Antimicrobial Sensitivity Testing of Mycoplasma bovis Isolates Derived from Western Canadian Feedlot Cattle

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Handling
2.2. Mycoplasma spp. Isolation
2.3. Species Identification
2.4. Antimicrobial Susceptibility Testing
2.5. Breakpoint Interpretation Guidelines
2.6. Data Analysis
3. Results
3.1. Sample History
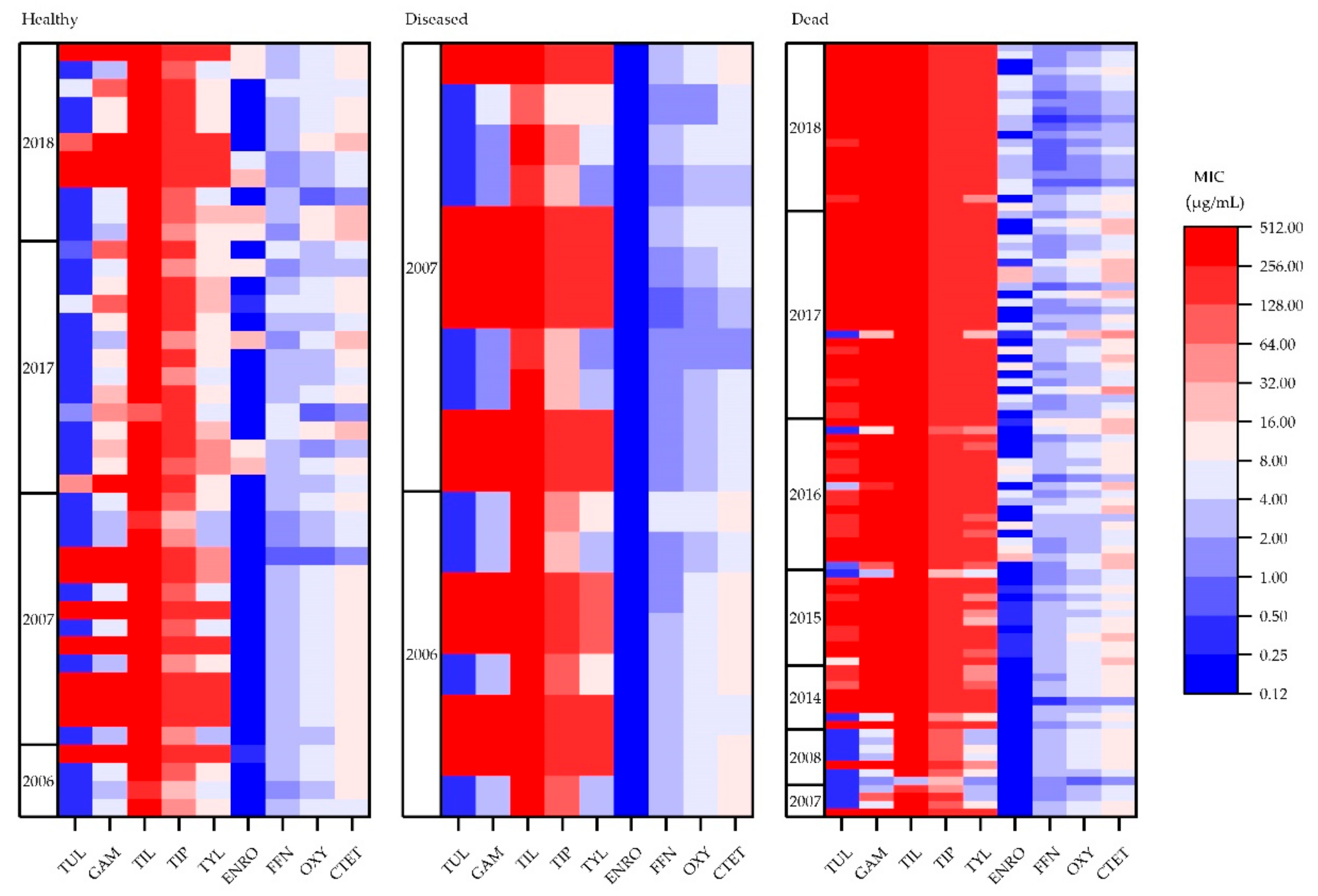
3.2. Antimicrobial Susceptibility Testing
3.3. Antimicrobial Susceptibility Testing―Anatomical Region
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Year | Feedlot | Cohort | Isolate ID | Animal # | Anatomical Region | Health Status | DOF |
|---|---|---|---|---|---|---|---|
| 2006 | F6 | C13 | MPLM0648 | 1 | DNP | Diseased | 90 |
| MPLM0654 | 2 | DNP | Diseased | 90 | |||
| F14 | C21 | MPLM0649 | 3 | DNP | Diseased | 90 | |
| MPLM0786 | 4 | DNP | Diseased | 90 | |||
| F16 | C23 | MPLM0845 | 5 | DNP | Healthy | ||
| F20 | C29 | MPLM0810 | 6 | DNP | Diseased | 150 | |
| MPLM0642 | 7 | DNP | Diseased | 150 | |||
| MPLM0651 | 8 | DNP | Diseased | 150 | |||
| F30 | C40 | MPLM0643 | 9 | DNP | Healthy | 140 | |
| MPLM0811 | 10 | DNP | Healthy | 14 | |||
| MPLM0813 | 11 | DNP | Diseased | ||||
| MPLM0640 | 12 | DNP | Healthy | 14 | |||
| N = 5 | N = 5 | N = 12 | N = 12 | ||||
| 2007 | F1 | C1 | MPLM0788 | 13 | Lung | Dead | 47 |
| F8 | C15 | MPLM0632 | 14 | DNP | Dead | ||
| F9 | C16 | MPLM0795 | 15 | Lung | Dead | ||
| MPLM0631 | Joint | Dead | 60 | ||||
| F15 | C22 | MPLM0664 | 16 | DNP | Diseased | 15 | |
| MPLM0660 | 17 | DNP | Diseased | 0 | |||
| MPLM0665 | 18 | DNP | Diseased | 21 | |||
| MPLM0644 | 19 | DNP | Diseased | 21 | |||
| MPLM0669 | 20 | DNP | Dead | 41 | |||
| MPLM0661 | 21 | DNP | Diseased | 12 | |||
| MPLM0647 | 22 | DNP | Diseased | 16 | |||
| MPLM0657 | 23 | DNP | Diseased | 45 | |||
| MPLM0713 | 24 | DNP | Healthy | 15 | |||
| MPLM0714 | 25 | DNP | Healthy | 15 | |||
| MPLM0684 | 26 | DNP | Healthy | 15 | |||
| MPLM0652 | 27 | DNP | Diseased | 10 | |||
| MPLM0662 | 28 | DNP | Diseased | 21 | |||
| MPLM0700 | 29 | DNP | Healthy | 15 | |||
| MPLM0703 | 30 | DNP | Healthy | 15 | |||
| MPLM0715 | 31 | DNP | Healthy | 15 | |||
| MPLM0706 | 32 | DNP | Healthy | 15 | |||
| MPLM0645 | 33 | DNP | Diseased | 10 | |||
| MPLM0666 | 34 | DNP | Diseased | 10 | |||
| MPLM0692 | 35 | DNP | Healthy | 15 | |||
| MPLM0698 | 36 | DNP | Healthy | 15 | |||
| F16 | C24 | MPLM0670 | 37 | DNP | Healthy | 14 | |
| MPLM0671 | 38 | DNP | Healthy | 14 | |||
| C25 | MPLM0667 | 39 | DNP | Healthy | 90 | ||
| MPLM0668 | 40 | DNP | Healthy | 90 | |||
| MPLM0646 | 41 | DNP | Healthy | 90 | |||
| N = 5 | N = 6 | N = 30 | N = 29 | ||||
| 2008 | F7 | C14 | MPLM0781 | 42 | Lung | Dead | |
| MPLM0602 | DNP | Dead | 60 | ||||
| MPLM0784 | 43 | Lung | Dead | ||||
| F8 | C15 | MPLM0630 | 44 | Joint | Dead | 30 | |
| F22 | C31 | MPLM0789 | 45 | Lung | Dead | ||
| MPLM0792 | 46 | Lung | Dead | ||||
| F23 | C32 | MPLM0783 | 47 | Lung | Dead | ||
| MPLM0790 | 48 | Lung | Dead | ||||
| N = 4 | N = 4 | N = 8 | N = 7 | ||||
| 2014 | F2 | C5 | MYCO066 | 49 | Lung | Dead | 35 |
| MYCO096 | 50 | Lung | Dead | 96 | |||
| F3 | C6 | MP0219 | 51 | Lung | Dead | 39 | |
| MP0209 | Joint | Dead | 39 | ||||
| F13 | C20 | MYCO081 | 52 | Joint | Dead | 31 | |
| F17 | C26 | MP0064 | 53 | Lung | Dead | 193 | |
| F19 | C28 | MPLM0636 | 54 | Lung | Dead | 93 | |
| F21 | C30 | MYCO062 | 55 | Lung | Dead | 76 | |
| MYCO076 | 56 | Lung | Dead | 24 | |||
| N = 6 | N = 6 | N = 9 | N = 8 | ||||
| 2015 | F3 | C7 | MP0071 | 57 | Lung | Dead | 42 |
| MP0063 | 58 | Lung | Dead | 42 | |||
| MPLM0637 | 59 | Lung | Dead | 31 | |||
| MP0058 | 60 | Lung | Dead | 31 | |||
| MP0073 | 61 | Lung | Dead | 67 | |||
| MP0077 | 62 | Lung | Dead | 61 | |||
| MP0079 | 63 | Lung | Dead | 67 | |||
| MPLM0638 | Joint | Dead | 67 | ||||
| MP0183 | 64 | Joint | Dead | 217 | |||
| F11 | C18 | MPLM0634 | 65 | Lung | Dead | ||
| F13 | C20 | MP0057 | 66 | Lung | Dead | 80 | |
| F21 | C30 | MP0070 | 67 | Lung | Dead | 20 | |
| F27 | C37 | MP0075 | 68 | Lung | Dead | 46 | |
| N = 5 | N = 5 | N = 13 | N = 12 | ||||
| 2016 | F1 | C2 | MPLM0041 | 69 | Lung | Dead | 46 |
| MPLM0042 | Joint | Dead | 46 | ||||
| MPLM0033 | 70 | Lung | Dead | 48 | |||
| MPLM0034 | Joint | Dead | 48 | ||||
| MPLM0029 | 71 | Lung | Dead | ||||
| MPLM0030 | Joint | Dead | |||||
| F18 | C27 | MPLM0021 | 72 | Lung | Dead | 55 | |
| MPLM0022 | Joint | Dead | 55 | ||||
| MPLM0009 | 73 | Lung | Dead | 22 | |||
| MPLM0010 | Joint | Dead | 22 | ||||
| MPLM0013 | 74 | Lung | Dead | 26 | |||
| MPLM0014 | Joint | Dead | 26 | ||||
| MPLM0025 | 75 | Lung | Dead | 15 | |||
| MPLM0026 | Joint | Dead | 15 | ||||
| MPLM0037 | 76 | Lung | Dead | 81 | |||
| MPLM0017 | 77 | Lung | Dead | 43 | |||
| MPLM0018 | Joint | Dead | 43 | ||||
| MPLM0015 | 78 | Lung | Dead | 31 | |||
| MPLM0016 | Joint | Dead | 31 | ||||
| MPLM0007 | 79 | Lung | Dead | 9 | |||
| MPLM0008 | Joint | Dead | 9 | ||||
| MPLM0035 | 80 | Lung | Dead | 38 | |||
| MPLM0036 | Joint | Dead | 38 | ||||
| MPLM0031 | 81 | Lung | Dead | 34 | |||
| MPLM0019 | 82 | Lung | Dead | 55 | |||
| MPLM0020 | Joint | Dead | 55 | ||||
| MPLM0011 | 83 | Lung | Dead | 22 | |||
| MPLM0012 | Joint | Dead | 22 | ||||
| MPLM0039 | 84 | Lung | Dead | 50 | |||
| MPLM0040 | Joint | Dead | 50 | ||||
| F19 | C28 | MPLM0134 | 85 | Lung | Dead | 60 | |
| MPLM0135 | Joint | Dead | 60 | ||||
| MPLM0132 | 86 | Lung | Dead | 30 | |||
| F28 | C38 | MPLM0003 | 87 | Lung | Dead | 28 | |
| MPLM0004 | Joint | Dead | 28 | ||||
| N = 4 | N = 4 | N = 35 | N = 19 | ||||
| 2017 | F1 | C3 | MPLM0114 | 88 | Lung | Dead | 33 |
| MPLM0102 | 89 | Lung | Dead | 40 | |||
| MPLM0103 | Joint | Dead | 40 | ||||
| MPLM0160 | 90 | Lung | Dead | 52 | |||
| MPLM0093 | 91 | Lung | Dead | 37 | |||
| MPLM0094 | Joint | Dead | 37 | ||||
| MPLM0090 | 92 | Lung | Dead | 45 | |||
| MPLM0091 | Joint | Dead | 45 | ||||
| MPLM0111 | 93 | Lung | Dead | 17 | |||
| MPLM0112 | Joint | Dead | 17 | ||||
| MPLM0087 | 94 | Lung | Dead | 60 | |||
| MPLM0088 | Joint | Dead | 60 | ||||
| MPLM0105 | 95 | Lung | Dead | 54 | |||
| MPLM0106 | Joint | Dead | 54 | ||||
| MPLM0064 | 96 | Joint | Dead | ||||
| MPLM0164 | 97 | Joint | Dead | 30 | |||
| MPLM0143 | 98 | Joint | Dead | 24 | |||
| MPLM0066 | 99 | Lung | Dead | ||||
| MPLM0067 | Joint | Dead | |||||
| MPLM0108 | 100 | Lung | Dead | 44 | |||
| MPLM0145 | 101 | Lung | Dead | 42 | |||
| MPLM0146 | Joint | Dead | 42 | ||||
| MPLM0154 | 102 | Lung | Dead | 40 | |||
| MPLM0155 | Joint | Dead | 40 | ||||
| MPLM0073 | 103 | Joint | Dead | ||||
| MPLM0157 | 104 | Lung | Dead | 46 | |||
| MPLM0158 | Joint | Dead | 46 | ||||
| MPLM0084 | 105 | Lung | Dead | 33 | |||
| MPLM0085 | Joint | Dead | 33 | ||||
| MPLM0148 | 106 | Lung | Dead | 48 | |||
| MPLM0149 | Joint | Dead | 48 | ||||
| MPLM0069 | 107 | Lung | Dead | ||||
| MPLM0070 | Joint | Dead | |||||
| F4 | C8 | MPLM0831 | 108 | DNP | Healthy | 0 | |
| F5 | C10 | MPLM0833 | 109 | DNP | Healthy | 0 | |
| MPLM0834 | 110 | DNP | Healthy | 0 | |||
| MPLM0837 | 111 | DNP | Healthy | 0 | |||
| MPLM0838 | 112 | DNP | Healthy | 0 | |||
| C11 | MPLM0832 | 113 | DNP | Healthy | 0 | ||
| MPLM0835 | 114 | DNP | Healthy | 0 | |||
| C12 | MPLM0839 | 115 | DNP | Healthy | 0 | ||
| F10 | C17 | MPLM0081 | 116 | Lung | Dead | 180 | |
| F12 | C19 | MPLM0167 | 117 | Joint | Dead | ||
| MPLM0057 | 118 | Lung | Dead | ||||
| MPLM0058 | Joint | Dead | |||||
| MPLM0060 | 119 | Lung | Dead | ||||
| MPLM0061 | Joint | Dead | |||||
| MPLM0054 | 120 | Lung | Dead | ||||
| MPLM0076 | 121 | Joint | Dead | ||||
| F24 | C33 | MPLM0815 | 122 | DNP | Healthy | 0 | |
| MPLM0819 | 123 | DNP | Healthy | 0 | |||
| MPLM0820 | 124 | DNP | Healthy | 0 | |||
| F26 | C36 | MPLM0821 | 125 | DNP | Healthy | 0 | |
| MPLM0822 | 126 | DNP | Healthy | 0 | |||
| F29 | C39 | MPLM0826 | 127 | DNP | Healthy | 0 | |
| N = 8 | N = 10 | N = 55 | N = 40 | ||||
| 2018 | F1 | C4 | MPLM0563 | 128 | Lung | Dead | |
| MPLM0533 | 129 | Lung | Dead | 65 | |||
| MPLM0534 | Joint | Dead | 65 | ||||
| MPLM0535 | DNP | Dead | 65 | ||||
| MPLM0569 | 130 | Lung | Dead | 27 | |||
| MPLM0552 | 131 | Lung | Dead | 36 | |||
| MPLM0553 | DNP | Dead | 36 | ||||
| MPLM0611 | 132 | Lung | Dead | 15 | |||
| MPLM0622 | 133 | DNP | Dead | 33 | |||
| MPLM0566 | 134 | Lung | Dead | 30 | |||
| MPLM0567 | DNP | Dead | 30 | ||||
| MPLM0537 | 135 | Lung | Dead | 27 | |||
| MPLM0538 | Joint | Dead | 27 | ||||
| MPLM0539 | DNP | Dead | 27 | ||||
| MPLM0593 | 136 | Lung | Dead | 44 | |||
| MPLM0555 | 137 | Lung | Dead | 60 | |||
| MPLM0556 | Joint | Dead | 60 | ||||
| MPLM0549 | 138 | Lung | Dead | 34 | |||
| MPLM0587 | 139 | Lung | Dead | 34 | |||
| MPLM0588 | DNP | Dead | 34 | ||||
| MPLM0584 | 140 | Lung | Dead | 44 | |||
| MPLM0624 | 141 | Lung | Dead | 33 | |||
| MPLM0625 | DNP | Dead | 33 | ||||
| MPLM0541 | 142 | Lung | Dead | 71 | |||
| MPLM0542 | Joint | Dead | 71 | ||||
| MPLM0543 | DNP | Dead | 71 | ||||
| MPLM0545 | 143 | Lung | Dead | 56 | |||
| MPLM0546 | Joint | Dead | 56 | ||||
| MPLM0547 | DNP | Dead | 56 | ||||
| MPLM0608 | 144 | Lung | Dead | 61 | |||
| MPLM0609 | DNP | Dead | 61 | ||||
| MPLM0582 | 145 | DNP | Dead | 61 | |||
| MPLM0559 | 146 | Lung | Dead | 33 | |||
| MPLM0560 | Joint | Dead | 33 | ||||
| MPLM0578 | 147 | Lung | Dead | 15 | |||
| MPLM0579 | DNP | Dead | 15 | ||||
| F4 | C9 | MPLM0827 | 148 | DNP | Healthy | 0 | |
| MPLM0828 | 149 | DNP | Healthy | 0 | |||
| MPLM0829 | 150 | DNP | Healthy | 0 | |||
| MPLM0830 | 151 | DNP | Healthy | 0 | |||
| F24 | C34 | MPLM0816 | 152 | DNP | Healthy | 0 | |
| MPLM0817 | 153 | DNP | Healthy | 0 | |||
| MPLM0818 | 154 | DNP | Healthy | 0 | |||
| F25 | C35 | MPLM0627 | 155 | Lung | Dead | ||
| MPLM0628 | DNP | Dead | |||||
| F29 | C40 | MPLM0823 | 156 | DNP | Healthy | 0 | |
| MPLM0824 | 157 | DNP | Healthy | 0 | |||
| MPLM0825 | 158 | DNP | Healthy | 0 | |||
| F31 | C12 | MPLM0836 | 159 | DNP | Healthy | 0 | |
| N = 6 | N = 6 | N = 49 | N = 32 |
References
- Griffin, D.; Chengappa, M.M.; Kuszak, J.; McVey, D.S. Bacterial pathogens of the bovine respiratory disease complex. Vet. Clin. N. Am. Food Anim. 2010, 26, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Panciera, R.J.; Confer, A.W. Pathogenesis and pathology of bovine pneumonia. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 191–214. [Google Scholar] [CrossRef] [PubMed]
- Nickell, J.S.; White, B.J. Metaphylactic antimicrobial therapy for bovine respiratory disease in stocker and feedlot cattle. Vet. Clin. N. Am. Food Anim. 2010, 26, 285–301. [Google Scholar] [CrossRef] [PubMed]
- Brault, S.A.; Hannon, S.J.; Gow, S.P.; Otto, S.J.G.; Booker, C.W.; Morley, P.S. Calculation of antimicrobial use indicators in beef feedlots—Effects of choice of metric and standardized values. Front. Vet. Sci. 2019, 6, 330. [Google Scholar] [CrossRef]
- Brault, S.A.; Hannon, S.J.; Gow, S.P.; Warr, B.N.; Withell, J.; Song, J.; Williams, C.M.; Otto, S.J.G.; Booker, C.W.; Morley, P.S. Antimicrobial use on 36 beef feedlots in western Canada: 2008–2012. Front. Vet. Sci. 2019, 6, 329. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, R.A.J.; Ayling, R.D. Mycoplasma bovis: Disease, diagnosis, and control. Res. Vet. Sci. 2003, 74, 105–112. [Google Scholar] [CrossRef]
- Gagea, M.I.; Bateman, K.G.; Shanahan, R.A.; van Dreumel, T.; McEwen, B.J.; Carman, S.; Archambault, M.; Caswell, J.L. Naturally occurring mycoplasma bovis—Associated pneumonia and polyarthritis in feedlot beef calves. J. Vet. Diagn. Investig. 2006, 18, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horwood, P.; Schibrowski, M.; Fowler, E.; Gibson, J.; Barnes, T.; Mahony, T. Is mycoplasma bovis a missing component of the bovine respiratory disease complex in australia? Aust. Vet. J. 2014, 92, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Grissett, G.P.; White, B.J.; Larson, R.L. Structured literature review of responses of cattle to viral and bacterial pathogens causing bovine respiratory disease complex. J. Vet. Intern. Med. 2015, 29, 770–780. [Google Scholar] [CrossRef]
- Booker, C.W.; Abutarbush, S.M.; Morley, P.S.; Jim, G.K.; Pittman, T.J.; Schunicht, O.C.; Perrett, T.; Wildman, B.K.; Fenton, R.K.; Guichon, P.T.; et al. Microbiological and histopathological findings in cases of fatal bovine respiratory disease of feedlot cattle in western canada. Can. Vet. J. 2008, 49, 473–481. [Google Scholar]
- Radaelli, E.; Luini, M.; Loria, G.R.; Nicholas, R.A.J.; Scanziani, E. Bacteriological, serological, pathological and immunohistochemical studies of mycoplasma bovis respiratory infection in veal calves and adult cattle at slaughter. Res. Vet. Sci. 2008, 85, 282–290. [Google Scholar] [CrossRef]
- Caswell, J.L.; Bateman, K.G.; Cai, H.Y.; Castillo-Alcala, F. Mycoplasma bovis in respiratory disease of feedlot cattle. Vet. Clin. N. Am. Food Anim. 2010, 26, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, R.A.J. Bovine mycoplasmosis: Silent and deadly. Vet. Rec. 2011, 168, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Caswell, J.L.; Archambault, M. Mycoplasma bovis pneumonia in cattle. Anim. Health Res. Rev. 2007, 8, 161–186. [Google Scholar] [CrossRef] [PubMed]
- Romváry, J.; Rózsa, J.; Stipkovits, L.; Mészaros, J. Incidence of diseases due to Mycoplasma bovis in a cattle herd. Pneumo-arthritis syndrome in calves. Acta Vet. Acad. Sci. Hung 1979, 27, 29–37. [Google Scholar]
- Francoz, D.; Fortin, M.; Fecteau, G.; Messier, S. Determination of mycoplasma bovis susceptibilities against six antimicrobial agents using the E test method. Vet. Microbiol. 2005, 105, 57–64. [Google Scholar] [CrossRef]
- Cai, H.Y.; McDowall, R.; Parker, L.; Kaufman, E.I.; Caswell, J.L. Changes in antimicrobial susceptibility profiles of mycoplasma bovis over time. Can. Vet. J. 2019, 83, 34–41. [Google Scholar]
- Hendrick, S.H.; Bateman, K.G.; Rosengren, L.B. The effect of antimicrobial treatment and preventive strategies on bovine respiratory disease and genetic relatedness and antimicrobial resistance of mycoplasma bovis isolates in a western canadian feedlot. Can. Vet. J. 2013, 54, 1146. [Google Scholar]
- Anholt, R.M.; Klima, C.; Allan, N.; Matheson-Bird, H.; Schatz, C.; Ajitkumar, P.; Otto, S.J.; Peters, D.; Schmid, K.; Olson, M.; et al. Antimicrobial susceptibility of bacteria that cause bovine respiratory disease complex in alberta, canada. Front. Vet. Sci. 2017, 4, 207. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Robinson, D.; Bébéar, C. Antibiotic susceptibilities of mycoplasmas and treatment of mycoplasmal infections. J. Antimicrob. Chemother. 1997, 40, 622–630. [Google Scholar] [CrossRef] [Green Version]
- Lysnyansky, I.; Ayling, R.D. Mycoplasma bovis: Mechanisms of resistance and trends in antimicrobial susceptibility. Front. Microbiol. 2016, 7, 595. [Google Scholar] [CrossRef] [PubMed]
- Register, K.B.; Boatwright, W.D.; Gesy, K.M.; Thacker, T.C.; Jelinski, M.D. Mistaken identity of an open reading frame proposed for PCR-based identification of Mycoplasma bovis and the effect of polymorphisms and insertions on assay performance. J. Vet. Diagn. Investig. 2018, 30, 637–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, K.; McAuliffe, L.; Ayling, R.D.; Nicholas, R.A.J. Rapid detection of Mycoplasma dispar and M. bovirhinis using allele specific polymerase chain reaction protocols. FEMS Microbiol. Lett. 2004, 241, 103–107. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals, 5th ed.; CLSI standard VET01; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- CLSI. Methods for Antimicrobial Susceptibility Testing for Human Mycoplasmas Approved Guideline; CLSI document M43-A.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals, 4th ed.; CLSI supplement VET08; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Lerner, U.; Amram, E.; Ayling, R.D.; Mikula, I.; Gerchman, I.; Harrus, S.; Teff, D.; Yogev, D.; Lysnyansky, I. Acquired resistance to the 16-membered macrolides tylosin and tilmicosin by Mycoplasma bovis. Vet. Microbiol. 2014, 168, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Government of Canada, Notice of Compliance Information. Available online: https://health-products.canada.ca/noc-ac/index-eng.jsp (accessed on 10 December 2019).
- Rosenbusch, R.F.; Kinyon, J.M.; Apley, M.; Funk, N.D.; Smith, S.; Hoffman, L.J. In vitro antimicrobial inhibition profiles of mycoplasma bovis isolates recovered from various regions of the united states from 2002 to 2003. J. Vet. Diagn. Investig. 2005, 17, 436–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier-Bouchardon, A.V.; Ferré, S.; Le Grand, D.; Paoli, A.; Gay, E.; Poumarat, F. Overall decrease in the susceptibility of mycoplasma bovis to antimicrobials over the past 30 years in france. PLoS ONE 2014, 9, e87672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulyok, K.M.; Kreizinger, Z.; Fekete, L.; Hrivnák, V.; Magyar, T.; Jánosi, S.; Schweitzer, N.; Turcsányi, I.; Makrai, L.; Erdélyi, K.; et al. Antibiotic susceptibility profiles of mycoplasma bovis strains isolated from cattle in hungary, central europe. BMC Vet. Res. 2014, 10, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, J.W.; Viel, L.; Bateman, K.G.; Rosendal, S. Changes in the bacterial flora of the upper and lower respiratory tracts and bronchoalveolar lavage differential cell counts in feedlot calves treated for respiratory diseases. Can. Vet. J. 1992, 56, 177–183. [Google Scholar]
- Ayling, R.D.; Baker, S.E.; Nicholas, R.A.J.; Peek, M.L.; Simon, A.J. Comparison of in vitro activity of danofloxacin, florfenicol, oxytetracycline, spectinomycin and tilmicosin against recent field isolates of mycoplasma bovis. Vet. Rec. 2000, 146, 745–747. [Google Scholar] [CrossRef]
- Maunsell, F.P.; Donovan, G.A. Mycoplasma bovis infections in young calves. Vet. Clin. N. Am. Food Anim. 2009, 25, 139–177. [Google Scholar] [CrossRef] [PubMed]
- Sulyok, K.M.; Kreizinger, Z.; Wehmann, E.; Lysnyansky, I.; Bányai, K.; Marton, S.; Jerzsele, Á.; Rónai, Z.; Turcsányi, I.; Makrai, L.; et al. Mutations associated with decreased susceptibility to seven antimicrobial families in field and laboratory-derived mycoplasma bovis strains. Antimicrob. Agents Chemother. 2017, 61, e01983-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautier-Bouchardon, A.V. Antimicrobial resistance in mycoplasma spp. Microbiol. Spectrum. 2018, 6, 425–446. [Google Scholar] [CrossRef] [PubMed]
- Who Critically Important Antimicrobials for Human Medicine. 5th Revision. Available online: https://www.who.int/foodsafety/publications/cia2017.pdf?ua=1 (accessed on 21 December 2019).
- Cao, B.; Zhao, C.; Yin, Y.; Zhao, F.; Song, S.; Bai, L.; Zhang, J.; Liu, Y.; Zhang, Y.; Wang, H.; et al. High prevalence of macrolide resistance in mycoplasma pneumoniae isolates from adult and adolescent patients with respiratory tract infection in china. Clin. Infect. Dis. 2010, 51, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chironna, M.; Sallustio, A.; Esposito, S.; Perulli, M.; Chinellato, I.; Di Bari, C.; Quarto, M.; Cardinale, F. Emergence of macrolide-resistant strains during an outbreak of Mycoplasma pneumoniae infections in children. J. Antimicrob. Chemother. 2011, 66, 734–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, N.M.; Poehlsgaard, J.; Warrass, R.; Douthwaite, S. Inhibition of protein synthesis on the ribosome by tildipirosin compared with other veterinary macrolides. Antimicrob. Agents Chemother. 2012, 56, 6033–6036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poehlsgaard, J.; Andersen, N.M.; Warrass, R.; Douthwaite, S. Visualizing the 16-Membered ring macrolides tildipirosin and tilmicosin bound to their ribosomal site. ACS Chem. Biol. 2012, 7, 1351–1355. [Google Scholar] [CrossRef]
- Gerchman, I.; Levisohn, S.; Mikula, I.; Lysnyansky, I. In vitro antimicrobial susceptibility of mycoplasma bovis isolated in israel from local and imported cattle. Vet. Microbiol. 2009, 137, 268–275. [Google Scholar] [CrossRef]
- CLSI. Generation, Presentation, and Application of Antimicrobial Susceptibility Test Data for Bacteria of Animal Origin: A Report; CLSI Document VET05-R; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011. [Google Scholar]
- Amram, E.; Mikula, I.; Schnee, C.; Ayling, R.D.; Nicholas, R.A.J.; Rosales, R.S.; Harrus, S.; Lysnyansky, I. 16S rRNA gene mutations associated with decreased susceptibility to tetracycline in mycoplasma bovis. Antimicrob. Agents Chemother. 2015, 59, 796–802. [Google Scholar] [CrossRef] [Green Version]
- Singh, U.M.; Doig, P.A.; Ruhnke, H.L. Mycoplasma arthritis in calves. Can. Vet. J. 1971, 12, 183–185. [Google Scholar]
- Haines, D.M.; Martin, K.M.; Clark, E.G.; Jim, G.K.; Janzen, E.D. The immunohistochemical detection of mycoplasma bovis and bovine viral diarrhea virus in tissues of feedlot cattle with chronic, unresponsive respiratory disease and/or arthritis. Can. Vet. J. 2001, 42, 857–860. [Google Scholar]
- Watts, J.L.; Sweeney, M.T. Antimicrobial resistance in bovine respiratory disease pathogens: Measures, trends, and impact on efficacy. Vet. Clin. N. Am. Food Anim. 2010, 26, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Timsit, E.; Arcangioli, M.-A.; Bareille, N.; Seegers, H.; Assié, S. Transmission dynamics of mycoplasma bovis in newly received beef bulls at fattening operations. J. Vet. Diagn. Investig. 2012, 24, 1172–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereyre, S.; Charron, A.; Hidalgo-Grass, C.; Touati, A.; Moses, A.E.; Nir-Paz, R.; Bébéar, C. The spread of mycoplasma pneumoniae is polyclonal in both an endemic setting in france and in an epidemic setting in israel. PLoS ONE 2012, 7, e38585. [Google Scholar] [CrossRef] [Green Version]
- Heuvelink, A.; Reugebrink, C.; Mars, J. Antimicrobial susceptibility of mycoplasma bovis isolates from veal calves and dairy cattle in the netherlands. Vet. Microbiol. 2016, 189, 1–7. [Google Scholar] [CrossRef]
- Soehnlen, M.K.; Kunze, M.E.; Karunathilake, K.E.; Henwood, B.M.; Kariyawasam, S.; Wolfgang, D.R.; Jayarao, B.M. In vitro antimicrobial inhibition of mycoplasma bovis isolates submitted to the pennsylvania animal diagnostic laboratory using flow cytometry and a broth microdilution method. J. Vet. Diagn. Investig. 2011, 23, 547–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| Sample | Feedlot Production Year | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 2006 | 2007 | 2008 | 2014 | 2015 | 2016 | 2017 | 2018 | Total | |
| Anatomical Region | |||||||||
| DNP | 12 | 27 | 1 | 0 | 0 | 0 | 14 | 24 | 78 |
| Lung | 0 | 2 | 6 | 7 | 11 | 19 | 20 | 19 | 84 |
| Joint | 0 | 1 | 1 | 2 | 2 | 16 | 21 | 6 | 49 |
| Total | 12 | 30 | 8 | 9 | 13 | 35 | 55 | 49 | 211 |
| Health Status | |||||||||
| Healthy | 4 | 14 | 0 | 0 | 0 | 0 | 14 | 11 | 43 |
| Diseased | 8 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 19 |
| Dead | 0 | 5 | 8 | 9 | 13 | 35 | 41 | 38 | 149 |
| Total | 12 | 30 | 8 | 9 | 13 | 35 | 55 | 49 | 211 |
| Antibiotic | Class | ≤0.12 | 0.25 | 0.50 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | ≥256 | %R | MIC Range (µg/mL) | MIC50 (µg/mL) | MIC90 (µg/mL) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tulathromycin | Macrolide | 52 | 2 | 2 | 1 | 2 | 2 | 3 | 4 | 25 | 118 | 71.1 | ≤0.25–≥256 | ≥256 | ≥256 | ||
| Gamithromycin | Macrolide | 5 | 17 | 17 | 9 | 3 | 1 | 6 | 3 | 150 | 81.5 | 1–≥256 | ≥256 | ≥256 | |||
| Tilmicosin | Macrolide | 1 | 2 | 6 | 202 | 99.5 | 2–≥256 | ≥256 | ≥256 | ||||||||
| Tildipirosin | Macrolide | 1 | 8 | 13 | 20 | 169 | 100 | 8–≥128 | ≥128 | ≥128 | |||||||
| Tylosin | Macrolide | 3 | 10 | 13 | 22 | 6 | 12 | 11 | 134 | 87.7 | 1–≥128 | ≥128 | ≥128 | ||||
| Enrofloxacin | Fluoroquinolone | 120 | 15 | 20 | 30 | 19 | 7 | 36.0 | ≤0.12–16 | 0.25 | 4 | ||||||
| Florfenicol | Phenicol | 2 | 18 | 72 | 100 | 18 | 1 | 9.0 | ≤0.25–8 | 2 | 2 | ||||||
| Oxytetracycline | Tetracycline | 6 | 30 | 75 | 85 | 14 | 1 | 47.4 | 0.50–16 | 2 | 4 | ||||||
| Chlortetracycline | Tetracycline | 9 | 31 | 62 | 74 | 32 | 3 | 81.0 | ≤1–32 | 8 | 16 |
| Sample | TUL | GAM | TIL | TIP | TYL | ENRO | FFN | OXY | CTET |
|---|---|---|---|---|---|---|---|---|---|
| Healthy (N = 43) | |||||||||
| 2006–2007 (n = 18) | 24.39 | 21.31 | 21.17 | 18.61 | 21.83 | 16.92 | 19.64 | 22.67 | 22.86 |
| 2017–2018 (n = 25) | 20.28 | 22.50 | 22.60 | 24.44 | 22.12 | 25.66 | 23.70 | 21.52 | 21.38 |
| p = 0.225 | p = 0.753 | p = 0.403 | p = 0.091 | p = 0.939 | p = 0.004 | p = 0.206 | p = 0.745 | p = 0.676 | |
| Dead (N = 47) | |||||||||
| 2007–2008 (n = 8) | 7.31 | 6.94 | 24.00 | 9.38 | 4.56 | 10.50 | 31.88 | 32.50 | 29.13 |
| 2017–2018 (n = 39) | 27.42 | 27.50 | 24.00 | 27.00 | 27.99 | 26.77 | 22.38 | 22.26 | 22.95 |
| p < 0.001 | p < 0.001 | p = 1.000 | p < 0.001 | p < 0.001 | p = 0.001 | p = 0.060 | p = 0.044 | p = 0.231 | |
| Dead (N = 38) | |||||||||
| 2016 (n = 19) | 14.84 | 18.00 | 19.50 | 19.00 | 17.61 | 18.08 | 25.61 | 25.16 | 25.97 |
| 2018 (n = 19) | 24.16 | 21.00 | 19.50 | 20.00 | 21.39 | 20.92 | 13.39 | 13.84 | 13.03 |
| p = 0.002 | p = 0.075 | p = 1.000 | p = 0.317 | p = 0.097 | p = 0.413 | p < 0.001 | p = 0.001 | p < 0.001 |
| Antibiotic | Class | ≤0.12 | 0.25 | 0.50 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | ≥256 | %R | MIC Range (µg/mL) | MIC50 (µg/mL) | MIC90 (µg/mL) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tulathromycin | Macrolide | 26 | 1 | 1 | 2 | 1 | 1 | 11 | 30.2 | ≤0.25–≥256 | ≤0.25 | ≥256 | |||||
| Gamithromycin | Macrolide | 8 | 9 | 7 | 2 | 1 | 3 | 13 | 60.5 | 2–≥256 | 8 | ≥256 | |||||
| Tilmicosin | Macrolide | 1 | 2 | 40 | 100 | 64–≥256 | ≥256 | ≥256 | |||||||||
| Tildipirosin | Macrolide | 2 | 8 | 8 | 25 | 100 | 16–≥128 | ≥128 | ≥128 | ||||||||
| Tylosin | Macrolide | 3 | 7 | 15 | 4 | 4 | 10 | 76.7 | 2–≥128 | 8 | ≥128 | ||||||
| Enrofloxacin | Fluoroquinolone | 31 | 2 | 1 | 5 | 4 | 23.3 | ≤0.12–16 | ≤0.12 | 8 | |||||||
| Florfenicol | Phenicol | 1 | 8 | 29 | 5 | 11.6 | 0.5–4 | 2 | 4 | ||||||||
| Oxytetracycline | Tetracycline | 3 | 1 | 12 | 23 | 4 | 62.8 | ≤0.5–8 | 4 | 4 | |||||||
| Chlortetracycline | Tetracycline | 3 | 2 | 10 | 23 | 5 | 88.4 | ≤1–16 | 8 | 16 |
| Antibiotic | Class | ≤0.12 | 0.25 | 0.50 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | ≥256 | %R | MIC Range (µg/mL) | MIC50 (µg/mL) | MIC90 (µg/mL) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tulathromycin | Macrolide | 13 | 1 | 1 | 1 | 1 | 20 | 60 | 83.5 | ≤0.25–≥256 | ≥256 | ≥256 | |||||
| Gamithromycin | Macrolide | 1 | 4 | 5 | 1 | 1 | 2 | 1 | 82 | 89.7 | 1–≥256 | ≥256 | ≥256 | ||||
| Tilmicosin | Macrolide | 1 | 1 | 95 | 99.0 | 2–≥256 | ≥256 | ≥256 | |||||||||
| Tildipirosin | Macrolide | 2 | 2 | 7 | 86 | 100 | 16–≥128 | ≥128 | ≥128 | ||||||||
| Tylosin | Macrolide | 1 | 4 | 2 | 4 | 2 | 7 | 6 | 71 | 92.8 | ≤1–≥128 | ≥128 | ≥128 | ||||
| Enrofloxacin | Fluoroquinolone | 47 | 10 | 13 | 18 | 6 | 3 | 41.2 | ≤0.12–16 | 0.25 | 4 | ||||||
| Florfenicol | Phenicol | 2 | 10 | 33 | 44 | 7 | 1 | 8.2 | ≤0.25–8 | 2 | 2 | ||||||
| Oxytetracycline | Tetracycline | 3 | 13 | 37 | 36 | 7 | 1 | 45.4 | ≤0.5–≥16 | 2 | 4 | ||||||
| Chlortetracycline | Tetracycline | 4 | 15 | 27 | 34 | 15 | 2 | 80.4 | ≤1–32 | 8 | 16 |
| ENRO | TIP | GAM | TUL | FFN | OXY | CTET | TYL | TIL | |
|---|---|---|---|---|---|---|---|---|---|
| ENRO | 0.166 * | 0.262 ** | 0.295 ** | −0.357 ** | −0.170 * | −0.112 | 0.389 ** | 0.163 | |
| TIP | 0.166 * | 0.846 ** | 0.690 ** | −0.027 | −0.091 | −0.047 | 0.720 ** | 0.336 ** | |
| GAM | 0.262 ** | 0.846 ** | 0.874 ** | −0.183 * | −0.139 | −0.097 | 0.876 ** | 0.333 ** | |
| TUL | 0.295 ** | 0.690 ** | 0.874 ** | −0.255 ** | −0.187 * | −0.145 | 0.838 ** | 0.238 ** | |
| FFN | −0.357 ** | −0.027 | −0.183 * | −0.255 ** | 0.694 ** | 0.653 ** | −0.225 ** | 0.085 | |
| OXY | −0.170 * | −0.091 | −0.139 | −0.187 * | 0.694 ** | 0.895 ** | −0.102 | 0.215 * | |
| CTET | −0.112 | −0.047 | −0.097 | ‒0.145 | 0.653 ** | 0.895 ** | −0.018 | 0.171 * | |
| TYL | 0.389 ** | 0.720 ** | 0.876 ** | 0.838 ** | −0.225 ** | −0.102 | −0.018 | 0.335 ** | |
| TIL | 0.163 | 0.336 ** | 0.333 ** | 0.238 ** | 0.085 | 0.215 * | 0.171 * | 0.335 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jelinski, M.; Kinnear, A.; Gesy, K.; Andrés-Lasheras, S.; Zaheer, R.; Weese, S.; McAllister, T.A. Antimicrobial Sensitivity Testing of Mycoplasma bovis Isolates Derived from Western Canadian Feedlot Cattle. Microorganisms 2020, 8, 124. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010124
Jelinski M, Kinnear A, Gesy K, Andrés-Lasheras S, Zaheer R, Weese S, McAllister TA. Antimicrobial Sensitivity Testing of Mycoplasma bovis Isolates Derived from Western Canadian Feedlot Cattle. Microorganisms. 2020; 8(1):124. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010124
Chicago/Turabian StyleJelinski, Murray, Andrea Kinnear, Karen Gesy, Sara Andrés-Lasheras, Rahat Zaheer, Scott Weese, and Tim A. McAllister. 2020. "Antimicrobial Sensitivity Testing of Mycoplasma bovis Isolates Derived from Western Canadian Feedlot Cattle" Microorganisms 8, no. 1: 124. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010124