Bioactive Compounds of Nigella Sativa Essential Oil as Antibacterial Agents against Chlamydia Trachomatis D

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Cell Line
2.2. Essential Oil and Active Compounds
2.3. Cytotoxicity Assay
2.4. Anti-Chlamydial Assay
2.5. Determination of Minimal Inhibitory Concentrations
2.6. Statistical Analysis
3. Results
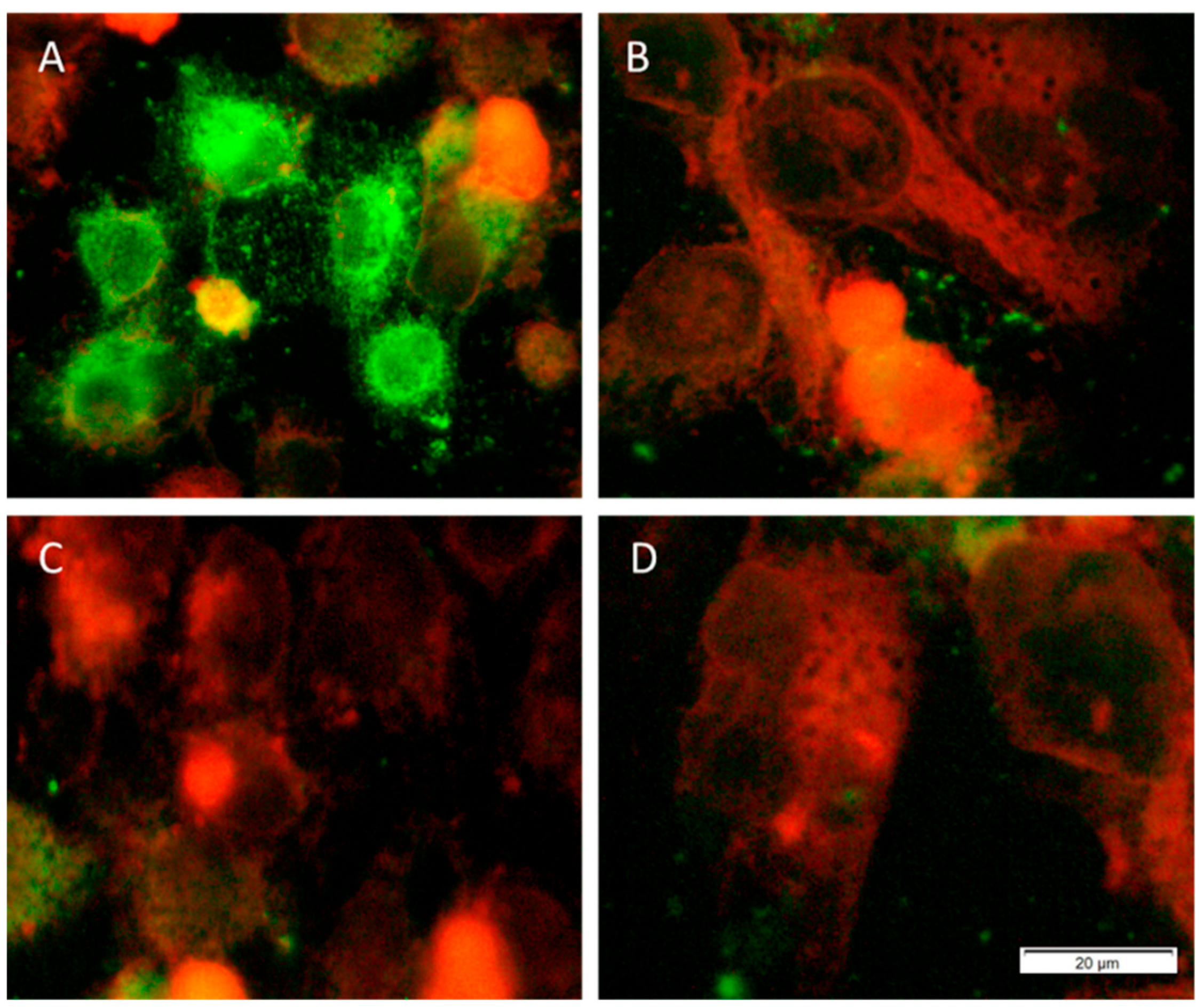
3.1. Cytotoxicity Assay
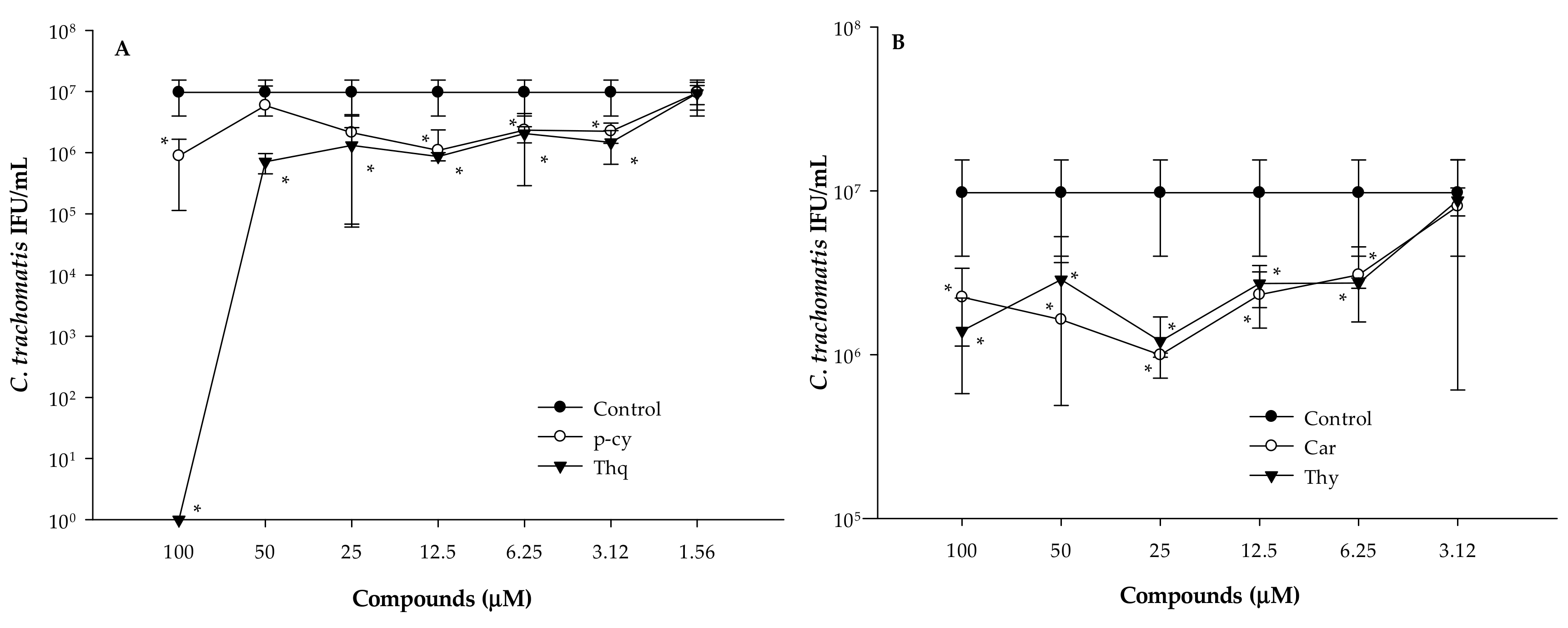
3.2. Anti-Chlamydial Assay
3.3. Determination of Minimal Inhibitory Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Allcock, S.; Young, E.H.; Holmes, M.; Gurdasani, D.; Dougan, G.; Sandhu, M.S.; Solomon, L.; Török, M.E. Antimicrobial resistance in human populations: Challenges and opportunities. Glob. Health Epidemiol. Genom. 2017, 2, e4. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.R.; Rosenthal, K.S.; Pfaller, M.A. Medical Microbiology, 8th ed.; Elsevier: Philadelphia, PA, USA, 2016; ISBN 978-0-323-29956-5. [Google Scholar]
- Elwell, C.; Mirrashidi, K.; Engel, J. Chlamydia cell biology and pathogenesis. Nat. Rev. Microbiol. 2016, 14, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Mestrovic, T.; Ljubin-Sternak, S. Molecular mechanisms of Chlamydia trachomatis resistance to antimicrobial drugs. Front. Biosci. 2018, 23, 656–670. [Google Scholar] [CrossRef]
- Binet, R.; Maurelli, A.T. Frequency of Development and Associated Physiological Cost of Azithromycin Resistance in Chlamydia psittaci 6BC and C. trachomatis L2. Antimicrob. Agents Chemother. 2007, 51, 4267–4275. [Google Scholar] [CrossRef] [PubMed]
- Dessus-Babus, S.; Bébéar, C.M.; Charron, A.; Bébéar, C.; de Barbeyrac, B. Sequencing of gyrase and topoisomerase IV quinolone-resistance-determining regions of Chlamydia trachomatis and characterization of quinolone-resistant mutants obtained In vitro. Antimicrob. Agents Chemother. 1998, 42, 2474–2481. [Google Scholar] [CrossRef] [PubMed]
- Marti, H.; Kim, H.; Joseph, S.J.; Dojiri, S.; Read, T.D.; Dean, D. Tet(C) Gene Transfer between Chlamydia suis Strains Occurs by Homologous Recombination after Co-infection: Implications for Spread of Tetracycline-Resistance among Chlamydiaceae. Front. Microbiol. 2017, 8, 156. [Google Scholar] [CrossRef]
- Schoborg, R.V. Chlamydia persistence—A tool to dissect chlamydia–host interactions. Microbes Infect. 2011, 13, 649–662. [Google Scholar] [CrossRef]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- Warnke, P.H.; Becker, S.T.; Podschun, R.; Sivananthan, S.; Springer, I.N.; Russo, P.A.J.; Wiltfang, J.; Fickenscher, H.; Sherry, E. The battle against multi-resistant strains: Renaissance of antimicrobial essential oils as a promising force to fight hospital-acquired infections. J. Craniomaxillofac. Surg. 2009, 37, 392–397. [Google Scholar] [CrossRef]
- Forouzanfar, F.; Bazzaz, B.S.F.; Hosseinzadeh, H. Black cumin (Nigella sativa) and its constituent (thymoquinone): A review on antimicrobial effects. Iran. J. Basic Med. Sci. 2014, 17, 929–938. [Google Scholar]
- Mouwakeh, A.; Kincses, A.; Nové, M.; Mosolygó, T.; Mohácsi-Farkas, C.; Kiskó, G.; Spengler, G. Nigella sativa essential oil and its bioactive compounds as resistance modifiers against Staphylococcus aureus. Phytother. Res. 2019, 33, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Shaaban, H.A.; Sadek, Z.; Edris, A.E.; Saad-Hussein, A. Analysis and antibacterial activity of Nigella sativa essential oil formulated in microemulsion system. J. Oleo Sci. 2015, 64, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Mouwakeh, A.; Telbisz, Á.; Spengler, G.; Mohácsi-Farkas, C.; Kiskó, G. Antibacterial and Resistance Modifying Activities of Nigella sativa Essential Oil and its Active Compounds Against Listeria monocytogenes. In Vivo 2018, 32, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Marchese, A.; Orhan, I.E.; Daglia, M.; Barbieri, R.; Di Lorenzo, A.; Nabavi, S.F.; Gortzi, O.; Izadi, M.; Nabavi, S.M. Antibacterial and antifungal activities of thymol: A brief review of the literature. Food Chem. 2016, 210, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Sessa, R.; Di Pietro, M.; De Santis, F.; Filardo, S.; Ragno, R.; Angiolella, L. Effects of Mentha suaveolens essential oil on Chlamydia trachomatis. BioMed Res. Int. 2015, 2015, 508071. [Google Scholar] [CrossRef] [PubMed]
- Burián, K.; Hegyesi, H.; Buzás, E.; Endrész, V.; Kis, Z.; Falus, A.; Gönczöl, E. Chlamydophila (Chlamydia) pneumoniae induces histidine decarboxylase production in the mouse lung. Immunol. Lett. 2003, 89, 229–236. [Google Scholar] [CrossRef]
- Balogh, E.P.; Faludi, I.; Virók, D.P.; Endrész, V.; Burián, K. Chlamydophila pneumoniae induces production of the defensin-like MIG/CXCL9, which has in vitro antichlamydial activity. Int. J. Med. Microbiol. 2011, 301, 252–259. [Google Scholar] [CrossRef]
- Raffai, T.; Burián, K.; Janovák, L.; Bogdanov, A.; Hegemann, J.H.; Endrész, V.; Virok, D.P. Vaginal Gel Component Hydroxyethyl Cellulose Significantly Enhances the Infectivity of Chlamydia trachomatis Serovars D and E. Antimicrob. Agents Chemother. 2019, 63, e02034–e020318. [Google Scholar] [CrossRef] [PubMed]
- Żesławska, E.; Kincses, A.; Unger, V.; Tóth, V.; Spengler, G.; Nitek, W.; Tejchman, W. Exocyclic Sulfur and Selenoorganic Compounds Towards Their Anticancer Effects: Crystallographic and Biological Studies. Anticancer Res. 2018, 38, 4577–4584. [Google Scholar] [CrossRef] [PubMed]
- Eszik, I.; Lantos, I.; Önder, K.; Somogyvári, F.; Burián, K.; Endrész, V.; Virok, D.P. High dynamic range detection of Chlamydia trachomatis growth by direct quantitative PCR of the infected cells. J. Microbiol. Methods 2016, 120, 15–22. [Google Scholar] [CrossRef]
- Newman, L.; Rowley, J.; Vander Hoorn, S.; Wijesooriya, N.S.; Unemo, M.; Low, N.; Stevens, G.; Gottlieb, S.; Kiarie, J.; Temmerman, M. Global Estimates of the Prevalence and Incidence of Four Curable Sexually Transmitted Infections in 2012 Based on Systematic Review and Global Reporting. PLoS ONE 2015, 10, e0143304. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zheng, H.; Mai, Z.; Qin, X.; Chen, W.; Huang, T.; Chen, D.; Zheng, L. An in vitro model of azithromycin-induced persistent Chlamydia trachomatis infection. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Kincses, A.; Varga, B.; Csonka, Á.; Sancha, S.; Mulhovo, S.; Madureira, A.M.; Ferreira, M.-J.U.; Spengler, G. Bioactive compounds from the African medicinal plant Cleistochlamys kirkii as resistance modifiers in bacteria. Phytother. Res. 2018, 32, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.J.; Kincses, A.; Gajdács, M.; Spengler, G.; dos Santos, D.J.V.A.; Molnár, J.; Ferreira, M.-J.U. Terpenoids from Euphorbia pedroi as Multidrug-Resistance Reversers. J. Nat. Prod. 2018, 81, 2032–2040. [Google Scholar] [CrossRef] [PubMed]
- Burt, S.A.; Ojo-Fakunle, V.T.A.; Woertman, J.; Veldhuizen, E.J.A. The Natural Antimicrobial Carvacrol Inhibits Quorum Sensing in Chromobacterium violaceum and Reduces Bacterial Biofilm Formation at Sub-Lethal Concentrations. PLoS ONE 2014, 9, e93414. [Google Scholar] [CrossRef]
- Marchese, A.; Arciola, C.R.; Barbieri, R.; Silva, A.S.; Nabavi, S.F.; Tsetegho Sokeng, A.J.; Izadi, M.; Jafari, N.J.; Suntar, I.; Daglia, M.; et al. Update on Monoterpenes as Antimicrobial Agents: A Particular Focus on p-Cymene. Materials 2017, 10, 947. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 |
|---|---|
| p-cymene | >100 μM |
| thymoquinone | >100 μM |
| carvacrol | >100 μM |
| thymol | >100 μM |
| N. sativa essential oil | 0.009% (v/v) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mosolygó, T.; Mouwakeh, A.; Hussein Ali, M.; Kincses, A.; Mohácsi-Farkas, C.; Kiskó, G.; Spengler, G. Bioactive Compounds of Nigella Sativa Essential Oil as Antibacterial Agents against Chlamydia Trachomatis D. Microorganisms 2019, 7, 370. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090370
Mosolygó T, Mouwakeh A, Hussein Ali M, Kincses A, Mohácsi-Farkas C, Kiskó G, Spengler G. Bioactive Compounds of Nigella Sativa Essential Oil as Antibacterial Agents against Chlamydia Trachomatis D. Microorganisms. 2019; 7(9):370. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090370
Chicago/Turabian StyleMosolygó, Tímea, Ahmad Mouwakeh, Munira Hussein Ali, Annamária Kincses, Csilla Mohácsi-Farkas, Gabriella Kiskó, and Gabriella Spengler. 2019. "Bioactive Compounds of Nigella Sativa Essential Oil as Antibacterial Agents against Chlamydia Trachomatis D" Microorganisms 7, no. 9: 370. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7090370