Enhancing the 1-Aminocyclopropane-1-Carboxylate Metabolic Rate of Pseudomonas sp. UW4 Intensifies Chemotactic Rhizocompetence


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbes and Plants
2.2. Bacterial Promoters
2.3. Construction of Expression Vectors
2.4. Three-Parent Mating and Transconjugant Identification
2.5. Growth Curves
2.6. Quantitative Real-Time Reverse Transcription PCR
2.7. AcdS Enzyme Activity Assay
2.8. Swimming Plate Chemotaxis Assay
2.9. Axenic Tissue Culture Bottles Trial
2.10. Pot Trial
2.11. Statistical Analysis
3. Results
3.1. Identification of the Pseudomonas sp. UW4 ΔAcdS Transconjugants with AcdS Expression Vectors Controlled by Different Promoters
3.2. The Growth Curves of Pseudomonas sp. UW4 Strains
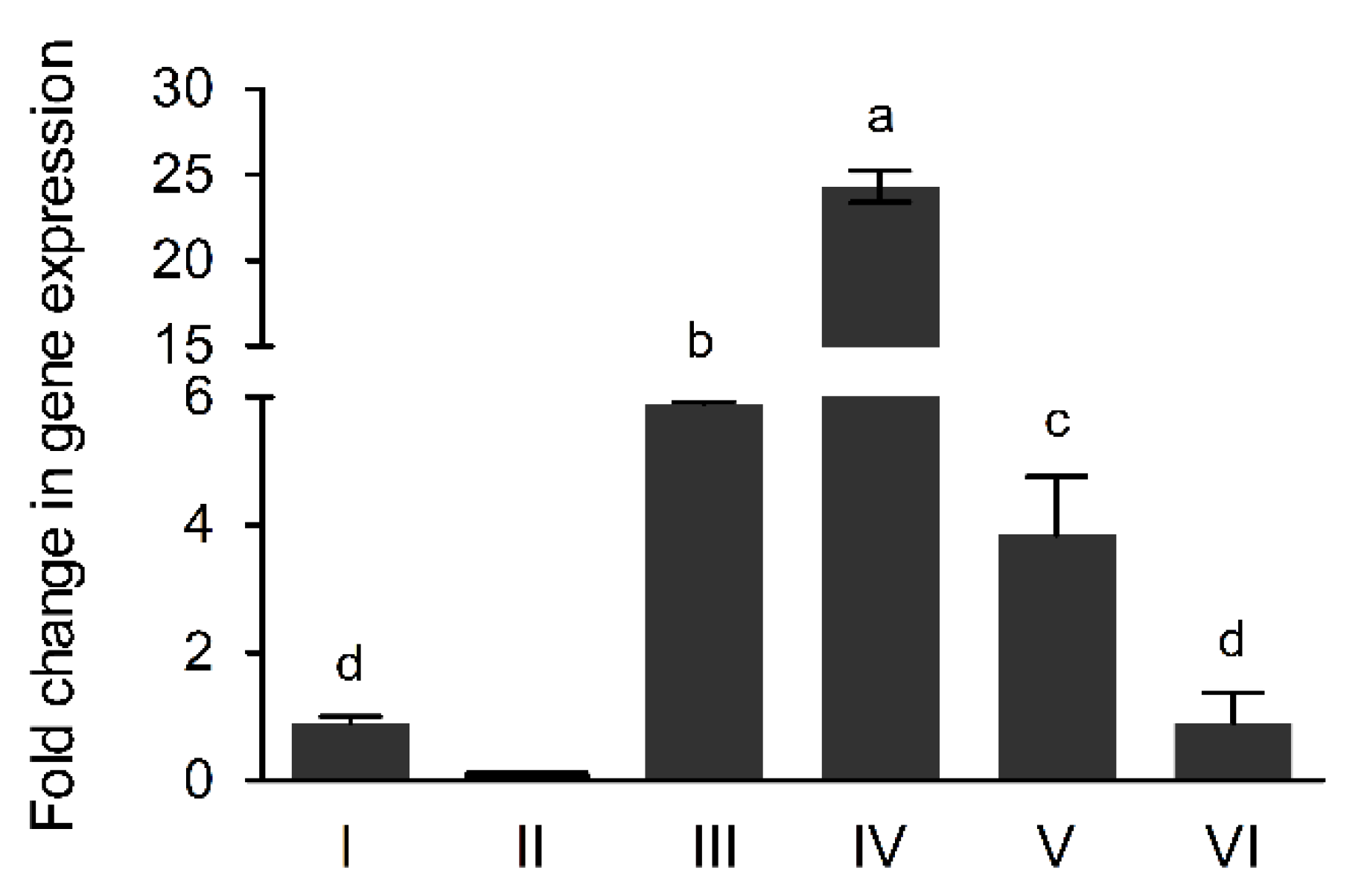
3.3. The AcdS Gene Expression Levels in Pseudomonas sp. UW4 Strains
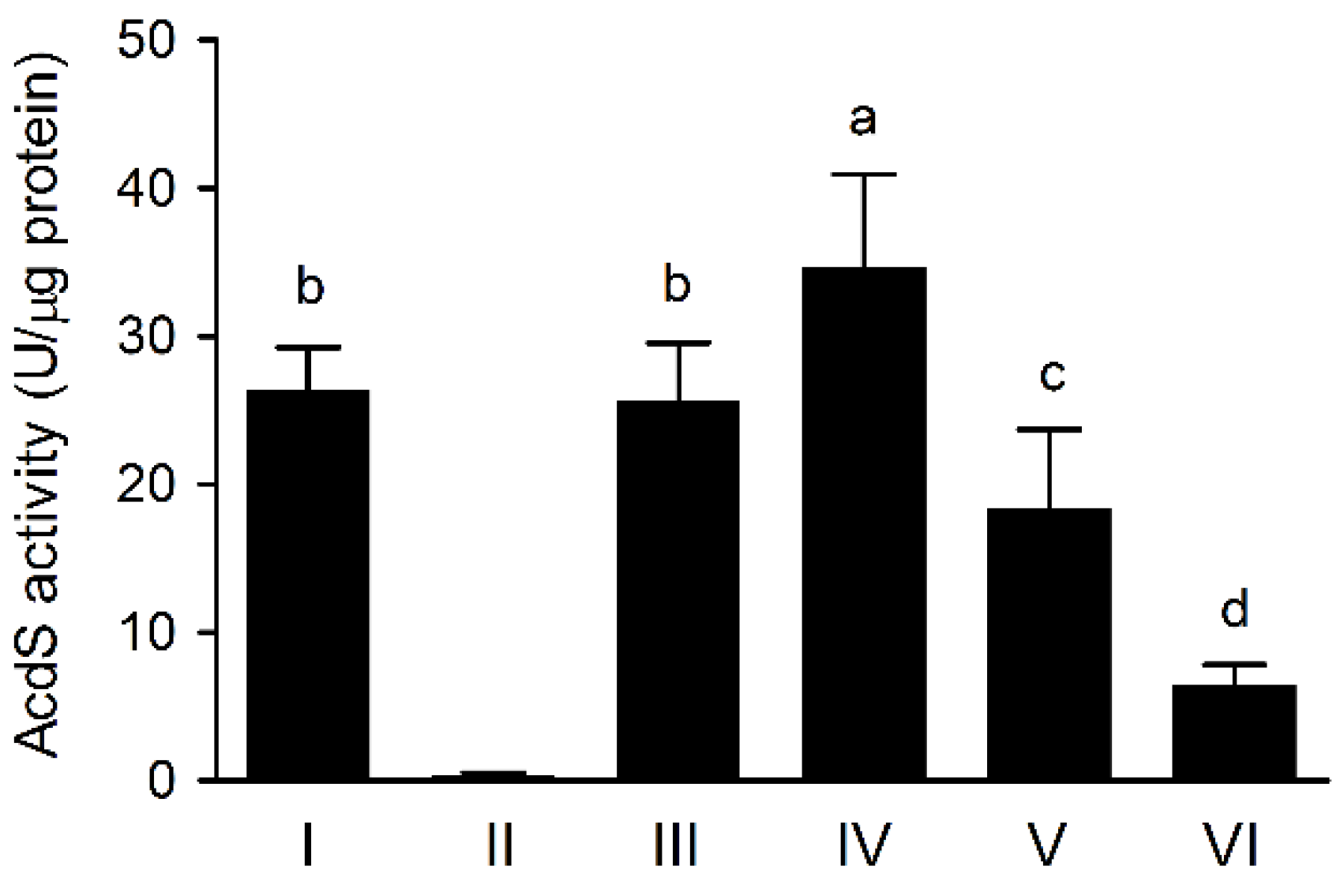
3.4. The AcdS Activities of Pseudomonas sp. UW4 Strains
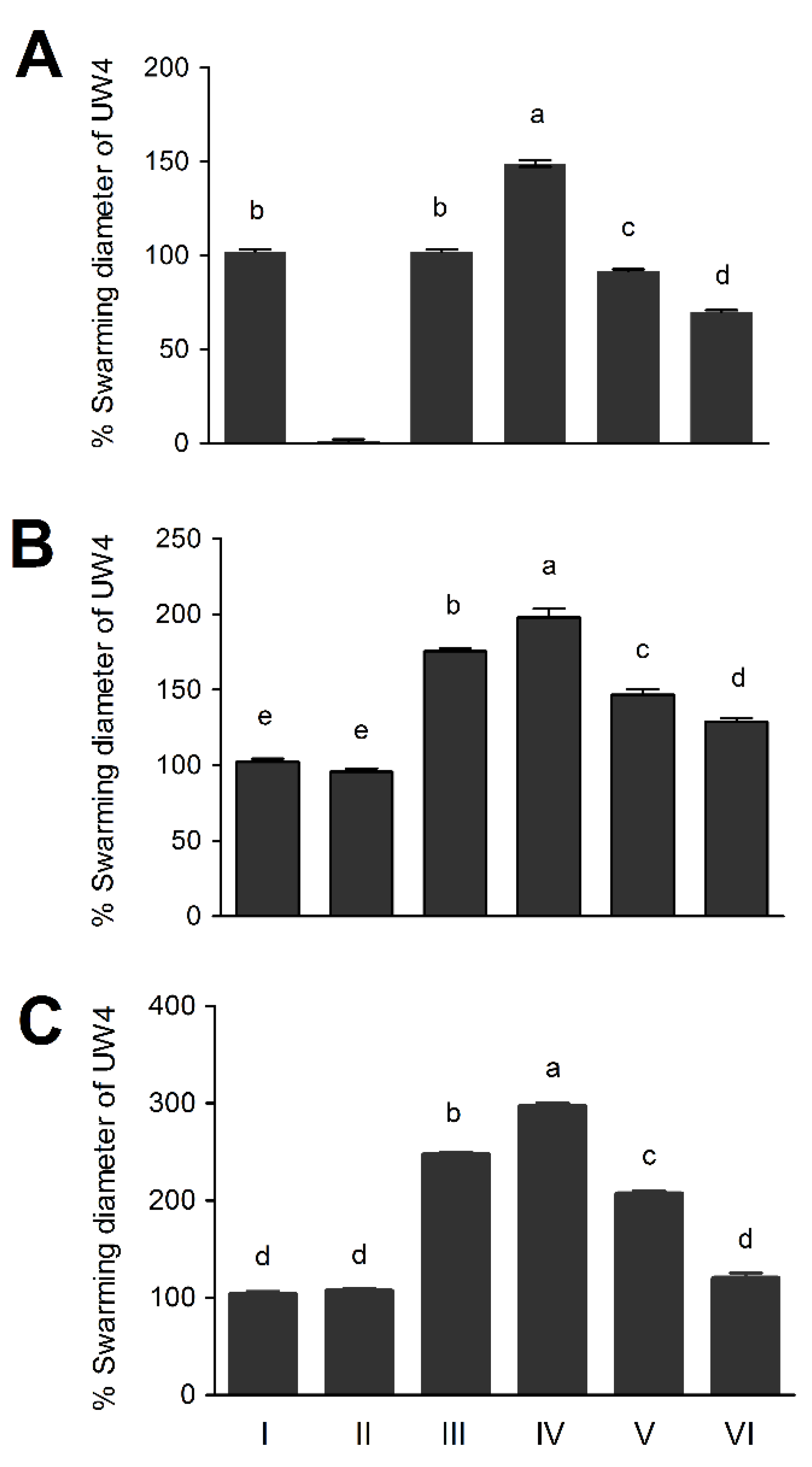
3.5. The Chemotaxis Response of Pseudomonas sp. UW4 Strains to Selected Chemoattractants
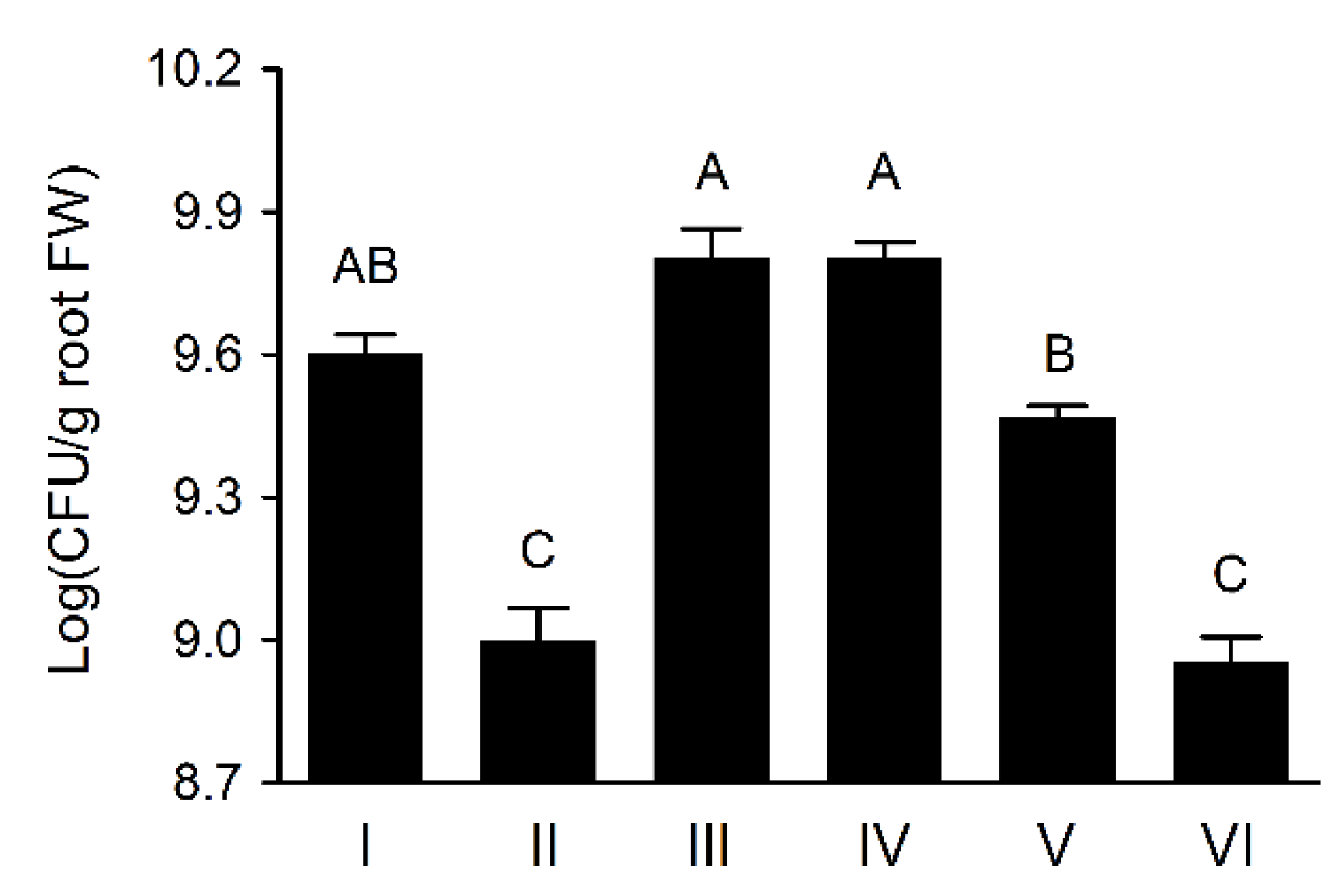
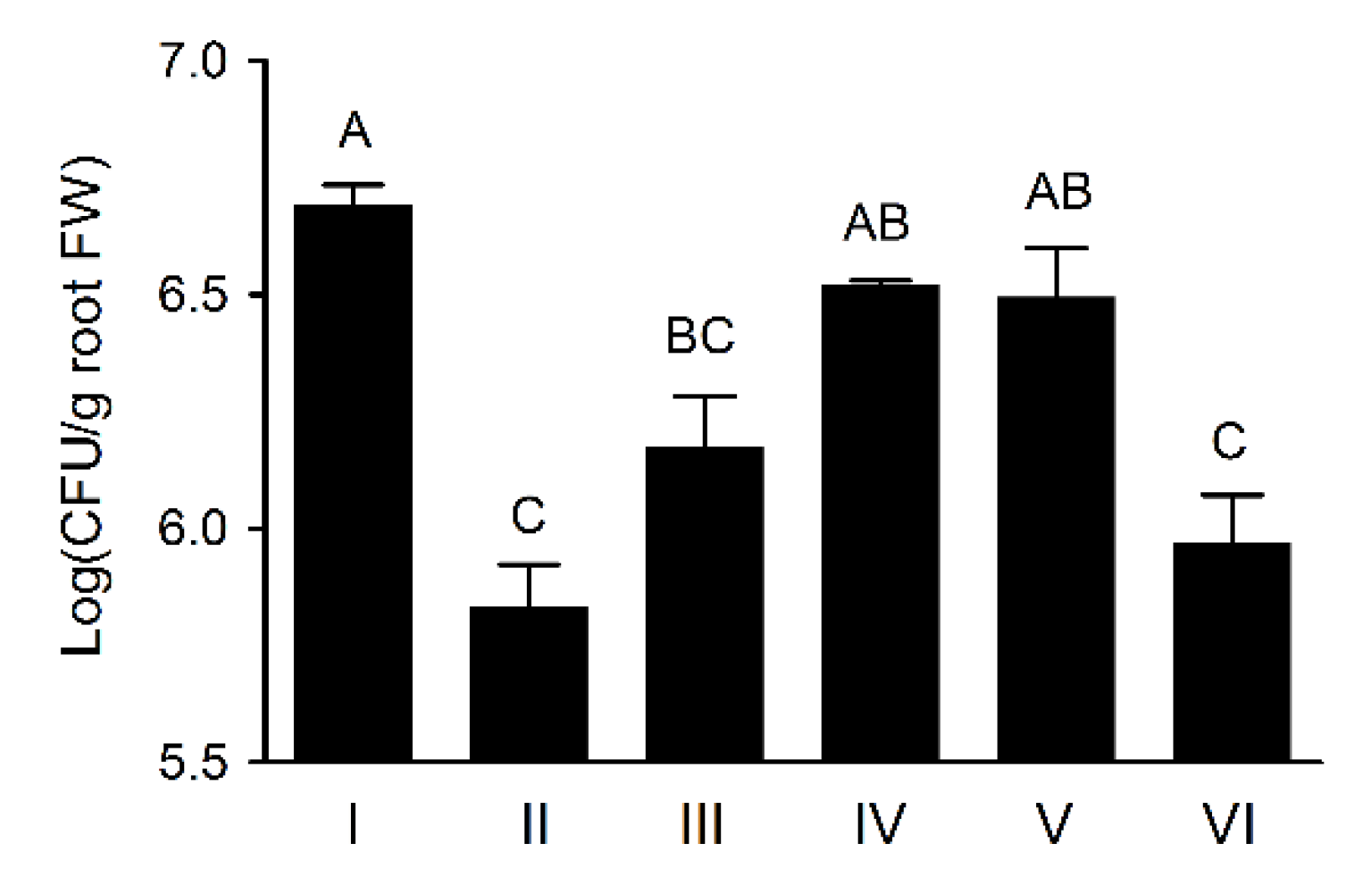
3.6. Wheat Rhizosphere Colonization by Pseudomonas sp. UW4 Strains in Axenic Tissue Culture Bottles
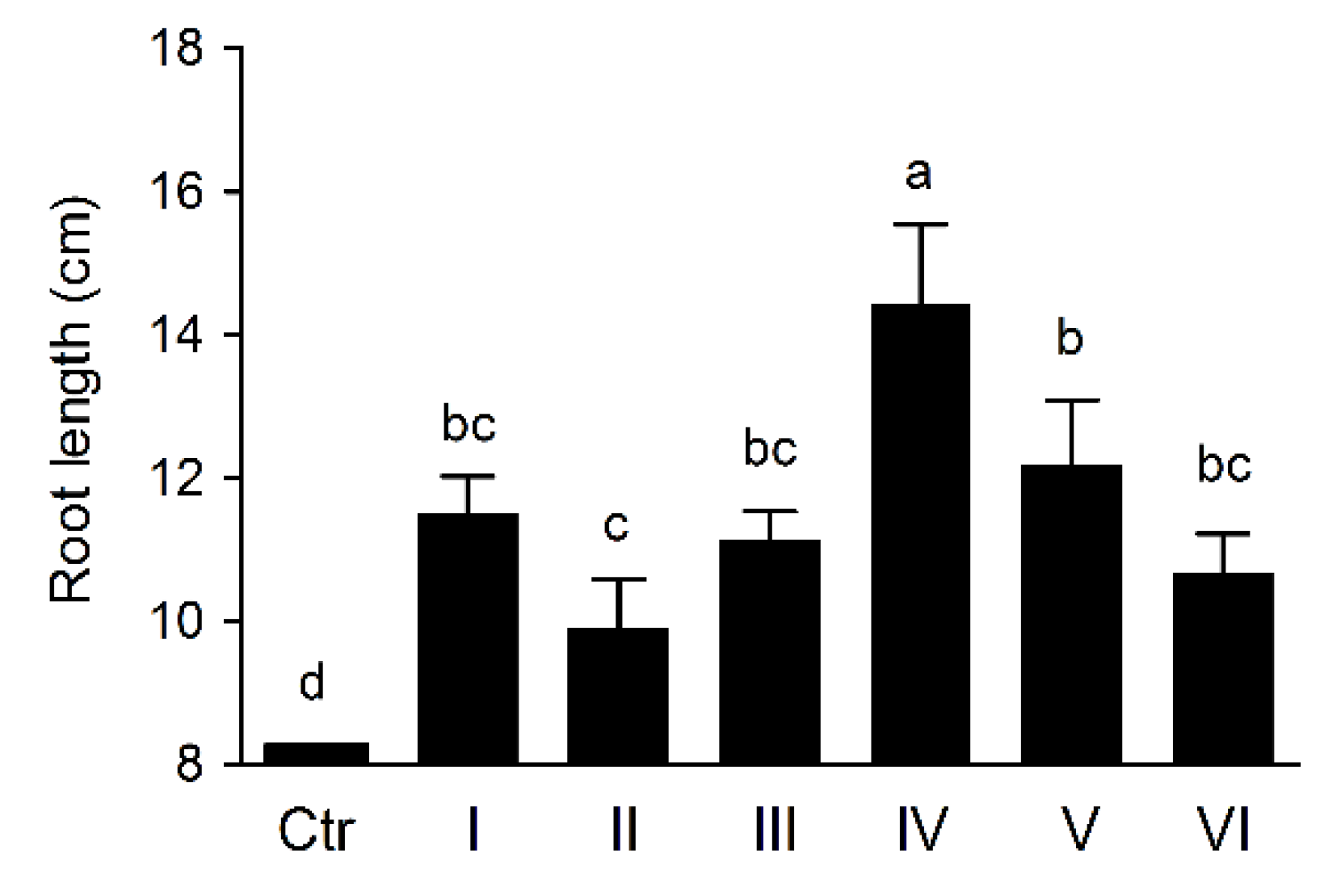
3.7. Wheat Rhizosphere Colonization and Growth Promoting by Pseudomonas sp. UW4 Strains in Pot Trial
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Allard-Massicotte, R.; Tessier, L.; Lécuyer, F.; Lakshmanan, V.; Lucier, J.F.; Garneau, D.; Caudwell, L.; Vlamakis, H.; Bais, H.P.; Beauregard, P.B. Bacillus subtilis early colonization of Arabidopsis thaliana roots involves multiple chemotaxis receptors. mBio 2016, 7, e01664-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SubbaRao, N.S. Biofetilizers in Agriculture and Forestry, 3rd ed.; International Science Publishers: New York, NY, USA, 1993; p. 242. [Google Scholar]
- Kaur, C.; Selvakumar, G.; Ganeshamurthy, A.N. Rhizocompetence of applied bioinoculants. In Plant-Microbe Interactions in Agro-Ecological Perspectives; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Springer: Singapore, 2017; Volume 1, pp. 501–511. [Google Scholar]
- Berninger, T.; González López, Ó.; Bejarano, A.; Preininger, C.; Sessitsch, A. Maintenance and assessment of cell viability in formulation of non-sporulating bacterial inoculants. Microb. Biotechnol. 2018, 11, 277–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Zhang, J.; Shen, C.; Li, H.; Qiu, L. 1-aminocyclopropane-1-carboxylate: A novel and strong chemoattractant for the plant beneficial rhizobacterium Pseudomonas putida UW4. Mol. Plant Microbe Interact. 2019, 32, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Scharf, B.E.; Hynes, M.F.; Alexandre, G.M. Chemotaxis signaling systems in model beneficial plant-bacteria associations. Plant Mol. Biol. 2016, 90, 549–559. [Google Scholar] [CrossRef]
- Grichko, V.P.; Glick, B.R. Amelioration of flooding stress by ACC deaminase-containing plant growth-promoting bacteria. Plant Physiol. Biochem. 2001, 39, 11–17. [Google Scholar] [CrossRef]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Gamalero, E.; Berta, G.; Massa, N.; Glick, B.R.; Lingua, G. Synergistic interactions between the ACC deaminase-producing bacterium Pseudomonas putida UW4 and the AM fungus Gigaspora rosea positively affect cucumber plant growth. FEMS Microbiol. Ecol. 2008, 64, 459–467. [Google Scholar] [CrossRef]
- Holguin, G.; Glick, B.R. Expression of the ACC deaminase gene from Enterobacter cloacae UW4 in Azospirillum brasilense. Microb. Ecol. 2001, 41, 281–288. [Google Scholar] [CrossRef]
- Nascimento, F.; Brígido, C.; Alho, L.; Glick, B.R.; Oliveira, S. Enhanced chickpea growth-promotion ability of a Mesorhizobium strain expressing an exogenous ACC deaminase gene. Plant Soil 2012, 353, 221–230. [Google Scholar] [CrossRef]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Changes in the population of seed bacteria of transgenerationally Cd-exposed Arabidopsis thaliana. Plant Biol. 2013, 15, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Pashapour, S.; Besharati, H.; Rezazade, M.; Alimadadi, A.; Ebrahimi, N. Activity screening of plant growth promoting rhizobacteria isolated from alfalfa rhizosphere. Biol. J. Microorg. 2016, 4, 65–68. [Google Scholar]
- Wong-Ng, J.; Celani, A.; Vergassola, M. Exploring the function of bacterial chemotaxis. Curr. Opin. Microbial. 2018, 45, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Baumgartner, J.W.; Hazelbauer, G.L. High-and low-abundance chemoreceptors in Escherichia coli: Differential activities associated with closely related cytoplasmic domains. J. Bacteriol. 1997, 179, 6714–6720. [Google Scholar] [CrossRef] [Green Version]
- Kalinin, Y.; Neumann, S.; Sourjik, V.; Wu, M. Responses of Escherichia coli bacteria to two opposing chemoattractant gradients depend on the chemoreceptor ratio. J. Bacteriol. 2010, 192, 1796–1800. [Google Scholar] [CrossRef] [Green Version]
- Lybarger, S.R.; Nair, U.; Lilly, A.A.; Hazelbauer, G.L.; Maddock, J.R. Clustering requires modified methyl-accepting sites in low-abundance but not high-abundance chemoreceptors of Escherichia coli. Mol. Microbiol. 2005, 56, 1078–1086. [Google Scholar] [CrossRef]
- Zhulin, I.B.; Armitage, J.P. Motility, chemokinesis, and methylation-independent chemotaxis in Azospirillum brasilense. J. Bacteriol. 1993, 175, 952–958. [Google Scholar] [CrossRef] [Green Version]
- Braatsch, S.; Helmark, S.; Kranz, H.; Koebmann, B.; Ruhdal Jensen, P. Escherichia coli strains with promoter libraries constructed by Red/ET recombination pave the way for transcriptional fine-tuning. BioTechniques 2008, 45, 335–337. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Glick, B.R. Transcriptional regulation of the Enterobacter cloacae UW4 1-aminocyclopropane-1-carboxylate (ACC) deaminase gene (acdS). Can. J. Microbiol. 2001, 47, 359–367. [Google Scholar] [CrossRef]
- Kovach, M.E.; Phillips, R.W.; Elzer, P.H.; Peterson, K.M. pBBR1MCS: A broad-host-range cloning vector. BioTechniques 1994, 16, 800–802. [Google Scholar]
- Li, J.; Ovakim, D.H.; Charles, T.C.; Glick, B.R. An ACC deaminase minus mutant of Enterobacter cloacae UW4 no longer promotes root elongation. Curr. Microbiol. 2000, 41, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Figurski, D.H.; Helinski, D.R. Replication of an origin-containing derivative of plasmid RK2 dependent on a plasmid function provided in trans. Proc. Natl. Acad. Sci. USA 1979, 76, 1648–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukunaga, S.; Horiguchi, H.; Iwamoto, M.; Shigenobu, Y.; Shinozawa, T. Chemotaxis for methamphetamine in Escherichia coli. Microbio. Immunol. 1988, 32, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Kumita, W.; Saito, R.; Sato, K.; Ode, T.; Moriya, K.; Koike, K.; Chida, T.; Okamura, N. Molecular characterizations of carbapenem and ciprofloxacin resistance in clinical isolates of Pseudomonas putida. J. Infect. Chemother. 2009, 15, 6–12. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Blaha, D.; Prigent-Combaret, C.; Mirza, M.S.; Moënne-Loccoz, Y. Phylogeny of the 1-aminocyclopropane-1-carboxylic acid deaminase-encoding gene acdS in phytobeneficial and pathogenic Proteobacteria and relation with strain biogeography. FEMS Microbiol. Ecol. 2006, 56, 455–470. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Qiu, C.; Huang, T.; Zhou, W.; Qi, Y.; Gao, Y.; Shen, J.; Qiu, L. Effect of 1-aminocyclopropane-1-carboxylic acid deaminase producing bacteria on the hyphal growth and primordium initiation of Agaricusbisporus. Fungal Ecol. 2013, 6, 110–118. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Loyola-Vargas, V.M.; Broeckling, C.D.; Badri, D.; Vivanco, J.M. Effect of transporters on the secretion of phytochemicals by the roots of Arabidopsis thaliana. Planta 2007, 225, 301–310. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth promoting rhizobacteria. Physiol. Plant 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Fischer, S.E.; Jofré, E.C.; Cordero, P.V.; Manero, F.J.G.; Mori, G.B. Survival of native Pseudomonas in soil and wheat rhizosphere and antagonist activity against plant pathogenic fungi. Antonie van Leeuwenhoek 2010, 97, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, H.; Niu, Y.; Hu, M.; Qi, F.; Zhang, M.; Huang, J.; Tao, Y. Determination of temperature sensitive plasmid copy number in Escherichia coli by absolute and relative real time quantitation PCR. Int. Cardivasc. Res. J. 2017, 10, e14600. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Endres, R.G. A minimal model for metabolism-dependent chemotaxis in Rhodobacter sphaeroides. Interface Focus 2014, 4, 20140002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, B.L. Role of proton motive force in sensory transduction in bacteria. Ann. Rev. Microbiol. 1983, 37, 551–573. [Google Scholar] [CrossRef] [PubMed]
- Bal, H.B.; Nayak, L.; Das, S.; Adhya, T.K. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2013, 366, 93–105. [Google Scholar] [CrossRef]
- Shaharoona, B.; Arshad, M.; Zahir, Z.A. Effect of plant growth promoting rhizobacteria containing ACC-deaminase on maize (Zea mays L.) growth under axenic conditions and on nodulation in mung bean (Vigna radiata L.). Lett. Appl. Microbiol. 2006, 42, 155–159. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Zahir, Z.A.; Naveed, M.; Arshad, M. Preliminary investigations on inducing salt tolerance in maize through inoculation with rhizobacteria containing ACC deaminase activity. Can. J. Microbiol. 2007, 53, 1141–1149. [Google Scholar] [CrossRef]
- Belimov, A.A.; Dodd, I.C.; Safronova, V.I.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Davies, W.J.; Tikhonovich, I.A. Rhizobacteria that produce auxins and contain 1-amino-cyclopropane-1-carboxylic acid deaminase decrease amino acid concentrations in the rhizosphere and improve growth and yield of well-watered and water-limited potato (Solanum tuberosum). Ann. Appl. Biol. 2015, 167, 11–25. [Google Scholar] [CrossRef]
- O’Banion, B.; O’Neal, L.; Alexandre, G.; Lebeis, S. Bridging the gap between single-strain and community-level plant-microbe chemical interactions. Mol. Plant-Microbe Interact. 2019, in press. [Google Scholar] [CrossRef] [Green Version]
- Begonia, M.F.T.; Kremer, R.J. Chemotaxis of deleterious rhizobacteria to velvetleaf (Abutilon theophrasti Medik.) seeds and seedlings. FEMS Microbial. Ecol. 1994, 15, 227–235. [Google Scholar] [CrossRef]
- Yang, Y.M.; Pollard, A.; Höfler, C.; Poschet, G.; Wirtz, M.; Hell, R.; Sourjik, V. Relation between chemotaxis and consumption of amino acids in bacteria. Mol. Microbiol. 2015, 96, 1272–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Li, T.; Liu, W.; Zhang, Y.; Shang, D.; Gao, Y.; Qi, Y.; Qiu, L. Enhancing the 1-Aminocyclopropane-1-Carboxylate Metabolic Rate of Pseudomonas sp. UW4 Intensifies Chemotactic Rhizocompetence. Microorganisms 2020, 8, 71. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010071
Gao X, Li T, Liu W, Zhang Y, Shang D, Gao Y, Qi Y, Qiu L. Enhancing the 1-Aminocyclopropane-1-Carboxylate Metabolic Rate of Pseudomonas sp. UW4 Intensifies Chemotactic Rhizocompetence. Microorganisms. 2020; 8(1):71. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010071
Chicago/Turabian StyleGao, Xiyang, Tao Li, Wenliang Liu, Yan Zhang, Di Shang, Yuqian Gao, Yuancheng Qi, and Liyou Qiu. 2020. "Enhancing the 1-Aminocyclopropane-1-Carboxylate Metabolic Rate of Pseudomonas sp. UW4 Intensifies Chemotactic Rhizocompetence" Microorganisms 8, no. 1: 71. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010071