Short Communication: Preliminary Differences Identified in Genes Responsible for Biofilm Formation in Poultry Isolates of Salmonella enterica Heidelberg, Enteritidis, and Kentucky


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Pellicle Formation
2.2. Quantitative Reverse-Transcriptase PCR (qRT-PCR) Assay
2.3. Statistical Analysis
3. Results
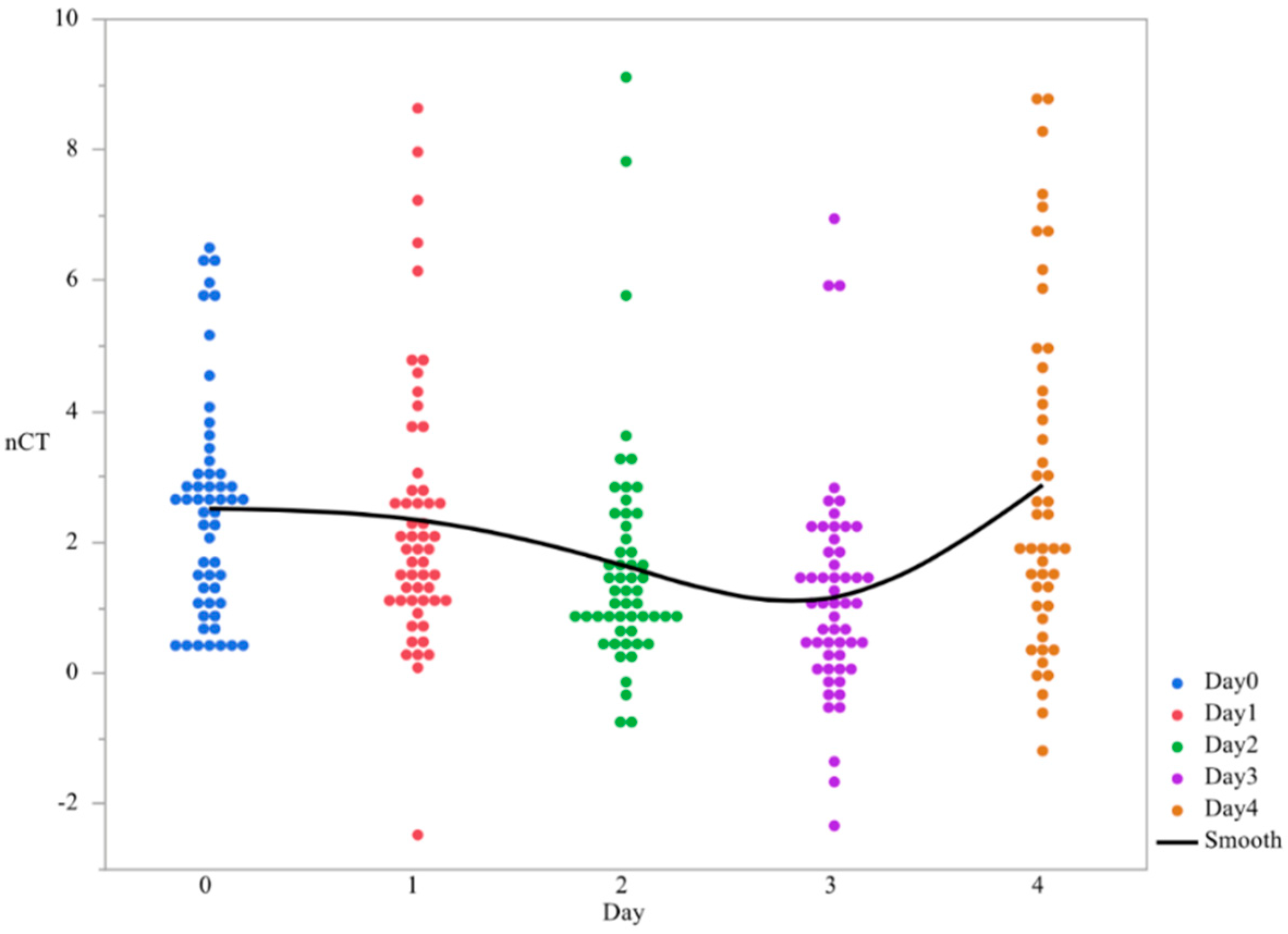
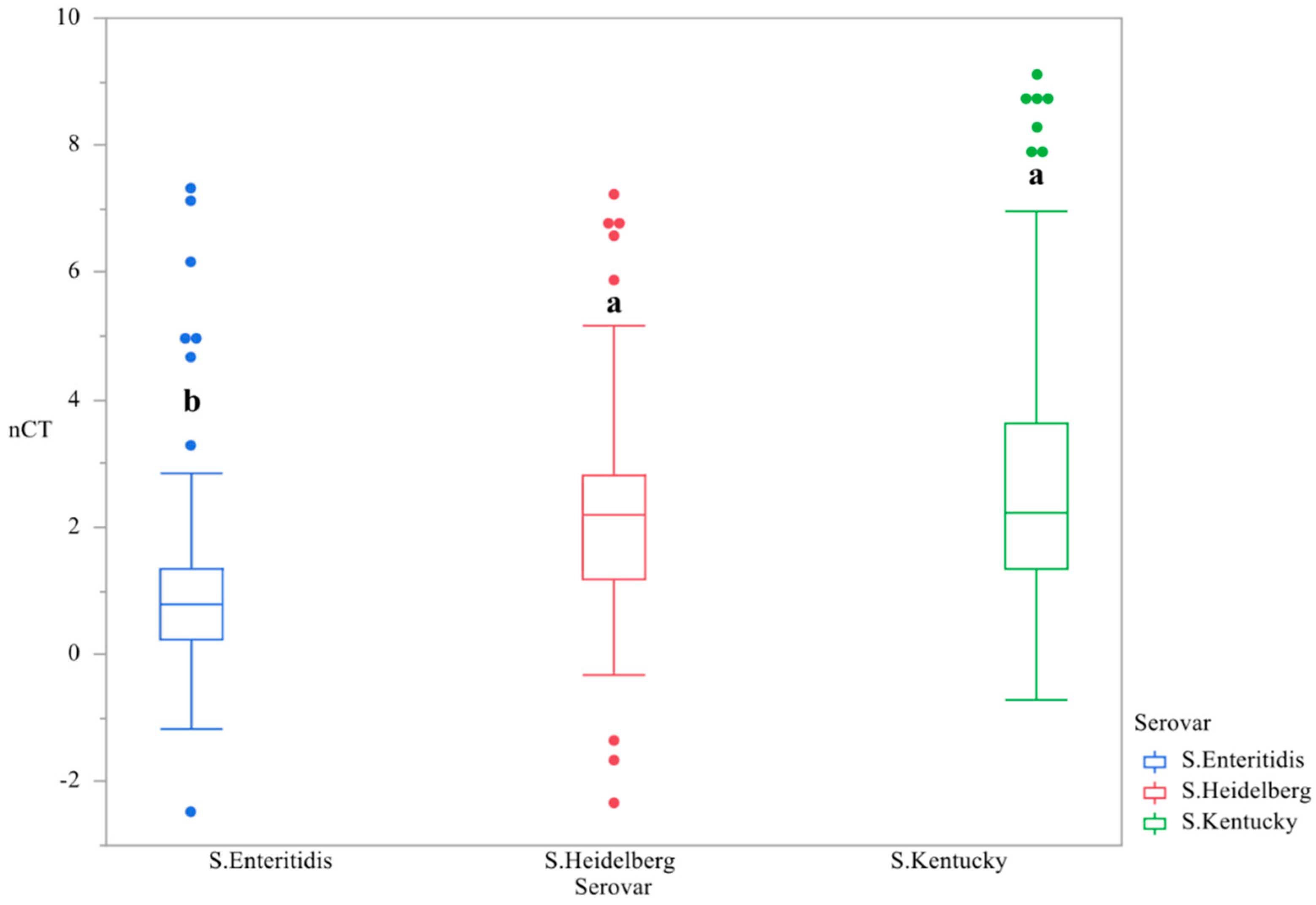
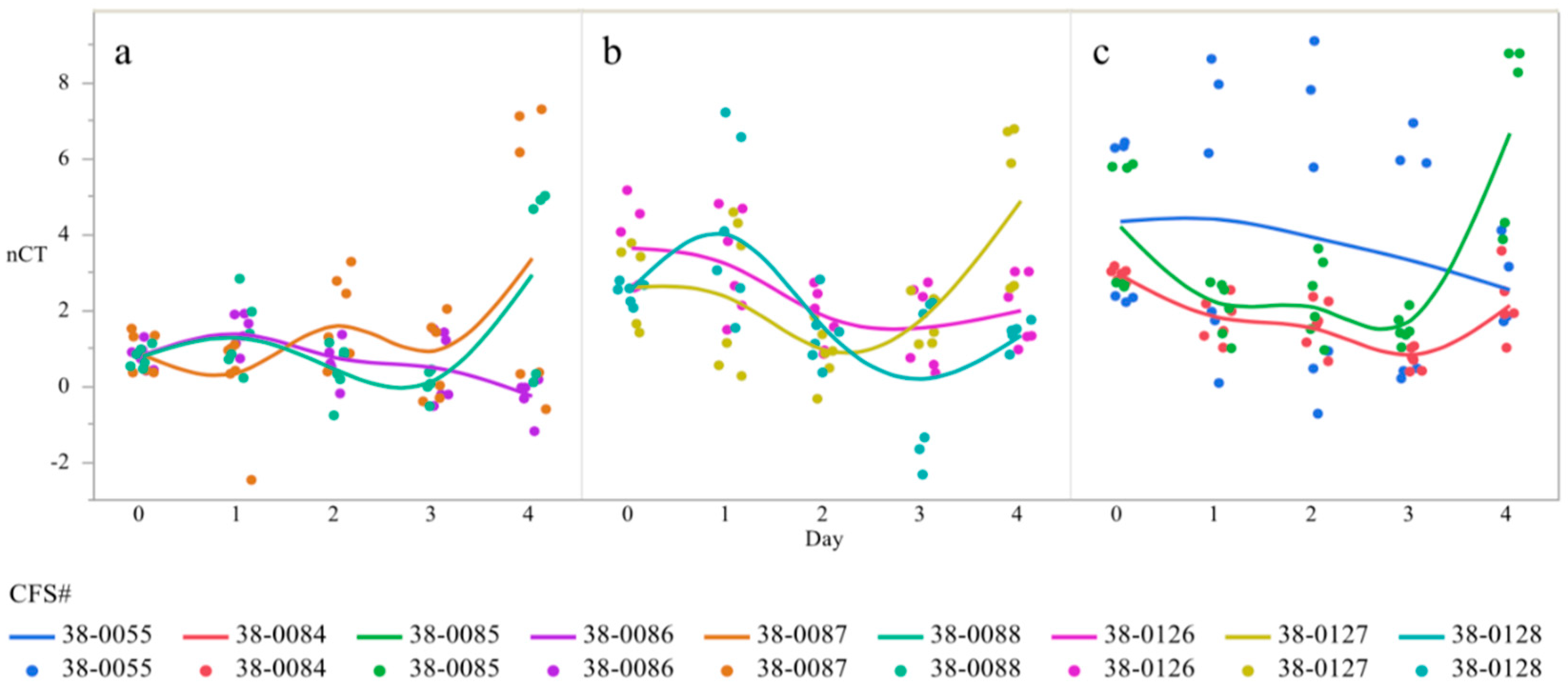
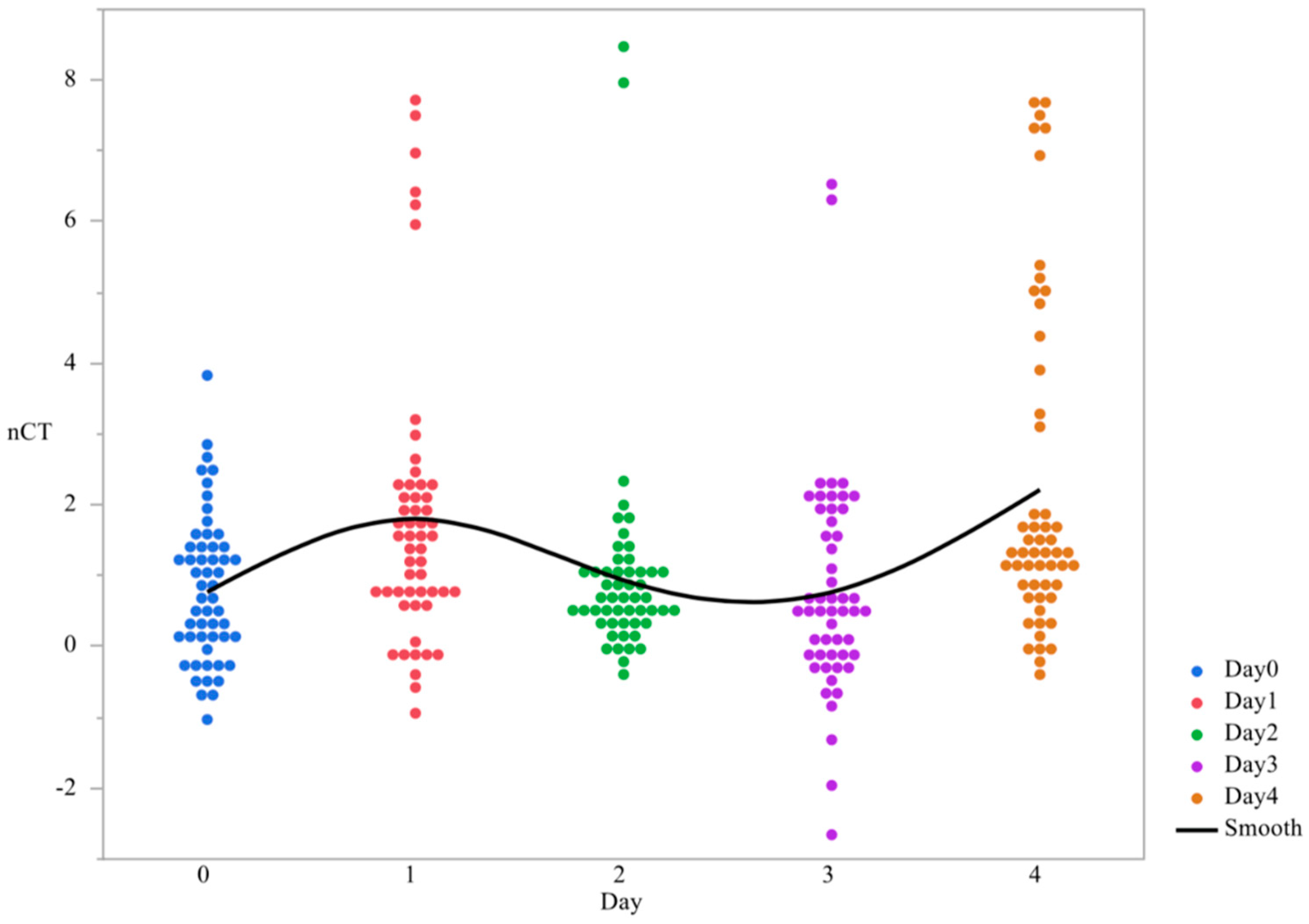
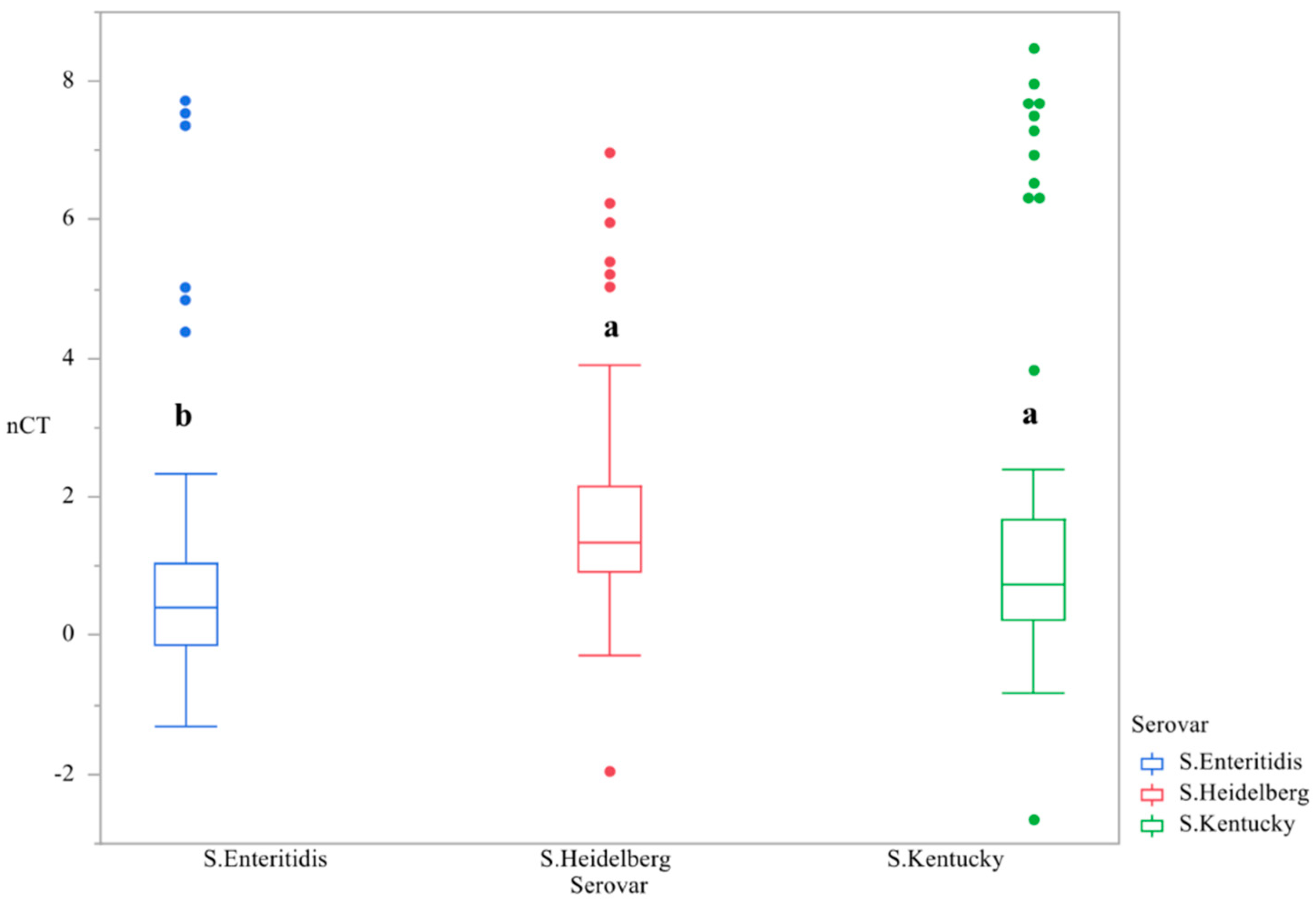
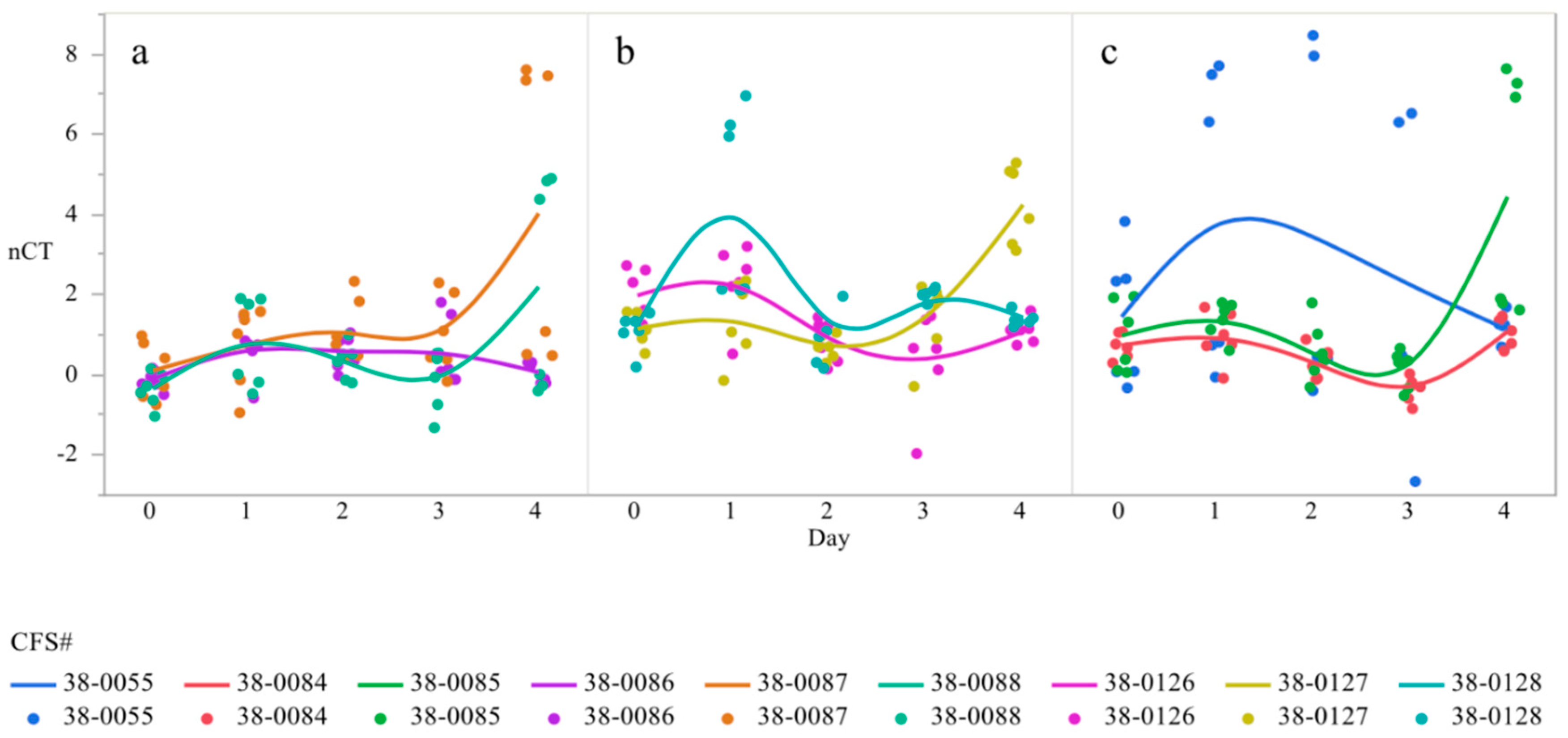
3.1. CsgD Expression
3.2. BcsA Expression
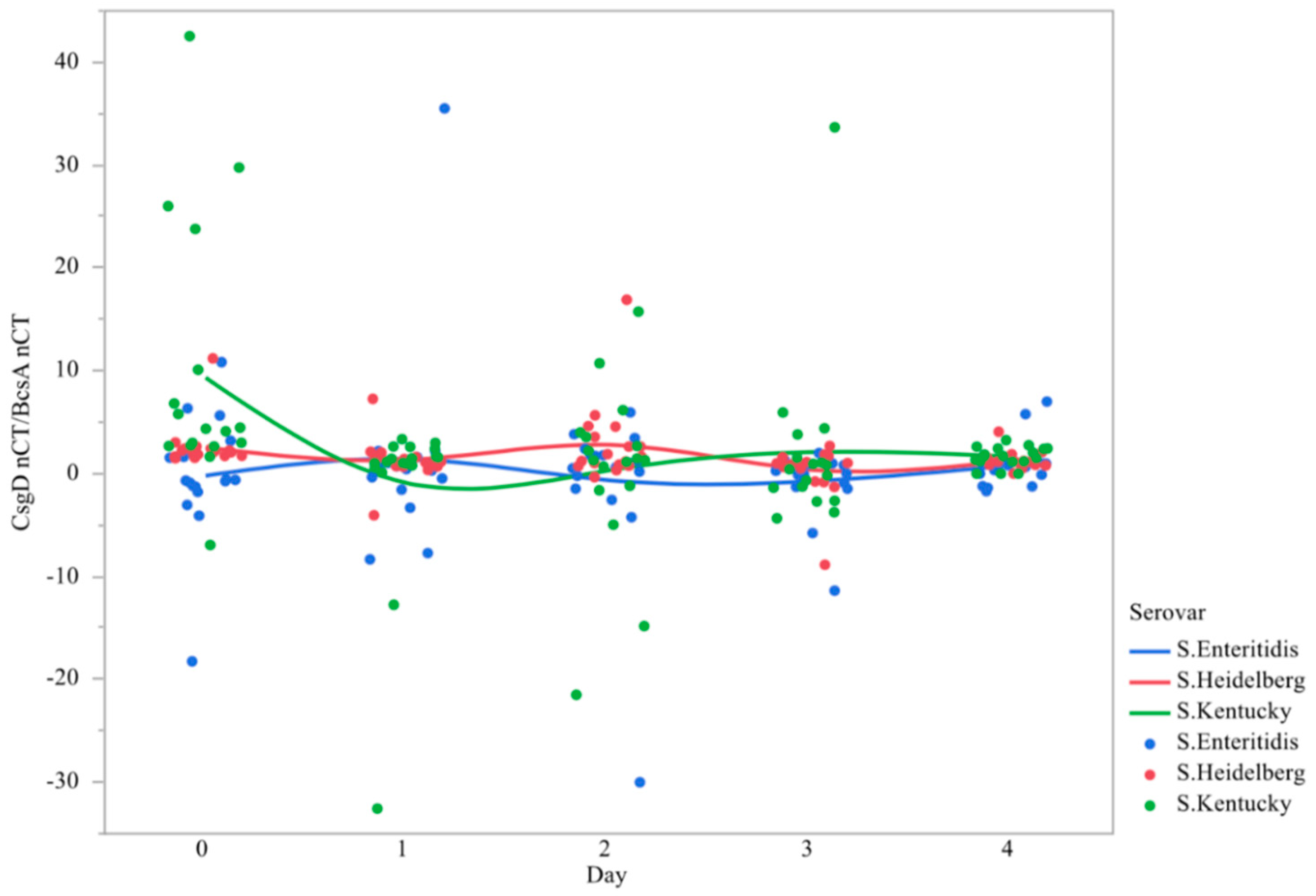
3.3. Ratio of csgD to bcsA Expression
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.L.; Lynne, A.M.; Nayak, R. Salmonella challenges: Prevalence in swine and poultry and potential pathogenicity of such isolates. J. Anim. Sci. 2008, 86, E149–E162. [Google Scholar] [CrossRef] [PubMed]
- Hanning, I.B.; Nutt, J.D.; Ricke, S.C. Salmonellosis outbreaks in the United States due to fresh produce: Sources and potential intervention measures. Foodborne Pathog. Dis. 2009, 6, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.L.; Nayak, R.; Hanning, I.B.; Johnson, T.J.; Han, J.; Ricke, S.C. Population dynamics of Salmonella enterica serotypes in commercial egg and poultry production. Appl. Environ. Microbiol. 2011, 77, 4273–4279. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella pathogenicity and host adaptation in chicken-associated serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA); Food Safety and Inspection Service (FSIS). Serotypes Profile of Salmonella Isolates from Meat and Poultry Products, January 1998 through December 2014; United States Department of Agriculture, 2016. Available online: https://www.fsis.usda.gov/wps/wcm/connect/3866026a-582d-4f0e-a8ce-851b39c7390f/Salmonella-Serotype-Annual-2014.pdf?MOD=AJPERES (accessed on 5 April 2019).
- Finstad, S.; O'Bryan, C.A.; Marcy, J.A.; Crandall, P.G.; Ricke, S.C. Salmonella and broiler processing in the United States: Relationship to foodborne salmonellosis. Food Res. Int. 2012, 45, 789–794. [Google Scholar] [CrossRef]
- Howard, Z.R.; O'Bryan, C.A.; Crandall, P.G.; Ricke, S.C. Salmonella Enteritidis in shell eggs: Current issues and prospects for control. Food Res. Int. 2012, 45, 755–764. [Google Scholar] [CrossRef]
- Rabsch, W.; Hargis, B.M.; Tsolis, R.M.; Kingsley, R.A.; Hinz, K.-H.; Tschäpe, H.; Bäumler, A.J. Competitive exclusion of Salmonella enteritidis by Salmonella gallinarum in Poultry. Emerg. Infect. Dis. 2000, 6, 443–448. [Google Scholar] [CrossRef]
- Ricke, S.C. Insights and challenges of Salmonella infection of laying hens. Curr. Opin. Food Sci. 2017, 18, 43–49. [Google Scholar] [CrossRef]
- Gast, R.K.; Guard-Bouldin, J.; Holt, P.S. Colonization of reproductive organs and internal contamination of eggs after experimental infection of laying hens with Salmonella heidelberg and Salmonella enteritidis. Avian Dis. 2004, 48, 863–869. [Google Scholar] [CrossRef]
- Gast, R.K.; Guraya, R.; Guard-Bouldin, J.; Holt, P.S.; Moore, R.W. Colonization of specific regions of the reproductive tract and deposition at different locations inside eggs laid by hens infected with Salmonella Enteritidis or Salmonella Heidelberg. Avian Dis. 2007, 51, 40–44. [Google Scholar] [CrossRef]
- Kaldhone, P.R.; Foley, S.L.; Ricke, S.C. Salmonella Heidelberg in layer hens and egg production: Incidence and potential issues. In Producing Safe. Eggs; Ricke, S.C., Gast, R.K., Eds.; Elsevier Inc.: San Diego, CA, USA, 2017; Chapter 12; pp. 235–256. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/B9780128025826000124 (accessed on 1 May 2019).
- Centers for Disease Control and Prevention (CDC). Reports of Selected Salmonella Outbreak Investigations 2018. Available online: https://www.cdc.gov/salmonella/outbreaks.html (accessed on 15 April 2019).
- Kingsley, R.A.; Bäumler, A.J. Host adaptation and the emergence of infectious disease: The Salmonella paradigm. Mol. Microbiol. 2000, 36, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Uzzau, S.; Brown, D.J.; Wallis, T.; Rubino, S.; Leori, G.; Bernard, S.; Casadesús, J.; Platt, D.J.; Olsen, J.E. Host adapted serotypes of Salmonella enterica. Epidemiol. Infect. 2000, 125, 229–255. [Google Scholar] [CrossRef] [PubMed]
- McClelland, M.; Sanderson, K.E.; Clifton, S.W.; Latreille, P.; Porwollik, S.; Sabo, A.; Meyer, R.; Bieri, T.; Ozersky, P.; McLellan, M. Comparison of genome degradation in Paratyphi A and Typhi, human-restricted serovars of Salmonella enterica that cause typhoid. Nat. Genet. 2004, 36, 1268. [Google Scholar] [CrossRef] [PubMed]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal Salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Donlan, R.M. Biofilms: Microbial Life on Surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Steenackers, H.; Hermans, K.; Vanderleyden, J.; De Keersmaecker, S.C.J. Salmonella biofilms: An overview on occurrence, structure, regulation and eradication. Food Res. Int. 2012, 45, 502–531. [Google Scholar] [CrossRef]
- Vestby, L.K.; Møretrø, T.; Langsrud, S.; Heir, E.; Nesse, L.L. Biofilm forming abilities of Salmonella are correlated with persistence in fish meal-and feed factories. BMC Vet. Res. 2009, 5, 20. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Solano, C.; García, B.; Valle, J.; Berasain, C.; Ghigo, J.M.; Gamazo, C.; Lasa, I. Genetic analysis of Salmonella enteritidis biofilm formation: Critical role of cellulose. Mol. Microbiol. 2002, 43, 793–808. [Google Scholar] [CrossRef]
- Zogaj, X.; Nimtz, M.; Rohde, M.; Bokranz, W.; Römling, U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 2001, 39, 1452–1463. [Google Scholar] [CrossRef] [PubMed]
- White, A.; Gibson, D.; Kim, W.; Kay, W.; Surette, M. Thin aggregative fimbriae and cellulose enhance long-term survival and persistence of Salmonella. J. Bacteriol. 2006, 188, 3219–3227. [Google Scholar] [CrossRef] [PubMed]
- Barnhart, M.M.; Chapman, M.R. Curli biogenesis and function. Annu. Rev. Microbiol. 2006, 60, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Verstraeten, N.; Braeken, K.; Debkumari, B.; Fauvart, M.; Fransaer, J.; Vermant, J.; Michiels, J. Living on a surface: Swarming and biofilm formation. Trends Microbiol. 2008, 16, 496–506. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | |||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |
| CsgD nCT | 2.157 ± 0.231 ab | 2.346 ± 0.285 ab | 1.641 ± 0.242 bc | 1.151 ± 0.226 c | 2.884 ± 0.387 a |
| Day | |||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |
| S. Enteritidis | |||||
| 38-0086 | 0.788 ± 0.139 abc | 1.442 ± 0.190 abc | 0.667 ± 0.21 bc | 0.53 ± 0.385 bc | −0.28 ± 0.24 c |
| 38-0087 | 0.887 ± 0.227 abc | 0.218 ± 0.553 bc | 1.843 ± 0.472 abc | 0.722 ± 0.437 abc | 3.449 ± 1.545 a |
| 38-0088 | 0.762 ± 0.108 abc | 1.328 ± 0.389 abc | 0.438 ± 0.284 bc | 0.057 ± 0.141 c | 3.008 ± 1.142 ab |
| S. Heidelberg | |||||
| 38-0126 | 3.617 ± 0.461 abc | 3.267 ± 0.565 abcd | 1.767 ± 0.317 bcde | 1.552 ± 0.451 bcde | 2.002 ± 0.374 bcde |
| 38-0127 | 2.575 ± 0.452 abcde | 2.43 ± 0.81 abcde | 0.86 ± 0.305 de | 1.794 ± 0.261 bcde | 4.928 ± 0.956 a |
| 38-0128 | 2.483 ± 0.111 abcde | 4.182 ± 0.928 ab | 1.363 ± 0.343 cde | 0.155 ± 0.876 e | 1.383 ± 0.152 bcde |
| S. Kentucky | |||||
| 38-0055 | 4.333 ± 0.904 ab | 4.425 ± 1.476 ab | 3.898 ± 1.714 ab | 3.313 ± 1.33 ab | 2.538 ± 0.474 ab |
| 38-0084 | 2.955 ± 0.068 ab | 1.752 ± 0.235 b | 1.628 ± 0.263 b | 0.719 ± 0.117 b | 2.186 ± 0.421 b |
| 38-0085 | 4.256 ± 0.692 ab | 2.068 ± 0.296 b | 2.308 ± 0.427 b | 1.522 ± 0.155 b | 6.809 ± 1.114 a |
| Day | |||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |
| BcsA nCT | 0.766 ± 0.138 c | 1.815 ± 0.273 ab | 0.932 ± 0.219 bc | 0.766 ± 0.22 c | 2.217 ± 0.319 a |
| Day | |||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |
| S. Enteritidis | |||||
| 38-0086 | −0.115 ± 0.101 b | 0.645 ± 0.275 b | 0.557 ± 0.175 b | 0.558 ± 0.354 b | 0.059 ± 0.105 b |
| 38-0087 | 0.107 ± 0.296 b | 0.742 ± 0.423 b | 1.14 ± 0.315 b | 1.022 ± 0.402 b | 4.089 ± 1.521 a |
| 38-0088 | −0.372 ± 0.175 b | 0.823 ± 0.467 b | 0.343 ± 0.184 b | −0.103 ± 0.314 b | 2.247 ± 1.105 ab |
| S. Heidelberg | |||||
| 38-0126 | 1.948 ± 0.284 bc | 2.313 ± 0.39 b | 0.852 ± 0.219 bc | 0.392 ± 0.513 c | 1.097 ± 0.124 bc |
| 38-0127 | 1.133 ± 0.162 bc | 1.39 ± 0.402 bc | 0.702 ± 0.118 bc | 1.433 ± 0.39 bc | 4.281 ± 0.4 a |
| 38-0128 | 1.098 ± 0.194 bc | 4.263 ± 0.961 a | 0.899 ± 0.321 bc | 2.017 ± 0.071 bc | 1.39 ± 0.068 bc |
| S. Kentucky | |||||
| 38-0055 | 1.403 ± 0.688 abc | 3.842 ± 1.511 ab | 3.382 ± 1.982 abc | 2.228 ± 1.805 abc | 1.134 ± 0.159 abc |
| 38-0084 | 0.727 ± 0.128 abc | 0.938 ± 0.26 abc | 0.332 ± 0.16 bc | −0.359 ± 0.126 c | 1.126 ± 0.149 abc |
| 38-0085 | 0.963 ± 0.361 abc | 1.378 ± 0.183 abc | 0.587 ± 0.302 abc | 0.16 ± 0.192 bc | 4.532 ± 1.237 a |
| Day | |||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |
| S. Enteritidis | −0.230 ± 1.450 b | 1.485 ± 2.126 b | −0.696 ± 1.808 b | 0.741 ± 0.741 b | 0.768 ± 0.530 b |
| S. Heidelberg | 2.575 ± 0.516 ab | 1.235 ± 0.472 b | 2.936 ± 0.963 ab | 0.2638 ± 0.620 b | 1.384 ± 0.205 b |
| S. Kentucky | 9.400 ± 2.960 a | −1.083 ± 2.026 b | 0.406 ± 2.062 b | 2.101 ± 2.087 ab | 1.646 ± 0.227 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Z.; Dittoe, D.K.; Feye, K.M.; Kogut, M.; Ricke, S.C. Short Communication: Preliminary Differences Identified in Genes Responsible for Biofilm Formation in Poultry Isolates of Salmonella enterica Heidelberg, Enteritidis, and Kentucky. Microorganisms 2019, 7, 196. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070196
Shi Z, Dittoe DK, Feye KM, Kogut M, Ricke SC. Short Communication: Preliminary Differences Identified in Genes Responsible for Biofilm Formation in Poultry Isolates of Salmonella enterica Heidelberg, Enteritidis, and Kentucky. Microorganisms. 2019; 7(7):196. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070196
Chicago/Turabian StyleShi, Zhaohao, Dana K. Dittoe, Kristina M. Feye, Mike Kogut, and Steven C. Ricke. 2019. "Short Communication: Preliminary Differences Identified in Genes Responsible for Biofilm Formation in Poultry Isolates of Salmonella enterica Heidelberg, Enteritidis, and Kentucky" Microorganisms 7, no. 7: 196. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7070196