Biochar and Rhizobacteria Amendments Improve Several Soil Properties and Bacterial Diversity

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Biochar and PGPR Characterization
2.3. Experimental Design and Soil Sampling
2.4. Analysis of Soil Physicochemical Properties and Bacterial Community
2.5. Data Analysis
3. Results
3.1. Effects of Biochar and PGPR on Soil Physicochemical Properties
3.2. Effects of Biochar and PGPR on Microbial Richness and Diversity Indices
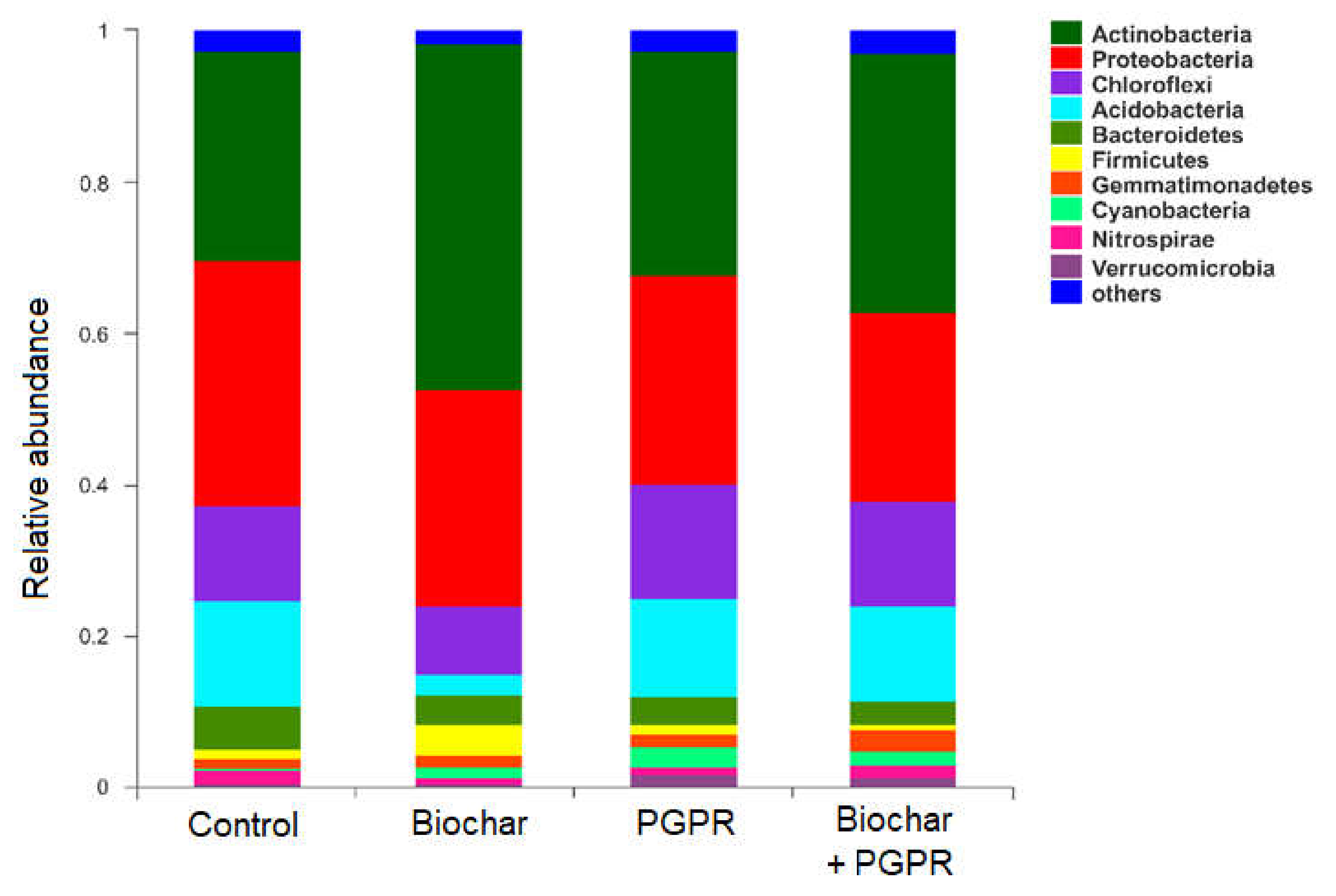
3.3. Effects of Biochar and PGPR on Soil Bacterial Community Composition (Phylum Level)
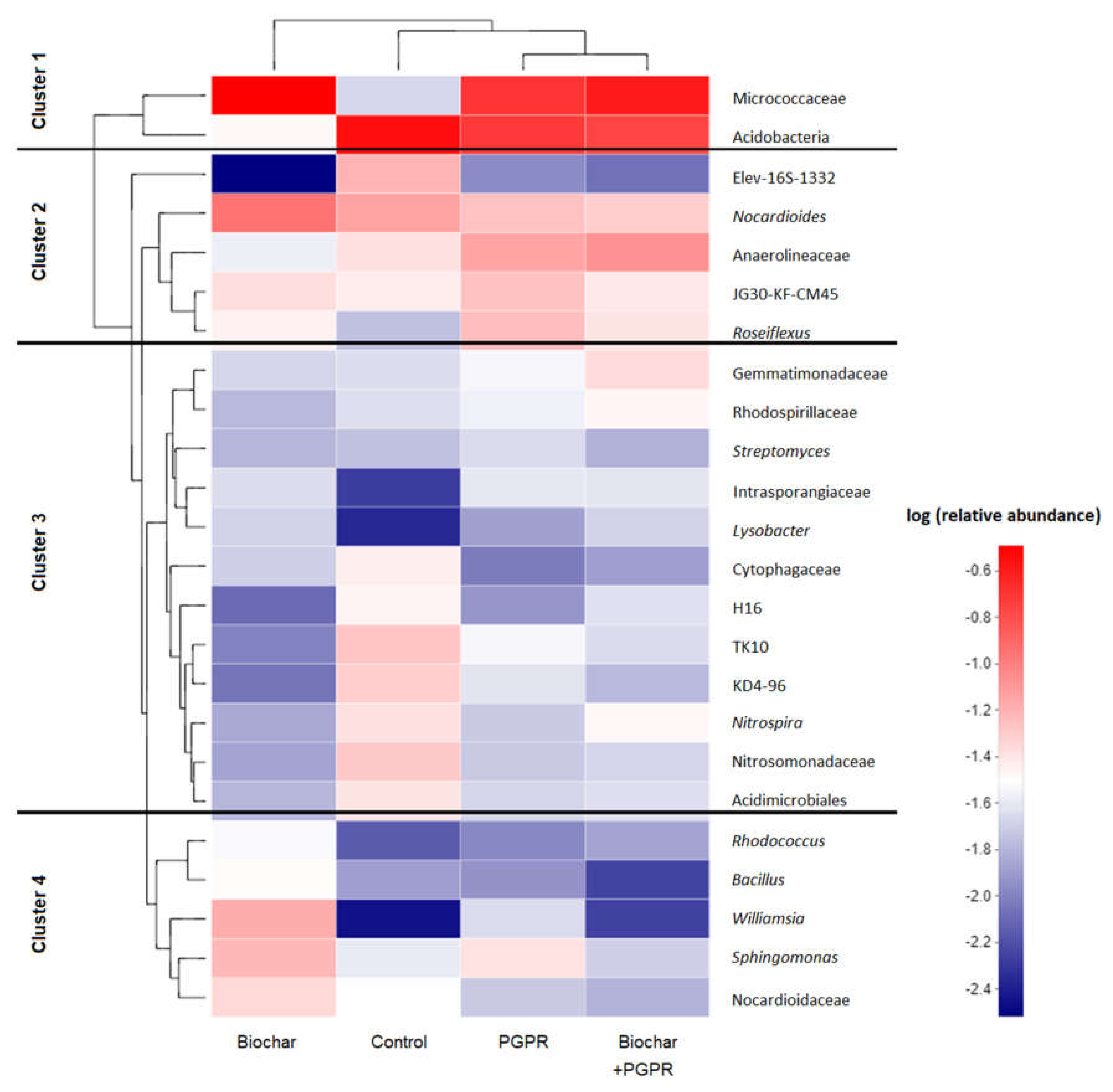
3.4. Effects of Biochar and PGPR on Soil Bacterial Community Composition (Genus Level)
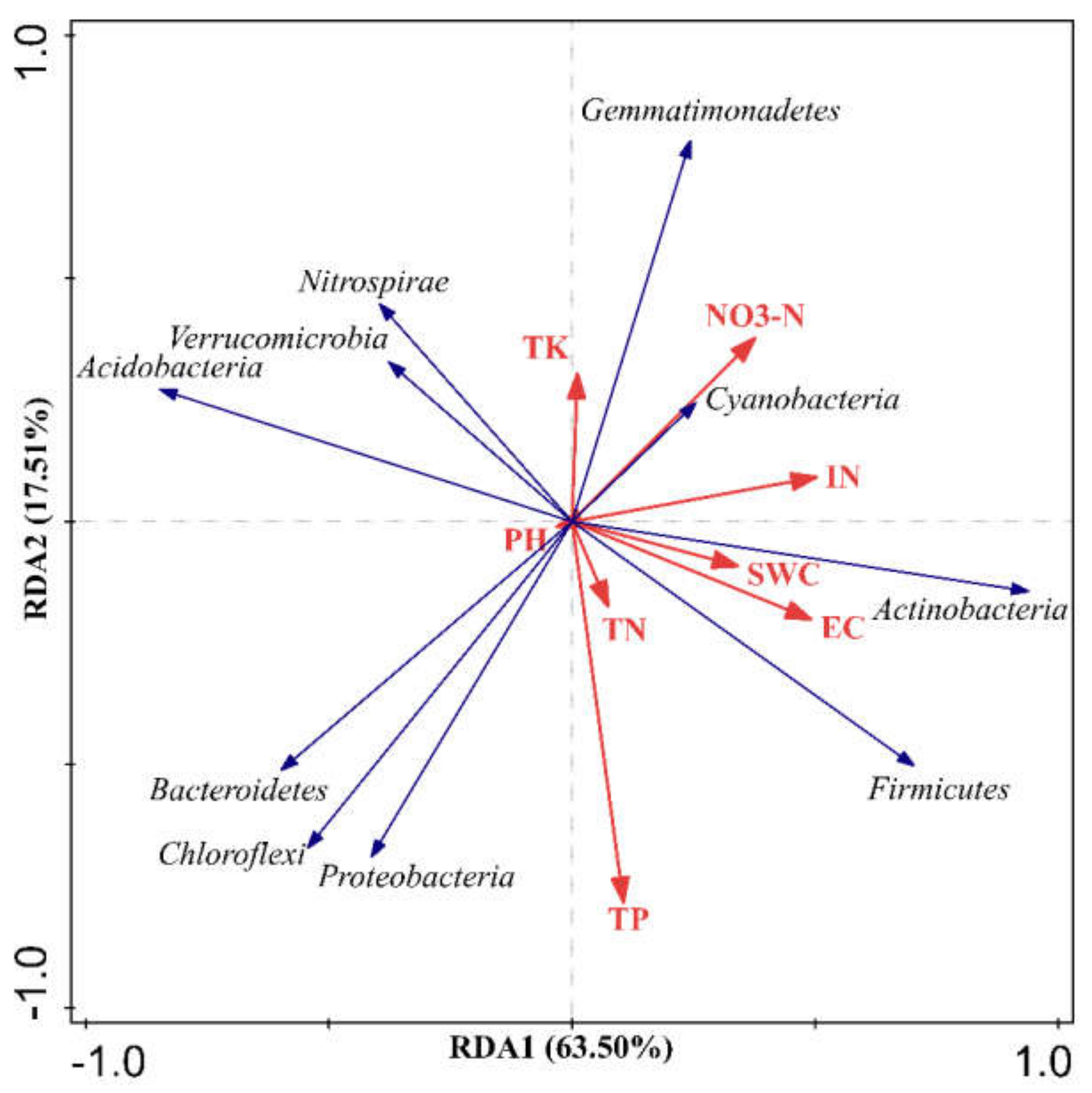
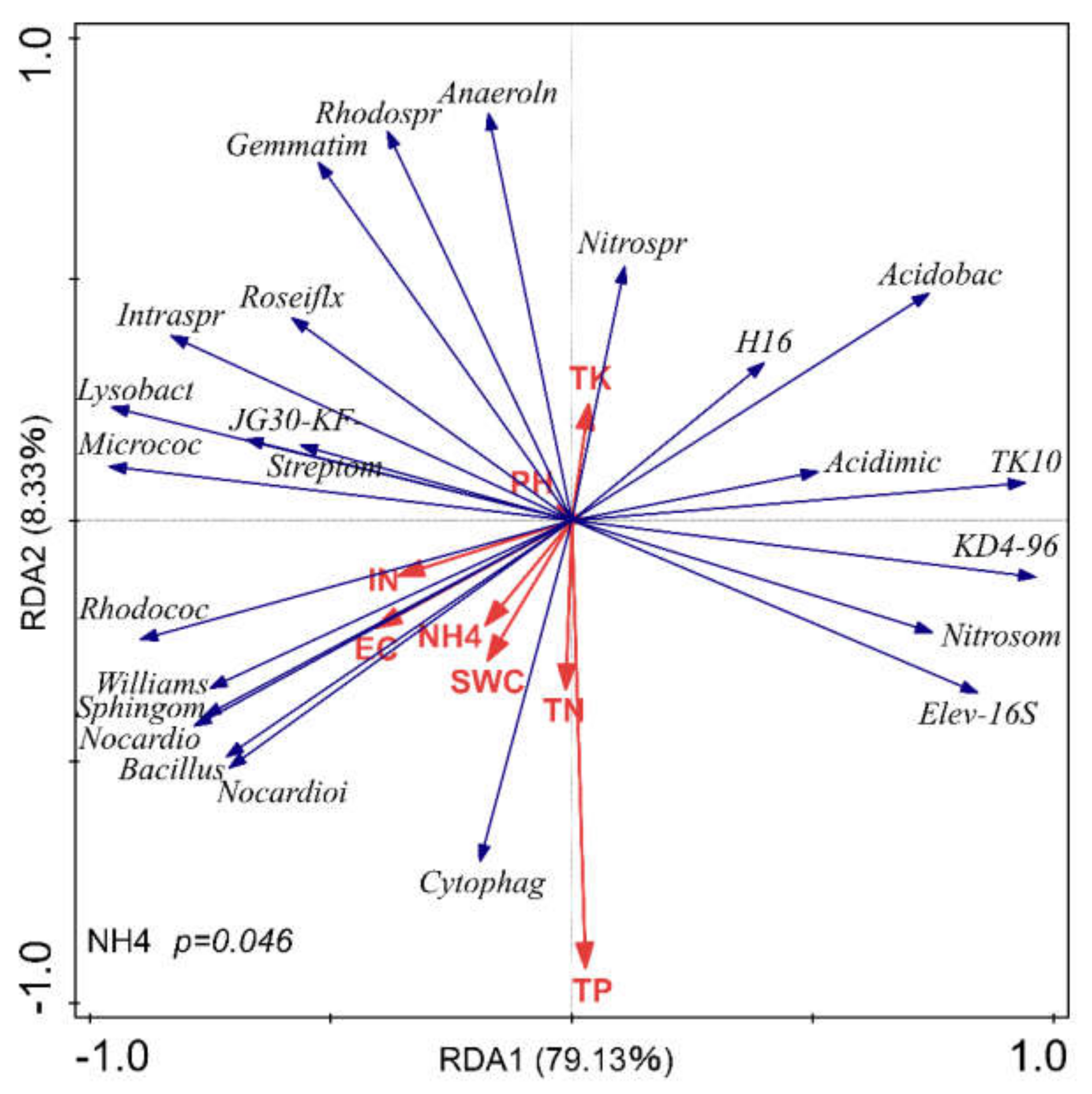
3.5. Correlations between Soil Physicochemical Properties and Soil Bacterial Community Composition
4. Discussion
4.1. Effect of Biochar and PGPR on Soil Nutrient Content
4.2. Soil Microbial α-Diversity Indices
4.3. Soil Bacterial Community Composition (Phylum Level)
4.4. Soil Bacterial Community Composition (Genus Level)
4.5. Suggestion for Using Biochar and PGPR to Improve Soil Properties and Bacterial Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chan, K.Y.; van Zwieten, L.; Meszaros, I.; Downie, A.; Joseph, S. Using poultry litter biochars as soil amendments. Soil Res. 2008, 46, 437–444. [Google Scholar] [CrossRef]
- Glaser, B.; Lehmann, J.; Zech, W. Ameliorating physical and chemical properties of highly weathered soils in the tropics with charcoal—A review. Biol. Fertil. Soils 2002, 35, 219–230. [Google Scholar] [CrossRef]
- Lehmann, J.; Gaunt, J.; Rondon, M. Bio-char sequestration in terrestrial ecosystems—A review. Mitig. Adapt. Strateg. Glob. Chang. 2006, 11, 403–427. [Google Scholar] [CrossRef]
- Abiven, S.; Hund, A.; Martinsen, V.; Cornelissen, G. Biochar amendment increases maize root surface areas and branching: A shovelomics study in Zambia. Plant Soil 2015, 395, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Dugan, E.; Verhoef, A.; Robinson, S. Bio-char from sawdust, maize stover and charcoal: Impact on water holding capacities (WHC) of three soils from Ghana. In Proceedings of the 19th World Congress of Soil Science: Soil solutions for a changing world, Symposium 4.2.2 Soil and water-global change, Brisbane, Australia, 1–6 August 2010; pp. 9–12. [Google Scholar]
- Jindo, K.; Mizumoto, H.; Sawada, Y.; Sanchez-Monedero, M.A.; Sonoki, T. Physical and chemical characterization of biochars derived from different agricultural residues. Biogeosciences Discuss. 2014, 11, 6613–6621. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Godínez, L.J.; Fernandez-Valverde, S.L.; Romero, J.C.M.; Martínez-Romero, E. Metatranscriptomics and nitrogen fixation from the rhizoplane of maize plantlets inoculated with a group of PGPRs. Syst. Appl. Microbiol. 2019, 42, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1–19. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Y.; Shen, F.; Tian, D.; Zeng, Y.; Yang, G.; Zhang, Y.; Deng, S. Partitioning biochar properties to elucidate their contributions to bacterial and fungal community composition of purple soil. Sci. Total Environ. 2019, 648, 1333–1341. [Google Scholar] [CrossRef]
- Nishizawa, T.; Tago, K.; Uei, Y.; Ishii, S.; Isobe, K.; Otsuka, S.; Senoo, K. Advantages of functional single-cell isolation method over standard agar plate dilution method as a tool for studying denitrifying bacteria in rice paddy soil. AMB Express 2012, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, P.; Tribedi, P. Functional diversity performs a key role in the isolation of nitrogen-fixing and phosphate-solubilizing bacteria from soil. Folia Microbiol. 2019, 64, 461–470. [Google Scholar] [CrossRef]
- Boon, N.; Marlé, C.; Top, E.M.; Verstraete, W. Comparison of the spatial homogeneity of physico-chemical parameters and bacterial 16S rRNA genes in sediment samples from a dumping site for dredging sludge. Appl. Microbiol. Biotechnol. 2000, 53, 742–747. [Google Scholar] [CrossRef]
- Handelsman, J.; Rondon, M.R.; Brady, S.F.; Clardy, J.; Goodman, R.M. Molecular biological access to the chemistry of unknown soil microbes: A new frontier for natural products. Chem. Biol. 1998, 5, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Fortuna, A.M.; Marsh, T.L.; Honeycutt, C.W.; Halteman, W.A. Use of primer selection and restriction enzymes to assess bacterial community diversity in an agricultural soil used for potato production via terminal restriction fragment length polymorphism. Appl. Microbiol. Biotechnol. 2011, 91, 1193–1202. [Google Scholar] [CrossRef]
- Chhabra, S.; Brazil, D.; Morrissey, J.; Burke, J.; O’Gara, F.; Dowling, D.N. Fertilization management affects the alkaline phosphatase bacterial community in barley rhizosphere soil. Biol. Fertil. Soils 2013, 49, 31–39. [Google Scholar] [CrossRef]
- Chen, F.; Zheng, H.; Zhang, K.; Ouyang, Z.; Wu, Y.; Shi, Q.; Li, H. Non-linear impacts of Eucalyptus plantation stand age on soil microbial metabolic diversity. J. Soils Sediments 2013, 13, 887–894. [Google Scholar] [CrossRef]
- Tabassum, B.; Khan, A.; Tariq, M.; Ramzan, M.; Iqbal Khan, M.S.; Shahid, N.; Aaliya, K. Bottlenecks in commercialisation and future prospects of PGPR. Appl. Soil Ecol. 2017, 121, 102–117. [Google Scholar] [CrossRef]
- Ren, H.; Lv, C.; Fernández-García, V.; Huang, B.; Yao, J. Biochar and PGPR amendments influence soil enzyme activities and nutrient concentrations in a eucalyptus seedling plantation. Biomass Convers. Biorefinery 2019. [Google Scholar] [CrossRef]
- Li, Y.; Wen, H.; Chen, L.; Yin, T. Succession of bacterial community structure and diversity in soil along a chronosequence of reclamation and re-vegetation on coal mine spoils in China. PLoS ONE 2014, 9, e115024. [Google Scholar] [CrossRef]
- Agegnehu, G.; Bass, A.M.; Nelson, P.N.; Bird, M.I. Benefits of biochar, compost and biochar-compost for soil quality, maize yield and greenhouse gas emissions in a tropical agricultural soil. Sci. Total Environ. 2016, 543, 295–306. [Google Scholar] [CrossRef]
- Basso, A.S.; Miguez, F.E.; Laird, D.A.; Horton, R.; Westgate, M. Assessing potential of biochar for increasing water-holding capacity of sandy soils. GCB Bioenergy 2013, 5, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Crombie, K.; Maek, O.; Cross, A.; Sohi, S. Biochar—Synergies and trade-offs between soil enhancing properties and C sequestration potential. GCB Bioenergy 2015, 7, 1161–1175. [Google Scholar] [CrossRef] [Green Version]
- Graber, E.R.; Harel, Y.M.; Kolton, M.; Cytryn, E.; Silber, A.; David, D.R.; Tsechansky, L.; Borenshtein, M.; Elad, Y. Biochar impact on development and productivity of pepper and tomato grown in fertigated soilless media. Plant Soil 2010, 337, 481–496. [Google Scholar] [CrossRef]
- Asai, H.; Samson, B.K.; Stephan, H.M.; Songyikhangsuthor, K.; Homma, K.; Kiyono, Y.; Inoue, Y.; Shiraiwa, T.; Horie, T. Biochar amendment techniques for upland rice production in Northern Laos. 1. Soil physical properties, leaf SPAD and grain yield. Field Crop. Res. 2009, 111, 81–84. [Google Scholar] [CrossRef]
- Yuanyuan, L.; Longqian, C. 454 Pyrosequencing Analysis of Bacterial Diversity Revealed by a Comparative Study of Soils from Mining Subsidence and Reclamation Areas. J. Microbiol. Biotechnol. 2014, 24, 313–323. [Google Scholar]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Ren, H.; Qin, X.; Huang, B.; Fernández-García, V.; Chengqun, L. Responses of soil enzyme activities and plant growth in a eucalyptus seedling plantation amended with bacterial fertilizers. Arch. Microbiol. 2020. [Google Scholar] [CrossRef]
- Docherty, K.M.; Balser, T.C.; Bohannan, B.J.M.; Gutknecht, J.L.M. Soil microbial responses to fire and interacting global change factors in a California annual grassland. Biogeochemistry 2012, 109, 63–83. [Google Scholar] [CrossRef]
- Zhang, N.; Wan, S.; Guo, J.; Han, G.; Gutknecht, J.; Schmid, B.; Yu, L.; Liu, W.; Bi, J.; Wang, Z.; et al. Precipitation modifies the effects of warming and nitrogen addition on soil microbial communities in northern Chinese grasslands. Soil Biol. Biochem. 2015, 89, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Calderón, K.; Spor, A.; Breuil, M.C.; Bru, D.; Bizouard, F.; Violle, C.; Barnard, R.L.; Philippot, L. Effectiveness of ecological rescue for altered soil microbial communities and functions. ISME J. 2017, 11, 272–283. [Google Scholar] [CrossRef] [Green Version]
- Kimetu, J.M.; Lehmann, J. Stability and stabilisation of biochar and green manure in soil with different organic carbon contents. Aust. J. Soil Res. 2010, 48, 577–585. [Google Scholar] [CrossRef]
- Lu, W.; Ding, W.; Zhang, J.; Li, Y.; Luo, J.; Bolan, N.; Xie, Z. Biochar suppressed the decomposition of organic carbon in a cultivated sandy loam soil: A negative priming effect. Soil Biol. Biochem. 2014, 76, 12–21. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil. 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Gaskin, J.W.; Steiner, C.; Harris, K.; Das, K.C.; Bibens, B. Effect of low-temperature pyrolysis conditions on biochar for agricultural use. Trans. ASABE 2008, 51, 2061–2069. [Google Scholar] [CrossRef]
- Sharma, R.; Sharma, P.; Chauhan, A.; Walia, A.; Shirkot, C.K. Plant growth promoting activities of rhizobacteria isolated from Podophyllum hexandrum growing in North-West regions of the Himalaya. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 1443–1457. [Google Scholar] [CrossRef]
- Clough, T.J.; Condron, L.M.; Kammann, C.; Müller, C. A Review of Biochar and Soil Nitrogen Dynamics. Agronomy 2013, 3, 275–293. [Google Scholar] [CrossRef] [Green Version]
- Oram, N.J.; Van de Voorde, T.F.J.; Ouwehand, G.J.; Bezemer, T.M.; Mommer, L.; Jeffery, S.; van Groenigen, J.W. Soil amendment with biochar increases the competitive ability of legumes via increased potassium availability. Agric. Ecosyst. Environ. 2014, 191, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xing, W.; Bao, H.; Wang, J.; Tong, X.; Zhang, H.; Luo, W.; Wu, F. Impact of pine leaf biochar amendment on bacterial dynamics and correlation of environmental factors during pig manure composting. Bioresou. Technol. 2019, 293, 1–10. [Google Scholar] [CrossRef]
- Qayyum, M.F.; Ashraf, I.; Abid, M.; Steffens, D. Effect of biochar, lime, and compost application on phosphorus adsorption in a Ferralsol. J. Plant Nutr. Soil Sci. 2015, 178, 576–581. [Google Scholar] [CrossRef]
- Muhammad, N.; Dai, Z.; Xiao, K.; Meng, J.; Brookes, P.C.; Liu, X.; Wang, H.; Wu, J.; Xu, J. Changes in microbial community structure due to biochars generated from different feedstocks and their relationships with soil chemical properties. Geoderma 2014, 226–227, 270–278. [Google Scholar] [CrossRef]
- Rondon, M.A.; Lehmann, J.; Ramírez, J.; Hurtado, M. Biological nitrogen fixation by common beans (Phaseolus vulgaris L.) increases with bio-char additions. Biol. Fertil. Soils 2007, 43, 699–708. [Google Scholar] [CrossRef]
- Varin, T.; Lovejoy, C.; Jungblut, A.D.; Vincent, W.F.; Corbeil, J. Metagenomic profiling of Arctic microbial mat communities as nutrient scavenging and recycling systems. Limnol. Oceanogr. 2010, 55, 1901–1911. [Google Scholar] [CrossRef] [Green Version]
- Kolb, S.E.; Fermanich, K.J.; Dornbush, M.E. Effect of Charcoal Quantity on Microbial Biomass and Activity in Temperate Soils. Soil Sci. Soc. Am. J. 2009, 73, 1173–1181. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.C.; Shen, Y.Y.; Xu, G.P.; Huang, Y.Q.; Zhang, D.N.; Sun, Y.J.; Li, Y.Q.; He, W.; Zhou, L.W. Effects of Reclamation on Soil Nutrients and Microbial Activities in the Huixian Karst Wetland in Guilin. Huanjing Kexue Environ. Sci. 2018, 39, 1813–1823. [Google Scholar]
- Ogawa, M.; Okimori, Y. Pioneering works in biochar research, Japan. Aust. J. Soil Res. 2010, 48, 489–500. [Google Scholar] [CrossRef] [Green Version]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global Biogeography and Quantitative Seasonal Dynamics of Gemmatimonadetes in Soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [Green Version]
- Khodadad, C.L.M.; Zimmerman, A.R.; Green, S.J.; Uthandi, S.; Foster, J.S. Taxa-specific changes in soil microbial community composition induced by pyrogenic carbon amendments. Soil Biol. Biochem. 2011, 43, 385–392. [Google Scholar] [CrossRef]
- Martínez-Pérez, C.; Mohr, W.; Löscher, C.R.; Dekaezemacker, J.; Littmann, S.; Yilmaz, P.; Lehnen, N.; Fuchs, B.M.; Lavik, G.; Schmitz, R.A.; et al. The small unicellular diazotrophic symbiont, UCYN-A, is a key player in the marine nitrogen cycle. Nat. Microbiol. 2016, 1, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Marcarelli, A.M.; Wurtsbaugh, W.A.; Griset, O. Salinity controls phytoplankton response to nutrient enrichment in the Great Salt Lake, Utah, USA. Can. J. Fish. Aquat. Sci. 2006, 63, 2236–2248. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Paerl, H.W.; Zhu, G.; Qin, B.; Hall, N.S.; Zhu, M. Long-term nutrient trends and harmful cyanobacterial bloom potential in hypertrophic Lake Taihu, China. Hydrobiologia 2016, 787, 1–14. [Google Scholar] [CrossRef]
- Koch, H.; Lücker, S.; Albertsen, M.; Kitzinger, K.; Herbold, C.; Spieck, E.; Nielsen, P.H.; Wagner, M.; Daims, H. Expanded metabolic versatility of ubiquitous nitrite-oxidizing bacteria from the genus Nitrospira. Proc. Natl. Acad. Sci. USA 2015, 112, 11371–11376. [Google Scholar] [CrossRef] [Green Version]
- Wagner, M.; Horn, M. The Planctomycetes, Verrucomicrobia, Chlamydiae and sister phyla comprise a superphylum with biotechnological and medical relevance. Curr. Opin. Biotechnol. 2006, 17, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Pol, A.; Heijmans, K.; Harhangi, H.R.; Tedesco, D.; Jetten, M.S.M.; Op Den Camp, H.J.M. Methanotrophy below pH 1 by a new Verrucomicrobia species. Nature 2007, 450, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Islam, T.; Jensen, S.; Reigstad, L.J.; Larsen, O.; Birkeland, N.K. Methane oxidation at 55 °C and pH 2 by a thermoacidophilic bacterium belonging to the Verrucomicrobia phylum. Proc. Natl. Acad. Sci. USA 2008, 105, 300–304. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Grogan, P.; Caporaso, J.G.; Zhang, H.; Lin, X.; Knight, R.; Chu, H. PH is a good predictor of the distribution of anoxygenic purple phototrophic bacteria in Arctic soils. Soil Biol. Biochem. 2014, 74, 193–200. [Google Scholar] [CrossRef]
- Hou, J.; Liu, W.; Wang, B.; Wang, Q.; Luo, Y.; Franks, A.E. PGPR enhanced phytoremediation of petroleum contaminated soil and rhizosphere microbial community response. Chemosphere 2015, 138, 592–598. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Jiang, X.; Tong, T.; Miao, S.; Huang, J.; Xie, S. Sulfadiazine degradation in soils: Dynamics, functional gene, antibiotic resistance genes and microbial community. Sci. Total Environ. 2019, 691, 1072–1081. [Google Scholar] [CrossRef]
- Deslippe, J.R.; Hartmann, M.; Simard, S.W.; Mohn, W.W. Long-term warming alters the composition of Arctic soil microbial communities. FEMS Microbiol. Ecol. 2012, 82, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, G.; Hai, X.; Li, J.; Shangguan, Z.; Peng, C.; Deng, L. Long-term forest succession improves plant diversity and soil quality but not significantly increase soil microbial diversity: Evidence from the Loess Plateau. Ecol. Eng. 2020, 142, 1–13. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fixed C (g kg−1) | Av.P (g kg−1) | Av. K (g kg−1) | Bulk (g cm−3) | SA (m2 g−1) | Porosity (%) | pH | EC (mS cm−1) | CEC (cmol kg−1) |
|---|---|---|---|---|---|---|---|---|
| 650 | 10.20 | 55.65 | 0.19 | 9 | 67.03 | 10.24 | 4.68 | 60.80 |
| Aimed Object | Primer | Sequence (5’–3’) |
|---|---|---|
| Bacterial16S rRNA gene | 338F | ACTCCTACGGGAGGCAGCAG |
| 806R | GGACTACHVGGGTWTCTAAT |
| PCR Reaction System (20 μL) | Addition (μL) | Amplification System |
|---|---|---|
| 5×FastPfu Buffer | 4 | Denaturation at 95 ℃ for 3 min |
| 2.5mM dNTPs | 2 | Degeneration at 95 ℃ for 30 s |
| Forward Primer (5 µM) | 0.8 | Annealing at 55 ℃ for 30 s |
| Reverse Primer (5 µM) | 0.8 | Extension at 72 ℃ for 45 s |
| FastPfu Polymerase | 0.4 | 25 recycling |
| BSA | 0.2 | Extension at 72 ℃ for 10 min |
| Template DNA | 10 ng | |
| Add ddH2O to | 20 | Stored at 10 ℃ |
| Treatment | NO3−-N (mg g−1) | NH4+-N (mg g−1) | IN (mg g−1) | TN (mg g−1) | TP (mg g−1) | TK(mg g−1) | pH | EC (dS m−1) | SWC (%) |
|---|---|---|---|---|---|---|---|---|---|
| Control | 0.038 ± 0.002 c | 0.028 ± 0.001 a | 0.066 ± 0.003 c | 1.03 ± 0.05 c | 2.31 ± 0.12 a | 1.45 ± 0.07 c | 8.33 ± 0.42 ab | 105 ± 5 b | 13 ± 1 c |
| Biochar | 0.050 ± 0.003 b | 0.025 ± 0.001 b | 0.075 ± 0.004 b | 1.28 ± 0.06 b | 1.99 ± 0.10 b | 1.58 ± 0.08 c | 7.84 ± 0.39 b | 128 ± 6 a | 14 ± 1 b |
| PGPR | 0.024 ± 0.001 d | 0.025 ± 0.001 b | 0.049 ± 0.003 d | 2.06 ± 0.10 a | 1.86 ± 0.09 b | 1.77 ± 0.09 b | 8.78 ± 0.44 a | 88 ± 4 c | 13 ± 1 bc |
| Biochar+PGPR | 0.064 ± 0.003 a | 0.025 ± 0.001 ab | 0.089 ± 0.005 a | 1.06 ± 0.05 c | 1.41 ± 0.07 c | 2.25 ± 0.11 a | 7.89 ± 0.39 b | 135 ± 7 a | 17 ± 1 a |
| Treatments | Reads | OTUs | Coverage | Richness and Diversity Indices | ||
|---|---|---|---|---|---|---|
| Simpson | ACE | Chao | ||||
| Control | 26784 ± 1339 c | 1864 ± 58 c | 0.98 ± 0.00 b | 0.0032 ± 0.0005 d | 2176 ± 26 b | 2188 ± 16 c |
| Biochar | 35153 ± 1758 b | 2185 ± 71 b | 0.99 ± 0.00 a | 0.0190 ± 0.0047 a | 2615 ± 29 a | 2631 ± 1 b |
| PGPR | 41703 ± 2085 a | 2408 ± 205 a | 0.99 ± 0.00 a | 0.0066 ± 0.0005 c | 2714 ± 211 a | 2696 ± 277 ab |
| Biochar+PGPR | 35036 ± 1752 b | 2324 ± 1 a | 0.99 ± 0.01 ab | 0.0107 ± 0.0008 b | 2721 ± 110 a | 2767 ± 135 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, H.; Huang, B.; Fernández-García, V.; Miesel, J.; Yan, L.; Lv, C. Biochar and Rhizobacteria Amendments Improve Several Soil Properties and Bacterial Diversity. Microorganisms 2020, 8, 502. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040502
Ren H, Huang B, Fernández-García V, Miesel J, Yan L, Lv C. Biochar and Rhizobacteria Amendments Improve Several Soil Properties and Bacterial Diversity. Microorganisms. 2020; 8(4):502. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040502
Chicago/Turabian StyleRen, Han, Baoling Huang, Víctor Fernández-García, Jessica Miesel, Li Yan, and Chengqun Lv. 2020. "Biochar and Rhizobacteria Amendments Improve Several Soil Properties and Bacterial Diversity" Microorganisms 8, no. 4: 502. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040502