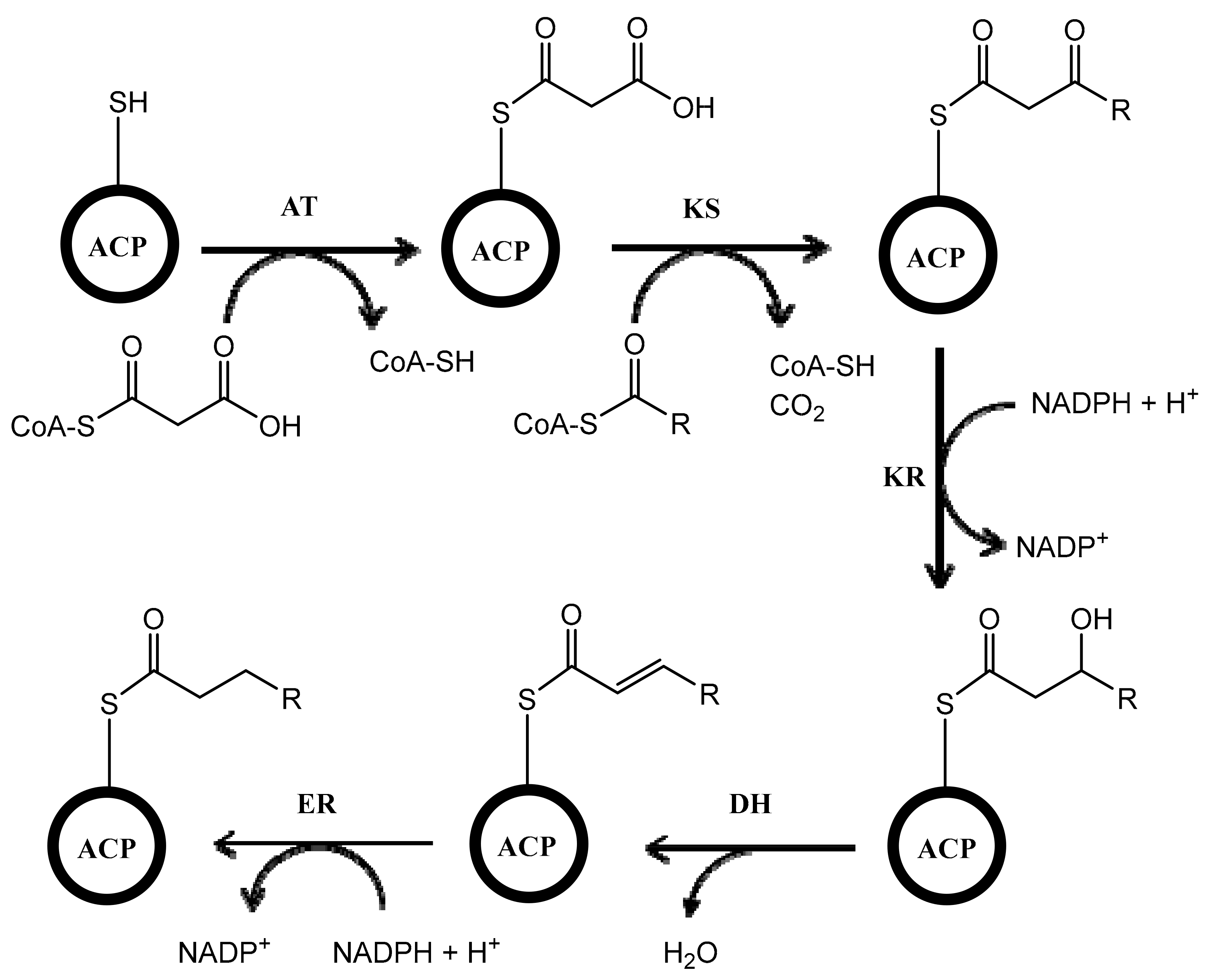
Biosynthesis of polyketides is very complex because the process involves multifunctional enzymes called polyketide synthases (PKSs). The mechanism of PKS is similar to fatty acid synthase (FAS). The process includes many enzymatic reactions with different enzymes such as acyltransferase (AT), which has a role in catalyzing the attachment of the substrate (e.g., acetyl or malonyl) to the acyl carrier protein (ACP), and ketosynthase (KS), which catalyzes the condensation of substrates attached in ACP. After condensation of the substrates, the reaction continues by incorporating ketoreductase (KR), which reduces keto ester, dehydratase (DH), which dehydrates the compound, and enoylreductase (ER), which reduces the carbon-carbon double bond in the molecule (Figure 1). Unlike in FAS, the process catalyzed by KR, DH, and ER is optional in PKSs, which can give the various structures of polyketides with keto groups, hydroxy groups, and/or double bonds in different locations of the molecule [24,25,26]. In Streptomyces, there are three types of PKSs (type I, type II, and type III) [27,28,29]. This review describes the biosynthesis of polyketides in Streptomyces with three distinct types of PKSs. The focus is only on the Streptomyces genus because it is one of the most important producers of bioactive compounds and one of the most well-studied microbes in terms of polyketide biosynthesis. To the best of our knowledge, this is the first review that describes the three types of PKSs that are involved in the biosynthesis of polyketides in Streptomyces.
2. Polyketide Synthases Type I
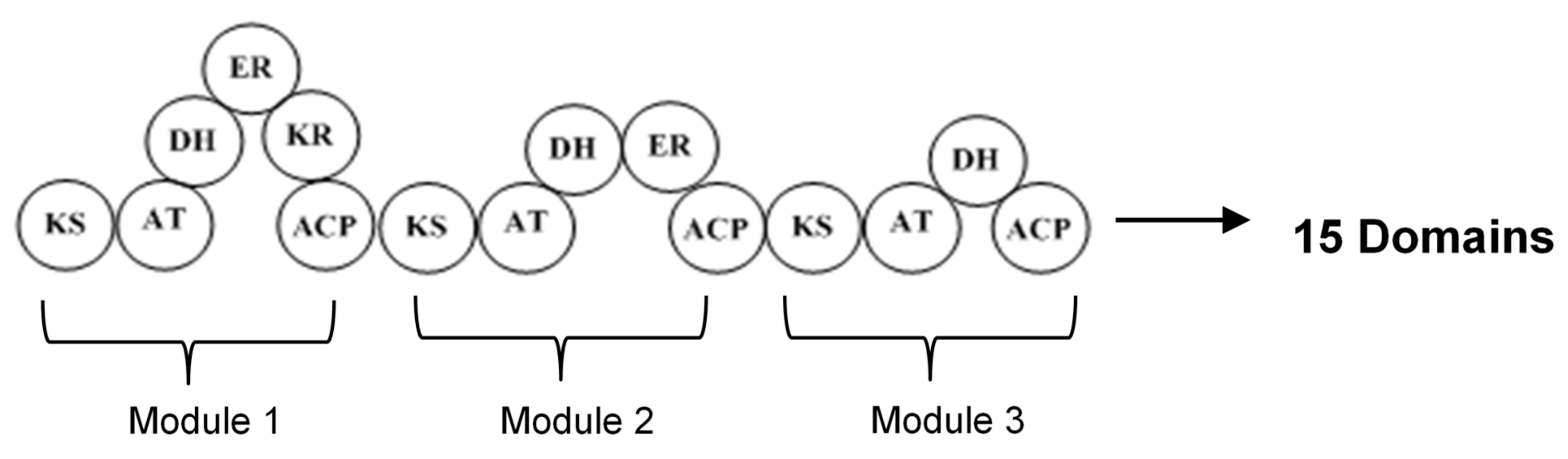
The type I polyketide synthases (type I PKSs) involve huge multifunctional proteins that have many modules containing some domains, in which a particular enzymatic reaction occurs (Figure 2). Each module has the responsibility of performing one condensation cycle in a non-iterative way. Because this system works with some modules, it is also called modular PKS. The essential domains existing in each module are acyltransferase (AT), keto synthase (KS), and acyl carrier protein (ACP) which collaborate to produce β-keto ester intermediate. In addition, the other domains that may be present in the module are β-ketoreductase (KR), dehydratase (DH), and enoyl reductase (ER), which are responsible for keto group modification. In the process of producing polyketide, the expanding polyketide chain is transferred from one module to another module until the completed molecule is liberated from the last module by a special enzyme [2,26,31].
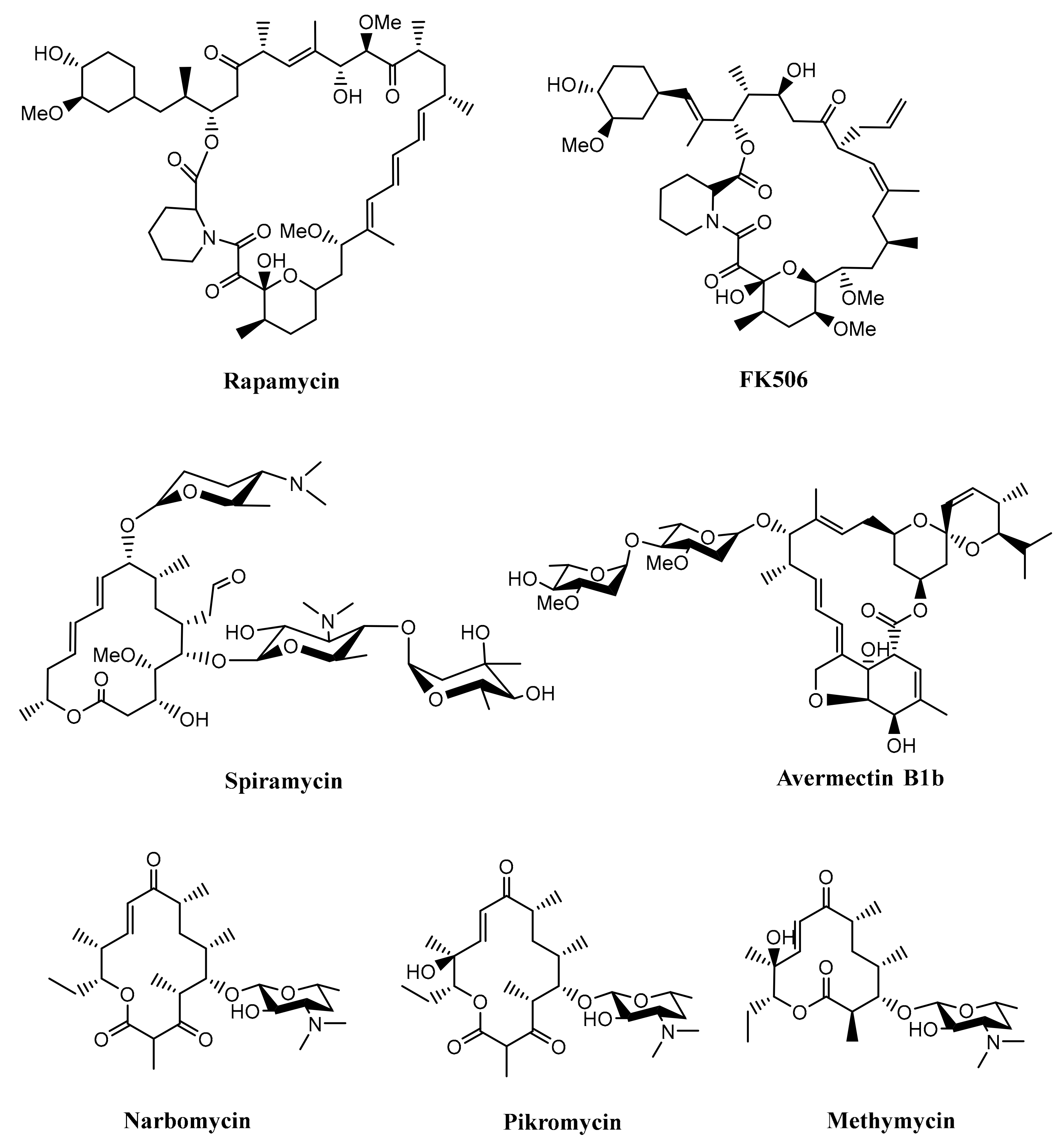
Furthermore, type I PKSs are generally responsible for producing macrocyclic polyketides (macrolides), although there was also a study reporting that type I PKSs are also involved in the biosynthesis of linear polyketide tautomycetin [32]. Macrolide belongs to a polyketide compound characterized by a macrocyclic lactone ring, which has various bioactivities such as antibacterial, antifungal, immunosuppressing, and anticancer. As an antibacterial agent, macrolide works by inhibiting protein synthesis by binding to the 50S ribosomal subunit and blocking the translocation steps of protein synthesis [8,27,33]. Some examples of macrolides produced by Streptomyces are rapamycin, FK506, spiramycin, avermectin, methymycin, narbomycin, and pikromycin, as shown in Figure 3 [34,35,36,37]. These compounds were produced by multifunctional polypeptides encoded by a biosynthetic gene cluster. The list of some polyketides produced by Streptomyces with their huge multifunctional proteins can be seen in Table 1.
2.1. Biosynthesis of Rapamycin
Rapamycin is a 31-membered ring macrolide produced by Streptomyces hygroscopicus isolated firstly from the soil of Easter Island (Chile) in the South Pacific Ocean. It is a hydrophobic compound and known as an antifungal compound against Candida albicans, Cryptococcus neoformans, Aspergillus fumigatus, Fusarium oxysporum, and some pathogenic species from the genus Penicillium. The antifungal mechanism of this compound has been described by diffusing into the cell and binding to intracellular receptor immunophilin FKB12. The FKBP12-rapamycin complexes inhibit enzymes required for signal transduction and cell growth. These enzymes are TOR (target of rapamycin) kinases that are conserved and very important for cell cycle progression. Interestingly, it was also reported that rapamycin has not only antifungal activity, but also anticancer and immunosuppressant activity [8,27,38,39].
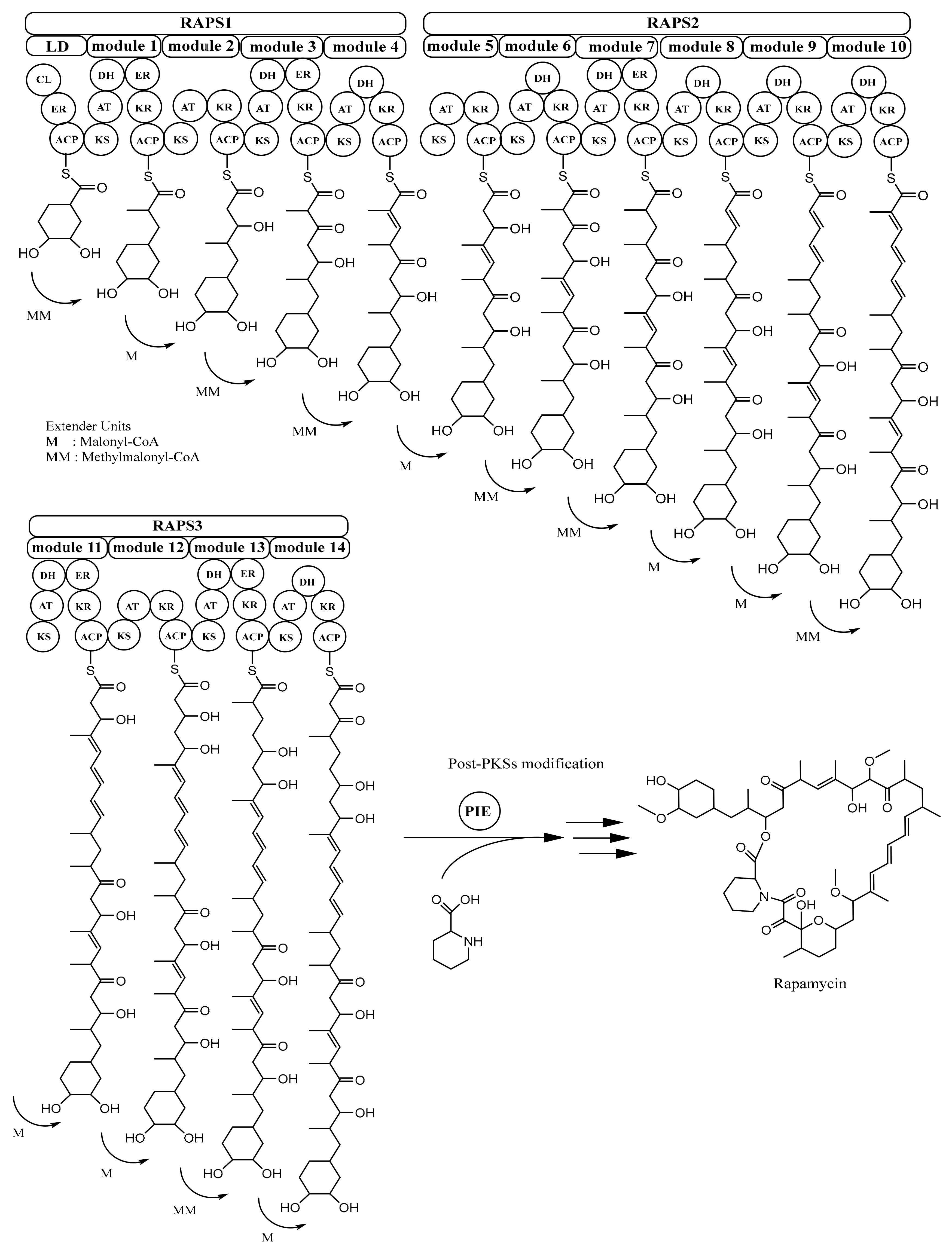
Rapamycin is synthesized by type I PKS rapamycin synthase (RAPS) [40]. The rapamycin-PKS gene cluster (rapPKS) is 107.3 kb in size and has three remarkably large ORFs (open reading frames), rapA, rapB, and rapC which encode multifunctional protein RAPS1 (~900 kDa), RAPS2 (~1.07 MDa), and RAPS3 (~660 kDa), respectively. Protein RAPS1 comprises four modules for polyketide chain extension; protein RAPS2 contains six modules responsible for continuing the process of polyketide chain elongation until C-16; and RAPS3 possesses four modules that have a role in completing the polyketide fraction of the rapamycin molecule. Overall, these three giant proteins encompass 70 domains or enzymatic functions, and because of this, rapamycin PKSs are considered as the most complex multienzyme system discovered so far [26,27,34].
In rapamycin PKSs, there is a loading domain (LD) before module 1. In LD, there are three domains, i.e., coenzyme A ligase (CL), enoylreductase (ER), and acyl carrier protein (ACP) domain, which are considered to play a role in activating, reducing a free shikimic-acid-derived moiety starter unit, and finally passing it to the ketosynthase (KS) domain of the first module, respectively. The extender units required for producing rapamycin are malonyl-CoA and methylmalonyl-CoA. The mechanism of transferring from the last domain in rapamycin PKSs and cyclisation of polyketide molecule is assisted by pipecolate-incorporating enzyme (PIE), as depicted in Figure 4. This enzyme (170 kDa) is encoded by gene rapP, which is also located in the rapPKS gene cluster [26,27,34].
2.2. Biosynthesis of Avermectin
Avermectin is a 16-membered ring macrolide and one of the notable anthelmintic compounds produced by Streptomyces avermitilis [41,49]. The biosynthesis of avermectin involves type I PKSs (AVES1, AVES2, AVES3, and AVES4). AVES1 (414 kDa) contains one loading domain and two modules; AVES2 (666 kDa) consists of four modules; AVES 3 (575 kDa) comprises three modules; and AVES4 (510 kDa) has three modules. The process of avermectin biosynthesis includes assembling of the polyketide-derived initial aglycon (6, 8a-seco-6, 8a-deoxy-5-oxoavermectin aglycons) by AVES1–4, alteration of the initial aglycon to avermectin aglycons, and, as the last step, the glycosylation of avermectin aglycons to produce avermectins. The starter unit for avermectin biosynthesis is isobutyryl-CoA (derived from valine) or 2-methylbutyryl-CoA (derived from isoleucine), whereas the extender units involved in the production of avermectin are seven malonyl-CoAs (for acetate units) and five methylmalonyl-CoAs (for propionate units). The nucleotide sequence of the avermectin biosynthetic gene cluster comprises 18 ORFs spanning a distance of 82 kb, in which four large ORFs encode the avermectin polyketide synthase (AVES1, AVES2, AVES3, and AVES4) and some of the 14 ORFs encode polypeptides having important roles in avermectin biosynthesis [41].
2.3. Biosynthesis of Candicidin
Candicidin is a 38-membered ring polyene macrolide produced by Streptomyces griseus IMRU 3570 that has antifungal activity. Like the other polyene compounds, the antifungal mechanism of candicidin is also by disrupting the fungal cell membrane. Candicidin has both the amino sugar mycosamine and the aromatic component p-aminoacetophenone in its macrolide structure [43,50].
The candicidin biosynthetic gene cluster (<205 kb) was cloned and partially sequenced. Four genes, canP1, canP3, canP2 (incomplete), and canPF (incomplete), were determined as genes encoding parts of type I PKSs (CanP1, CanP2, CanP3, and CanPF). CanP1 contains one loading domain and one module; CanP2 consists of three modules; and CanP2 comprises six modules. CanPF hypothetically serves as one end of the PKS gene cluster. The starter unit is PABA (p-aminobenzoic acid), and the extender units are four methylmalonyl-CoAs and 17 malonyl-CoAs. At the end of the process in PKS, the molecule is released by thioesterase (CanT). In the next step, the compound is cyclized to become candicidin aglycone, oxidized by P450 monooxygenase (CanC) with aid from ferredoxin (CanF). The last step is glycosylation by adding mycosamine to the structure [43,44].
2.4. Biosynthesis of Tautomycetin
Tautomycetin, firstly isolated from Streptomyces griseochromogenes and then from Streptomyces sp. CK4412, is an antifungal compound and an activated T cell-specific immunosuppressive compound. The inhibition of T-cells’ proliferation is by the apoptosis mechanism. Unlike the other type I polyketide-derived compounds, tautomycetin has a linear structure [32,51].
The tautomycetin (TMC) biosynthetic gene cluster (~70 kb) has two ORFs that encode type I PKSs (Tmc A and TmcB). TmcA has six modules including the loading module, and TmcB has four modules, the TE (thioesterase) domain of which is located in TmcB. TE domain is responsible for releasing the intermediate chain of the compound from the PKS. The biosynthesis of TMC requires malonyl-CoA as a starter unit and the extender units such as 4 malonyl-CoAs, 4 methylmalonyl-CoAs, and 1 ethylmalonyl-CoA. After being released from PKS, the intermediate compound is modified by post-PKS mechanisms such as hydroxylation, decarboxylation, dehydration, and esterification with the cyclic C8 dialkylmaleic anhydride moiety [32].
3. Polyketide Synthases Type II
The type II polyketide synthases (type II PKSs) are responsible for producing aromatic polyketide. Based on the polyphenolic ring system and their biosynthetic pathways, the aromatic polyketides produced by type II PKSs generally are classified into seven groups, i.e., anthracyclines, angucyclines, aureolic acids, tetracyclines, tetracenomycins, pradimicin-type polyphenols, and benzoisochromanequinones [52].
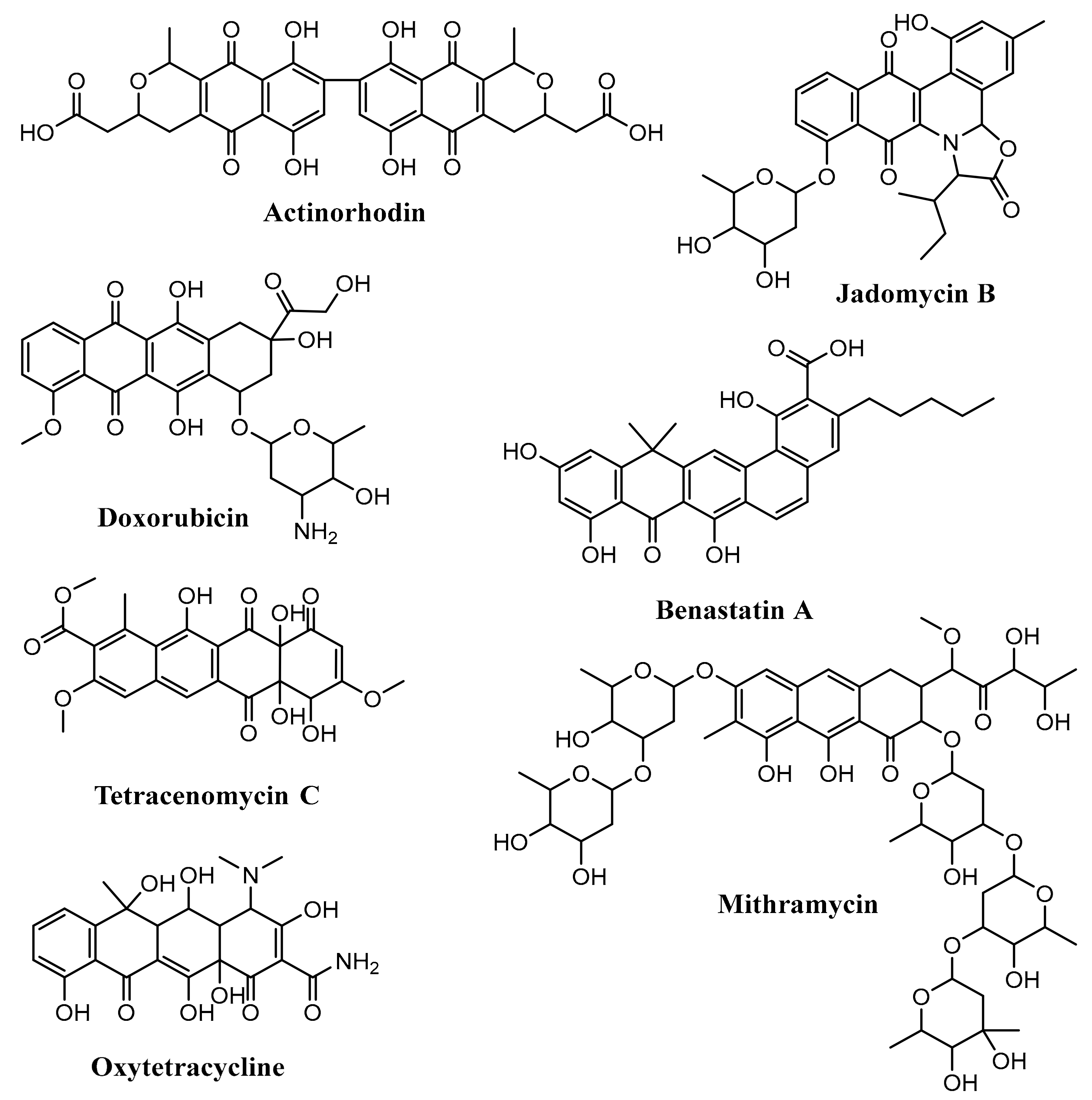
Anthracyclines consists of a linear tetracyclic ring system with quinone–hydroquinone groups in rings B and C. Angucyclines have an angular tetracyclic ring system. The aureolic acids have a tricyclic chromophore. Tetracyclines contain a linear tetracyclic ring system without quinone–hydroquinone groups in rings B and C. Tetracenomycins have a linear tetracyclic ring system with the quinone group in ring B. Pradimicin-type polyphenols are considered as extended angucyclines. Benzoisochromanequinones contain a quinone derivative from the isochroman structure [52]. Some examples of aromatic polyketide produced by Streptomyces are actinorhodin (benzoisochromanequinones), doxorubicin (anthracyclines), jadomycin B (angucyclines), oxytetracycline (tetracyclines), mithramycin (aureolic acids), tetracenomycin C (tetracenomycins), and benastatin A (pradimicin-type polyphenols) (Figure 5) [28,52,53,54,55,56,57].
Unlike type I PKSs that involve huge multifunctional proteins that have many modules containing domains and perform the enzymatic reaction in a non-iterative way, the type II PKSs have monofunctional polypeptides and work iteratively to produce aromatic polyketide. However, like the type I PKS, the type II PKSs also comprise the acyl carrier protein (ACP) that functions as an anchor for the nascent polyketide chain. In addition to possessing ACP, the type II PKSs also consists of two ketosynthases units (KSα and KSβ) that work cooperatively to produce the poly-β-keto chain. The KSα unit catalyzes the condensation of the precursors; on the other hand, the role of KSβ in the type II PKSs is as a chain length-determining factor. The three major systems (ACP, KSα, and KSβ) are called “minimal PKS” that work iteratively to produce aromatic polyketide. The other additional enzymes such as ketoreductases, cyclases, and aromatases cooperate to transform the poly-β-keto chain into the aromatic compound core. Furthermore, the post-tailoring process is conducted by oxygenases and glycosyl and methyl transferases [52,58,59,60]. The list of some aromatic polyketides produced by Streptomyces with their type II PKSs can be seen in Table 2.
3.1. Biosynthesis of Doxorubicin
Doxorubicin was isolated from Streptomyces peucetius in the early of 1960s. It belongs to anthracyclines that have a tetracyclic ring containing quinone and a hydroquinone group in their structure. Doxorubicin is one of the important drugs for the treatment of cancer such as breast cancer, childhood solid tumors, soft tissue sarcomas, and aggressive lymphomas. There are some proposed mechanisms for how doxorubicin kills the cancer cells: (i) intercalation of DNA and interference of topoisomerase-II-mediated DNA repair; and (ii) formation of free radicals and their deterioration of cell components such as cellular membranes, DNA, and proteins [61,62,63].
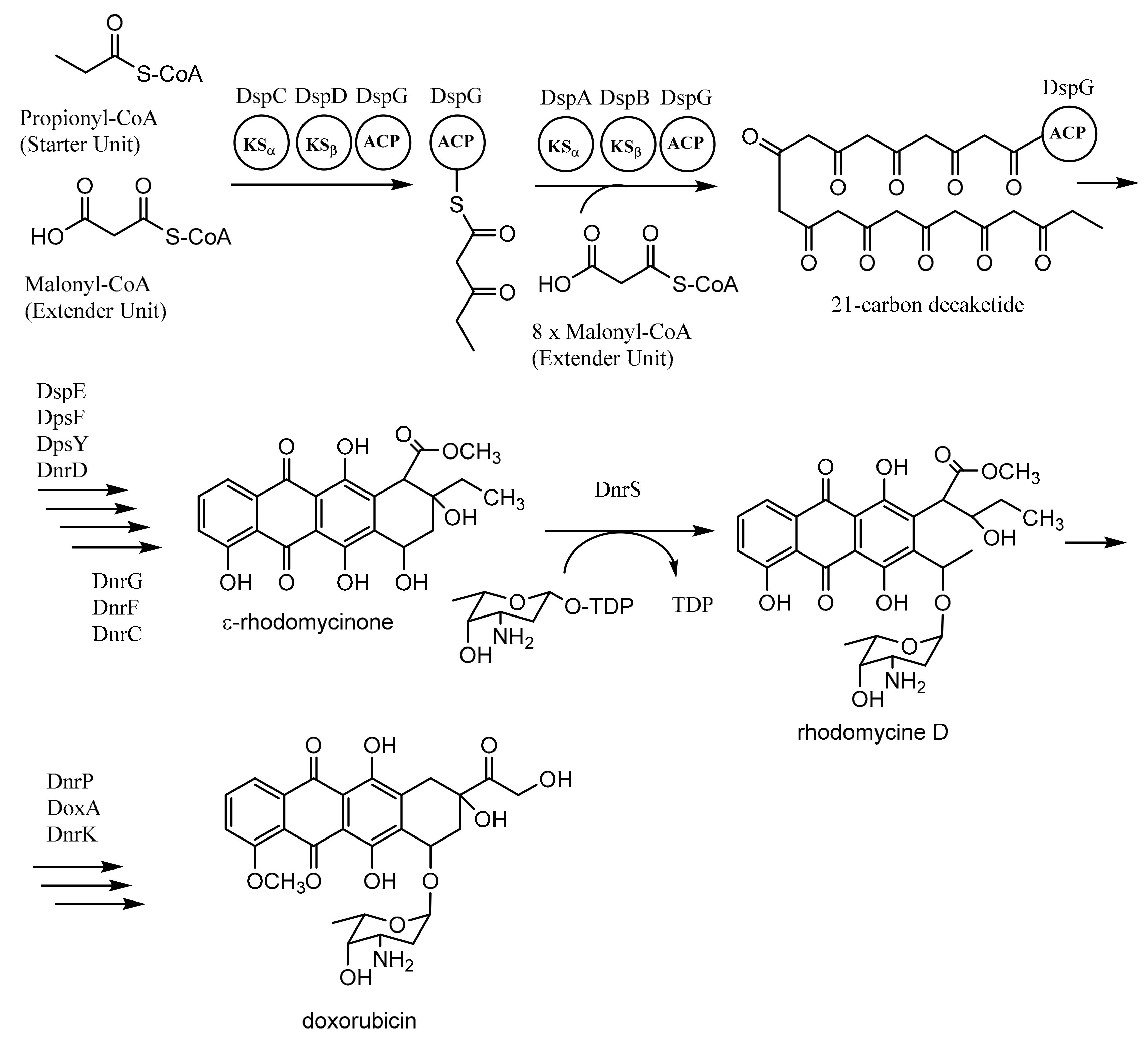
Daunorubicin (DNR)-doxorubicin (DXR) type II PKSs, encoded by dps genes in Streptomyces peucetius, are involved in the formation of doxorubicin. The biosynthesis of doxorubicin requires one propionyl-CoA as the starter unit and nine malonyl-CoAs as the extender units. The process involves two “minimal PKSs” (DpsC-DpsD-DpsG and DpsA-DpsB-DpsG) to produce a 21-carbon decaketide as an intermediate compound. The repetitive process is conducted by KSα (DpsA), KSβ (DpsB), and ACP (DpsG). The next process employs several enzymes such as ketoreductase (DpsE), cyclases (DpsF, DpsY, and DnrD), oxygenase (DnrG and DnrF), and methyl transferase (DnrC) to produce ε-rhodomycinone, an important intermediate of doxorubicin biosynthesis. The remaining steps to synthesize doxorubicin utilize glycosyltransferase (DnrS) with the thymidine-diphospho (TDP) derivative of l-daunosamine, methyl esterase (DnrP), oxygenase (DoxA), and methyl transferase (DnrK) (Figure 6) [60,64,65,66,67,68].
3.2. Biosynthesis of Medermycin
Medermycin is a benzoisochromanequinone (BIQ) antibiotic, isolated from Streptomyces sp. K73. It has high activity against some Gram-positive bacteria such as Staphylococcus aureus, Staphylococcus epidermidis, Sarcina lutea, Bacillus subtilis, and Bacillus cereus. Besides antibiotic activity, medermycin also has potent activity as a platelet aggregation inhibitor. Because of its unique ability to give different colors in acidic and alkaline aqueous solution, medermycin is considered as an indicator type antibiotic [69,70,84].
Biosynthesis of medermycin requires eight malonyl-CoAs and a sugar molecule, angolosamine, which is derived from the deoxyhexose (DOH) pathway. In the first step of biosynthesis, the minimal PKS that consists of ACP (encoded by the med-ORF23 gene), KSα (encoded by med-ORF1), and KSβ (encoded by med-ORF2) forms an octaketide moiety. The next process employs several enzymes such as keto reductase, aromatase, cyclase, enoyl reductase, and oxygenase/hydroxylase to produce the aglycone compound dihydrokalafungin. The aglycone structure then is combined by C-glycosyl transferase with an angolosamine structure to yield the final structure medermycin [69].
3.3. Biosynthesis of Hedamycin
Hedamycin is a pluramycin antitumor antibiotic, produced by Streptomyces griseoruber. This aromatic polyketide has a planar anthrapyrantrione chromophore, two amino sugars in its structure (α-l-N,N-dimethylvancosamine and β-d-angolosamine), and a bisepoxide-containing a side chain. The compound could inhibit 50% of human cancer cell growth at a subnanomolar concentration in three days. It is a monofunctional DNA alkylating agent, and because of its low therapeutic index, hedamycin is not clinically used [81,85].
Biosynthesis of hedamycin uses twelve malonyl-CoAs and two amino sugars, vancosamine and an angolosamine moiety. The minimal type II PKSs of hedamycin biosynthesis consist of HedC (KSα), HedD (CLF), and HedE (ACP). Uniquely, the initial process involves type I PKSs (HedT and HedU proteins) that produce the 2,4-hexadienyl primer unit from three malonyl-CoAs, and then, it is transferred to the minimal type II PKSs of hedamycin biosynthesis. After that, a dodecaketide structure is formed by processing nine malonyl-CoAs. The structure then is modified with keto reductase, aromatase/cyclase, and oxygenase into the aglycone compound. In the last step, two glycosyltransferases are used for incorporating two amino sugars to produce hedamycin [80,81].
3.4. Biosynthesis of Fredericamycin
Fredericamycin, isolated from Streptomyces griseus ATCC 49344, is an aromatic polyketide that contains a spirocyclic structure. It has moderate antitumor and cytotoxic activity in various cell lines. These bioactivities are suggested because of the blockage of topoisomerases I and II or the peptidyl-prolyl cis-trans isomerase Pin1 [83].
The biosynthesis of fredericamycin employs the minimal type II PKSs that contains KSα (FdmF and FdmS), KSβ (FdmG), and ACP (FdmH). There are two alternative mechanisms for chain initiation in the biosynthesis of fredericamycin. The first one requires acetyl-CoA and two malonyl-CoAs to produce the hexadienyl-priming unit. The second mechanism is by utilizing butyryl- or crotonyl-CoA and one malonyl-CoA to yield the hexadienyl-priming unit. The next step is carried out by processing twelve malonyl-CoAs as extender units to give the pentadecaketide intermediate, and then, the cyclases and oxygenases modify the intermediate compound into the final product [82,83,86].
4. Polyketide Synthases Type III
Unlike the type I and type II PKSs, the type III PKSs do not utilize ACP as an anchor for the production of polyketide metabolite. In this case, acyl-CoAs are used directly as substrates for generating polyketide compounds. In order to create polyketides, this system contains enzymes that construct homodimers and catalyzes many reactions such as priming, extension, and cyclization in an iterative way. With this fact, the type III PKSs are the simplest structures among the other types of PKSs. The type III PKSs found in bacteria were reported the first time in1999, and before that time, the type III PKSs were known only to be detected in plants [87,88,89].
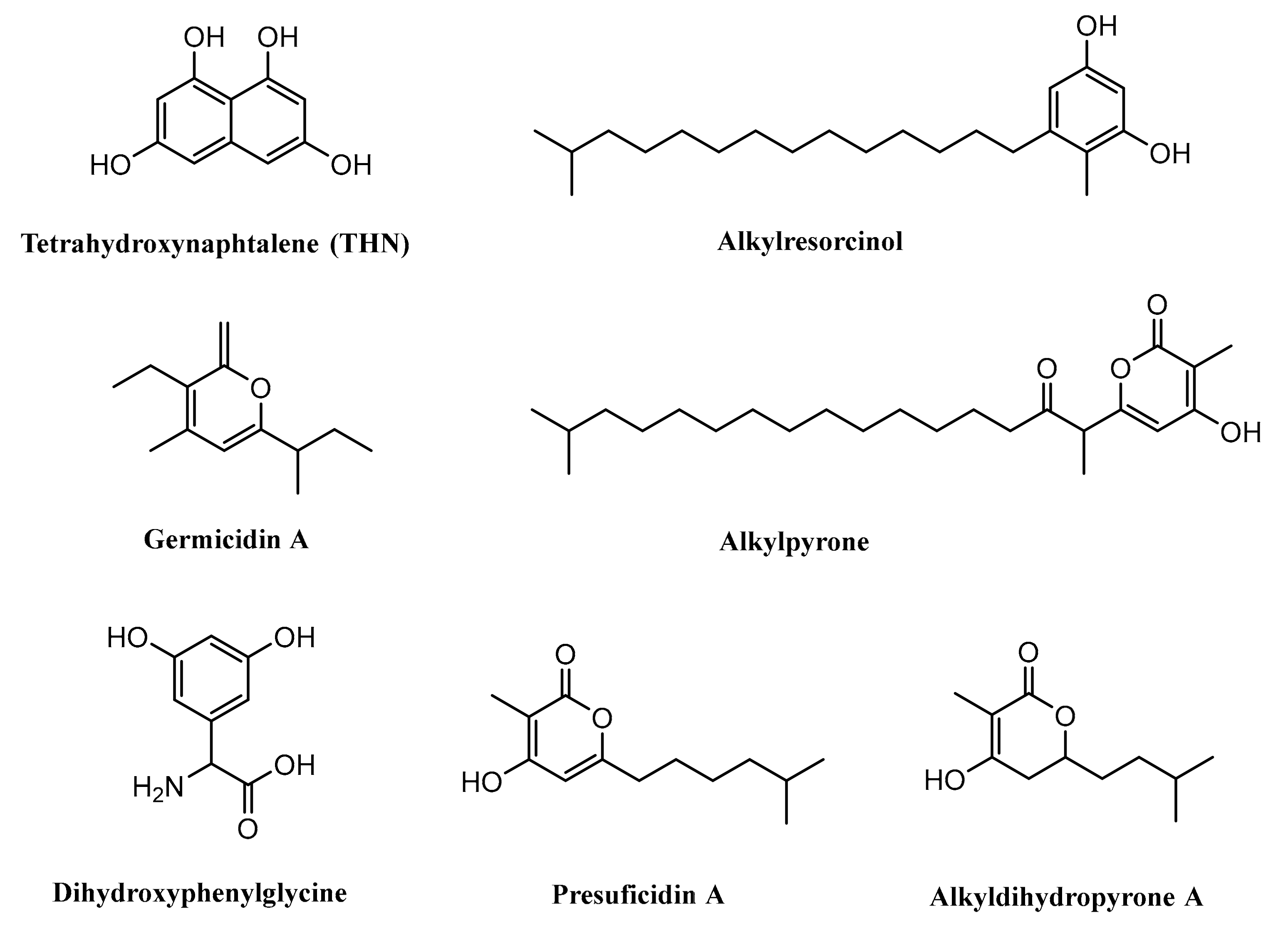
Some studies previously revealed that type III PKSs could also be identified in the Streptomyces such as RppA, found in Streptomyces griseus, which is responsible in the synthesis of 1,3,6,8-tetrahydroxynaphthalene (THN) [90]. Gcs, identified in Streptomyces coelicolor A3(2), is reported to have an important role in the biosynthesis of germicidin [91]. SrsA, encoded by the srsA gene and isolated from Streptomyces griseus, is known to have an important role in the biosynthesis of phenolic lipids, i.e., alkylresorcinols and alkylpyrones [29].
The type III PKS Ken2, isolated from Streptomyces violaceoruber, was suggested to be involved in the production of 3,5-dihydroxyphenylglycine (3,5-DHPG). This compound is a nonproteinogenic amino acid needed for the formation of kendomycin and several other glycopeptide antibiotics such as balhimycin, chloroeremomycin, and also vancomycin [92]. Cpz6, encoded by the cpz6 gene and isolated from Streptomyces sp. MK730–62F2, was reported to be engaged in the biosynthesis of caprazamycins by producing a group of new triketidepyrenes (presulficidins) [93]. Moreover, another finding also suggested that DpyA catalyzes the formation of alkyldihydropyrones in Streptomyces reveromyceticus (Figure 7) [94].
4.1. Biosynthesis of Germicidin
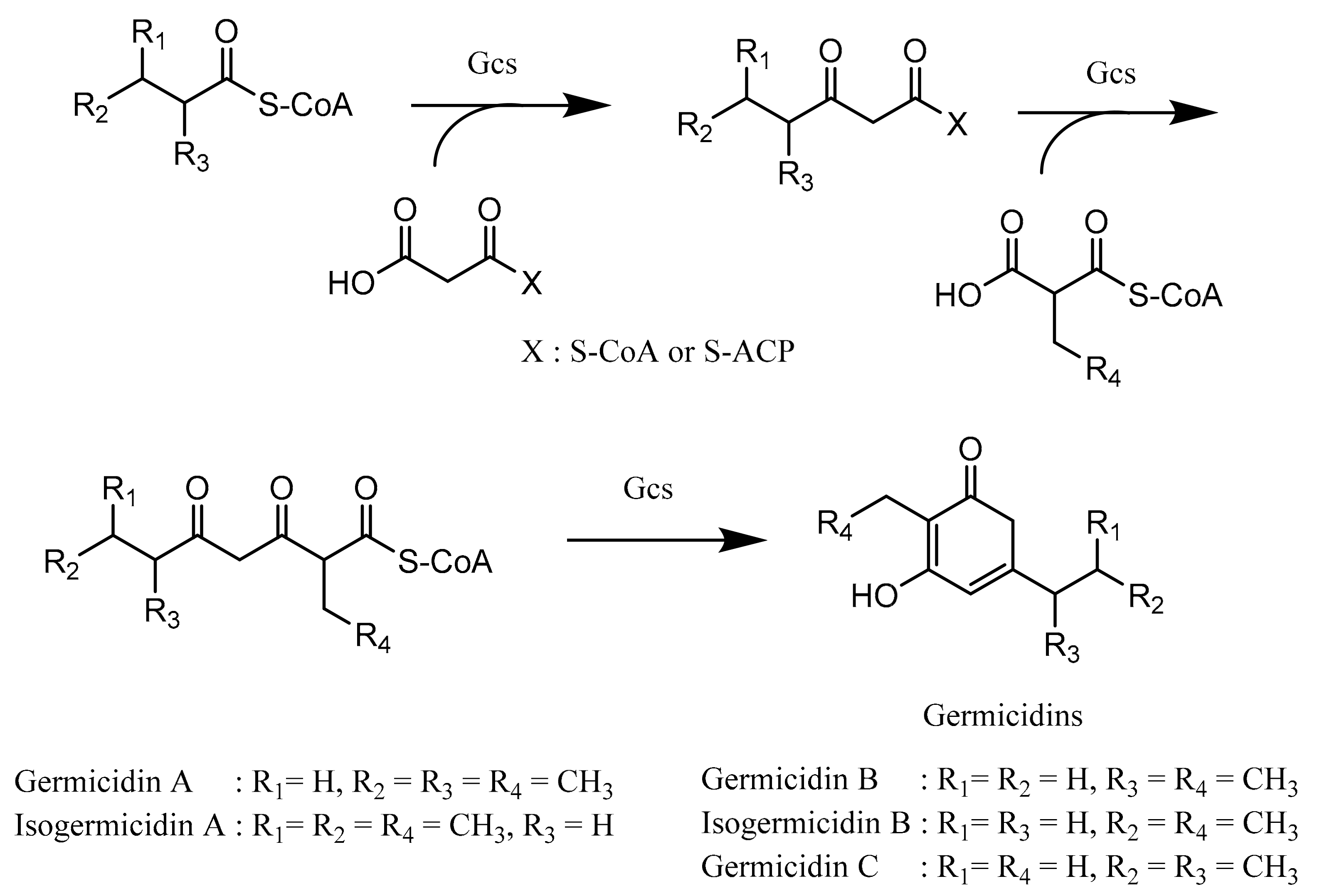
Germicidin, a pyrone-derived polyketide, is produced by a type III PKS germicidin synthase (Gcs) and is known to inhibit spore germination. Germicidin A, produced by Streptomyces viridochromogenes and Streptomyces coelicolor, prevents the spore germination reversibly at a very low concentration (40 pg/mL). The mechanism of inhibition is suggested by affecting the sporal respiratory chain and blocking Ca2+-activated ATPase, thus resulting in inadequate energy for spore germination. Furthermore, germicidin A also has antibacterial properties against various Gram-positive bacteria [95,96].
Although many bacterial type III PKSs use only malonyl-CoA as both starter and extender units, the type III PKS Gcs, which is responsible for germicidin biosynthesis, is suggested to have the ability to utilize either acyl-ACP or acyl-CoA such as medium-chain acyl-CoAs (C4–C8) as starter units and malonyl-CoA, methylmalonyl-CoA, and ethylmalonyl-CoA as extender units [97,98]. In the first step, the starter unit is transacylated onto the cysteine residue of Gcs, and then, Gcs catalyzes the condensation reaction between the starter unit and extender unit concomitantly with the decarboxylation process, resulting in β-ketoacyl-CoA. The process continues with β-ketoacyl-CoA, which transacylates back onto the cysteine residue of Gcs (repetitive process) and subsequently undergoes a condensation reaction with either methylmalonyl-CoA or ethylmalonyl-CoA simultaneously with decarboxylation to formulate β,δ-diketothioester of CoA or a triketide intermediate. In the end of the reaction, cyclization of the β,δ-diketothioester of CoA is carried out to produce various types of germicidins (Figure 8) [91].
4.2. Biosynthesis of Tetrahydroxynaphthalene
Tetrahydroxynaphthalene or THN is a small aromatic compound that is produced by utilizing type III PKSs (RppA). The biosynthesis process of THN requires five molecules of malonyl-CoA to form a pentaketide intermediate structure, and then, it is cyclized and aromatized to yield THN product. Spontaneous oxidation of THN may result flaviolin (red pigment) [90,99,100].
4.3. Biosynthesis of Dihydroxyphenylglycine
In order to synthesize 3,5-dihydroxyphenylglycine (3,5-DHPG), four malonyl-CoAs are needed, and the process is catalyzed by type III PKS (Ken2 or DpgA), which leads to the formation of the intermediate tetraketide compound. The tetraketide further is modified by hydratase/dehydratase, and oxidase/thioesterase to form 3,5-dihydroxyphenylacetic acid. The final step involves transaminase and tyrosine, as the amino group donor, to yield 3,5-DHPG, which is known as a nonproteinogenic amino acid [92,100,101].
4.4. Biosynthesis of Alkylresorcinol
The alkylresorcinol biosynthesis in Streptomyces griseus is catalyzed by SrsA. The reaction needs fatty acid (starter unit), one methylmalonyl-CoA, and two malonyl-CoAs (extender unit), and the intermediate structure is tetraketide. The tetraketide structure then transforms into the aromatic compound nonenzymatically (alkylresorcinol). This reaction may occur because of the nucleophilic attack on the thioester group by the methine carbon of the intermediate tetraketide compound [29].
5. Conclusions
Streptomyces has various systems in order to produce polyketides with different structures and functions. Knowing the polyketide structures, activities, producing enzymes, starter units, extender units, and the structural genes are very important in the development of new drugs. Some mechanisms of polyketide biosynthesis in Streptomyces that have been reported previously could provide strong basic knowledge not only for the biosynthesis investigation of the new polyketides, but also engineering the producing system in the future.
Acknowledgments
The authors gratefully acknowledge support from German Federal Ministry of Education and Research (BMBF) under the German-Indonesian anti-infective cooperation (GINAICO) project, a fellowship awarded by the German Academic Exchange Service (German: Deutscher Akademischer Austauschdienst or DAAD), and The President’s Initiative and Networking Funds of the Helmholtz Association of German Research Centres (German: Helmholtz Gemeinschaft Deutscher Forschungszentren or HGF) under Contract Number VH-GS-202.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Moore, B.S.; Hopke, J.N. Discovery of a new bacterial polyketide biosynthetic pathway. ChemBioChem 2001, 2, 35–38. [Google Scholar] [CrossRef]
- Rokem, J.S.; Lantz, A.E.; Nielsen, J. Systems biology of antibiotic production by microorganisms. Nat. Prod. Rep. 2007, 24, 1262. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Funa, N.; Miyahisa, I.; Horinouchi, S. Synthesis of unnatural flavonoids and stilbenes by exploiting the plant biosynthetic pathway in Escherichia coli. Chem. Biol. 2007, 14, 613–621. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A.; Rice, L.B. Antifungal agents: Mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [CrossRef]
- Tacar, O.; Sriamornsak, P.; Dass, C.R. Doxorubicin: An update on anticancer molecular action, toxicity and novel drug delivery systems. J. Pharm. Pharmacol. 2013, 65, 157–170. [Google Scholar] [CrossRef]
- Shushni, M.A.M.; Singh, R.; Mentel, R.; Lindequist, U. Balticolid: A new 12-membered macrolide with antiviral activity from an Ascomycetous fungus of marine origin. Mar. Drugs 2011, 9, 844–851. [Google Scholar] [CrossRef]
- Li, J.; Kim, S.G.; Blenis, J. Rapamycin: One drug, many effects. Cell Metab. 2014, 19, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Van de Donk, N.W.C.J.; Kamphuis, M.M.J.; Lokhorst, H.M.; Bloem, A.C. The cholesterol lowering drug lovastatin induces cell death in myeloma plasma cells. Leukemia 2002, 16, 1362–1371. [Google Scholar] [CrossRef]
- Hleba, L.; Charousova, I.; Cisarova, M.; Kovacik, A.; Kormanec, J.; Medo, J.; Bozik, M.; Javorekova, S. Rapid identification of Streptomyces tetracycline producers by MALDI-TOF mass spectrometry. J. Environ. Sci. Heal. Part A 2018. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, R.; Sathiyabama, M. Lovastatin-producing endophytic fungus isolated from a medicinal plant Solanum xanthocarpum. Nat. Prod. Res. 2015, 29, 2282–2286. [Google Scholar] [CrossRef]
- Huang, Q.; Lu, G.; Shen, H.-M.; Chung, M.C.M.; Ong, C.N. Anti-cancer properties of anthraquinones from rhubarb. Med. Res. Rev. 2006, 27, 609–630. [Google Scholar] [CrossRef]
- Kohli, G.S.; John, U.; Figueroa, R.I.; Rhodes, L.L.; Harwood, D.T.; Groth, M.; Bolch, C.J.S.; Murray, S.A. Polyketide synthesis genes associated with toxin production in two species of Gambierdiscus (Dinophyceae). BMC Genomics 2015, 16, 410. [Google Scholar] [CrossRef]
- Florian, P.; Monika, H. Polyketides in insects: Ecological role of these widespread chemicals and evolutionary aspects of their biogenesis. Biol. Rev. 2008, 83, 209–226. [Google Scholar]
- Adele, C.; Guido, C.; Guido, V.; Angelo, F. Shaping the polypropionate biosynthesis in the solar-powered mollusc Elysia viridis. ChemBioChem 2008, 10, 315–322. [Google Scholar]
- Müller, R.; Wink, J. Future potential for anti-infectives from bacteria—How to exploit biodiversity and genomic potential. Int. J. Med. Microbiol. 2014, 304, 3–13. [Google Scholar] [CrossRef]
- Widyastuti, Y.; Lisdiyanti, P.; Ratnakomala, S.; Kartina, G.; Ridwan, R.; Rohmatussolihat, R.; Rosalinda Prayitno, N.; Triana, E.; Widhyastuti, N.; et al. Genus diversity of Actinomycetes in Cibinong Science Center, West Java, Indonesia. Microbiol. Indones. 2012, 6, 165–172. [Google Scholar] [CrossRef]
- Lucas, X.; Senger, C.; Erxleben, A.; Grüning, B.A.; Döring, K.; Mosch, J.; Flemming, S.; Günther, S. StreptomeDB: A resource for natural compounds isolated from Streptomyces species. Nucleic Acids Res. 2013, 41, D1130–D1136. [Google Scholar] [CrossRef]
- Igarashi, M.; Takahashi, Y.; Shitara, T.; Nakamura, H.; Naganawa, H.; Miyake, T.; Akamatsu, Y. Caprazamycins, novel lipo-nucleoside antibiotics, from Streptomyces sp. J. Antibiot. 2005, 58, 327–337. [Google Scholar] [CrossRef]
- Dutta, S.; Basak, B.; Bhunia, B.; Chakraborty, S.; Dey, A. Kinetics of rapamycin production by Streptomyces hygroscopicus MTCC 4003. 3 Biotech. 2014, 4, 523–531. [Google Scholar] [CrossRef]
- Rodríguez, L.; Rodríguez, D.; Olano, C.; Braña, A.F.; Méndez, C.; Salas, J.A. Functional analysis of OleY L-oleandrosyl 3-O-methyltransferase of the oleandomycin biosynthetic pathway in Streptomyces antibioticus. J. Bacteriol. 2001, 183, 5358–5363. [Google Scholar] [CrossRef] [PubMed]
- Elibol, M. Optimization of medium composition for actinorhodin production by Streptomyces coelicolor A3(2) with response surface methodology. Process Biochem. 2004, 39, 1057–1062. [Google Scholar] [CrossRef]
- Pokhrel, A.R.; Chaudhary, A.K.; Nguyen, H.T.; Dhakal, D.; Le, T.T.; Shrestha, A.; Liou, K.; Sohng, J.K. Overexpression of a pathway specific negative regulator enhances production of daunorubicin in bldA deficient Streptomyces peucetius ATCC 27952. Microbiol. Res. 2016, 192, 96–102. [Google Scholar] [CrossRef]
- Shelest, E.; Heimerl, N.; Fichtner, M.; Sasso, S. Multimodular type I polyketide synthases in algae evolve by module duplications and displacement of AT domains in trans. BMC Genomics 2015, 16, 1–15. [Google Scholar] [CrossRef]
- Hopwood, D.A. Cracking the polyketide code. PLoS Biol. 2004, 2, 166–169. [Google Scholar] [CrossRef]
- Staunton, J.; Weissman, K.J. Polyketide biosynthesis: A millennium review. Nat. Prod. Rep. 2001, 18, 380–416. [Google Scholar] [CrossRef]
- Lal, R.; Kumari, R.; Kaur, H.; Khanna, R.; Dhingra, N.; Tuteja, D. Regulation and manipulation of the gene clusters encoding type I PKSs. Trends Biotechnol. 2000, 18, 264–274. [Google Scholar] [CrossRef]
- Okamoto, S.; Taguchi, T.; Ochi, K.; Ichinose, K. Biosynthesis of actinorhodin and related antibiotics: Discovery of alternative routes for quinone formation encoded in the act gene cluster. Chem. Biol. 2009, 16, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Funabashi, M.; Funa, N.; Horinouchi, S. Phenolic lipids synthesized by type III polyketide synthase confer penicillin resistance on Streptomyces griseus. J. Biol. Chem. 2008, 283, 13983–13991. [Google Scholar] [CrossRef]
- Vance, S.; Tkachenko, O.; Thomas, B.; Bassuni, M.; Hong, H.; Nietlispach, D.; Broadhurst, W. Sticky swinging arm dynamics: Studies of an acyl carrier protein domain from the mycolactone polyketide synthase. Biochem. J. 2016, 473, 1097–1110. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Stassi, D.; Lax, S.A.; Katz, L. A second type I PKS gene cluster isolated from Streptomyces hygroscopicus ATCC 29253, a rapamycin-producing strain. Gene 1997, 203, 1–9. [Google Scholar] [CrossRef]
- Choi, S.S.; Hur, Y.A.; Sherman, D.H.; Kim, E.S. Isolation of the biosynthetic gene cluster for tautomycetin, a linear polyketide T cell-specific immunomodulator from Streptomyces sp. CK4412. Microbiology 2007, 153, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Mazzei, T.; Mini, E.; Noveffi, A.; Periti, P. Chemistry and mode of action of macrolides. J. Antimicrob. Chemother. 1993, 31, 1–9. [Google Scholar] [CrossRef]
- Schwecke, T.; Aparicio, J.F.; Molnár, I.; König, A.; Khaw, L.E.; Haydock, S.F.; Oliynyk, M.; Caffrey, P.; Cortés, J.; Lester, J.B. The biosynthetic gene cluster for the polyketide immunosuppressant rapamycin. Proc. Natl. Acad. Sci. USA 1995, 92, 7839–7843. [Google Scholar] [CrossRef] [PubMed]
- Motamedi, H.; Shafiee, A. The biosynthetic gene cluster for the macrolactone ring of the immunosuppressant FK506. Eur. J. Biochem. 1998, 256, 528–534. [Google Scholar] [CrossRef]
- Karray, F.; Darbon, E.; Oestreicher, N.; Dominguez, H.; Tuphile, K.; Gagnat, J.; Blondelet-Rouault, M.H.; Gerbaud, C.; Pernodet, J.L. Organization of the biosynthetic gene cluster for the macrolide antibiotic spiramycin in Streptomyces ambofaciens. Microbiology 2007, 153, 4111–4122. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zhao, L.; Liu, H.W.; Sherman, D.H. A gene cluster for macrolide antibiotic biosynthesis in Streptomyces venezuelae: Architecture of metabolic diversity. Proc. Natl. Acad. Sci. USA 1998, 95, 12111–12116. [Google Scholar] [CrossRef]
- Cruz, M.C.; Cavallo, L.M.; Görlach, J.M.; Cox, G.; Perfect, J.R.; Cardenas, M.E.; Heitman, J. Rapamycin antifungal action is mediated via conserved complexes with FKBP12 and TOR kinase homologs in Cryptococcus neoformans. Mol. Cell Biol. 1999, 19, 4101–4112. [Google Scholar] [CrossRef]
- Bastidas, R.J.; Shertz, C.A.; Lee, S.C.; Heitman, J.; Cardenas, M.E. Rapamycin exerts antifungal activity in vitro and in vivo against Mucor circinelloides via FKBP12-dependent inhibition of tor. Eukaryot. Cell 2012, 11, 270–281. [Google Scholar] [CrossRef]
- Kwan, D.H.; Schulz, F. The stereochemistry of complex polyketide biosynthesis by modular polyketide synthases. Molecules 2011, 16, 6092–6115. [Google Scholar] [CrossRef]
- Ikeda, H.; Nonomiya, T.; Usami, M.; Ohta, T.; Omura, S. Organization of the biosynthetic gene cluster for the polyketide anthelmintic macrolide avermectin in Streptomyces avermitilis. Proc. Natl. Acad. Sci. USA 1999, 96, 9509–9514. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.L.; Hu, Z.; Schirmer, A.; Reid, R.; Revill, W.P.; Reeves, C.D.; Petrakovsky, O.V.; Dong, S.D.; Katz, L. Chalcomycin biosynthesis gene cluster from Streptomyces bikiniensis: Novel features of an unusual ketolide produced through expression of the chm polyketide synthase in Streptomyces fradiae. Antimicrob. Agents Chemother. 2004, 48, 4703–4712. [Google Scholar] [CrossRef]
- Gil, J.A.; Campelo-Diez, A.B. Candicidin biosynthesis in Streptomyces griseus. Appl. Microbiol. Biotechnol. 2003, 60, 633–642. [Google Scholar] [CrossRef]
- Campelo, A.B.; Gil, J.A. The candicidin gene cluster from Streptomyces griseus IMRU 3570. Microbiology 2002, 148, 51–59. [Google Scholar] [CrossRef]
- Kino, T.; Hatanaka, H.; Hashimoto, M.; Nishiyama, M.; Goto, T.; Okuhara, M.; Kohsaka, M.; Aoki, H.; Imanaka, H. FK-506, a novel immunosuppressant isolated from a Streptomyces. I. Fermentation, isolation, and physico-chemical and biological characteristics. J. Antibiot. (Tokyo) 1987, 40, 1249–1255. [Google Scholar] [CrossRef]
- Wu, K.; Chung, L.; Revill, W.P.; Katz, L.; Reeves, C.D. The FK520 gene cluster of Streptomyces hygroscopicus var. ascomyceticus (ATCC 14891) contains genes for biosynthesis of unusual polyketide extender units. Gene 2000, 251, 81–90. [Google Scholar] [CrossRef]
- Aparicio, J.; Colina, A.; Ceballos, E.; Martin, J. The biosynthetic gene cluster for the 26-membered ring polyene macrolide pimaricin. A new polyketide synthase organization encoded by two subclusters separated by functionalization genes. J. Biol. Chem. 1999, 274, 10133–10139. [Google Scholar] [CrossRef]
- Fiers, W.D.; Dodge, G.J.; Li, Y.; Smith, J.L.; Fecik, R.A.; Aldrich, C.C. Tylosin polyketide synthase module 3: Stereospecificity, stereoselectivity and steady-state kinetic analysis of β-processing domains via diffusible, synthetic substrates. Chem. Sci. 2015, 6, 5027–5033. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Z.; Li, M.; Wen, Y.; Song, Y.; Li, J. Construction of ivermectin producer by domain swaps of avermectin polyketide synthase in Streptomyces avermitilis. Appl. Microbiol. Biotechnol. 2006, 72, 986–994. [Google Scholar] [CrossRef]
- Hammond, S.M.; Kliger, B.N. Mode of action of the polyene antibiotic candicidin: Binding factors in the wall of Candida albicans. Antimicrob. Agents Chemother. 1976, 9, 561–568. [Google Scholar] [CrossRef]
- Cheng, X.-C.; Kihara, T.; Ying, X.; Uramoto, M.; Osada, H.; Kusakabe, H.; Wang, B.-N.; Kobayashi, Y.; Ko, K.; Yamaguchi, I.; Shen, Y.-C.; Isono, K. A new antibiotic, tautomycetin. J. Antibiot. 1989, 42, 141–144. [Google Scholar]
- Hertweck, C.; Luzhetskyy, A.; Rebets, Y.; Bechthold, A. Type II polyketide synthases: Gaining a deeper insight into enzymatic teamwork. Nat. Prod. Rep. 2007, 24, 162–190. [Google Scholar] [CrossRef]
- Malla, S.; Prasad Niraula, N.; Singh, B.; Liou, K.; Kyung Sohng, J. Limitations in doxorubicin production from Streptomyces peucetius. Microbiol. Res. 2010, 165, 427–435. [Google Scholar] [CrossRef]
- Jakeman, D.L.; Bandi, S.; Graham, C.L.; Reid, T.R.; Wentzell, J.R.; Douglas, S.E. Antimicrobial activities of jadomycin B and structurally related analogues. Antimicrob. Agents Chemother. 2009, 53, 1245–1247. [Google Scholar] [CrossRef]
- Lombó, F.; Menéndez, N.; Salas, J.A.; Méndez, C. The aureolic acid family of antitumor compounds: Structure, mode of action, biosynthesis, and novel derivatives. Appl. Microbiol. Biotechnol. 2006, 73, 1–14. [Google Scholar] [CrossRef]
- Petkovic, H.; Cullum, J.; Hranueli, D.; Hunter, I.S.; Peric-Concha, N.; Pigac, J.; Thamchaipenet, A.; Vujaklija, D.; Long, P.F. Genetics of Streptomyces rimosus, the oxytetracycline producer. Microbiol. Mol. Biol. Rev. 2006, 70, 704–728. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Hutchinson, C.R. Triple hydroxylation of tetracenomycin A2 to tetracenomycin C in Streptomyces glaucescens. Overexpression of the tcmG gene in Streptomyces lividans and characterization of the tetracenomycin A2 oxygenase. J. Biol. Chem. 1994, 1326, 30726–30733. [Google Scholar]
- Zhang, Z.; Pan, H.-X.; Tang, G.-L. New insights into bacterial type II polyketide biosynthesis. F1000Research 2017, 6, 172. [Google Scholar] [CrossRef] [PubMed]
- Komaki, H.; Harayama, S. Sequence diversity of type II polyketide synthase genes in Streptomyces. Actinomycetologica 2006, 20, 42–48. [Google Scholar] [CrossRef]
- Chan, Y.A.; Podevels, A.M.; Kevany, B.M.; Thomas, M.G. Biosynthesis of polyketide synthase extender units. Nat. Prod. Rep. 2009, 26, 90–114. [Google Scholar] [CrossRef] [PubMed]
- Minotti, G. Anthracyclines: Molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol. Rev. 2004, 56, 185–229. [Google Scholar] [CrossRef]
- Yang, F.; Teves, S.S.; Kemp, C.J.; Henikoff, S. Doxorubicin, DNA torsion, and chromatin dynamics. Biochim. Biophys. Acta 2014, 1845, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin pathways: Pharmacodynamics and adverse effects. Pharmacogenet. Genomics 2011, 21, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Sheldon, P.J.; Wendt-Pienkowski, E.; Richard Hutchinson, C. The Streptomyces peucetius dpsC gene determines the choice of starter unit in biosynthesis of the daunorubicin polyketide. J. Bacteriol. 1999, 181, 4690–4695. [Google Scholar]
- Grimm, A.; Madduri, K.; Ali, A.; Hutchinson, C.R. Characterization of the Streptomyces peucetius ATCC 29050 genes encoding doxorubicin polyketide synthase. Gene 1994, 151, 1–10. [Google Scholar] [CrossRef]
- Rajgarhia, V.B.; Strohl, W.R. Minimal Streptomyces sp. strain C5 daunorubicin polyketide biosynthesis genes required for aklanonic acid biosynthesis. J. Bacteriol. 1997, 179, 2690–2696. [Google Scholar] [CrossRef]
- Hutchinson, C.R. Biosynthetic studies of daunorubicin and tetracenomycin C. Chem. Rev. 1997, 97, 2525–2536. [Google Scholar] [CrossRef]
- Lomovskaya, N.; Doi-Katayama, Y.; Filippini, S.; Nastro, C.; Fonstein, L.; Gallo, M.; Colombo, A.L.; Hutchinson, C.R. The Streptomyces peucetius dpsY and dnrX genes govern early and late steps of daunorubicin and doxorubicin biosynthesis. J. Bacteriol. 1998, 180, 2379–2386. [Google Scholar]
- Ichinose, K.; Ozawa, M.; Itou, K.; Kunieda, K.; Ebizuka, Y. Cloning, sequencing and heterologous expression of the medermycin biosynthetic gene cluster of Streptomyces sp. AM-7161: Towards comparative analysis of the benzoisochromanequinone gene clusters. Microbiology 2003, 149, 1633–1645. [Google Scholar] [CrossRef] [PubMed]
- Takano, S.; Hasuda, K.; Ito, A.; Koide, Y.; Ishii, F. A new antibiotic, medermycin. J. Antibiot. (Tokyo) 1976, 29, 765–768. [Google Scholar] [CrossRef]
- Pickens, L.B.; Tang, Y. Oxytetracycline biosynthesis. J. Biol. Chem. 2010, 285, 27509–27515. [Google Scholar] [CrossRef]
- Fischer, C.; Lipata, F.; Rohr, J. The complete gene cluster of the antitumor agent gilvocarcin V and its implication for the biosynthesis of the gilvocarcins. J. Am. Chem. Soc. 2003, 125, 7818–7819. [Google Scholar] [CrossRef] [PubMed]
- Lombó, F.; Abdelfattah, M.S.; Braça, A.F.; Salas, J.A. Elucidation of oxygenation steps during oviedomycin biosynthesis and generation of derivatives with increased antitumor activity. ChemBioChem 2009, 10, 296–303. [Google Scholar] [CrossRef]
- Xu, Z.; Jakobi, K.; Welzel, K.; Hertweck, C. Biosynthesis of the antitumor agent chartreusin involves the oxidative rearrangement of an anthracyclic polyketide. Chem. Biol. 2005, 12, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Bretschneider, T.; Zocher, G.; Unger, M.; Scherlach, K.; Stehle, T.; Hertweck, C. A ketosynthase homolog uses malonyl units to form esters in cervimycin biosynthesis. Nat. Chem. Biol. 2011, 8, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Herold, K.; Xu, Z.; Gollmick, F.A.; Gräfe, U.; Hertweck, C. Biosynthesis of cervimycin C, an aromatic polyketide antibiotic bearing an unusual dimethylmalonyl moiety. Organ. Biomol. Chem. 2004, 2, 2411–2414. [Google Scholar] [CrossRef]
- Jakobi, K.; Hertweck, C. A gene cluster encoding resistomycin biosynthesis in Streptomyces resistomycificus; exploring polyketide cyclization beyond linear and angucyclic patterns. J. Am. Chem. Soc. 2004, 126, 2298–2299. [Google Scholar] [CrossRef] [PubMed]
- Menendez, N.; Nur-e-Alam, M.; Bran, A.F.; Rohr, J.; Salas, J.A.; Mendez, C. Biosynthesis of the antitumor chromomycin A3 in Streptomyces griseus: Analysis of the gene cluster and rational design of novel chromomycin analogs. Chem. Biol. 2004, 11, 21–32. [Google Scholar]
- Sun, L.; Zeng, J.; Cui, P.; Wang, W.; Yu, D.; Zhan, J. Manipulation of two regulatory genes for efficient production of chromomycins in Streptomyces reseiscleroticus. J. Biol. Eng. 2018, 12, 1–11. [Google Scholar] [CrossRef]
- Das, A.; Khosla, C. In vivo and in vitro analysis of the hedamycin polyketide synthase. Chem. Biol. 2009, 16, 1197–1207. [Google Scholar] [CrossRef]
- Bililign, T.; Hyun, C.-G.; Williams, J.S.; Czisny, A.M.; Thorson, J.S. The hedamycin locus implicates a novel aromatic PKS priming mechanism. Chem. Biol. 2004, 11, 959–969. [Google Scholar] [CrossRef]
- Das, A.; Szu, P.; Fitzgerald, J.T.; Khosla, C. Mechanism and engineering of polyketide chain initiation in fredericamycin biosynthesis. J. Am. Chem. Soc. 2010, 132, 8831–8833. [Google Scholar] [CrossRef]
- Chen, Y.; Wendt-Pienkowski, E.; Shen, B. Identification and utility of FdmR1 as a Streptomyces antibiotic regulatory protein activator for fredericamycin production in Streptomyces griseus ATCC 49344 and heterologous hosts. J. Bacteriol. 2008, 190, 5587–5596. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, A.; Fukamachi, N.; Yamaki, K.; Hayashi, M.; Oh-ishi, S.; Kobayashi, B.; Omura, S. Inhibition of platelet aggregation by medermycin and it’s related isochromanequinone antibiotics. J. Antibiot. 1987, 40, 1075–1076. [Google Scholar] [CrossRef]
- Tu, L.C.; Melendy, T.; Beerman, T.A. DNA damage responses triggered by a highly cytotoxic monofunctional DNA alkylator, hedamycin, a pluramycin antitumor antibiotic. Mol. Cancer Ther. 2004, 3, 577–585. [Google Scholar] [PubMed]
- Szu, P.-H.; Govindarajan, S.; Meehan, M.J.; Das, A.; Nguyen, D.D.; Dorrestein, P.C.; Minshull, J.; Khosla, C. Analysis of the ketosynthase-chain length factor heterodimer from the fredericamycin polyketide synthase. Acta Neurobiol. Exp. (Wars) 2011, 18, 1021–1031. [Google Scholar] [CrossRef]
- Shen, B. Polyketide biosynthesis beyond the type I, II and III polyketide synthase paradigms. Curr. Opin. Chem. Biol. 2003, 7, 285–295. [Google Scholar] [CrossRef]
- Yu, D.; Xu, F.; Zeng, J.; Zhan, J. Type III polyketide synthases in natural product biosynthesis. IUBMB Life 2012, 64, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Nakano, C.; Ozawa, H.; Akanuma, G.; Funa, N.; Horinouchi, S. Biosynthesis of aliphatic polyketides by type III polyketide synthase and methyltransferase in Bacillus subtilis. J. Bacteriol. 2009, 191, 4916–4923. [Google Scholar] [CrossRef] [PubMed]
- Funa, N.; Funabashi, M.; Ohnishi, Y.; Horinouchi, S. Biosynthesis of hexahydroxyperylenequinone melanin via oxidative aryl coupling by cytochrome P-450 in Streptomyces griseus. J. Bacteriol. 2005, 187, 8149–8155. [Google Scholar] [CrossRef]
- Song, L.; Barona-Gomez, F.; Corre, C.; Xiang, L.; Udwary, D.W.; Austin, M.B.; Noel, J.P.; Moore, B.S.; Challis, G.L. Type III polyketide synthase β-ketoacyl-ACP starter unit and ethylmalonyl-coA extender unit selectivity discovered by Streptomyces coelicolor genome mining. J. Am. Chem. Soc. 2006, 128, 14754–14755. [Google Scholar] [CrossRef]
- Wenzel, S.C.; Bode, H.B.; Kochems, I.; Müller, R. A type I/type III polyketide synthase hybrid biosynthetic pathway for the structurally unique ansa compound kendomycin. ChemBioChem 2008, 9, 2711–2721. [Google Scholar] [CrossRef]
- Tang, X.; Eitel, K.; Kaysser, L.; Kulik, A.; Grond, S.; Gust, B. A two-step sulfation in antibiotic biosynthesis requires a type III polyketide synthase. Nat. Chem. Biol. 2013, 9, 610–615. [Google Scholar] [CrossRef]
- Aizawa, T.; Kim, S.Y.; Takahashi, S.; Koshita, M.; Tani, M.; Futamura, Y.; Osada, H.; Funa, N. Alkyldihydropyrones, new polyketides synthesized by a type III polyketide synthase from Streptomyces reveromyceticus. J. Antibiot. (Tokyo) 2014, 67, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Čihák, M.; Kameník, Z.; Šmídová, K.; Bergman, N.; Benada, O.; Kofroňová, O.; Petříčková, K.; Bobek, J. Secondary metabolites produced during the germination of Streptomyces coelicolor. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Petersen, F.; Zähner, H.; Metzger, J.W.; Freund, S.; Hummel, R.P. Germicidin, an autoregulative germination inhibitor of Streptomyces viridochromogenes NRRL B-1551. J. Antibiot. (Tokyo) 1993, 46, 1126–1138. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.P.; Go, M.K.; Yew, W.S. Exploiting the biosynthetic potential of type III polyketide synthases. Molecules 2016, 21, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Chemler, J.A.; Buchholz, T.J.; Geders, T.W.; Akey, D.L.; Rath, C.M.; Chlipala, G.E.; Smith, J.L.; Sherman, D.H. Biochemical and structural characterization of germicidin synthase: Analysis of a type III polyketide synthase that employs acyl-ACP as a starter unit donor. J. Am. Chem. Soc. 2012, 134, 7359–7366. [Google Scholar] [CrossRef]
- Izumikawa, M.; Shipley, P.R.; Hopke, J.N.; O’Hare, T.; Xiang, L.; Noel, J.P.; Moore, B.S. Expression and characterization of the type III polyketide synthase 1,3,6,8-tetrahydroxynaphthalene synthase from Streptomyces coelicolor A3(2). J. Ind. Microbiol. Biotechnol. 2003, 30, 510–515. [Google Scholar] [CrossRef]
- Chen, H.; Tseng, C.C.; Hubbard, B.K.; Walsh, C.T. Glycopeptide antibiotic biosynthesis: Enzymatic assembly of the dedicated amino acid monomer (S)-3,5-dihydroxyphenylglycine. Proc. Natl. Acad. Sci. USA 2001, 98, 14901–14906. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, V.; Nicholson, G.J.; Ries, J.; Recktenwald, J.; Schefer, A.B.; Shawky, R.M.; Schröder, J.; Wohlleben, W.; Pelzer, S. A polyketide synthase in glycopeptide biosynthesis: The biosynthesis of the non-proteinogenic amino acid (S)-3,5-dihydroxyphenylglycine. J. Biol. Chem. 2001, 276, 38370–38377. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Scheme of the reaction occurring in polyketide synthases (PKSs). ACP, acyl carrier protein; AT, acyltransferase; KS, ketosynthase; KR, ketoreductase; DH, dehydratase; ER, enoylreductase. Adapted with permission of Portland Press, from Vance, S.; Tkachenko, O.; Thomas, B.; Bassuni, M.; Hong, H.; Nietlispach, D.; Broadhurst, W. Sticky swinging arm dynamics: studies of an acyl carrier protein domain from the mycolactone polyketide synthase. Biochem. J. 2016, 473, 1097–1110 [30].
Figure 1.
Scheme of the reaction occurring in polyketide synthases (PKSs). ACP, acyl carrier protein; AT, acyltransferase; KS, ketosynthase; KR, ketoreductase; DH, dehydratase; ER, enoylreductase. Adapted with permission of Portland Press, from Vance, S.; Tkachenko, O.; Thomas, B.; Bassuni, M.; Hong, H.; Nietlispach, D.; Broadhurst, W. Sticky swinging arm dynamics: studies of an acyl carrier protein domain from the mycolactone polyketide synthase. Biochem. J. 2016, 473, 1097–1110 [30].

Figure 2.
Structure of type I PKSs with three modules and 15 domains. ACP, acyl carrier protein; AT, acyltransferase; KS, ketosynthase; KR, ketoreductase; DH, dehydratase; ER, enoylreductase.
Figure 2.
Structure of type I PKSs with three modules and 15 domains. ACP, acyl carrier protein; AT, acyltransferase; KS, ketosynthase; KR, ketoreductase; DH, dehydratase; ER, enoylreductase.

Figure 3.
Some of the macrolides produced by Streptomyces.

Figure 4.
Biosynthesis of rapamycin. ACP, acyl carrier protein; AT, acyltransferase; KS, ketosynthase; KR, ketoreductase; DH, dehydratase; ER, enoylreductase; PIE, pipecolate-incorporating enzyme. Adapted with permission from Schwecke, T.; Aparicio, J.F.; Molnár, I.; König, A; Khaw, L.E.; Haydock, S.F.; Oliynyk, M.; Caffrey, P.; Cortés, J.; Lester, J.B. The biosynthetic gene cluster for the polyketide immunosuppressant rapamycin. Proc. Natl. Acad. Sci. USA 1995, 92, 7839–7843, doi:10.1073/pnas.92.17.7839 [34]. Copyright (1995) National Academy of Sciences, U.S.A. Adapted with permission of The Royal Society of Chemistry 2001, from Staunton, J.; Weissman, K.J. Polyketide biosynthesis: A millennium review. Nat. Prod. Rep. 2001, 18, 380–416 [26]; permission conveyed through Copyright Clearance Center, Inc.
Figure 4.
Biosynthesis of rapamycin. ACP, acyl carrier protein; AT, acyltransferase; KS, ketosynthase; KR, ketoreductase; DH, dehydratase; ER, enoylreductase; PIE, pipecolate-incorporating enzyme. Adapted with permission from Schwecke, T.; Aparicio, J.F.; Molnár, I.; König, A; Khaw, L.E.; Haydock, S.F.; Oliynyk, M.; Caffrey, P.; Cortés, J.; Lester, J.B. The biosynthetic gene cluster for the polyketide immunosuppressant rapamycin. Proc. Natl. Acad. Sci. USA 1995, 92, 7839–7843, doi:10.1073/pnas.92.17.7839 [34]. Copyright (1995) National Academy of Sciences, U.S.A. Adapted with permission of The Royal Society of Chemistry 2001, from Staunton, J.; Weissman, K.J. Polyketide biosynthesis: A millennium review. Nat. Prod. Rep. 2001, 18, 380–416 [26]; permission conveyed through Copyright Clearance Center, Inc.

Figure 5.
Some aromatic polyketides produced by Streptomyces.

Figure 6.
Biosynthesis of doxorubicin. Adapted with permission of The Royal Society of Chemistry 2009, from Chan, Y.A.; Podevels, A.M.; Kevany, B.M.; Thomas, M.G. Biosynthesis of polyketide synthase extender units. Nat. Prod. Rep. 2009, 26, 90–114 [60]; permission conveyed through Copyright Clearance Center, Inc.
Figure 6.
Biosynthesis of doxorubicin. Adapted with permission of The Royal Society of Chemistry 2009, from Chan, Y.A.; Podevels, A.M.; Kevany, B.M.; Thomas, M.G. Biosynthesis of polyketide synthase extender units. Nat. Prod. Rep. 2009, 26, 90–114 [60]; permission conveyed through Copyright Clearance Center, Inc.

Figure 7.
Some compounds produced by type III PKSs.

Figure 8.
Biosynthesis of germicidins. Gcs: germicidin synthase. Adapted with permission from Song, L.; Barona-Gomez, F.; Corre, C.; Xiang, L.; Udwary, D.W.; Austin, M.B.; Noel, J.P.; Moore, B.S.; Challis, G.L. Type III polyketide synthase β-ketoacyl-ACP starter unit and ethylmalonyl-coA extender unit selectivity discovered by Streptomyces coelicolor genome mining. J. Am. Chem. Soc. 2006, 128, 14754–14755 [91]. Copyright 2006 American Chemical Society.
Figure 8.
Biosynthesis of germicidins. Gcs: germicidin synthase. Adapted with permission from Song, L.; Barona-Gomez, F.; Corre, C.; Xiang, L.; Udwary, D.W.; Austin, M.B.; Noel, J.P.; Moore, B.S.; Challis, G.L. Type III polyketide synthase β-ketoacyl-ACP starter unit and ethylmalonyl-coA extender unit selectivity discovered by Streptomyces coelicolor genome mining. J. Am. Chem. Soc. 2006, 128, 14754–14755 [91]. Copyright 2006 American Chemical Society.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Some polyketides produced by Streptomyces and their type I PKSs.
| Polyketide | Structure | Producer | Type I PKSs | Ref. |
|---|---|---|---|---|
| Avermectin | 16-membered ring macrolide | Streptomyces avermitilis | AVES1-4 | [41] |
| Chalcomycin | 16-membered ring macrolide | Streptomyces bikiniensis | ChmGI-V | [42] |
| Candicidin | 38-membered ring polyene macrolide | Streptomyces griseus IMRU 3570 | CanP1-3, and CanPF | [43,44] |
| FK506 (Tacrolimus) | 23-membered ring macrolide | Streptomyces tsukubaensis, Streptomyces sp. MA6858 | FkbABC | [35,45] |
| FK520 (Ascomycin) | 23-membered ring macrolide | Streptomyces hygroscopicus var. ascomyceticus | FkbABC | [46] |
| Methymycin, Neomethymycin, Narbomycin, Pikromycin | 12-membered ring macrolide, 12-membered ring macrolide, 14-membered ring macrolide, 14- membered ring macrolide | Streptomyces venezuelae | PikAI-IV | [37] |
| Pimaricin | 26-membered ring polyene macrolide | Streptomyces natalensis | PIMS0 and PIMS1 | [47] |
| Rapamycin | 31- membered ring macrolide | Streptomyces hygroscopicus | RAPS1-3 | [34] |
| Spiramycin | 16- membered ring macrolide | Streptomyces ambofaciens | SrmGI-V | [36] |
| Tautomycetin | Linear | Streptomyces sp. CK4412 | TmcA and TmcB | [32] |
| Tylosin | 16- membered ring macrolide | Streptomyces fradiae | TYLGI-V | [48] |
Table 2.
Some polyketides produced by Streptomyces and their minimal type II PKSs.
| Polyketide | Intermediate Backbone Structure | Producer | Minimal Type II PKSs | Ref. |
|---|---|---|---|---|
| Medermycin | octaketide | Streptomyces sp. K73 | Med-1,2,23 | [69,70] |
| Doxorubicin | decaketide | Streptomyces peucetius | DpsABCDG | [60,65] |
| Oxytetracycline | decaketide | Streptomyces rimosus | OxyABCD | [71] |
| Gilvocarcin | decaketide | Streptomyces griseoflavus Gö 3592 | GilABC | [72] |
| Oviedomycin | decaketide | Streptomyces antibioticus | OvmPKS | [73] |
| Chartreusin | decaketide | Streptomyces chartreusis | ChaABC | [74] |
| Cervimycin | decaketide | Streptomyces tendae HKI-179 | CerABC | [75,76] |
| Resistomycin | decaketide | Streptomyces resistomycificus | RemABC | [77] |
| Chromomycin | decaketide | Streptomyces griseus subsp. griseus | CmmPKS | [78,79] |
| Hedamycin | dodecaketide | Streptomyces griseoruber | HedCDE | [80,81] |
| Fredericamycin | pentadecaketide | Streptomyces griseus ATCC 49344 | FdmFSGH | [82,83] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Risdian, C.; Mozef, T.; Wink, J. Biosynthesis of Polyketides in Streptomyces. Microorganisms 2019, 7, 124. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7050124
AMA Style
Risdian C, Mozef T, Wink J. Biosynthesis of Polyketides in Streptomyces. Microorganisms. 2019; 7(5):124. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7050124
Chicago/Turabian StyleRisdian, Chandra, Tjandrawati Mozef, and Joachim Wink. 2019. "Biosynthesis of Polyketides in Streptomyces" Microorganisms 7, no. 5: 124. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7050124
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.