Curative Treatment of Candidiasis by the Live Biotherapeutic Microorganism Lactobacillus rhamnosus Lcr35® in the Invertebrate Model Caenorhabditis elegans: First Mechanistic Insights

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Anti C. albicans Effects of L. rhamnosus Lcr35® on Caco-2 Cell Monolayer
2.1.1. Growth Inhibition of C. albicans
2.1.2. Inhibition of C. albicans Biofilm Formation
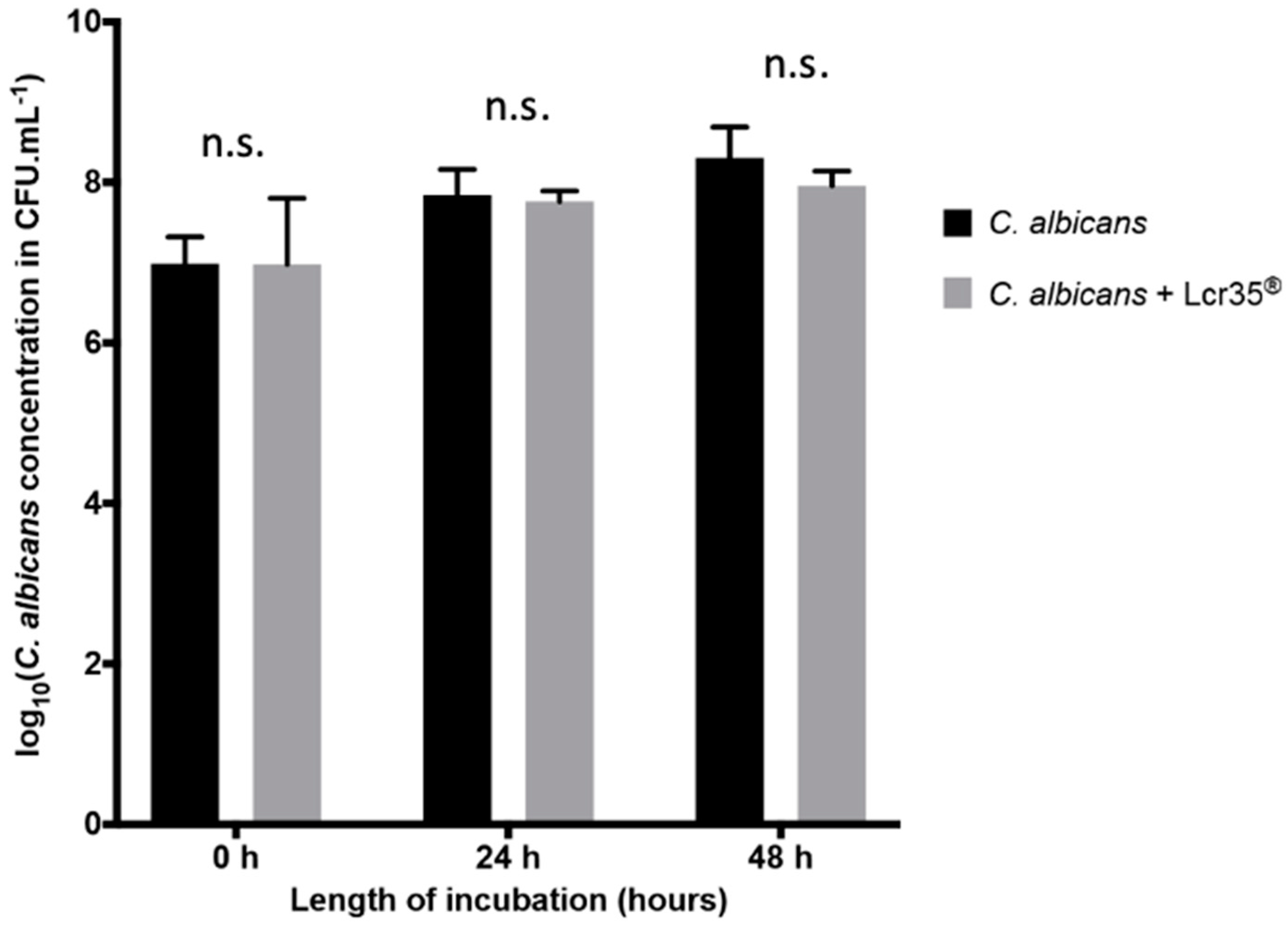
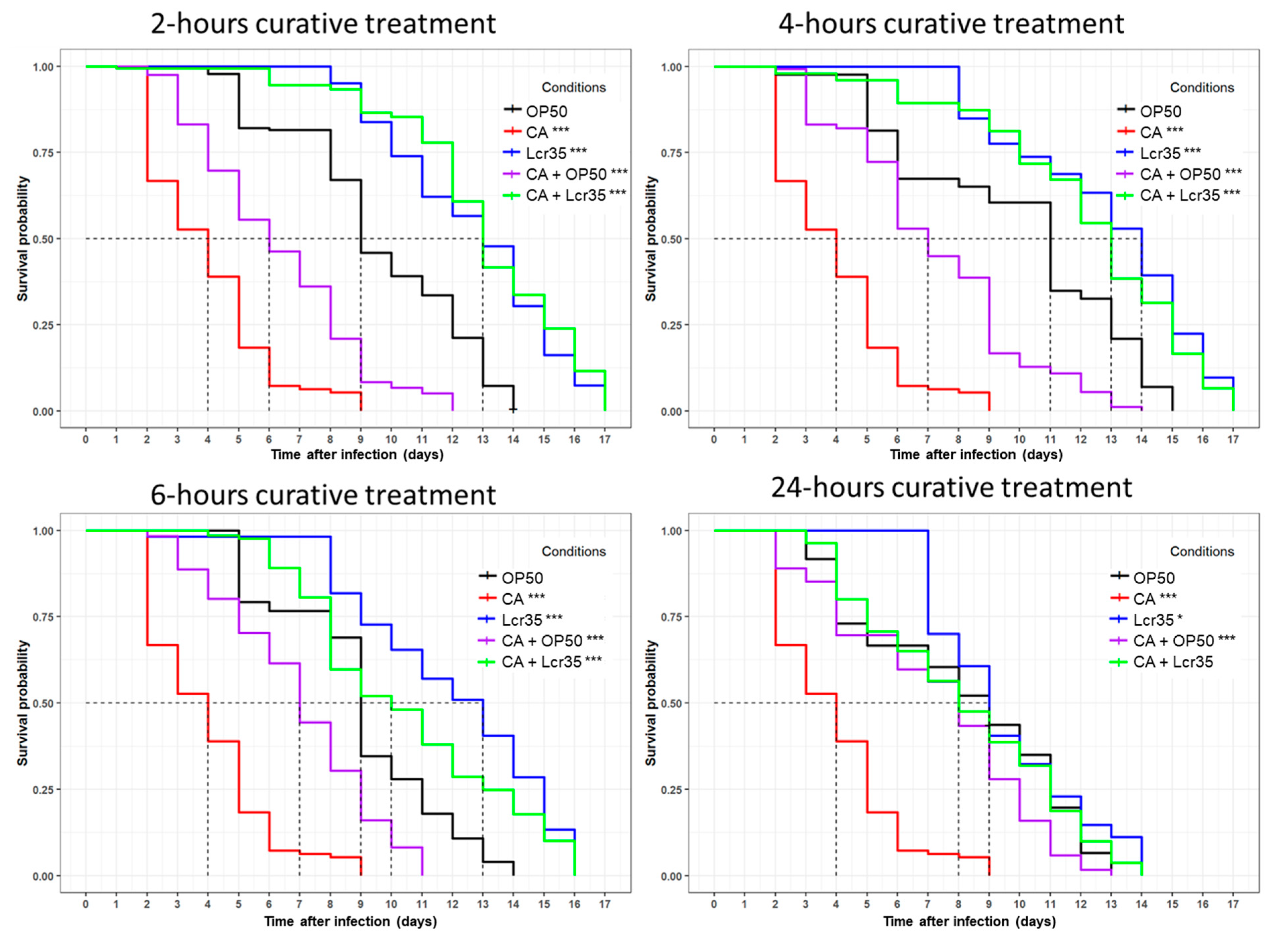
2.2. Effect of L. rhamnosus Lcr35® Curative Treatment on Candidiasis
2.2.1. Effect of L. rhamnosus Lcr35® on C. elegans Survival after C. albicans Exposure
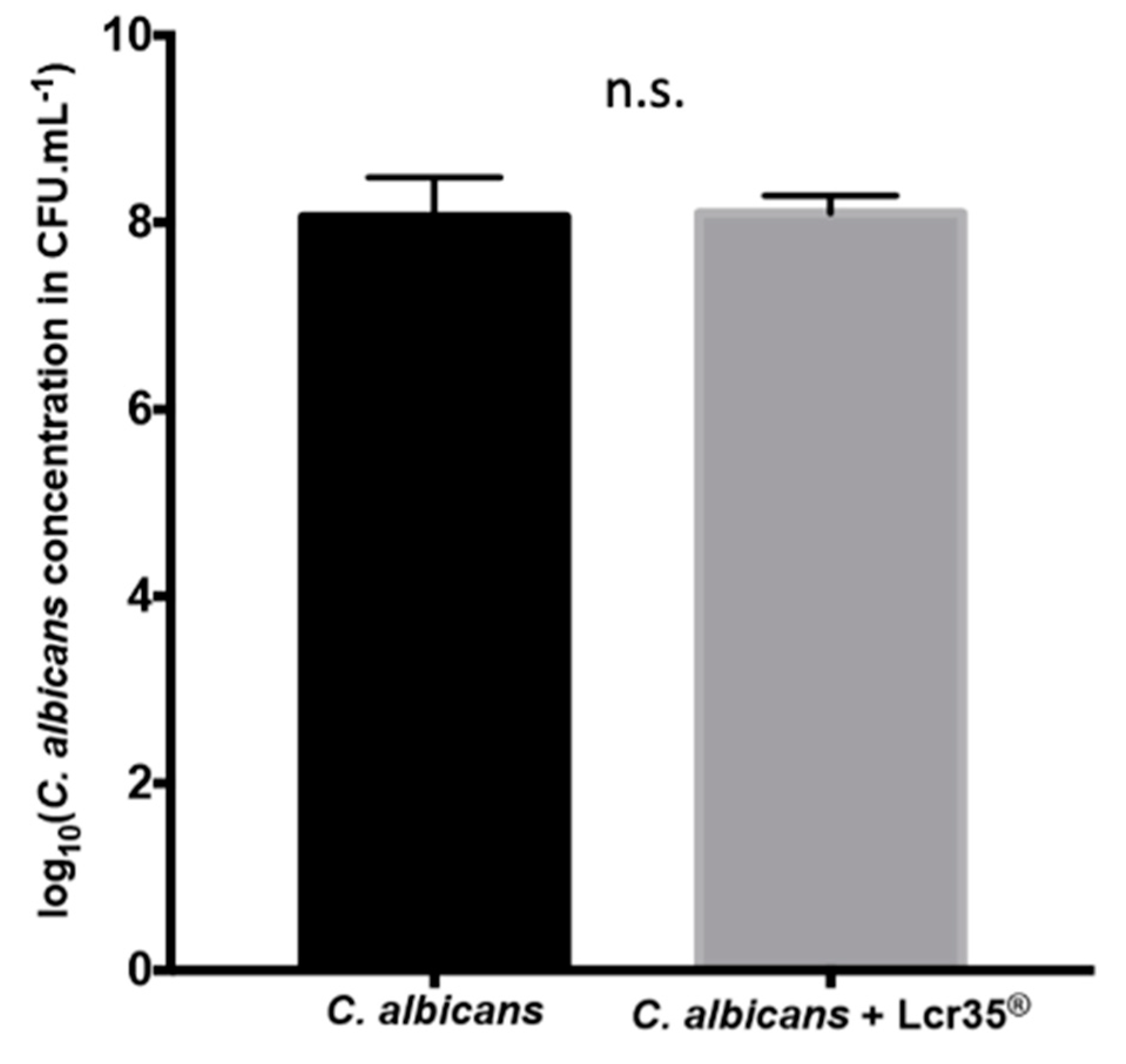
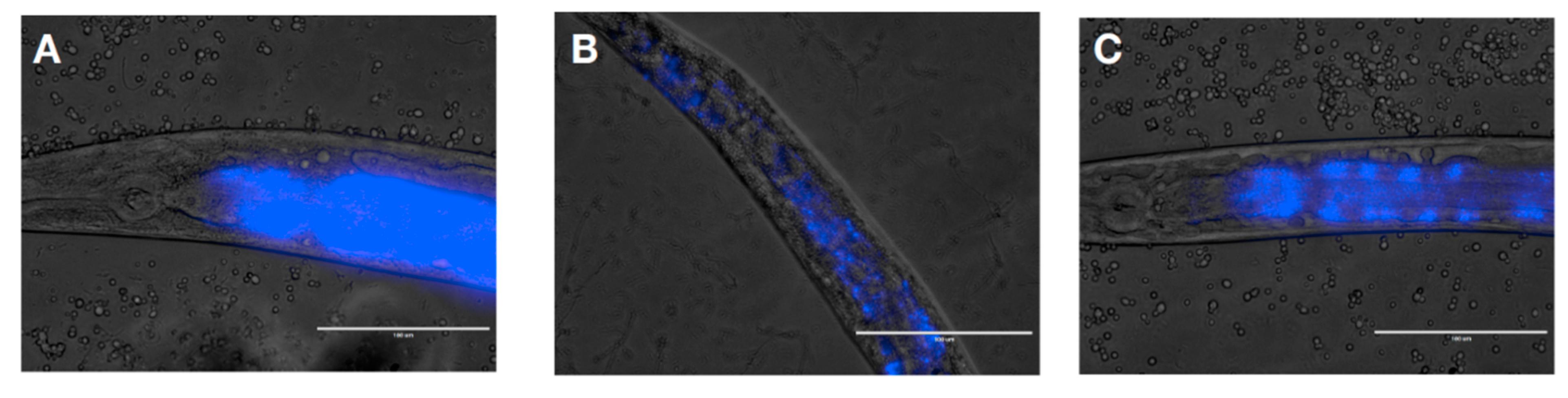
2.2.2. Influence of L. rhamnosus Lcr35® on C. albicans Presence in the Worm Gut
2.3. Effect of L. rhamnosus Lcr35® Cell-Free Supernatant on Candidiasis
2.4. Mechanistic Study
2.4.1. Modulation of C. elegans Genes Expression Induced by Lcr35® and C. albicans
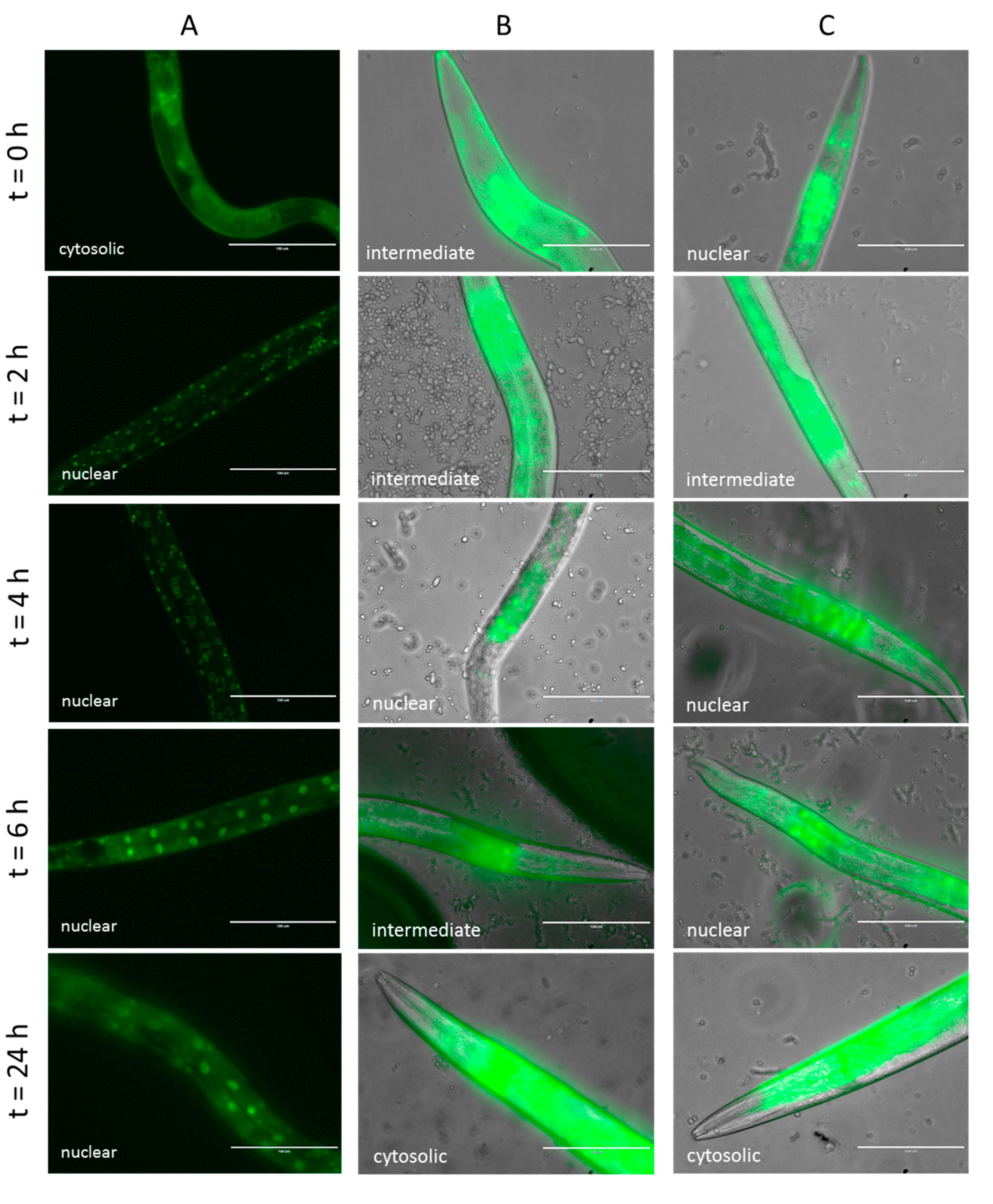
2.4.2. Effect of Lcr35® Curative Treatment on DAF-16 Nuclear Translocation
3. Discussion
4. Materials and Methods
4.1. Microbial Strains and Growth Conditions
4.2. Influence of L. rhamnosus Lcr35® on Growth of C. albicans on Caco-2 Cells Monolayer and on Biofilm Formation
4.3. C. elegans Maintenance
4.4. C. elegans Synchronization
4.5. Effects of L. rhamnosus Lcr35® on Candidiasis in C. elegans
4.5.1. Preparation of Plates Containing LBM or Pathogen Yeasts
4.5.2. Survival Assay: Curative Treatment
4.6. Study of L. rhamnosus Lcr35® Cell-Free Supernatant on Candidiasis in C. elegans
4.7. Visualization of C. albicans in C. elegans Intestine
4.8. RNA Isolation and Real-Time Quantitative PCR
4.9. DAF-16 Nuclear Localization
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pfaller, M.A.; Diekema, D.J. Epidemiology of Invasive Candidiasis: A Persistent Public Health Problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [Green Version]
- Ruhnke, M. Clinical Syndromes: Candida and Candidosis. In Clinically Relevant Mycoses; Presterl, E., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.; Benjamin, D.K., Jr.; Calandra, T.F.; Edwards, J.E., Jr.; Filler, S.G.; Fisher, J.F.; Kullberg, B.J.; Ostrosky-Zeichner, L.; et al. Clinical Practice Guidelines for the Management of Candidiasis: 2009 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 503–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, J.; Sobel, J.D.; Nyirjesy, P.; Sobel, R.; Williams, V.L.; Yu, Q.; Noverr, M.C.; Fidel, P.L. Current patient perspectives of vulvovaginal candidiasis: Incidence, symptoms, management and post-treatment outcomes. BMC Womens Health 2019, 19, 48. [Google Scholar] [CrossRef] [Green Version]
- Kumamoto, C. Inflammation and gastrointestinal Candida colonization Carol. Curr. Opin. Microbiol. 2011, 14, 386–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spettel, K.; Barousch, W.; Makristathis, A.; Zeller, I.; Nehr, M.; Selitsch, B.; Lackner, M.; Rath, P.M.; Steinmann, J.; Willinger, B. Analysis of antifungal resistance genes in Candida albicans and Candida glabrata using next generation sequencing. PLoS ONE 2019, 14, e0210397. [Google Scholar] [CrossRef] [PubMed]
- Moran, C.; Grussemeyer, C.A.; Spalding, J.R.; Benjamin, D.K.; Reed, S.D.; Reed, D.S.D. Comparison of costs, length of stay, and mortality associated with Candida glabrata and Candida albicans bloodstream infections. Am. J. Infect. Control 2010, 38, 78–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mac Dougall, C.; Polk, R.E. Antimicrobial Stewardship Programs in Health Care Systems. Clin. Microbiol. Rev. 2005, 18, 638–656. [Google Scholar] [CrossRef] [Green Version]
- Dreher-Lesnick, S.M.; Stibitz, S.; Carlson, P.E., Jr. US Regulatory Considerations for Development of Live Biotherapeutic Products as Drugs. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- O’Toole, P.W.; Marchesi, J.R.; Hill, C. Next-generation probiotics: The spectrum from probiotics to live biotherapeutics. Nat. Microbiol. 2017, 2, 17057. [Google Scholar] [CrossRef]
- Nivoliez, A.; Camares, O.; Paquet-Gachinat, M.; Bornes, S.; Forestier, C.; Veisseire, P. Influence of manufacturing processes on in vitro properties of the probiotic strain Lactobacillus rhamnosus Lcr35®. J. Biotechnol. 2012, 160, 236–241. [Google Scholar] [CrossRef]
- Poupet, C.; Saraoui, T.; Veisseire, P.; Bonnet, M.; Dausset, C.; Gachinat, M.; Camarès, O.; Chassard, C.; Nivoliez, A.; Bornes, S. Lactobacillus rhamnosus Lcr35 as an effective treatment for preventing Candida albicans infection in the invertebrate model Caenorhabditis elegans: First mechanistic insights. PLoS ONE 2019, 14, e0216184. [Google Scholar] [CrossRef] [PubMed]
- Patrignani, F.; Siroli, L.; Parolin, C.; Serrazanetti, D.I.; Vitali, B.; Lanciotti, R. Use of Lactobacillus crispatus to produce a probiotic cheese as potential gender food for preventing gynaecological infections. PLoS ONE 2019, 14, e0208906. [Google Scholar] [CrossRef] [PubMed]
- Romeo, M.G.; Romeo, D.M.; Trovato, L.; Oliveri, S.; Palermo, F.; Cota, F.; Betta, P. Role of probiotics in the prevention of the enteric colonization by Candida in preterm newborns: Incidence of late-onset sepsis and neurological outcome. J. Perinatol. 2011, 31, 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Bansal, A.; Chakrabarti, A.; Singhi, S. Evaluation of Efficacy of Probiotics in Prevention of Candida Colonization in a PICU—A Randomized Controlled Trial*. Crit. Care. Med. 2013, 41, 565–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Barros, P.P.; Scorzoni, L.; Ribeiro, F.C.; Fugisaki, L.R.O.; Fuchs, B.B.; Mylonakis, E.; Jorge, A.O.C.; Junqueira, J.C.; Rossoni, R.D. Lactobacillus paracasei 28.4 reduces in vitro hyphae formation of Candida albicans and prevents the filamentation in an experimental model of Caenorhabditis elegans. Microb. Pathog. 2018, 117, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.; Pooranachithra, M.; Balamurugan, K.; Goel, G. Probiotic mediated colonization resistance against E. coli infection in experimentally challenged Caenorhabditis elegans. Microb. Pathog. 2019, 127, 39–47. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, X.; Yu, H.; Yin, X.; Nie, S.-P.; Xie, M.-Y.; Chen, W.; Gong, J. Cell Signaling of Caenorhabditis elegans in Response to Enterotoxigenic Escherichia coli Infection and Lactobacillus zeae Protection. Front. Immunol. 2018, 9, 1745. [Google Scholar] [CrossRef] [Green Version]
- Schifano, E.; Zinno, P.; Guantario, B.; Roselli, M.; Marcoccia, S.; Devirgiliis, C.; Uccelletti, D. The Foodborne Strain Lactobacillus fermentum MBC2 Triggers pept-1-Dependent Pro-Longevity Effects in Caenorhabditis elegans. Microorganisms 2019, 7, 45. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhao, Y.; Liu, R.; Zheng, X.; Zhang, M.; Guo, H.; Zhang, H.; Ren, F. The transcription factor DAF-16 is essential for increased longevity in C. elegans Exposed to Bifidobacterium longum BB68. Sci. Rep. 2017, 7, 7408. [Google Scholar] [CrossRef]
- Pukkila-Worley, R.; Ausubel, F.M.; Mylonakis, E. Candida albicans infection of Caenorhabditis elegans induces antifungal immune defenses. PLoS Pathog. 2011, 7, e1002074. [Google Scholar] [CrossRef] [Green Version]
- Wegh, C.A.M.; Benninga, M.A.; Tabbers, M.M. Effectiveness of Probiotics in Children with Functional Abdominal Pain Disorders and Functional Constipation A Systematic Review. J. Clin. Gastroenterol. 2018, 52, S10–S26. [Google Scholar] [CrossRef] [PubMed]
- Wojtyniak, K.; Szajewska, H. Systematic Review: Probiotics for Functional Constipation in Children. J. Pediatr. 2017, 176, 1155–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojtyniak, K.; Horvath, A.; Dziechciarz, P.; Szajewska, H. Lactobacillus casei rhamnosus Lcr35 in the Management of Functional Constipation in Children: A Randomized Trial. J. Pediatr. 2017, 184, 101–105.e1. [Google Scholar] [CrossRef] [PubMed]
- Dausset, C.; Patrier, S.; Gajer, P.; Thoral, C.; Lenglet, Y.; Cardot, J.M.; Judlin, P.; Ravel, J.; Nivoliez, A. Comparative phase I randomized open-label pilot clinical trial of Gynophilus® (Lcr regenerans®) immediate release capsules versus slow release muco-adhesive tablets. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1869–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, A.; Motyl, I.; Śliżewska, K.; Libudzisz, Z.; Klewicka, E. Adherence of probiotic bacteria to human colon epithelial cells and inhibitory effect against enteric pathogens—In vitro study. Int. J. Dairy Technol. 2016, 69, 532–539. [Google Scholar]
- Allonsius, C.N.; van den Broek, M.F.L.; De Boeck, I.; Kiekens, S.; Oerlemans, E.F.M.; Kiekens, F.; Foubert, K.; Vandenheuvel, D.; Cos, P.; Delputte, P.; et al. Interplay between Lactobacillus rhamnosus GG and Candida and the involvement of exopolysaccharides. Microb. Biotechnol. 2017, 10, 1753–1763. [Google Scholar] [CrossRef]
- Irazoqui, J.E.; Troemel, E.R.; Feinbaum, R.L.; Luhachack, L.G.; Cezairliyan, B.O.; Ausubel, F.M. Distinct pathogenesis and host responses during infection of C. elegans by P. aeruginosa and S. aureus. PLoS Pathog. 2010, 6, e1000982. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Conly, J.; McClure, J.A.; Elsayed, S.; Louie, T.; Zhang, K. Caenorhabditis elegans as a host model for community-associated methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2010, 16, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Souza, A.C.R.; Fuchs, B.B.; Alves V de, S.; Jayamani, E.; Colombo, A.L.; Mylonakis, E. Pathogenesis of the Candida parapsilosis complex in the model host Caenorhabditis elegans. Genes 2018, 9, 401. [Google Scholar] [CrossRef] [Green Version]
- Pukkila-Worley, R.; Peleg, A.Y.; Tampakakis, E.; Mylonakis, E. Candida albicans hyphal formation and virulence assessed using a Caenorhabditis elegans infection model. Eukaryot. Cell 2009, 8, 1750–1758. [Google Scholar] [CrossRef] [Green Version]
- Park, M.R.; Ryu, S.; Maburutse, B.E.; Oh, N.S.; Kim, S.H.; Oh, S.; Jeong, S.Y.; Jeong, D.Y.; Oh, S.; Kim, Y. Probiotic Lactobacillus fermentum strain JDFM216 stimulates the longevity and immune response of Caenorhabditis elegans through a nuclear hormone receptor. Sci. Rep. 2018, 8, 7441. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Mylonakis, E. Caenorhabditis elegans immune conditioning with the probiotic bacterium Lactobacillus acidophilus strain ncfm enhances gram-positive immune responses. Infect. Immun. 2012, 80, 2500–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dausset, C. Détermination des mécanismes d’inhibition de souches de lactobacilles. In Influence du procédé industriel sur leurs caractéristiques probiotiques; AgroParisTech: Jouy-en-Josas, France, 2015. [Google Scholar]
- Graham, C.E.; Cruz, M.R.; Garsin, D.A.; Lorenz, M.C. Enterococcus faecalis bacteriocin EntV inhibits hyphal morphogenesis, biofilm formation, and virulence of Candida albicans. Proc. Natl. Acad. Sci. USA 2017, 114, 4507–4512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Lee, K.; Hsu, M.; Nau, G.; Mylonakis, E.; Ramratnam, B. Lactobacillus-derived extracellular vesicles enhance host immune responses against vancomycin-resistant enterococci. BMC Microbiol. 2017, 17, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komura, T.; Ikeda, T.; Yasui, C.; Saeki, S.; Nishikawa, Y. Mechanism underlying prolongevity induced by bifidobacteria in Caenorhabditis elegans. Biogerontology 2013, 14, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Aballay, A. Regulation of DAF-16-mediated Innate Immunity in Caenorhabditis elegans. J. Biol. Chem. 2009, 284, 35580–35587. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.; Robineleon, S.; Appay, M.D.; Kedinger, M.; Triadou, N.; Dussaulx, E.; Lacroix, B.; Simonassmann, P.; Haffen, K.; Fogh, J.; et al. Enterocyte-like differentiation and polarization of the human colon carcinoma cell line Caco-2 in culture. Biol. Cell 1983, 47, 323–330. [Google Scholar]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome. Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [Green Version]
- Semple, J.I.; Garcia-Verdugo, R.; Lehner, B. Rapid selection of transgenic C. elegans using antibiotic resistance. Nat. Methods. 2010, 7, 725–727. [Google Scholar] [CrossRef]
- Hoogewijs, D.; Houthoofd, K.; Matthijssens, F.; Vandesompele, J.; Vanfleteren, J.R. Selection and validation of a set of reliable reference genes for quantitative sod gene expression analysis in C. elegans. BMC Mol. Biol. 2008, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, H.; Shiozaki, T.; Kobatake, E.; Hosoya, T.; Moriya, T.; Sakai, F.; Taru, H.; Miyazaki, T. Effects and mechanisms of prolongevity induced by Lactobacillus gasseri SBT2055 in Caenorhabditis elegans. Aging Cell 2016, 15, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatima, S.; Haque, R.; Jadiya, P.; Shamsuzzama Kumar, L.; Nazir, A. Ida-1, the Caenorhabditis elegans orthologue of mammalian diabetes autoantigen IA-2, potentially acts as a common modulator between Parkinson’s disease and diabetes: Role of Daf-2/Daf-16 insulin like signalling pathway. PLoS ONE 2014, 9, e113986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.r-project.org/ (accessed on 10 September 2019).
- Therneau, T.M. A Package for Survival Analysis in R. 2015. Available online: https://github.com/therneau/survival (accessed on 10 September 2019).
- Kassambara, A.; Kosinski, M. Survminer: Drawing Survival Curves using “ggplot2.” 2017. Available online: http://www.sthda.com/english/rpkgs/survminer/ (accessed on 10 September 2019).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Lcr35® Supernatant Concentration (%) | Lcr35® Supernatant pH | C. elegans Mean Lifespan (days) | C. elegans Longevity (days) | Log Rank Test (p-Value) |
|---|---|---|---|---|---|
| E. coli OP50 | no supernatant | 9 | 14 | N/A | |
| Lcr35® | no supernatant | 13 | 17 | <2.10–16 | |
| C. albicans | no supernatant | 4 | 9 | <2.10–16 | |
| E. coli OP50 | 30 | 4,5 | 15 | 20 | <2.10–16 |
| 30 | 7 | 13 | 18 | <2.10–16 | |
| 50 | 4,5 | 12 | 15 | 2.10–9 | |
| 50 | 7 | 14 | 21 | <2.10–16 | |
| 100 | 4,5 | 1 | 8 | <2.10–16 | |
| 100 | 7 | 1 | 8 | <2.10–16 | |
| C. albicans | 30 | 4,5 | 5 | 6 | <2.10–16 |
| 30 | 7 | 2 | 3 | <2.10–16 | |
| 50 | 4,5 | 4 | 8 | <2.10–16 | |
| 50 | 7 | 2 | 3 | <2.10–16 | |
| 100 | 4,5 | 1 | 5 | <2.10–16 | |
| 100 | 7 | 2 | 3 | <2.10–16 | |
| Genes of Interest | |||||||
|---|---|---|---|---|---|---|---|
| Insulin Signaling Pathway | p38 MAPK Signaling Pathway | Antimicrobials | |||||
| Conditions | daf-2 | daf-16 | sek-1 | pmk-1 | abf-2 | cnc-4 | fipr-22/fipr-23 |
| E. coli OP50 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Lcr35® | 0.97 | 5.84 *** | 0.50 | 3.05 | 0.90 | 8.48 | 0.89 |
| C. albicans | 1.87 | 0.31 | 4.57 | 4.66 | 3.58 | 14.31 | 3.45 |
| C. albicans + E. coli OP50 | 0.52 | 0.41 | 1.01 | 2.57 | 0.95 | 7.04 | 0.26 |
| C. albicans + Lcr35® | 0.61 | 1.47 | 11.24 ** | 2.42 | 8.00 ** | 1.31 | 4.29 * |
| Gene Name | Gene Type | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Reference |
|---|---|---|---|---|
| cdc-42 | housekeeping | ATCCACAGACCGACGTGTTT | GTCTTTGAGCAATGATGCGA | [42] |
| Y45F10D.4 | housekeeping | CGAGAACCCGCGAAATGTCGGA | CGGTTGCCAGGGAAGATGAGGC | [43] |
| daf-2 | GOI | AAAAGATTTGGCTGGTCAGAGA | TTTCAGTACAAATGAGATTGTCAGC | [44] |
| daf-16 | GOI | TTCAATGCAAGGAGCATTTG | AGCTGGAGAAACACGAGACG | [44] |
| sek-1 | GOI | GCCGATGGAAAGTGGTTTTA | TAAACGGCATCGCCAATAAT | [44] |
| pmk-1 | GOI | CCGACTCCACGAGAAGGATA | AGCGAGTACATTCAGCAGCA | [44] |
| abf-2 | GOI | TCGTCCGTTCCCTTTTCCTT | CCTCTCTTAATAAGAGCACC | [12] |
| fipr-22/fipr-23 | GOI | CCCAATCCAGTATGAAGTTG | ATTTCAGTCTTCACACCGGA | [12] |
| cnc-4 | GOI | ATGCTTCGCTACATTCTCGT | TTACTTTCCAATGAGCATTC | [12] |
| cnc-7 | GOI | TTTTGTTGGCTCTGGTGGCA | ATGAGTCCAGGACGGTACAT | [12] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poupet, C.; Veisseire, P.; Bonnet, M.; Camarès, O.; Gachinat, M.; Dausset, C.; Chassard, C.; Nivoliez, A.; Bornes, S. Curative Treatment of Candidiasis by the Live Biotherapeutic Microorganism Lactobacillus rhamnosus Lcr35® in the Invertebrate Model Caenorhabditis elegans: First Mechanistic Insights. Microorganisms 2020, 8, 34. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010034
Poupet C, Veisseire P, Bonnet M, Camarès O, Gachinat M, Dausset C, Chassard C, Nivoliez A, Bornes S. Curative Treatment of Candidiasis by the Live Biotherapeutic Microorganism Lactobacillus rhamnosus Lcr35® in the Invertebrate Model Caenorhabditis elegans: First Mechanistic Insights. Microorganisms. 2020; 8(1):34. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010034
Chicago/Turabian StylePoupet, Cyril, Philippe Veisseire, Muriel Bonnet, Olivier Camarès, Marylise Gachinat, Caroline Dausset, Christophe Chassard, Adrien Nivoliez, and Stéphanie Bornes. 2020. "Curative Treatment of Candidiasis by the Live Biotherapeutic Microorganism Lactobacillus rhamnosus Lcr35® in the Invertebrate Model Caenorhabditis elegans: First Mechanistic Insights" Microorganisms 8, no. 1: 34. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010034