Evolutionarily-Related Helicobacter pylori Genotypes and Gastric Intraepithelial Neoplasia in a High-Risk Area of Northern Italy

 , and
, and 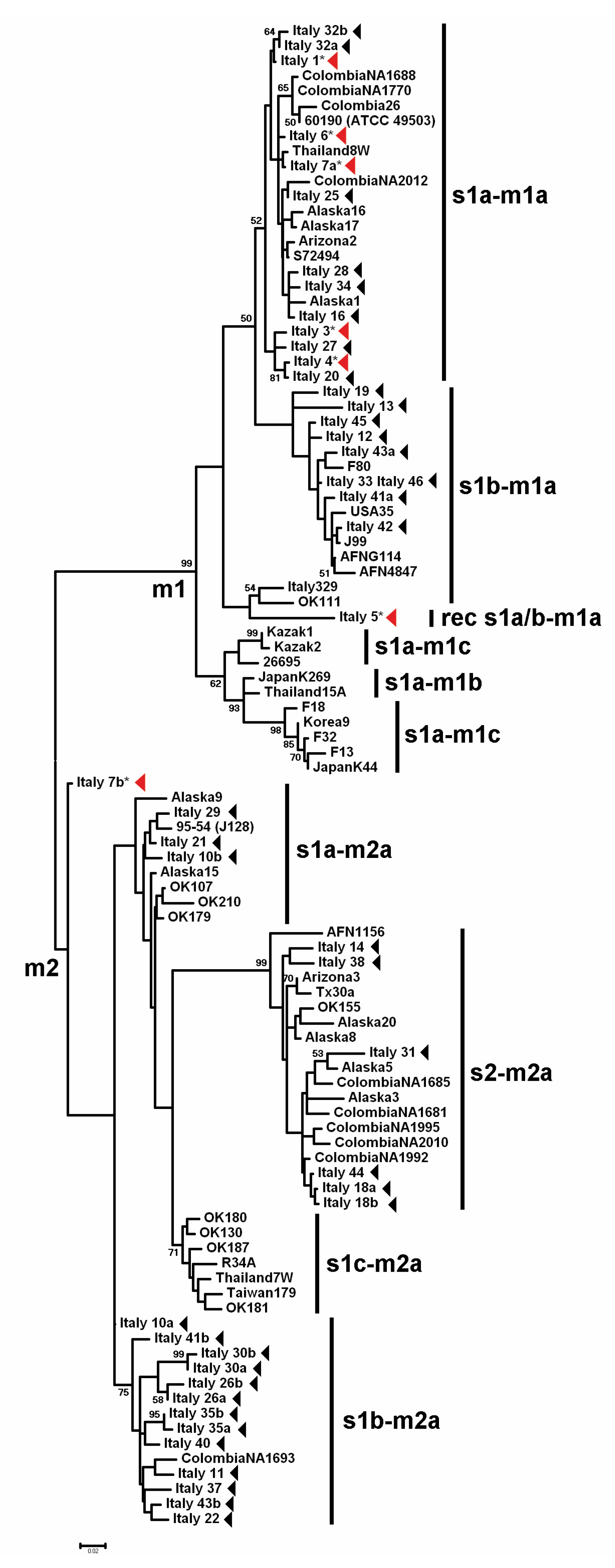{kind=link}
{kind=link}
{kind=link}
{kind=link}
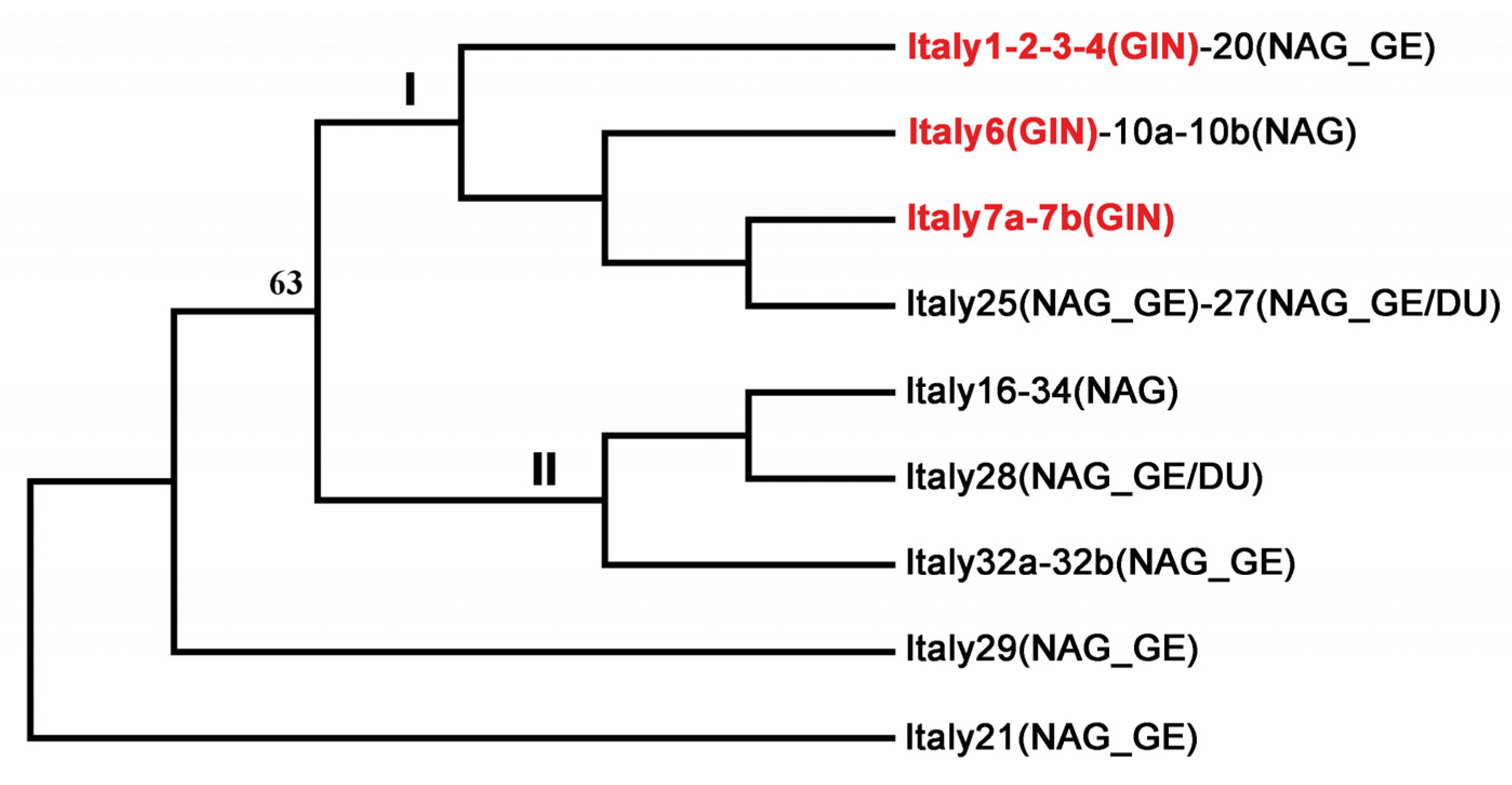{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cases and Histopathological Evaluation
2.2. DNA Extraction, Polymerase Chain Reaction (PCR) and Sequence Analyses
2.3. Data Analysis
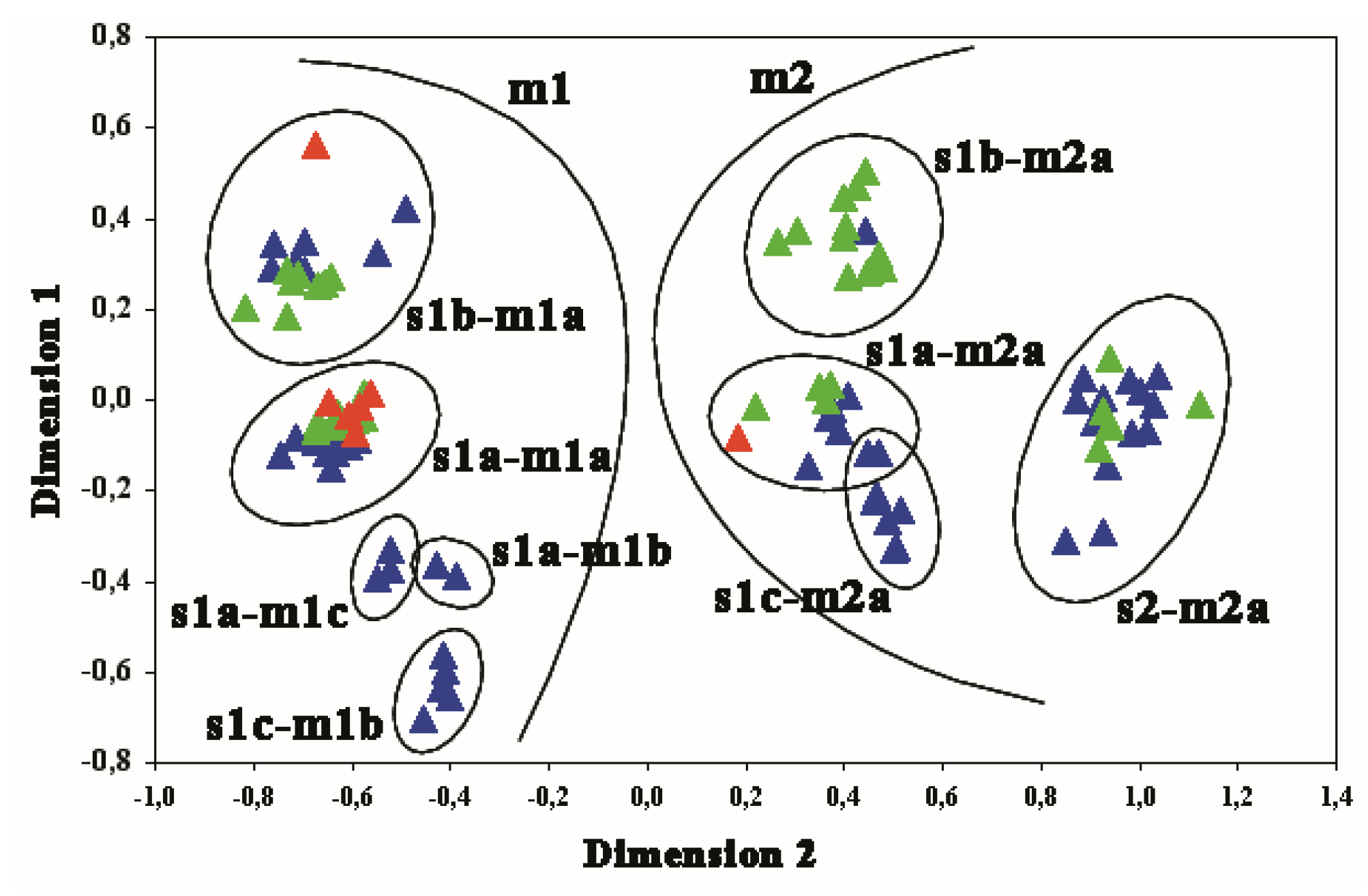
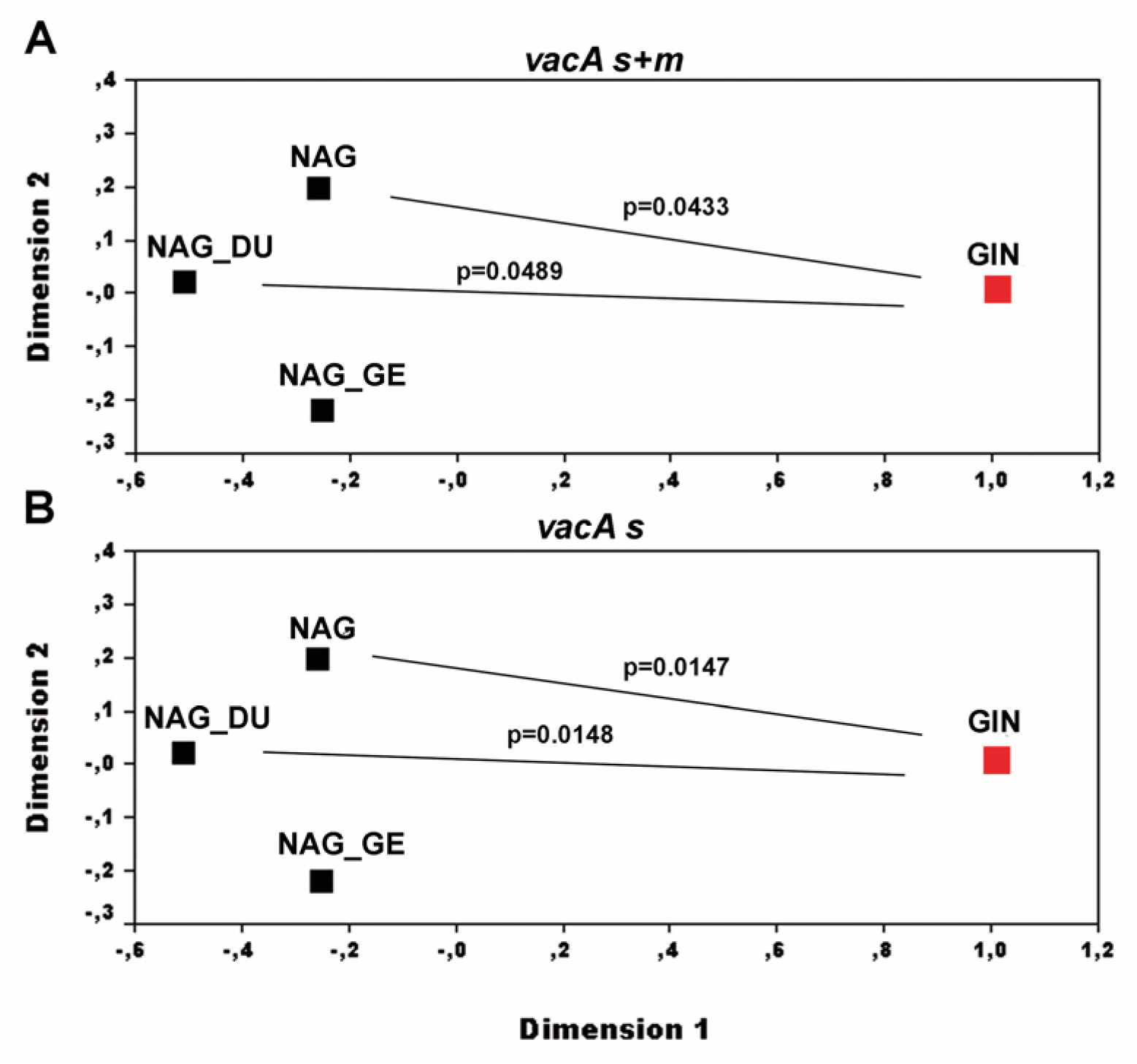
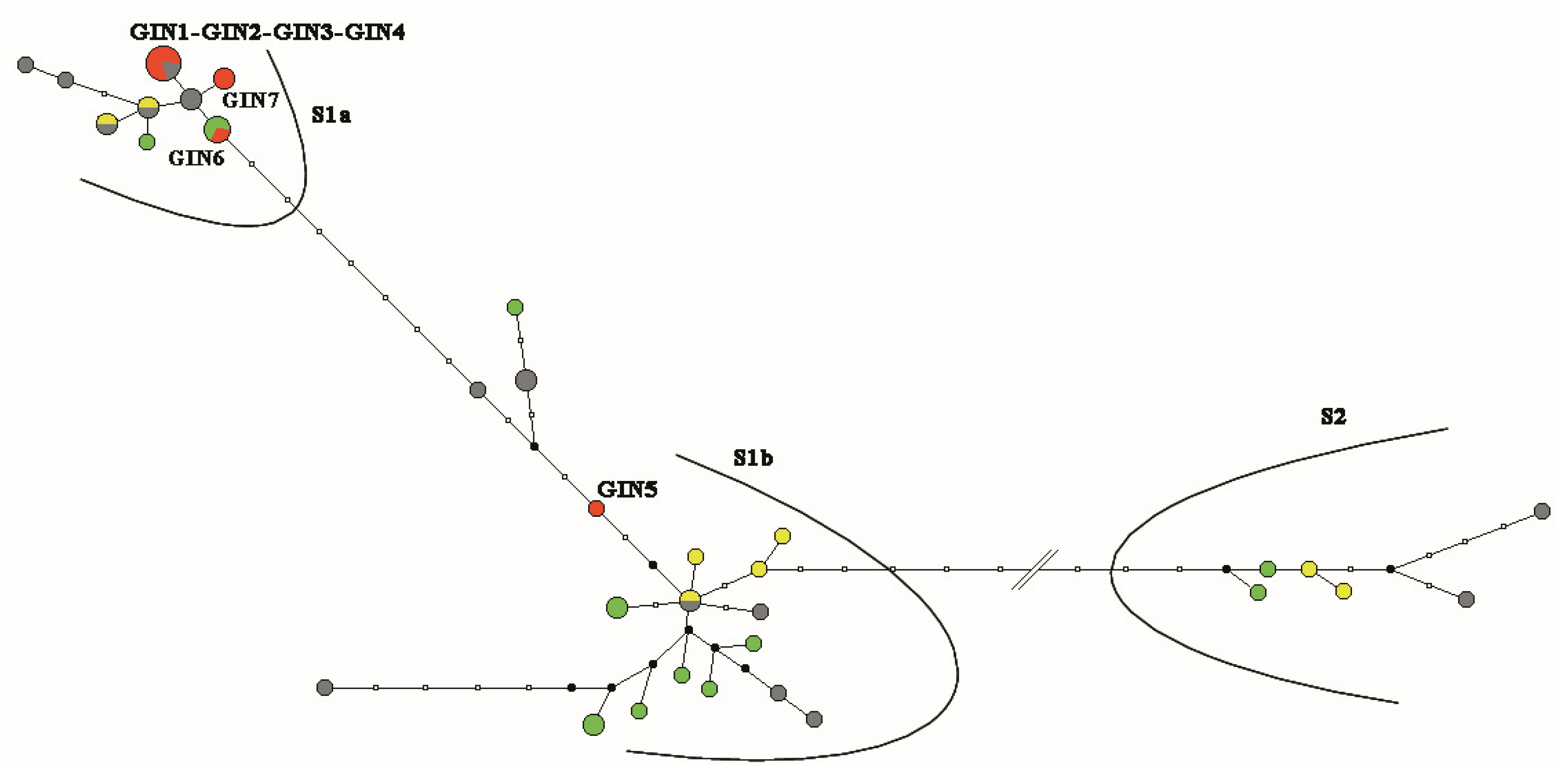
3. Results
3.1. GIN-Associated Hp Genotypes
3.2. NNGDL-Associated Hp Genotypes
3.3. Comparison between the Hp Genotypes of GINs and NNGDLs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Polk, D.B.; Peek, R.M., Jr. Helicobacter pylori: Gastric cancer and beyond. Nat. Rev. Cancer 2010, 10, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Parsonnet, J.; Friedman, G.D.; Vandersteen, D.P.; Chang, Y.; Vogelman, J.H.; Orentreich, N.; Sibley, R.K. Helicobacter pylori infection and the risk of gastric carcinoma. N. Engl. J. Med. 1991, 325, 1127–1131. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Biological Agents. A Review of Human Carcinogens; International Agency for Research on Cancer: Lyon, France, 2012; Volume 100, pp. 1–441. [Google Scholar]
- Lyons, K.; Le, L.C.; Pham, Y.T.; Borron, C.; Park, J.Y.; Tran, C.T.D.; Tran, T.V.; Tran, H.T.; Vu, K.T.; Do, C.D.; et al. Gastric cancer: Epidemiology, biology, and prevention: A mini review. Eur. J. Cancer Prev. Off. J. Eur. Cancer Prev. Organ. 2019, 28, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L. Helicobacter pylori diversity and gastric cancer risk. MBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Censini, S.; Lange, C.; Xiang, Z.; Crabtree, J.E.; Ghiara, P.; Borodovsky, M.; Rappuoli, R.; Covacci, A. Cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc. Natl. Acad. Sci. USA 1996, 93, 14648–14653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covacci, A.; Censini, S.; Bugnoli, M.; Petracca, R.; Burroni, D.; Macchia, G.; Massone, A.; Papini, E.; Xiang, Z.; Figura, N.; et al. Molecular characterization of the 128-kDa immunodominant antigen of Helicobacter pylori associated with cytotoxicity and duodenal ulcer. Proc. Natl. Acad. Sci. USA 1993, 90, 5791–5795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuck, D.; Kolmerer, B.; Iking-Konert, C.; Krammer, P.H.; Stremmel, W.; Rudi, J. Vacuolating cytotoxin of Helicobacter pylori induces apoptosis in the human gastric epithelial cell line AGS. Infect. Immun. 2001, 69, 5080–5087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galmiche, A.; Rassow, J.; Doye, A.; Cagnol, S.; Chambard, J.C.; Contamin, S.; de Thillot, V.; Just, I.; Ricci, V.; Solcia, E.; et al. The N-terminal 34 kDa fragment of Helicobacter pylori vacuolating cytotoxin targets mitochondria and induces cytochrome c release. EMBO J. 2000, 19, 6361–6370. [Google Scholar] [CrossRef] [PubMed]
- Atherton, J.C.; Cao, P.; Peek, R.M., Jr.; Tummuru, M.K.; Blaser, M.J.; Cover, T.L. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori. Association of specific vacA types with cytotoxin production and peptic ulceration. J. Biol. Chem. 1995, 270, 17771–17777. [Google Scholar] [CrossRef] [Green Version]
- Atherton, J.C.; Peek, R.M., Jr.; Tham, K.T.; Cover, T.L.; Blaser, M.J. Clinical and pathological importance of heterogeneity in vacA, the vacuolating cytotoxin gene of Helicobacter pylori. Gastroenterology 1997, 112, 92–99. [Google Scholar] [CrossRef]
- Palli, D.; Vaira, D.; Menegatti, M.; Saieva, C. A serologic survey of Helicobacter pylori infection in 3281 Italian patients endoscoped for upper gastrointestinal symptoms. The Italian Helicobacter Pylori Study Group. Aliment. Pharmacol. Ther. 1997, 11, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Gasbarrini, A.; Anti, M.; Franceschi, F.; Armuzzi, A.; Cotichini, R.; Ojetti, V.; Candelli, M.; Lippi, M.E.; Paolucci, M.; Cicconi, V.; et al. Prevalence of and risk factors for Helicobacter pylori infection among healthcare workers at a teaching hospital in Rome: The Catholic University Epidemiological Study. Eur. J. Gastroenterol. Hepatol. 2001, 13, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Zamani, M.; Ebrahimtabar, F.; Zamani, V.; Miller, W.H.; Alizadeh-Navaei, R.; Shokri-Shirvani, J.; Derakhshan, M.H. Systematic review with meta-analysis: The worldwide prevalence of Helicobacter pylori infection. Aliment. Pharmacol. Ther. 2018, 47, 868–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toracchio, S.; Marzio, L. Primary and secondary antibiotic resistance of Helicobacter pylori strains isolated in central Italy during the years 1998-2002. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2003, 35, 541–545. [Google Scholar] [CrossRef]
- Capocaccia, R.; De Angelis, R.; Frova, L.; Sant, M.; Buiatti, E.; Gatta, G.; Micheli, A.; Berrino, F.; Barchielli, A.; Conti, E.; et al. Estimation and projections of stomach cancer trends in Italy. Cancer Causes Control CCC 1995, 6, 339–346. [Google Scholar] [CrossRef]
- Ferlay, J.; Autier, P.; Boniol, M.; Heanue, M.; Colombet, M.; Boyle, P. Estimates of the cancer incidence and mortality in Europe in 2006. Ann. Oncol. Off. J. Eur. Soc. Med Oncol. 2007, 18, 581–592. [Google Scholar] [CrossRef]
- Forman, D.B.F.; Brewster, D.H.; Gombe Mbalawa, C.; Kohler, B.; Piñeros, M.; Steliarova-Foucher, E.; Swaminathan, R.; Ferlay, J. (Eds.) Cancer Incidence in Five Continents. In Cancer Incidence in Five Continents, 164th ed.; International Agency for Research on Cancer: Lyon, France, 2014; Volume X, pp. 714–779. [Google Scholar]
- Stroffolini, T.; Rosmini, F.; Ferrigno, L.; Fortini, M.; D’Amelio, R.; Matricardi, P.M. Prevalence of Helicobacter pylori infection in a cohort of Italian military students. Epidemiol. Infect. 1998, 120, 151–155. [Google Scholar] [CrossRef]
- Gallo, N.; Zambon, C.F.; Navaglia, F.; Basso, D.; Guariso, G.; Grazia Piva, M.; Greco, E.; Mazza, S.; Fogar, P.; Rugge, M.; et al. Helicobacter pylori infection in children and adults: A single pathogen but a different pathology. Helicobacter 2003, 8, 21–28. [Google Scholar] [CrossRef]
- Russo, F.; Berloco, P.; Cuomo, R.; Caruso, M.L.; Di Matteo, G.; Giorgio, P.; De Francesco, V.; Di Leo, A.; Ierardi, E. Helicobacter pylori strains and histologically-related lesions affect the outcome of triple eradication therapy: A study from southern Italy. Aliment. Pharmacol. Ther. 2003, 17, 421–428. [Google Scholar] [CrossRef]
- Zambon, C.F.; Navaglia, F.; Basso, D.; Rugge, M.; Plebani, M. Helicobacter pylori babA2, cagA, and s1 vacA genes work synergistically in causing intestinal metaplasia. J. Clin. Pathol. 2003, 56, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Figura, N.; Valassina, M.; Roviello, F.; Pinto, F.; Lenzi, C.; Giannace, R.; Marrelli, D.; Valentini, M.; Valensin, P.E. Helicobacter pylori cagA and vacA types and gastric carcinoma. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Study Liver 2000, 32, S182–S183. [Google Scholar] [CrossRef]
- Cellini, L.; Grande, R.; Di Campli, E.; Di Bartolomeo, S.; Capodicasa, S.; Marzio, L. Analysis of genetic variability, antimicrobial susceptibility and virulence markers in Helicobacter pylori identified in Central Italy. Scand. J. Gastroenterol. 2006, 41, 280–287. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, V.; Margiotta, M.; Zullo, A.; Hassan, C.; Valle, N.D.; Burattini, O.; D’Angelo, R.; Stoppino, G.; Cea, U.; Giorgio, F.; et al. Claritromycin resistance and Helicobacter pylori genotypes in Italy. J. Microbiol. 2006, 44, 660–664. [Google Scholar] [PubMed]
- Basso, D.; Zambon, C.F.; Letley, D.P.; Stranges, A.; Marchet, A.; Rhead, J.L.; Schiavon, S.; Guariso, G.; Ceroti, M.; Nitti, D.; et al. Clinical relevance of Helicobacter pylori cagA and vacA gene polymorphisms. Gastroenterology 2008, 135, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Grande, R.; Di Campli, E.; Di Bartolomeo, S.; Verginelli, F.; Di Giulio, M.; Baffoni, M.; Bessa, L.J.; Cellini, L. Helicobacter pylori biofilm: A protective environment for bacterial recombination. J. Appl. Microbiol. 2012, 113, 669–676. [Google Scholar] [CrossRef]
- Donida, B.M.; Tomasello, G.; Ghidini, M.; Buffoli, F.; Grassi, M.; Liguigli, W.; Maglietta, G.; Pergola, L.; Ratti, M.; Sabadini, G.; et al. Epidemiological, clinical and pathological characteristics of gastric neoplasms in the province of Cremona: The experience of the first population-based specialized gastric cancer registry in Italy. BMC Cancer 2019, 19, 212. [Google Scholar] [CrossRef] [Green Version]
- McColl, K.E.; el-Omar, E.; Gillen, D. Helicobacter pylori gastritis and gastric physiology. Gastroenterol. Clin. North Am. 2000, 29, 687–703. [Google Scholar] [CrossRef]
- Schlemper, R.J.; Riddell, R.H.; Kato, Y.; Borchard, F.; Cooper, H.S.; Dawsey, S.M.; Dixon, M.F.; Fenoglio-Preiser, C.M.; Flejou, J.F.; Geboes, K.; et al. The Vienna classification of gastrointestinal epithelial neoplasia. Gut 2000, 47, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Kato, M. Diagnosis and therapies for gastric non-invasive neoplasia. World J. Gastroenterol. 2015, 21, 12513–12518. [Google Scholar] [CrossRef]
- Dixon, M.F.; Genta, R.M.; Yardley, J.H.; Correa, P. Classification and grading of gastritis. The updated Sydney System. International Workshop on the Histopathology of Gastritis, Houston 1994. Am. J. Surg. Pathol. 1996, 20, 1161–1181. [Google Scholar] [CrossRef]
- Linee Guida. Tracciabilità, Raccolta, Trasporto, Conservazione e Archiviazione Di Cellule e Tessuti Per Indagini Diagnostiche Di Anatomia Patologica. Consiglio Superiore Di Sanità, Sezione I, Ministero Della Salute, Rome, Italy; 2015. Available online: http://www.salute.gov.it/imgs/C_17_pubblicazioni_2369_allegato.pdf (accessed on 6 January 2020).
- Rigoli, L.; Di Bella, C.; Verginelli, F.; Falchetti, M.; Bersiga, A.; Rocco, A.; Nardone, G.; Mariani-Costantini, R.; Caruso, R.A. Histological heterogeneity and somatic mtDNA mutations in gastric intraepithelial neoplasia. Mod. Pathol. Off. J. USA Can. Acad. Pathol. 2008, 21, 733–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocco, A.; Caruso, R.; Toracchio, S.; Rigoli, L.; Verginelli, F.; Catalano, T.; Neri, M.; Curia, M.C.; Ottini, L.; Agnese, V.; et al. Gastric adenomas: Relationship between clinicopathological findings, Helicobacter pylori infection, APC mutations and COX-2 expression. Ann. Oncol. Off. J. Eur. Soc. Med Oncol. 2006, 17, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. The genetical structure of populations. Ann. Eugen. 1951, 15, 323–354. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar]
- Polzin, T.D.; Daneshmand, S.V. On Steiner trees and minimum spanning trees in hypergraphs. Oper. Res. Lett. 2003, 12–20. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evol. Int. J. Org. Evol. 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, M. Helicobacter pylori CagA and gastric cancer: A paradigm for hit-and-run carcinogenesis. Cell Host Microbe 2014, 15, 306–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaoka, Y.; Orito, E.; Mizokami, M.; Gutierrez, O.; Saitou, N.; Kodama, T.; Osato, M.S.; Kim, J.G.; Ramirez, F.C.; Mahachai, V.; et al. Helicobacter pylori in North and South America before Columbus. FEBS Lett. 2002, 517, 180–184. [Google Scholar] [CrossRef] [Green Version]
- Covacci, A.; Telford, J.L.; Del Giudice, G.; Parsonnet, J.; Rappuoli, R. Helicobacter pylori virulence and genetic geography. Science 1999, 284, 1328–1333. [Google Scholar] [CrossRef] [Green Version]
- Kersulyte, D.; Mukhopadhyay, A.K.; Velapatino, B.; Su, W.; Pan, Z.; Garcia, C.; Hernandez, V.; Valdez, Y.; Mistry, R.S.; Gilman, R.H.; et al. Differences in genotypes of Helicobacter pylori from different human populations. J. Bacteriol. 2000, 182, 3210–3218. [Google Scholar] [CrossRef] [Green Version]
- Van Doorn, L.J.; Figueiredo, C.; Megraud, F.; Pena, S.; Midolo, P.; Queiroz, D.M.; Carneiro, F.; Vanderborght, B.; Pegado, M.D.; Sanna, R.; et al. Geographic distribution of vacA allelic types of Helicobacter pylori. Gastroenterology 1999, 116, 823–830. [Google Scholar] [CrossRef]
- Yamaoka, Y.; Kodama, T.; Gutierrez, O.; Kim, J.G.; Kashima, K.; Graham, D.Y. Relationship between Helicobacter pylori iceA, cagA, and vacA status and clinical outcome: Studies in four different countries. J. Clin. Microbiol. 1999, 37, 2274–2279. [Google Scholar] [CrossRef] [Green Version]
- Naumann, M.; Crabtree, J.E. Helicobacter pylori-induced epithelial cell signalling in gastric carcinogenesis. Trends Microbiol. 2004, 12, 29–36. [Google Scholar] [CrossRef]
- van Doorn, L.J.; Figueiredo, C.; Sanna, R.; Plaisier, A.; Schneeberger, P.; de Boer, W.; Quint, W. Clinical relevance of the cagA, vacA, and iceA status of Helicobacter pylori. Gastroenterology 1998, 115, 58–66. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toracchio, S.; Caruso, R.A.; Perconti, S.; Rigoli, L.; Betri, E.; Neri, M.; Verginelli, F.; Mariani-Costantini, R. Evolutionarily-Related Helicobacter pylori Genotypes and Gastric Intraepithelial Neoplasia in a High-Risk Area of Northern Italy. Microorganisms 2020, 8, 324. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030324
Toracchio S, Caruso RA, Perconti S, Rigoli L, Betri E, Neri M, Verginelli F, Mariani-Costantini R. Evolutionarily-Related Helicobacter pylori Genotypes and Gastric Intraepithelial Neoplasia in a High-Risk Area of Northern Italy. Microorganisms. 2020; 8(3):324. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030324
Chicago/Turabian StyleToracchio, Sonia, Rosario Alberto Caruso, Silvia Perconti, Luciana Rigoli, Enrico Betri, Matteo Neri, Fabio Verginelli, and Renato Mariani-Costantini. 2020. "Evolutionarily-Related Helicobacter pylori Genotypes and Gastric Intraepithelial Neoplasia in a High-Risk Area of Northern Italy" Microorganisms 8, no. 3: 324. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030324