Vegetable By-Product Lacto-Fermentation as a New Source of Antimicrobial Compounds

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Tomato, Carrot, and Melon By-Products Lacto-Fermentation
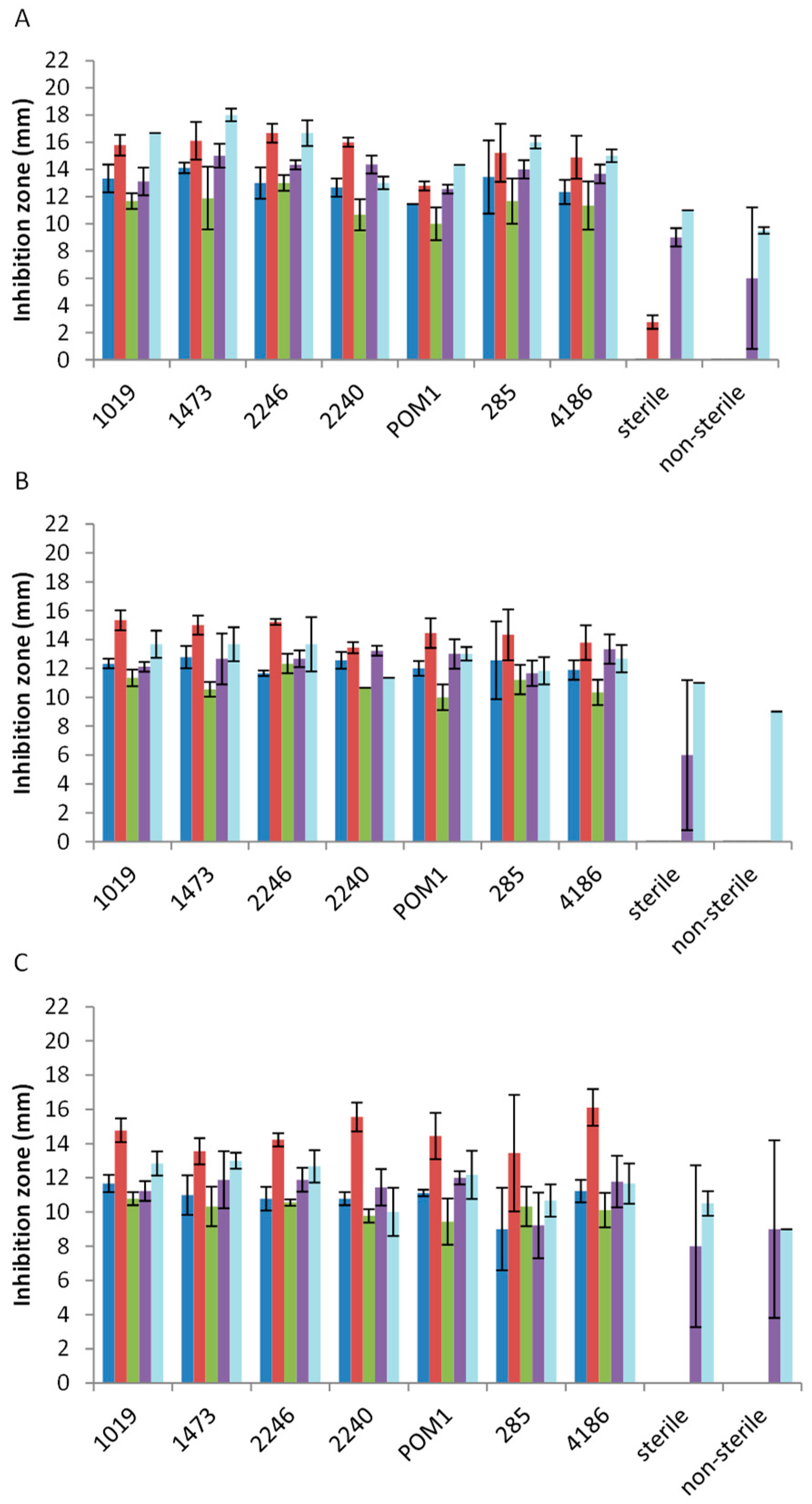
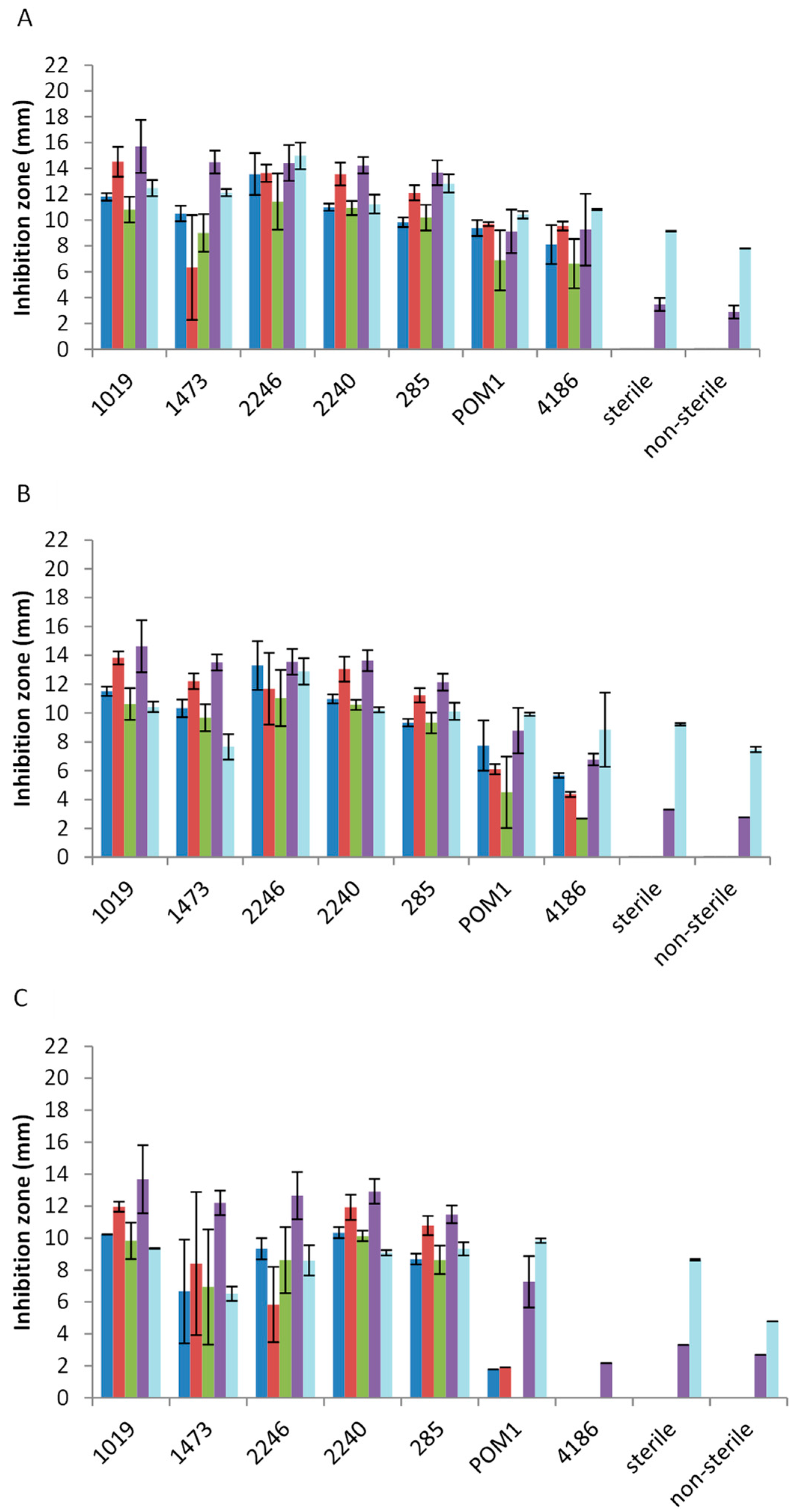
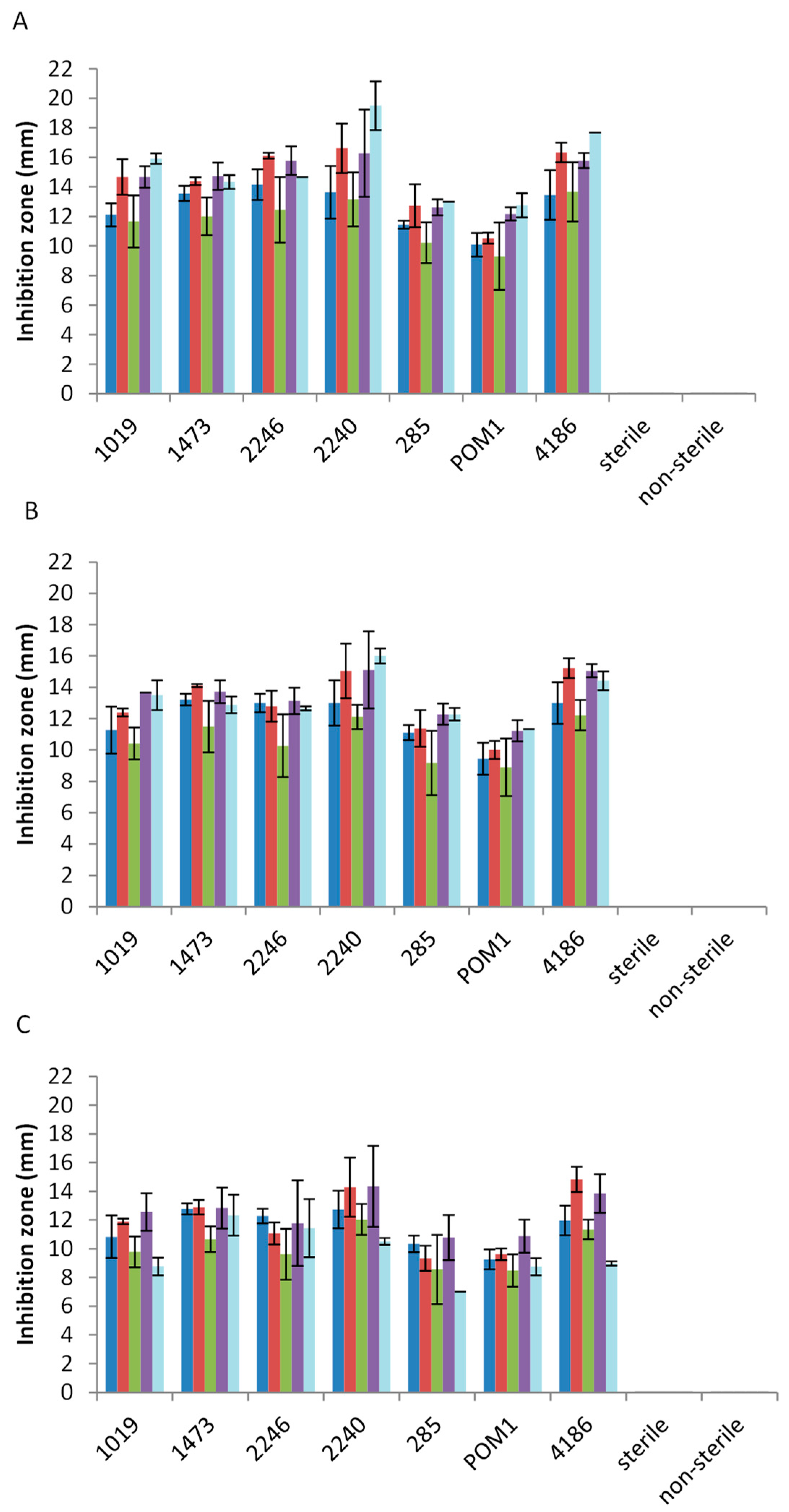
2.2. Antimicrobial Activity toward Foodborne Pathogens
3. Discussion
4. Materials and Methods
4.1. Tomato, Carrot, and Melon By-Products
4.2. Lactic Acid Bacteria Strains
4.3. Setup of Fermentation Conditions
4.4. Extract Production
4.5. Pathogenic Bacterial Strains
4.6. Evaluation of In Vitro Antimicrobial Activity
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Control 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Fernández-López, J.; Zhi, N.; Aleson-Carbonell, L.; Pérez-Alvarez, J.A.; Kuri, V. Antioxidant and antibacterial activities of natural extracts: Application in beef meatballs. Meat Sci. 2005, 69, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Zhong, Y. Novel antioxidants in food quality preservation and health promotion. Eur. J. Lipid Sci. Tech. 2010, 112, 930–940. [Google Scholar] [CrossRef]
- Tiwari, B.K.; Valdramidis, V.P.; O’Donnell, C.P.; Muthukumarappan, K.; Bourke, P.; Cullen, P.J. Application of natural antimicrobials for food preservation. J. Agric. Food Chem. 2009, 57, 5987–6000. [Google Scholar] [CrossRef]
- Elsser-Gravesen, D.; Elsser-Gravesen, A. Biopreservatives. In Biotechnology of Food and Feed Additives, 1st ed.; Zorn, H., Czermak, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 29–49. [Google Scholar]
- Irkin, R.; Esmer, O.K. Novel food packaging systems with natural antimicrobial agents. J. Food Sci. Tech. 2015, 52, 6095–6111. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, P.K.; Garg, G. Natural Products As Preservatives. Int. J. Pharma Bio Sci. 2010, 1, 601–612. [Google Scholar]
- Lucera, A.; Costa, C.; Conte, A.; Del Nobile, M.A. Food applications of natural antimicrobial compounds. Front. Microbiol. 2012, 3, 287. [Google Scholar] [CrossRef]
- Moayedi, A.; Hashemi, M.; Safari, M. Valorization of tomato waste proteins through production of antioxidant and antibacterial hydrolysates by proteolytic Bacillus subtilis: Optimization of fermentation conditions. J. Food Sci. Tech. 2016, 53, 391–400. [Google Scholar] [CrossRef]
- Viuda-Martos, M.; Ruiz-Navajas, Y.; Fernández-López, J.; Pérez-Álvarez, J.A. Effect of adding citrus waste water, thyme and oregano essential oil on the chemical, physical and sensory characteristics of a bologna sausage. Innov. Food Sci. Emerg. Technol. 2009, 10, 655–660. [Google Scholar] [CrossRef]
- Kaur, D.; Wani, A.A.; Oberoi, D.P.S.; Sogi, D.S. Effect of extraction conditions on lycopene extractions from tomato processing waste skin using response surface methodology. Food Chem. 2008, 108, 711–718. [Google Scholar] [CrossRef]
- García, M.L.; Calvo, M.M.; Selgasa, M.D. Beef hamburgers enriched in lycopene using dry tomato peel as an ingredient. Meat Sci. 2009, 83, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, S.; Renda, G.; La Barbera, L.; Amri, M.; Messina, C.M.; Santulli, A. Tunisian tomato by-products, as a potential source of natural bioactive compounds. Nat. Prod. Res. 2016, 31, 626–631. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#home (accessed on 15 September 2019).
- Schirmer, B.C.; Langsrud, S. Evaluation of Natural Antimicrobials on Typical Meat Spoilage Bacteria In Vitro and in Vacuum-Packed Pork Meat. J. Food Sci. 2010, 75, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Cho, A.R.; Han, J. Antioxidant and antimicrobial activities of leafy green vegetable extracts and their applications to meat product preservation. Food Control 2013, 29, 112–120. [Google Scholar] [CrossRef]
- Kabuki, T.; Nakajima, H.; Arai, M.; Ueda, S.; Kuwabara, Y.; Dosako, Y. Characterization of novel antimicrobial compounds from mango (Mangifera indica L.) kernel seeds. Food Chem. 2000, 71, 61–66. [Google Scholar] [CrossRef]
- Gedikoğlu, A.; Sökmen, M.; Çivit, A. Evaluation of Thymus vulgaris and Thymbra spicata essential oils and plant extracts for chemical composition, antioxidant, and antimicrobial properties. Food Sci. Nutr. 2019, 7, 1704–1714. [Google Scholar] [CrossRef]
- Battikh, H.; Bakhrouf, A.; Ammar, E. Antimicrobial effect of Kombucha analogues. LWT-Food Sci. Technol. 2012, 47, 71–77. [Google Scholar] [CrossRef]
- Wen, Y.L.; Yan, L.P.; Chen, C.S. Effects of fermentation treatment on antioxidant and antimicrobial activities of four common Chinese herbal medicinal residues by Aspergillus oryzae. J. Food Drug Anal. 2013, 21, 219–226. [Google Scholar] [CrossRef]
- Adams, M.R.; Nicolaides, L. Review of the sensitivity of different foodborne pathogens to fermentation. Food Control 2008, 8, 227–239. [Google Scholar] [CrossRef]
- Poutanen, K.; Flander, L.; Katina, K. Sourdough and cereal fermentation in a nutritional perspective. Food Microbiol. 2009, 26, 693–699. [Google Scholar] [CrossRef]
- Van Boekel, M.; Fogliano, V.; Pellegrini, N.; Stanton, C.; Scholz, G.; Lalljie, S.; Somoza, V.; Knorr, D.; Jasti, P.R.; Eisenbrand, G. A review on the beneficial aspects of food processing. Mol. Nutr. Food Res. 2010, 54, 1215–1247. [Google Scholar] [CrossRef] [PubMed]
- Gaggia, F.; Di Gioia, D.; Baffoni, L.; Biavati, B. The role of protective and probiotic cultures in food and feed and their impact in food safety. Trends Food Sci. Technol. 2011, 22, S58–S66. [Google Scholar] [CrossRef]
- Sicard, D.; Legras, J.L. Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. C. R. Biol. 2011, 334, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Jacques, N.; Casaregola, S. Safety assessment of dairy microorganisms: The hemiascomycetous yeasts. Int. J. Food Microbiol. 2008, 126, 321–326. [Google Scholar] [CrossRef]
- Ricci, A.; Cirlini, M.; Calani, L.; Bernini, V.; Neviani, E.; Del Rio, D.; Galaverna, G.; Lazzi, C. In vitro metabolism of elderberry juice polyphenols by lactic acid bacteria. Food Chem. 2019, 276, 692–699. [Google Scholar] [CrossRef]
- Valerio, F.; Lavermicocca, P.; Pascale, M.; Visconti, A. Production of phenyllactic acid by lactic acid bacteria: An approach to the selection of strains contributing to food quality and preservation. FEMS Microbiol. Lett. 2004, 233, 289–295. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Valerio, F.; Visconti, A. Antifungal Activity of Phenyllactic Acid against Molds Isolated from Bakery Products. Appl. Environ. Microbiol. 2003, 69, 634–640. [Google Scholar] [CrossRef]
- Ricci, A.; Cirlini, M.; Maoloni, A.; Del Rio, D.; Calani, L.; Bernini, V.; Galaverna, G.; Neviani, E.; Lazzi, C. Use of Dairy and Plant-Derived Lactobacilli as Starters for Cherry Juice Fermentation. Nutrients 2019, 11, 213. [Google Scholar] [CrossRef]
- Gardner, N.J.; Savard, T.; Obermeier, P.; Caldwell, G.; Champagne, C.P. Selection and characterization of mixed starter cultures for lactic acid fermentation of carrot, cabbage, beet and onion vegetable mixtures. Int. J. Food Microbiol. 2001, 64, 261–275. [Google Scholar] [CrossRef]
- Aukrust, T.W.; Blom, H.; Sandtory, B.F.; Slinde, E. Interactions between starter culture and raw material in lactic acid fermentation of sliced carrot. LWT-Food Sci. Technol. 1994, 27, 337–341. [Google Scholar] [CrossRef]
- Di Cagno, R.; Surico, R.F.; Siragusa, S.; De Angelis, M.; Paradiso, A.; Minervini, F.; De Gara, L.; Gobbetti, M. Selection and use of autochthonous mixed starter for lactic acid fermentation of carrots, French beans or marrows. Int. J. Food Microbiol. 2008, 127, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Peña, N.; de Chiara, M.L.V.; Amodio, M.L.; Colelli, G.; Spano, G. Probiotic lactic acid bacteria for the production of multifunctional fresh-cut cantaloupe. Food Res. Int. 2015, 77, 762–772. [Google Scholar] [CrossRef]
- Di Cagno, R.; Surico, R.F.; Paradiso, A.; De Angelis, M.; Salmon, J.C.; Buchin, S.; De Gara, L.; Gobbetti, M. Effect of autochthonous lactic acid bacteria starters on health-promoting and sensory properties of tomato juices. Int. J. Food Microbiol. 2009, 128, 473–483. [Google Scholar] [CrossRef]
- Filannino, P.; Azzi, L.; Cavoski, I.; Vincentini, O.; Rizzello, C.G.; Gobbetti, M.; Di Cagno, R. Exploitation of the health-promoting and sensory properties of organic pomegranate (Punica granatum L.) juice through lactic acid fermentation. Int. J. Food Microbiol. 2013, 163, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Di Cagno, R.; Trani, A.; Cantatore, V.; Gambacorta, G.; Gobbetti, M. Lactic acid fermentation enriches the profile of biogenic compounds and enhances the functional features of common purslane (Portulaca oleracea L.). J. Funct. Foods 2017, 39, 175–185. [Google Scholar] [CrossRef]
- Sirilun, S.; Sivamaruthi, B.S.; Kumar, N.; Kesika, P.; Peerajan, S.; Chaiyasut, C. Lactobacillus-fermented plant juice as a potential ingredient in cosmetics: Formulation and assessment of natural mouthwash. Asian J. Pharm Clin. Res. 2016, 9, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Hayouni, E.A.; Abedrabba, M.; Bouix, M.; Hamdi, M. The effects of solvents and extraction method on the phenolic contents and biological activities in vitro of Tunisian Quercus coccifera L. and Juniperus phoenicea L. fruit extracts. Food Chem. 2007, 105, 1126–1134. [Google Scholar] [CrossRef]
- Al-Zoreky, N.S. Antimicrobial activity of pomegranate (Punica granatum L.) fruit peels. Int. J. Food Microbiol. 2009, 134, 244–248. [Google Scholar] [CrossRef]
- Dall’Asta, C.; Cirlini, M.; Morini, E.; Rinaldi, M.; Ganino, T.; Chiavaro, E. Effect of chestnut flour supplementation on physico-chemical properties and volatiles in bread making. LWT-Food Sci. Technol. 2013, 53, 233–239. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity. J. Pharm. Biomed. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Tomato | Melon | Carrot | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Strain | T0 | T72 | Δ (T72 − T0) | T0 | T72 | Δ (T72 − T0) | T0 | T72 | Δ (T72 − T0) |
| L. plantarum POM1 | 7.04 ± 0.10 | 8.84 ± 0.24 | 1.80 | 6.93 ± 0.35 | 8.88 ± 0.13 | 1.94 | 7.26 ± 0.10 | 9.30 ± 0.08 | 2.04 |
| L. plantarum 285 | 7.73 ± 0.09 | 9.35 ± 0.08 | 1.62 | 7.62 ± 0.02 | 9.16 ± 0.08 | 1.54 | 7.64 ± 0.05 | 9.39 ± 0.05 | 1.75 |
| L. paracasei 4186 | 7.82 ± 0.15 | 8.66 ± 0.45 | 0.84 | 7.86 ± 0.26 | 9.08 ± 0.21 | 1.22 | 7.67 ± 0.07 | 9.42 ± 0.07 | 1.75 |
| L. casei 2246 | 7.21 ± 0.29 | 8.78 ± 0.38 | 1.57 | 7.59 ± 0.21 | 9.48 ± 0.31 | 1.89 | 7.60 ± 0.04 | 9.64 ± 0.06 | 2.03 |
| L. casei 2240 | 7.68 ± 0.10 | 9.07 ± 0.09 | 1.39 | 7.75 ± 0.07 | 9.37 ± 0.03 | 1.61 | 7.71 ± 0.03 | 9.68 ± 0.09 | 1.97 |
| L. rhamnosus 1473 | 7.80 ± 0.09 | 9.37 ± 0.20 | 1.57 | 7.03 ± 0.11 | 9.57 ± 0.08 | 2.55 | 7.43 ± 0.12 | 9.43 ± 0.11 | 2.00 |
| L. rhamnosus 1019 | 7.39 ± 0.29 | 8.40 ± 0.25 | 1.01 | 7.36 ± 0.23 | 9.87 ± 0.56 | 2.51 | 7.64 ± 0.05 | 9.50 ± 0.04 | 1.86 |
| Extract | Tomato | Melon | Carrot |
|---|---|---|---|
| L. plantarum POM1 | 3.08 ± 0.04 | 3.70 ± 0.02 | 3.66 ± 0.02 |
| L. plantarum 285 | 3.70 ± 0.08 | 3.61 ± 0.02 | 3.60 ± 0.00 |
| L. paracasei 4186 | 3.19 ± 0.80 | 3.57 ± 0.02 | 3.63 ± 0.02 |
| L. rhamnosus 1019 | 3.66 ± 0.03 | 3.64 ± 0.01 | 3.47 ± 0.07 |
| L. rhamnosus 1473 | 3.50 ± 0.14 | 3.62 ± 0.05 | 3.52 ± 0.05 |
| L. casei 2240 | 2.89 ± 1.19 | 3.52 ± 0.04 | 3.46 ± 0.01 |
| L. casei 2246 | 3.51 ± 0.47 | 3.54 ± 0.06 | 3.48 ± 0.02 |
| Sterile substrate | 4.06 ± 0.06 | 4.57 ± 0.10 | 4.33 ± 0.16 |
| Non-sterile substrate | 4.28 ± 0.04 | 4.53 ± 0.13 | 4.44 ± 0.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ricci, A.; Bernini, V.; Maoloni, A.; Cirlini, M.; Galaverna, G.; Neviani, E.; Lazzi, C. Vegetable By-Product Lacto-Fermentation as a New Source of Antimicrobial Compounds. Microorganisms 2019, 7, 607. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120607
Ricci A, Bernini V, Maoloni A, Cirlini M, Galaverna G, Neviani E, Lazzi C. Vegetable By-Product Lacto-Fermentation as a New Source of Antimicrobial Compounds. Microorganisms. 2019; 7(12):607. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120607
Chicago/Turabian StyleRicci, Annalisa, Valentina Bernini, Antonietta Maoloni, Martina Cirlini, Gianni Galaverna, Erasmo Neviani, and Camilla Lazzi. 2019. "Vegetable By-Product Lacto-Fermentation as a New Source of Antimicrobial Compounds" Microorganisms 7, no. 12: 607. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120607