Emergence of a Multidrug-Resistant Enterobacter hormaechei Clinical Isolate from Egypt Co-Harboring mcr-9 and blaVIM-4

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Identification
2.2. Antimicrobial Susceptibility Testing (AST)
2.3. Filter-Mating Conjugation
2.4. Whole Genome Sequencing (WGS) and Analysis
2.5. Nucleotide Sequence Accession Numbers
3. Results and Discussion
3.1. AST of E. hormaechei AMS-38, and Conjugation of mcr-9 and blaVIM-4-Carrying Plasmid
3.2. Characterization of the Genome of E. hormaechei AMS-38
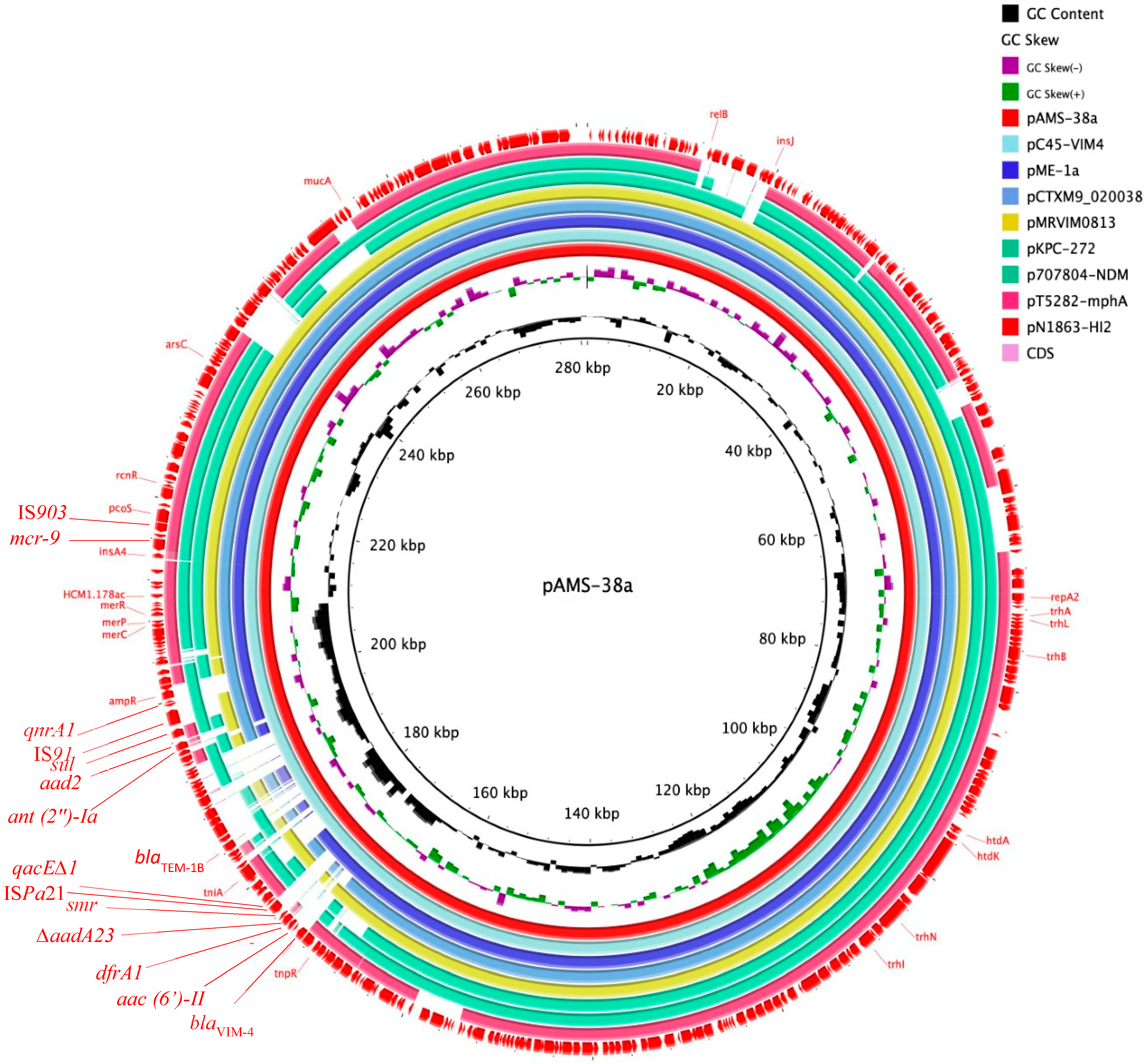
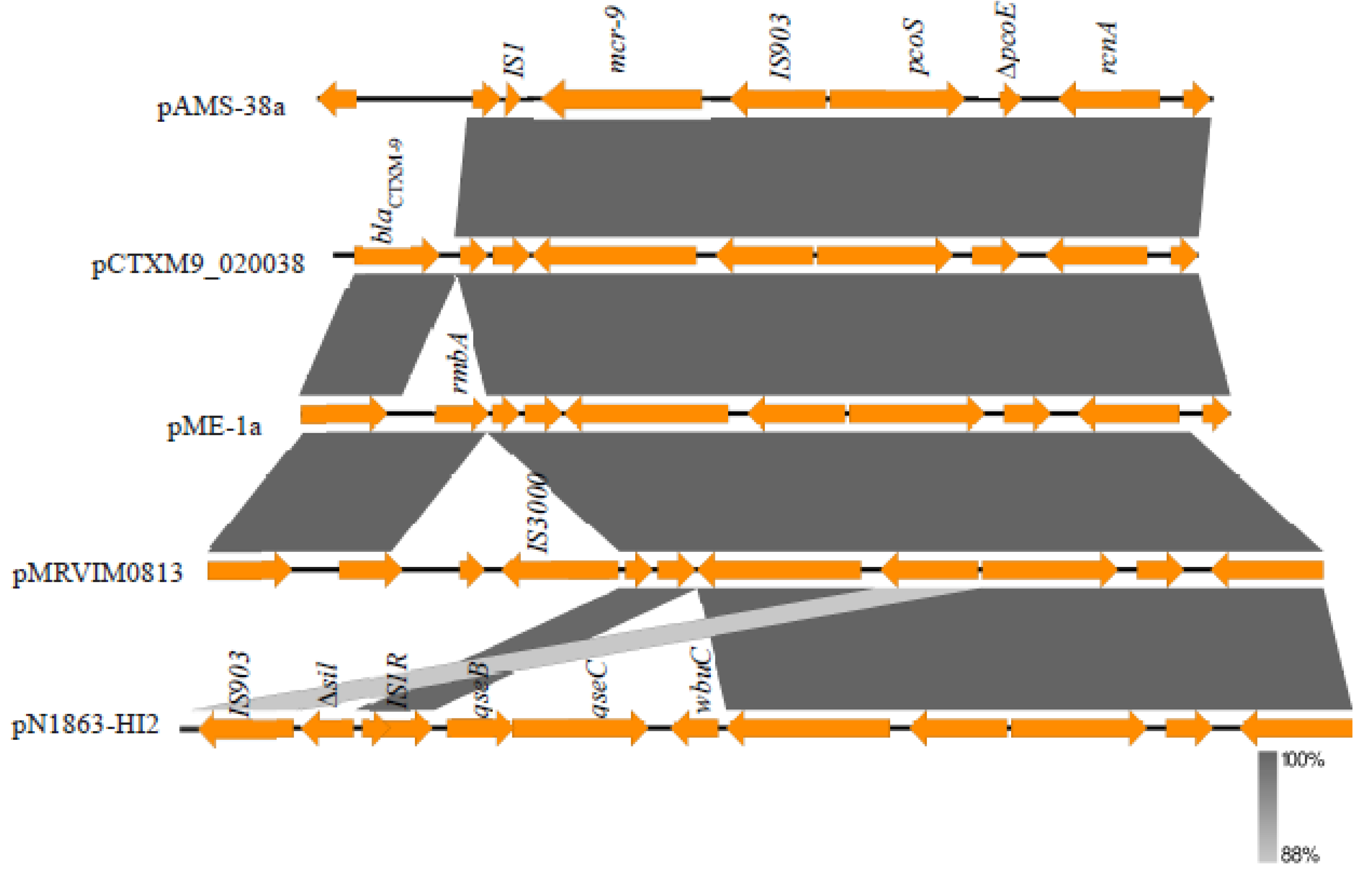
3.3. Plasmids and Resistome Analysis of the Strain E. hormaechei AMS-38
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Antimicrobial Resistance: Global Report on Surveillance 2014; WHO Press: Geneva, Switzerland, 2014. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Antibiotic resistance threats in the United States, 2013. Available online: http://www.cdc.gov/drugresistance/threat-report-2013/pdf/arthreats-2013-508.pdf (accessed on 24 March 2020).
- Hsu, L.Y.; Apisarnthanarak, A.; Khan, E.; Suwantarat, N.; Ghafur, A.; Tambyah, P.A. Carbapenem-resistant Acinetobacter baumannii and Enterobacteriaceae in South and Southeast Asia. Clin. Microbiol. Rev. 2016, 30, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Meunier, D.; Findlay, J.; Doumith, M.; Godoy, D.; Perry, C.; Pike, R.; Gronthoud, F.; Shryane, T.; Poirel, L.; Welfare, W.; et al. FRI-2 carbapenemase-producing Enterobacter cloacae complex in the UK. J. Antimicrob. Chemother. 2017, 72, 2478–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliman, A.M.; Zarad, H.O.; Nariya, H.; Shimamoto, T.; Shimamoto, T. Genetic analysis of carbapenemase-producing Gram-negative bacteria isolated from a university teaching hospital in Egypt. Infect. Genet. Evol. 2020, 77, 104065. [Google Scholar] [CrossRef] [PubMed]
- Hashem, H.; Hanora, A.; Abdalla, S.; Shaeky, A.; Saad, A. Dissemination of metallo-β-lactamase in Pseudomonas aeruginosa isolates in Egypt: Mutation in blaVIM-4. APMIS 2017, 125, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Sonnevend, A.; Ghazawi, A.; Yahfoufi, N.; Al-Baloushi, A.; Hashmey, R.; Mathew, M.; Tariq, W.Z.; Pal, T. VIM-4 carbapenemase-producing Enterobacter cloacae in the United Arab Emirates. Clin. Microb. Infect. 2012, 18, E494–E496. [Google Scholar] [CrossRef] [Green Version]
- Jamal, W.; Rotimi, V.O.; Albert, M.J.; Khodakhast, F.; Nordmann, P.; Poirel, L. High prevalence of VIM-4 and NDM-1 metallo-β-lactamase among carbapenem-resistant Enterobacteriaceae. J. Med. Microbiol. 2013, 62, 1239–1244. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Elnahriry, S.S.; Khalifa, H.O.; Soliman, A.M.; Ahmed, A.S.; Hussein, A.M.; Shimamoto, T.; Shimamoto, T. Emergence of plasmid-mediated colistin resistance gene mcr-1 in a clinical Escherichia coli isolate from Egypt. Antimicrob. Agents Chemother. 2016, 60, 3249–3250. [Google Scholar] [CrossRef] [Green Version]
- Kieffer, N.; Royer, G.; Decousser, J.W.; Bourrel, A.S.; Palmieri, M.; De La Rosa, J.M.; Jacquier, H.; Denamur, E.; Nordmann, P.; Poirel, L. mcr-9, an inducible gene encoding an acquired phosphoethanolamine transferase in Escherichia coli, and its origin. Antimicrob. Agents Chemother. 2019, AAC-00965. [Google Scholar] [CrossRef] [Green Version]
- Liang, Q.; Yin, Z.; Zhao, Y.; Liang, L.; Feng, J.; Zhan, Z.; Wang, H.; Song, Y.; Tong, Y.; Wu, W.; et al. Sequencing and comparative genomics analysis of the IncHI2 plasmids pT5282-mphA and p112298-catA and the IncHI5 plasmid pYNKP001-dfrA. Int. J. Antimicrob. Agents 2017, 49, 709–718. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Tanizawa, Y.; Fujisawa, T.; Nakamura, Y. DFAST: A flexible prokaryotic genome annotation pipeline for faster genome publication. Bioinformatics 2018, 34, 1037–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast genome and metagenome distance estimation using MinHash. Genome Biol. 2016, 17, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galata, V.; Fehlmann, T.; Backes, C.; Keller, A. PLSDB: A resource of complete bacterial plasmids. Nucleic Acids Res. 2019, 47, D195–D202. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Petty, N.K.; Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genomics 2011, 12, 402. [Google Scholar] [CrossRef] [Green Version]
- Chavda, K.D.; Westblade, L.F.; Satlin, M.J.; Hemmert, A.C.; Castanheira, M.; Jenkins, S.G.; Chen, L.; Kreiswirth, B.N. First report of blaVIM-4- and mcr-9-coharboring Enterobacter species isolated from a pediatric patient. mSphere 2019, 4, e00629-19. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Li, Y.; Wang, G.; Li, C.; Xiang, L.; She, J.; Yang, Y.; Zhong, F.; Zhang, L. Coproduction of MCR-9 and NDM-1 by colistin-resistant Enterobacter hormaechei isolated from bloodstream infection. Infect. Drug Resist. 2019, 12, 2979. [Google Scholar] [CrossRef] [Green Version]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of novel mobilized colistin resistance gene mcr-9 in a multidrug-resistant, colistin-susceptible Salmonella enterica serotype Typhimurium isolate. mBio 2019, 10, e00853-19. [Google Scholar] [CrossRef] [Green Version]
- Börjesson, S.; Greko, C.; Myrenås, M.; Landén, A.; Nilsson, O.; Pedersen, K. A link between the newly described colistin resistance gene mcr-9 and clinical Enterobacteriaceae isolates carrying blaSHV-12 from horses in Sweden. J. Glob. Antimicrob. Resist. 2020, 20, 285–289. [Google Scholar] [CrossRef]
- Faccone, D.; Martino, F.; Albornoz, E.; Gomez, S.; Corso, A.; Petroni, A. Plasmid carrying mcr-9 from an extensively drug-resistant NDM-1-producing Klebsiella quasipneumoniae subsp. quasipneumoniae clinical isolate. Infect. Genet. Evol. 2020, 81, 104273. [Google Scholar] [CrossRef]
- Wang, C.; Feng, Y.; Liu, L.; Wei, L.; Kang, M.; Zong, Z. Identification of novel mobile colistin resistance gene mcr-10. Emerg. Microbes Infect. 2020, 9, 508–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimude, J.U.; Amyes, S.G.B. Molecular characterisation and diversity in Enterobacter cloacae from Edinburgh and Egypt carrying blaCTX-M-14 and blaVIM-4 β-lactamase genes. Int. J. Antimicrob. Agents 2013, 41, 574–577. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample | Size (bp) | GC% | No. of CDSs | MLST or pMLST | Inc Type * | AMR Gene |
|---|---|---|---|---|---|---|
| Chromosome | 4,914,941 | 54.9 | 6279 | ST133 | ND | fosA, blaACT-7 |
| pAMS-38a | 281,121 | 46.8 | 361 | ST1 | IncHI2 | mcr-9, sul1, qnrA1, dfrA1, blaTEM-1B, blaVIM-4, aac(6’)-Il, aadA23, aadA2b, ant(2’’)-Ia |
| pAMS-38b | 109,015 | 50.8 | 144 | ND | IncFIB(pHCM2) | ND |
| pAMS-38c | 9525 | 51.7 | 15 | ND | ND | ND |
| pAMS-38d | 2511 | 51.8 | 2 | ND | ND | ND |
| Plasmid | Distance To pAMS-38a * | Bacterial Species | Isolation Year ** | Country | Accession No. | |
|---|---|---|---|---|---|---|
| 1 | pC45-VIM4 | 0.0022 | E. cloacae complex | 2014 | France | LT991958 |
| 2 | pME-1a | 0.0026 | E. hormaechei subsp. steigerwaltii | 2018 | USA | CP041734 |
| 3 | pCTXM9_020038 | 0.0048 | E. hormaechei | 2016 | China | CP031724 |
| 4 | pMRVIM0813 | 0.0049 | E. cloacae | 2015 | USA | KP975077 |
| 5 | pKPC-272 | 0.0079 | E. cloacae | 2012 | USA | NZ_CP008825.1 |
| 6 | p707804-NDM | 0.0080 | Leclercia adecarboxylata | ND | China | MH909331.1 |
| 7 | pT5282-mphA | 0.0108 | E. cloacae | 2012 | China | KY270852 |
| 8 | pN1863-HI2 | 0.0120 | E. cloacae | ND | China | MF344583 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soliman, A.M.; Maruyama, F.; Zarad, H.O.; Ota, A.; Nariya, H.; Shimamoto, T.; Shimamoto, T. Emergence of a Multidrug-Resistant Enterobacter hormaechei Clinical Isolate from Egypt Co-Harboring mcr-9 and blaVIM-4. Microorganisms 2020, 8, 595. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040595
Soliman AM, Maruyama F, Zarad HO, Ota A, Nariya H, Shimamoto T, Shimamoto T. Emergence of a Multidrug-Resistant Enterobacter hormaechei Clinical Isolate from Egypt Co-Harboring mcr-9 and blaVIM-4. Microorganisms. 2020; 8(4):595. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040595
Chicago/Turabian StyleSoliman, Ahmed M., Fumito Maruyama, Hoda O. Zarad, Atsushi Ota, Hirofumi Nariya, Toshi Shimamoto, and Tadashi Shimamoto. 2020. "Emergence of a Multidrug-Resistant Enterobacter hormaechei Clinical Isolate from Egypt Co-Harboring mcr-9 and blaVIM-4" Microorganisms 8, no. 4: 595. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8040595