First Report on the Prevalence and Subtype Distribution of Blastocystis sp. in Edible Marine Fish and Marine Mammals: A Large Scale-Study Conducted in Atlantic Northeast and on the Coasts of Northern France

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
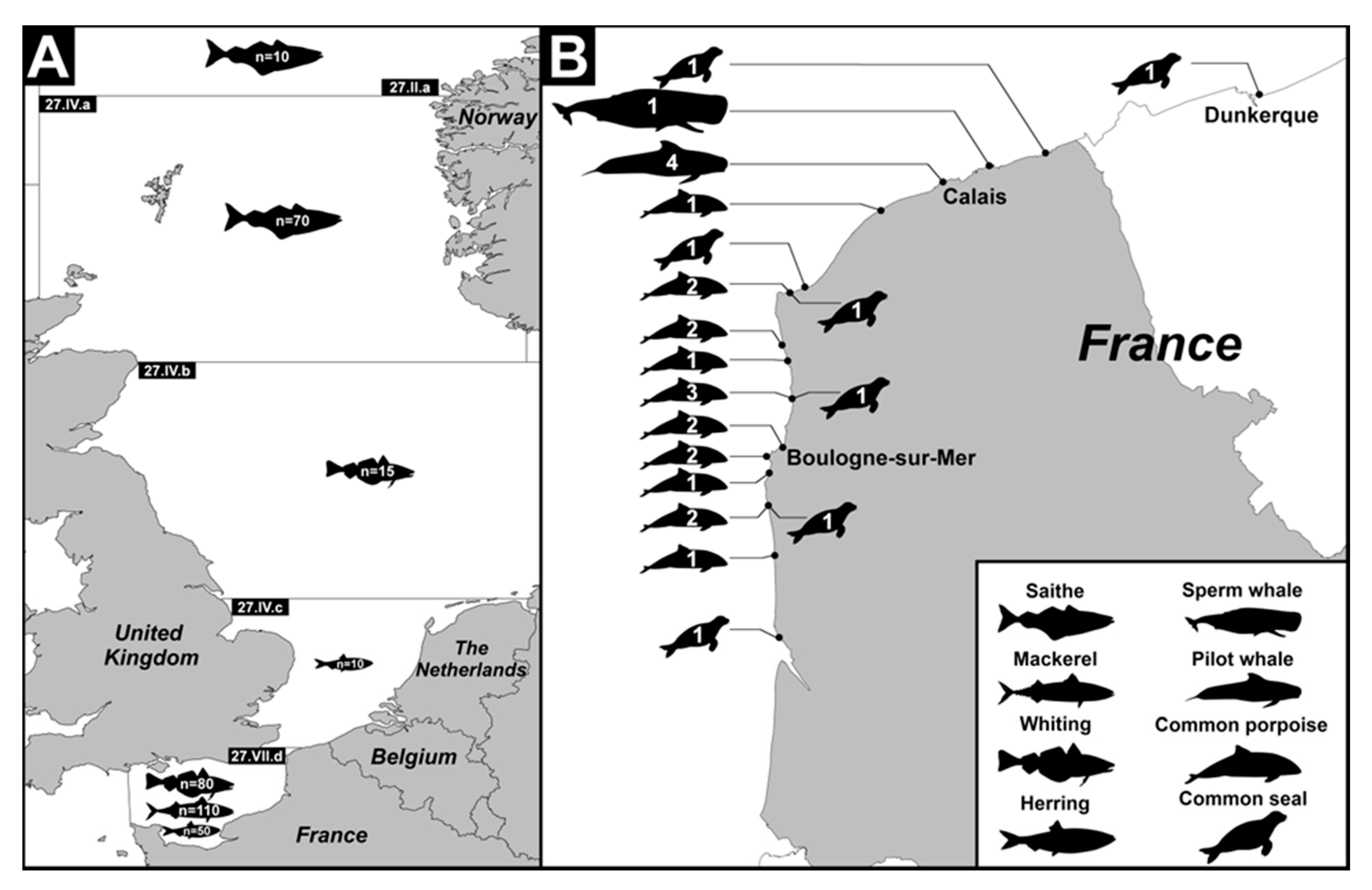
2.2. Fish Sampling
2.3. Marine Mammal Sampling
2.4. DNA Extraction
2.5. Amplification of the SSU rDNA Gene and Molecular Subtyping of Blastocystis sp. Isolates
2.6. Phylogenetic Analysis of Blastocystis sp. Isolates
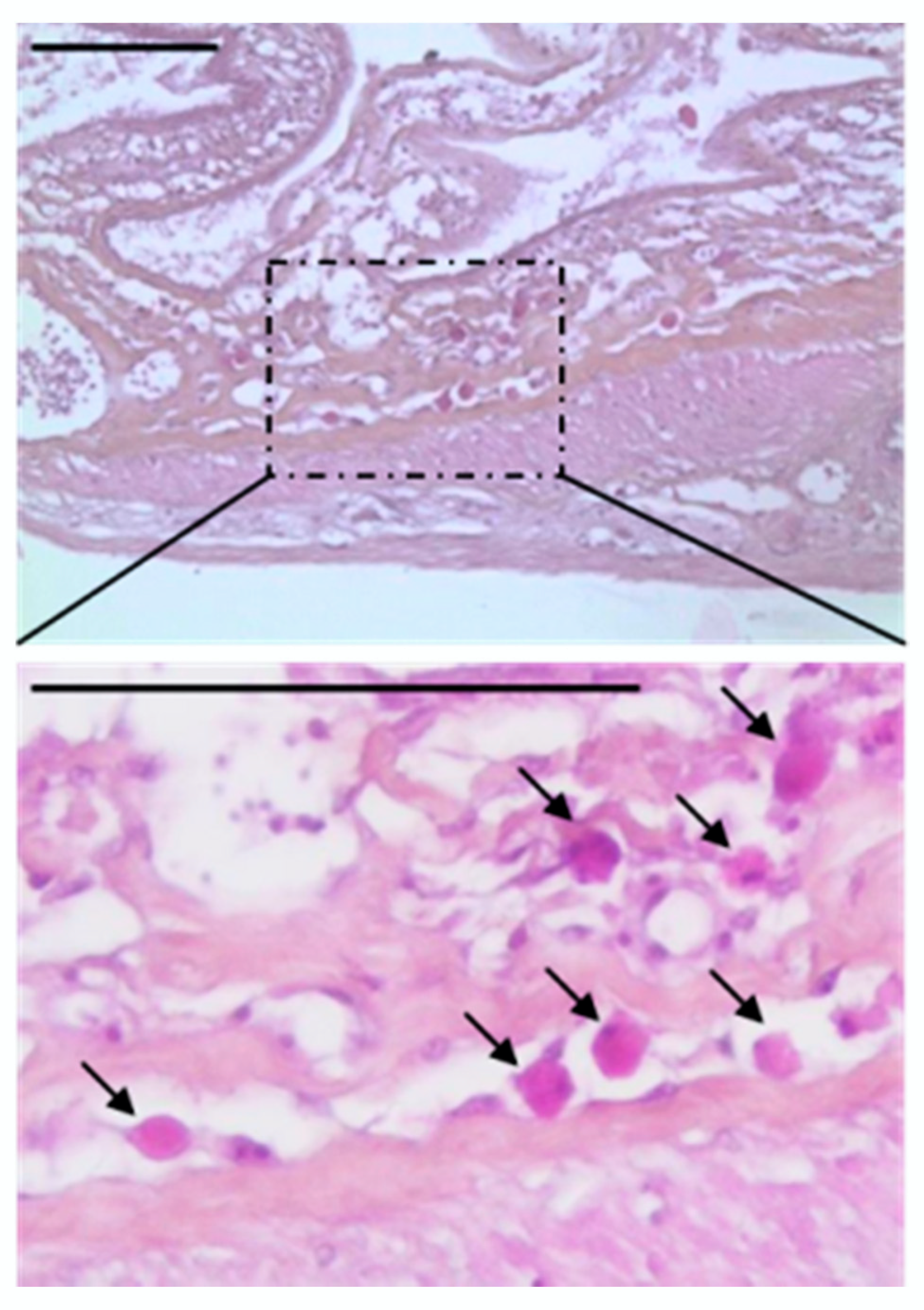
2.7. Histological Analysis
2.8. Statistical Analysis
3. Results and Discussion
3.1. Prevalence of Blastocystis sp. in Edible Marine Fish and Marine Mammal Samples
3.2. ST Distribution of Blastocystis sp. in Edible Marine Fish
3.3. ST Distribution of Blastocystis sp. in Marine Mammals
3.4. Colonisation and Circulation of Blastocystis sp. in Edible Marine Fish and Marine Mammals and Risk of Zoonotic Transmission
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boreham, P.F.L.; Stenzel, D.J. Blastocystis in humans and animals: Morphology, biology, and epizootiology. Adv. Parasitol. 1993, 32, 1–70. [Google Scholar]
- Tan, K.S.W. Blastocystis in humans and animals: New insights using modern methodologies. Vet. Parasitol. 2004, 126, 121–144. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Koyama, Y.; Tsuchiya, E.; Takami, K. Blastocystis phylogeny among various isolates from humans to insects. Parasitol. Int. 2016, 65, 750–759. [Google Scholar] [CrossRef] [PubMed]
- El Safadi, D.; Gaayeb, L.; Meloni, D.; Cian, A.; Poirier, P.; Wawrzyniak, I.; Delbac, F.; Dabboussi, F.; Delhaes, L.; Seck, M.; et al. Children of Senegal River Basin show the highest prevalence of Blastocystis sp. ever observed worldwide. BMC Infect. Dis. 2014, 14, 164. [Google Scholar] [CrossRef] [PubMed]
- Forsell, J.; Granlund, M.; Samuelsson, L.; Koskiniemi, S.; Edebro, H.; Evengard, B. High occurrence of Blastocystis sp. subtypes 1-3 and Giardia intestinalis assemblage B among patients in Zanzibar, Tanzania. Parasit. Vectors 2016, 9, 370. [Google Scholar] [CrossRef] [Green Version]
- Lokmer, A.; Cian, A.; Froment, A.; Gantois, N.; Viscogliosi, E.; Chabé, M.; Ségurel, L. Use of shotgun metagenomics for the identification of protozoa in the gut microbiota of healthy individuals from worldwide populations with various industrialization levels. PLoS ONE 2019, 14, e0211139. [Google Scholar] [CrossRef]
- Tan, K.S.W. New insights on classification, identification, and clinical relevance of Blastocystis spp. Clin. Microbiol. Rev. 2008, 21, 639–665. [Google Scholar] [CrossRef] [Green Version]
- Clark, C.G.; van der Giezen, M.; Alfellani, M.A.; Stensvold, C.R. Recent developments in Blastocystis research. Adv. Parasitol. 2013, 82, 1–32. [Google Scholar]
- Stensvold, C.R.; Clark, C.G. Current status of Blastocystis: A personal view. Parasitol. Int. 2016, 65, 763–771. [Google Scholar] [CrossRef]
- Ajjampur, S.S.; Tan, K.S.W. Pathogenic mechanisms in Blastocystis spp.—Interpreting results from in vitro and in vivo studies. Parasitol. Int. 2016, 65, 772–779. [Google Scholar] [CrossRef]
- Fréalle, E.; El Safadi, D.; Cian, A.; Aubry, E.; Certad, G.; Osman, M.; Wacrenier, A.; Dutoit, E.; Creusy, C.; Dubos, F.; et al. Acute Blastocystis-associated appendicular peritonitis in a child, Casablanca, Morocco. Emerg. Infect. Dis. 2015, 21, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Lepczynska, M.; Chen, W.C.; Dzika, E. Mysterious chronic urticarial caused by Blastocystis spp.? Int. J. Dermatol. 2015, 55, 259–266. [Google Scholar] [CrossRef]
- Noël, C.; Dufernez, F.; Gerbod, D.; Edgcomb, V.P.; Delgado-Viscogliosi, P.; Ho, L.C.; Singh, M.; Wintjens, R.; Sogin, M.L.; Capron, M.; et al. Molecular phylogenies of Blastocystis isolates from different hosts: Implications for genetic diversity, identification of species, and zoonosis. J. Clin. Microbiol. 2005, 43, 348–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stensvold, C.R.; Suresh, G.K.; Tan, K.S.; Thompson, R.C.; Traub, R.J.; Viscogliosi, E.; Yoshikawa, H.; Clark, C.G. Terminology for Blastocystis subtypes a consensus. Trends Parasitol. 2007, 23, 93–96. [Google Scholar] [CrossRef]
- Alfellani, M.A.; Taner-Mulla, D.; Jacob, A.S.; Imeede, C.A.; Yoshikawa, H.; Stensvold, C.R.; Clark, C.G. Genetic diversity of Blastocystis in livestock and zoo animals. Protist 2013, 164, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Cian, A.; El Safadi, D.; Osman, M.; Moriniere, R.; Gantois, N.; Benamrouz-Vanneste, S.; Delgado-Viscogliosi, P.; Guyot, K.; Li, L.L.; Monchy, S.; et al. Molecular epidemiology of Blastocystis sp. in various animal groups from two French zoos and evaluation of potential zoonotic risk. PLoS ONE 2017, 12, e0169659. [Google Scholar] [CrossRef] [Green Version]
- Alfellani, M.A.; Stensvold, C.R.; Vidal-Lapiedra, A.; Onuoha, E.S.; Fagbenro-Beyioku, A.F.; Clark, C.G. Variable geographic distribution of Blastocystis subtypes and its potential implications. Acta Trop. 2013, 126, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, J.D.; Sánchez, A.; Hernández, C.; Florez, C.; Bernal, M.C.; Giraldo, J.C.; Reyes, P.; Lopez, M.C.; Garcia, L.; Cooper, P.J.; et al. Geographic distribution of human Blastocystis subtypes in South America. Infect. Genet. Evol. 2016, 41, 32–35. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Alfellani, M.A.; Norskov-Lauritsen, S.; Prip, K.; Victory, E.L.; Maddox, C.; Nielsen, H.V.; Clark, C.G. Subtype distribution of Blastocystis isolates from synanthropic and zoo animals and identification of a new subtype. Int. J. Parasitol. 2009, 39, 473–479. [Google Scholar] [CrossRef] [Green Version]
- Parkar, U.; Traub, R.J.; Vitali, S.; Elliot, A.; Levecke, B.; Robertson, I.; Geurden, T.; Steele, J.; Drake, B.; Thompson, R.C.A. Molecular characterization of Blastocystis isolates from zoo animals and their animal-keepers. Vet. Parasitol. 2010, 169, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Owen, H.; Traub, R.J.; Cuttell, L.; Inpankaew, T.; Bielefeldt-Ohmann, H. Molecular epidemiology of Blastocystis in pigs and their in-contact humans in Southeast Queensland, Australia, and Cambodia. Vet. Parasitol. 2014, 203, 264–269. [Google Scholar] [CrossRef]
- Greige, S.; El Safadi, D.; Bécu, N.; Gantois, N.; Pereira, B.; Chabé, M.; Benamrouz-Vanneste, S.; Certad, G.; El Hage, R.; Chemaly, M.; et al. Prevalence and subtype distribution of Blastocystis sp. isolates from poultry in Lebanon and evidence of zoonotic potential. Parasit. Vectors 2018, 11, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, T.; Stark, D.; Harkness, J.; Ellis, J. Subtype distribution of Blastocystis isolates from a variety of animals from New South Wales, Australia. Vet. Parasitol. 2013, 196, 85–89. [Google Scholar] [CrossRef]
- Betts, E.L.; Gentekaki, E.; Thomasz, A.; Breakell, V.; Carpenter, A.I.; Tsaousis, A.D. Genetic diversity of Blastocystis in non-primate animals. Parasitology 2018, 145, 1228–1234. [Google Scholar] [CrossRef] [Green Version]
- Valença-Barbosa, C.; Bergamo do Bomfilm, T.C.; Rodrigues Teixeira, B.; Gentile, R.; Fraga da Costa Neto, S.; Souza Neil Magalhaes, B.; de Almeida Balthazar, D.; da Silva, F.A.; Biot, R.; Masini d’Avila Levy, C.; et al. Molecular epidemiology of Blastocystis isolated from animals in the state of Rio de Janeiro, Brazil. PLoS ONE 2019, 13, e0193860. [Google Scholar] [CrossRef] [Green Version]
- König, G.; Müller, H.E. Blastocystis hominis in animals: Incidence of four serogroups. Zent. Bakteriol. 1997, 286, 435–440. [Google Scholar] [CrossRef]
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular epidemiology of Anisakis and anisakiasis: An ecological and evolutionary road map. Adv. Parasitol. 2018, 99, 93–263. [Google Scholar] [PubMed]
- Certad, G.; Follet, J.; Gantois, N.; Hammouma-Ghelboun, O.; Guyot, K.; Benamrouz-Vanneste, S.; Fréalle, E.; Seesao, Y.; Delaire, B.; Creusy, C.; et al. Prevalence, molecular identification, and risk factors for Cryptosporidium infection in edible marine fish: A survey across sea areas surrounding France. Front. Microbiol. 2019, 10, 1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, P.; Wawrzyniak, I.; Albert, A.; El Alaoui, H.; Delbac, F.; Livrelli, V. Development and evaluation of a real-time PCR assay for detection and quantification of Blastocystis parasites in human stool samples: Prospective study of patients with hematological malignancies. J. Clin. Microbiol. 2011, 49, 975–983. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- El Safadi, D.; Cian, A.; Nourrisson, C.; Pereira, B.; Morelle, C.; Bastien, P.; Bellanger, A.P.; Botterel, F.; Candolfi, E.; Desoubeaux, G.; et al. Prevalence, risk factors for infection and subtype distribution of the intestinal parasite Blastocystis sp. from a large-scale multi-center study in France. BMC Infect. Dis. 2016, 16, 451. [Google Scholar] [CrossRef] [PubMed]
- Meloni, D.; Sanciu, G.; Poirier, P.; El Alaoui, H.; Chabe, M.; Delhaes, L.; Dei-Cas, E.; Delbac, F.; Fiori, P.L.; Di Cave, D.; et al. Molecular subtyping of Blastocystis sp. isolates from symptomatic patients in Italy. Parasitol. Res. 2011, 109, 613–619. [Google Scholar] [CrossRef]
- Segui, R.; Munoz-Antoli, C.; Klisiowicz, D.R.; Oishi, C.Y.; Koster, P.C.; De Lucio, A.; Hernandez-de-Mingo, M.; Puente, P.; Toledo, R.; Esteban, J.G.; et al. Prevalence of intestinal parasites, with emphasis on the molecular epidemiology of Giardia duodenalis and Blastocystis sp., in the Paranagua Bay, Brazil: A community survey. Parasit. Vectors 2018, 11, 490. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Lebbad, M.; Hansen, A.; Beser, J.; Belkessa, S.; Andersen, L.O.B.; Clark, C.G. Differentiation of Blastocystis and parasitic archamoebids encountered in untreated wastewater samples by amplicon-based next-generation sequencing. Parasite Epidemiol. Control. 2020, 9, e00131. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gong, B.; Liu, X.; Zhao, W.; Bu, T.; Zhang, W.; Liu, A.; Yanf, F. Distribution and genetic diversity of Blastocystis subtypes in various mammal and bird species in northeastern China. Parasit. Vectors 2018, 11, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, A.; Sumiyoshi, T.; Ohtaki, T.; Matsumoto, J. Prevalence and molecular subtyping of Blastocystis from dairy cattle in Kanagawa, Japan. Parasitol. Int. 2018, 67, 702–705. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.-H.; Seo, M.-G.; Kim, H.-Y.; Kim, J.W.; Lee, Y.-R.; Kim, J.H.; Kwon, O.-D.; Kwak, D. Occurrence and genetic diversity of Blastocystis in Korean cattle. Vet. Parasitol. 2018, 258, 70–73. [Google Scholar] [CrossRef]
- Anders, N.; Fernö, A.; Humborstad, O.-B.; Lokkebord, S.; Rieucau, G.; Utne-Palm, A.C. Size-dependent social attraction and repulsion explains the decision of Atlantic cod Gadus morhua to enter baited pots. J. Fish Biol. 2017, 91, 1569–1581. [Google Scholar] [CrossRef]
- Teow, W.L.; Ng, G.C.; Chan, P.P.; Chan, Y.C.; Yap, E.H.; Zaman, V.; Singh, M. A survey of Blastocystis in reptiles. Parasitol. Res. 1992, 78, 453–455. [Google Scholar] [CrossRef]
- Singh, M.; Ho, L.C.; Yap, A.L.L.; Ng, G.C.; Tan, S.W.; Moe, K.T.; Yap, E.H. Axenic culture of reptilian Blastocystis isolates in monophasic medium and speciation by karyotipic typing. Parasitol. Res. 1996, 82, 165–169. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Morimoto, K.; Nagashima, M.; Miyamoto, N. A survey of Blastocystis infection in anuran and urodele amphibians. Vet. Parasitol. 2004, 122, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Mladineo, I.; Poljak, V. Ecology and genetic structure of zoonotic Anisakis spp. from Adriatic commercial fish species. Appl. Environ. Microbiol. 2014, 80, 1281–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Cordon, G.; Rosales, M.J.; Gavira, M.M.; Valdez, R.A.; Vargas, F.; Cordova, O. Finding of Blastocystis sp. in bivalves of the genus Donax. Rev. Peru Biol. 2007, 14, 301–302. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Batch of Fish | Scientific Name | Common Name | Fishing Date | Fishing Area a | Number of Individuals Per Batch (Number of Fishes Analysed Per Batch) | Number of Blastocystis sp.-Positive Fishes |
|---|---|---|---|---|---|---|
| CH-I | Clupea harengus | Herring | 14/01/2014 | 27.VII.d Eastern English Channel | 32 (10) | 0 |
| CH-II | Clupea harengus | Herring | 02/12/2014 | 27.VII.d Eastern English Channel | 30 (10) | 4 |
| CH-III | Clupea harengus | Herring | 14/02/2015 | 27.VII.d Eastern English Channel | 30 (10) | 0 |
| CH-IV | Clupea harengus | Herring | 05/03/2015 | 27.VII.d Eastern English Channel | 30 (10) | 1 |
| CH-V | Clupea harengus | Herring | 08/12/2015 | 27.VII.d Eastern English Channel | 30 (10) | 0 |
| CH-VI | Clupea harengus | Herring | 08/12/2015 | 27.IV.c Southern North Sea | 30 (10) | 0 |
| MM-I | Merlangius merlangus | Whiting | 08/08/2014 | 27.IV.b Central North Sea | 30 (15) | 0 |
| MM-II | Merlangius merlangus | Whiting | 01/10/2014 | 27.VII.d Eastern English Channel | 30 (20) | 0 |
| MM-III | Merlangius merlangus | Whiting | 01/10/2014 | 27.VII.d Eastern English Channel | 30 (20) | 1 |
| MM-IV | Merlangius merlangus | Whiting | 27/01/2015 | 27.VII.d Eastern English Channel | 30 (10) | 1 |
| MM-V | Merlangius merlangus | Whiting | 01/04/2015 | 27.VII.d Eastern English Channel | 30 (10) | 0 |
| MM-VI | Merlangius merlangus | Whiting | 07/09/2015 | 27.VII.d Eastern English Channel | 30 (10) | 0 |
| MM-VII | Merlangius merlangus | Whiting | 07/09/2015 | 27.VII.d Eastern English Channel | 30 (10) | 0 |
| PV-I | Pollachius virens | Saithe | 16/05/2014 | 27.II.a Norwegian Sea | 22 (10) | 0 |
| PV-II | Pollachius virens | Saithe | 27/07/2014 | 27.IV.a Northern North Sea | 30 (30) | 0 |
| PV-III | Pollachius virens | Saithe | 02/10/2014 | 27.IV.a Northern North Sea | 30 (10) | 0 |
| PV-IV | Pollachius virens | Saithe | 27/11/2014 | 27.IV.a Northern North Sea | 30 (10) | 1 |
| PV-V | Pollachius virens | Saithe | 26/03/2015 | 27.IV.a Northern North Sea | 30 (10) | 1 |
| PV-VI | Pollachius virens | Saithe | 20/07/2015 | 27.IV.a Northern North Sea | 30 (10) | 0 |
| SS-I | Scomber scombrus | Mackerel | 07/08/2014 | 27.VII.d Eastern English Channel | 30 (30) | 0 |
| SS-II | Scomber scombrus | Mackerel | 17/09/2014 | 27.VII.d Eastern English Channel | 20 (20) | 2 |
| SS-III | Scomber scombrus | Mackerel | 01/10/2014 | 27.VII.d Eastern English Channel | 30 (20) | 1 |
| SS-IV | Scomber scombrus | Mackerel | 13/05/2015 | 27.VII.d Eastern English Channel | 30 (10) | 0 |
| SS-V | Scomber scombrus | Mackerel | 08/07/2015 | 27.VII.d Eastern English Channel | 30 (10) | 0 |
| SS-VI | Scomber scombrus | Mackerel | 07/09/2015 | 27.VII.d Eastern English Channel | 30 (10) | 0 |
| SS-VII | Scomber scombrus | Mackerel | 29/09/2015 | 27.VII.d Eastern English Channel | 30 (10) | 0 |
| Individual | Scientific Name | Common Name | Date of Stranding | Place of Stranding (French Department) a | Sex | Condition of Collected Sample b | Sequence Positive Samples for Blastocystis sp. |
|---|---|---|---|---|---|---|---|
| CET-01 | Phocoena phocoena | Common porpoise | 25/01/2014 | Saint-Etienne-au-Mont (62) | M | Fresh | 0 |
| CET-02 | Phocoena phocoena | Common porpoise | 06/03/2014 | Boulogne-sur-Mer (62) | F | Putrefied | 0 |
| CET-03 | Phocoena phocoena | Common porpoise | 18/03/2014 | Hardelot (62) | M | Putrefied | 0 |
| CET-04 | Phocoena phocoena | Common porpoise | 20/03/2014 | Wimereux (62) | F | Fresh | 0 |
| CET-05 | Phocoena phocoena | Common porpoise | 27/03/2014 | Boulogne-sur-Mer (62) | F | Putrefied | 0 |
| CET-06 | Phocoena phocoena | Common porpoise | 27/03/2014 | Saint-Etienne-au-Mont (62) | M | Fresh | 0 |
| CET-07 | Phocoena phocoena | Common porpoise | 10/04/2014 | Wimereux (62) | M | Fresh | 0 |
| CET-08 | Phocoena phocoena | Common porpoise | 21/08/2014 | Ambleteuse (62) | M | Fresh | 1 |
| CET-09 | Phocoena phocoena | Common porpoise | 01/09/2014 | Le Portel (62) | F | Very fresh | 3 |
| CET-10 | Phocoena phocoena | Common porpoise | 08/04/2014 | Audresselles (62) | F | Fresh | 0 |
| CET-11 | Globicephala melas | Long-finned pilot whale | 02/11/2015 | Calais (62) | M | Very fresh | 0 |
| CET-12 | Globicephala melas | Long-finned pilot whale | 02/11/2015 | Calais (62) | F | Very fresh | 0 |
| CET-13 | Globicephala melas | Long-finned pilot whale | 02/11/2015 | Calais (62) | M | Very fresh | 0 |
| CET-14 | Globicephala melas | Long-finned pilot whale | 02/11/2015 | Calais (62) | M | Very fresh | 0 |
| CET-15 | Physeter macrocephalus | Sperm whale | 02/02/2016 | Marck-en-Calaisis (62) | M | Putrefied | 1 |
| CET-16 | Phocoena phocoena | Common porpoise | 18/02/2016 | Sangatte (62) | F | Fresh | 0 |
| CET-17 | Phocoena phocoena | Common porpoise | 29/02/2016 | Audinghen (62) | M | Putrefied | 0 |
| CET-18 | Phocoena phocoena | Common porpoise | 28/02/2016 | Audinghen (62) | M | Putrefied | 0 |
| CET-19 | Phocoena phocoena | Common porpoise | 29/02/2016 | Audresselles (62) | F | Fresh | 0 |
| CET-20 | Phocoena phocoena | Common porpoise | 20/03/2016 | Equihen-Plage (62) | F | Fresh | 0 |
| CET-21 | Phocoena phocoena | Common porpoise | 26/05/2016 | Wimereux (62) | M | Putrefied | 0 |
| CET-22 | Phocoena phocoena | Common porpoise | 26/05/2016 | Le Portel (62) | F | Putrefied | 0 |
| PIN-01 | Phoca vitulina | Common seal | 25/01/2014 | Audinghen (62) | F | Fresh | 0 |
| PIN-02 | Phoca vitulina | Common seal | 06/03/2014 | Tardinghen (62) | M | Fresh | 0 |
| PIN-03 | Phoca vitulina | Common seal | 22/12/2015 | Camiers (62) | M | Fresh | 0 |
| PIN-04 | Phoca vitulina | Common seal | 18/01/2016 | Wimereux (62) | M | Fresh | 0 |
| PIN-05 | Phoca vitulina | Common seal | 17/01/2016 | Dunkerque (59) | F | Very fresh | 0 |
| PIN-06 | Phoca vitulina | Common seal | 11/03/2016 | Saint-Etienne-au-Mont (62) | F | Fresh | 2 |
| PIN-07 | Phoca vitulina | Common seal | 18/03/2016 | Oye-Plage (62) | M | Putrefied | 0 |
| Batch and Individual | Fishing Area | Analysed Sample a | Species | Blastocystis sp. ST best hit by BLAST | Sequence Identity with Best Hit b |
|---|---|---|---|---|---|
| CH-II-1 | Eastern English Channel | IC | Herring | ST8 | 100% |
| CH-II-1 | SIE | Herring | ST10 | 92% c | |
| (Untypable isolate ZLC7) d | (100%) | ||||
| CH-II-1 | SGE | Herring | ST8 | 100% | |
| CH-II-2 | Eastern English Channel | IC | Herring | ST8 | 100% |
| CH-II-7 | Eastern | IC | Herring | ST10 | 92% c |
| English Channel | (Untypable isolate ZLC7) d | (100%) | |||
| CH-II-7 | SIE | Herring | ST10 | 92% c | |
| (Untypable isolate ZLC7) d | (100%) | ||||
| CH-II-7 | SGE | Herring | ST8 | 100% | |
| CH-II-8 | Eastern | IC | Herring | ST10 | 92% c |
| English Channel | (Untypable isolate ZLC7) d | (100%) | |||
| CH-II-8 | SGE | Herring | ST7 | 98% | |
| CH-IV-7 | Eastern | IC | Herring | NMAST II e | 91% c |
| English Channel | (U-typable isolate ZLB27) d | (100%) | |||
| MM-III-3 | Eastern English Channel | IC | Whiting | ST2 | 100% |
| MM-IV-9 | Eastern | IC | Whiting | NMAST VII e | 85% c |
| English Channel | (Untypable isolate LPO12) d | (100%) | |||
| PV-IV-3 | Northern | IC | Saithe | ST10 | 92% c |
| North Sea | (Untypable isolate ZLC7) d | (100%) | |||
| PV-IV-3 | SGE | Saithe | ST8 | 100% | |
| PV-V-9 | Northern | IC | Saithe | ST10 | 92% c |
| North Sea | (Untypable isolate ZLC7) d | (100%) | |||
| SS-II-17 | Eastern | IC | Mackerel | ST8 | 91% c |
| English Channel | (Untypable isolate LPA3) d | (100%) | |||
| SS-II-46 | Eastern English Channel | IC | Mackerel | ST2 | 100% |
| SS-III-1 | Eastern | IC | Mackerel | ST10 | 92% c |
| English Channel | (Untypable isolate ZLC7) d | (100%) |
| Individual | Analysed Sample a | Species | Cloning | Blastocystis sp. ST Best Hit by BLAST | Sequence Identity with Best Hit b |
|---|---|---|---|---|---|
| CET-08 | SIE | Common porpoise | N.d. f | ST8 | 91% c |
| (Untypable isolate LPA3) d | (100%) | ||||
| CET-09 | SIE | Common porpoise | Clone 1 | ST4 | 99% |
| Clone 2 | NMAST VII e | 85% c | |||
| (Untypable isolate LPO12) d | (100%) | ||||
| Clone 3 | NMAST II e | 82% c | |||
| (Untypable isolate LPA10) | (99%) | ||||
| CET-09 | SGE-FU | Common porpoise | N.d. | ST2 | 99% |
| CET-09 | SGE-FO | Common porpoise | N.d. | ST3 | 99% |
| CET-15 | SIE | Sperm whale | N.d. | ST2 | 100% |
| PIN-06 | IC | Common seal | N.d. | ST4 | 100% |
| PIN-06 | SIE | Common seal | N.d. | ST4 | 100% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gantois, N.; Lamot, A.; Seesao, Y.; Creusy, C.; Li, L.-L.; Monchy, S.; Benamrouz-Vanneste, S.; Karpouzopoulos, J.; Bourgain, J.-L.; Rault, C.; et al. First Report on the Prevalence and Subtype Distribution of Blastocystis sp. in Edible Marine Fish and Marine Mammals: A Large Scale-Study Conducted in Atlantic Northeast and on the Coasts of Northern France. Microorganisms 2020, 8, 460. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030460
Gantois N, Lamot A, Seesao Y, Creusy C, Li L-L, Monchy S, Benamrouz-Vanneste S, Karpouzopoulos J, Bourgain J-L, Rault C, et al. First Report on the Prevalence and Subtype Distribution of Blastocystis sp. in Edible Marine Fish and Marine Mammals: A Large Scale-Study Conducted in Atlantic Northeast and on the Coasts of Northern France. Microorganisms. 2020; 8(3):460. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030460
Chicago/Turabian StyleGantois, Nausicaa, Angélique Lamot, Yuwalee Seesao, Colette Creusy, Luen-Luen Li, Sébastien Monchy, Sadia Benamrouz-Vanneste, Jacky Karpouzopoulos, Jean-Luc Bourgain, Célia Rault, and et al. 2020. "First Report on the Prevalence and Subtype Distribution of Blastocystis sp. in Edible Marine Fish and Marine Mammals: A Large Scale-Study Conducted in Atlantic Northeast and on the Coasts of Northern France" Microorganisms 8, no. 3: 460. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030460