Selection of Wild Lactic Acid Bacteria Strains as Promoters of Postbiotics in Gluten-Free Sourdoughs


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials, Chemicals, and Feedstocks
2.2. Prospection of Microorganisms and Culture Conditions
2.2.1. Enrichment and Fermentation Procedures for Microbial Prospection of Lactic Acid Bacteria
2.2.2. Microbiological Growth, Isolation, and Purification of Presumptive Lactic Acid Bacteria
2.2.3. Microbiological Identification of Presumptive Lactic Acid Bacteria to the Genus Level
2.2.4. Maintenance and Reactivation of Pure Cultures of Presumptive Lactic Acid Bacteria
2.2.5. Microbiological Growth, Isolation, and Purification of Indicator Microorganisms
2.3. Evaluation of the Capacity of Presumptive Lactic Acid Bacteria to Produce Exopolysaccharides
2.4. Evaluation of the Acidification Capacity of Presumptive Lactic Acid Bacteria
2.5. Evaluation of the Antimicrobial Properties of Flour Extracts Fermented with Presumptive Lactic Acid Bacteria
2.5.1. Antifungal Activity by Agar Well Diffusion Assay
2.5.2. Antibacterial Activity
2.5.3. Stability of the Antimicrobial Capacity of the Fermented Flour Extracts after Undergoing Thermal and Acidification Treatments
Antifungal Activity of Fermented Flour Extracts after Undergoing Thermal and Acidification Treatments
Antibacterial Activity of Fermented Flour Extracts after Undergoing Thermal and Acidification Treatments
2.6. Statistical Analysis
3. Results
3.1. Isolation of Wild Presumptive Lactic Acid Bacteria
3.2. Characterization of the Technological and Functional Properties of the Presumptive LAB Strains
3.2.1. Production of Exopolysaccharides by Presumptive Lactic Acid Bacteria
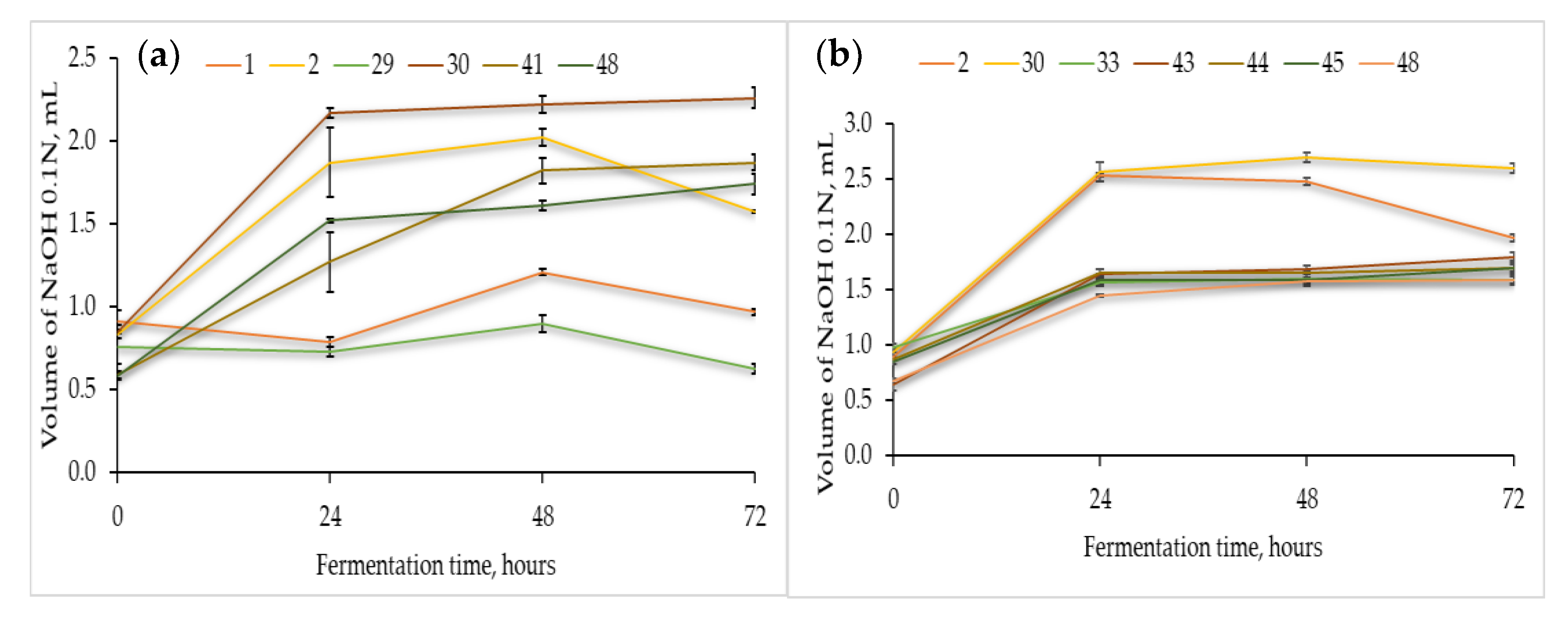
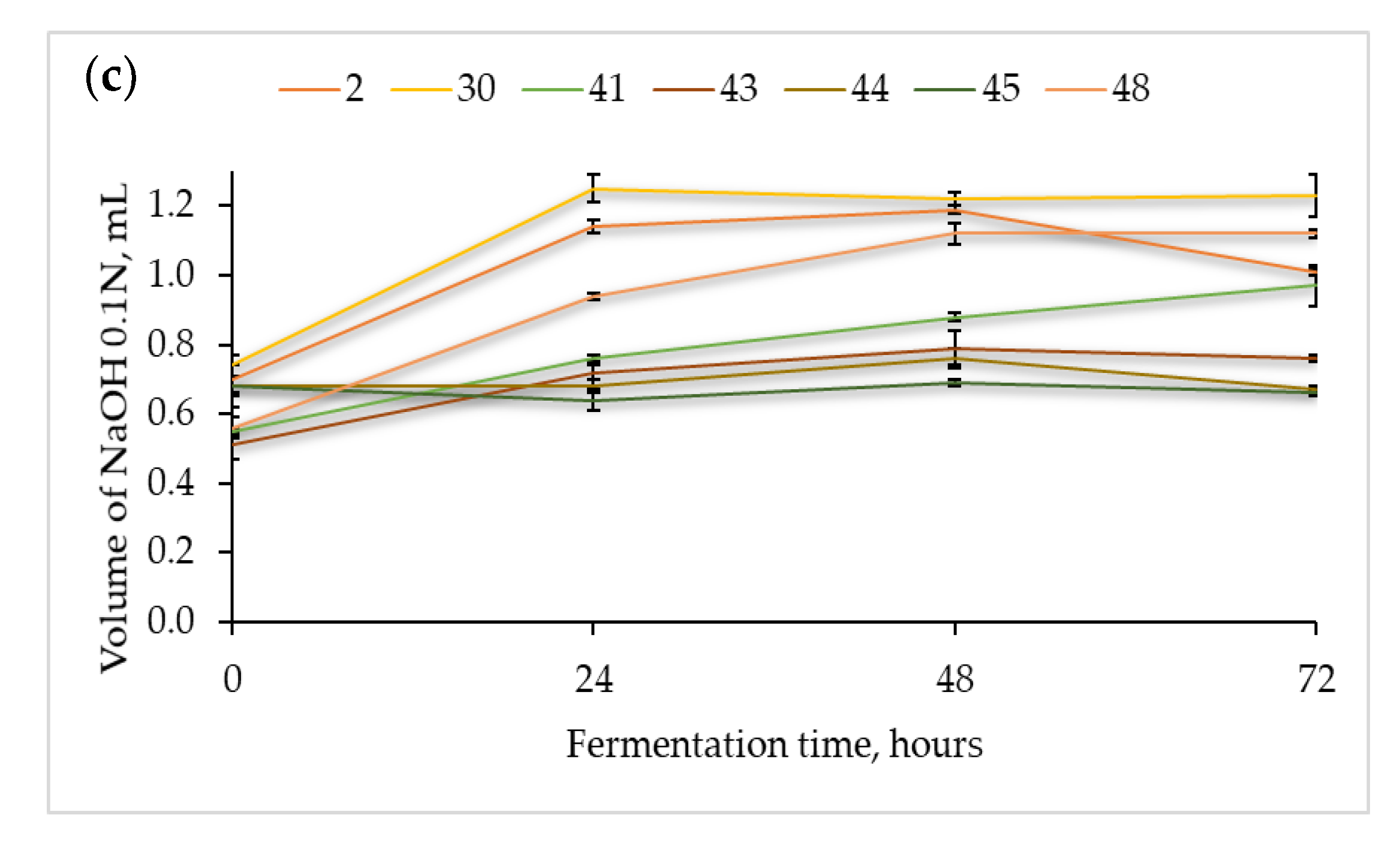
3.2.2. Acidification Capacity of the Presumptive Lactic Acid Bacteria
3.2.3. Antimicrobial Properties of Flours Fermented with Presumptive Lactic Acid Bacteria
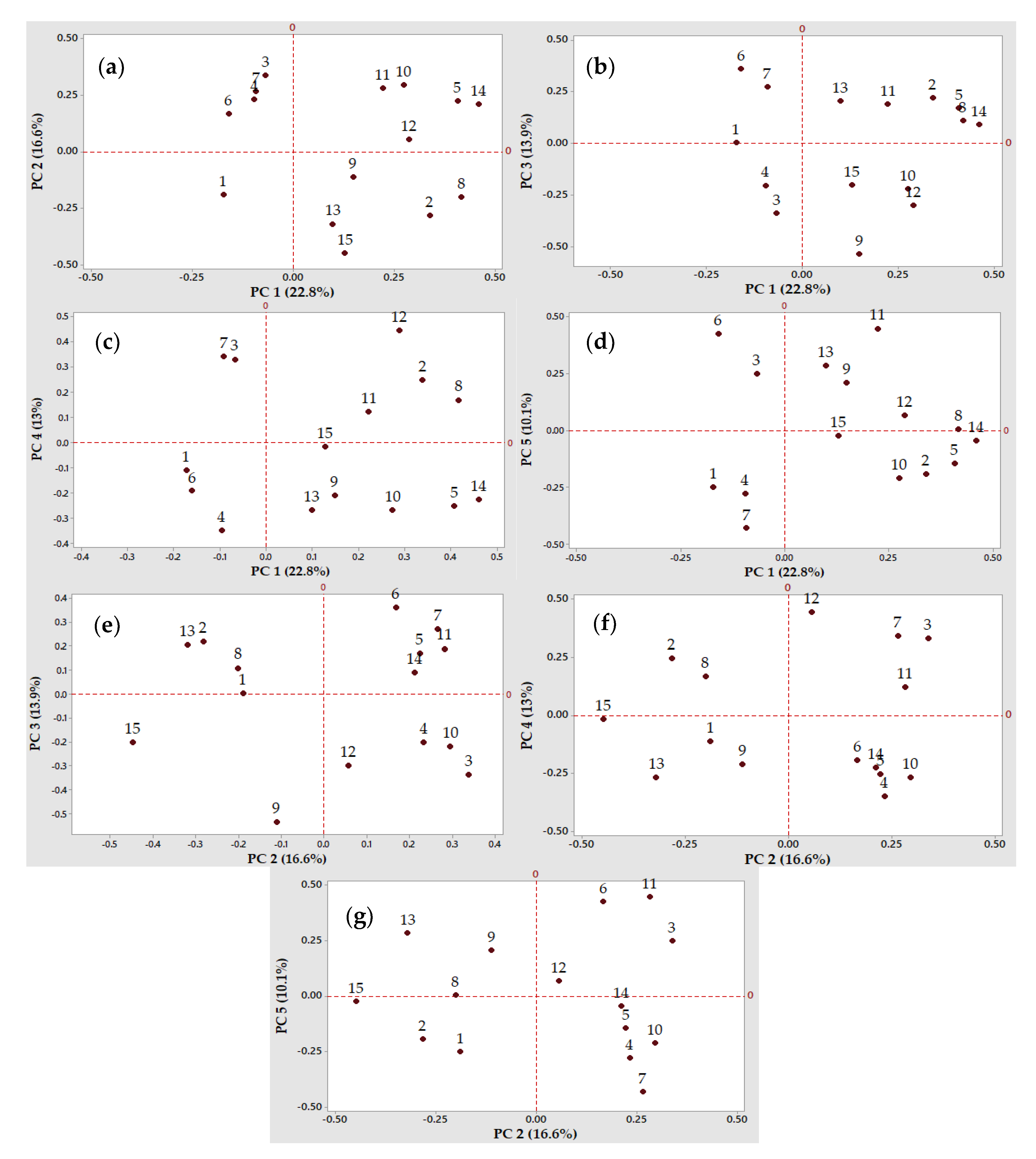
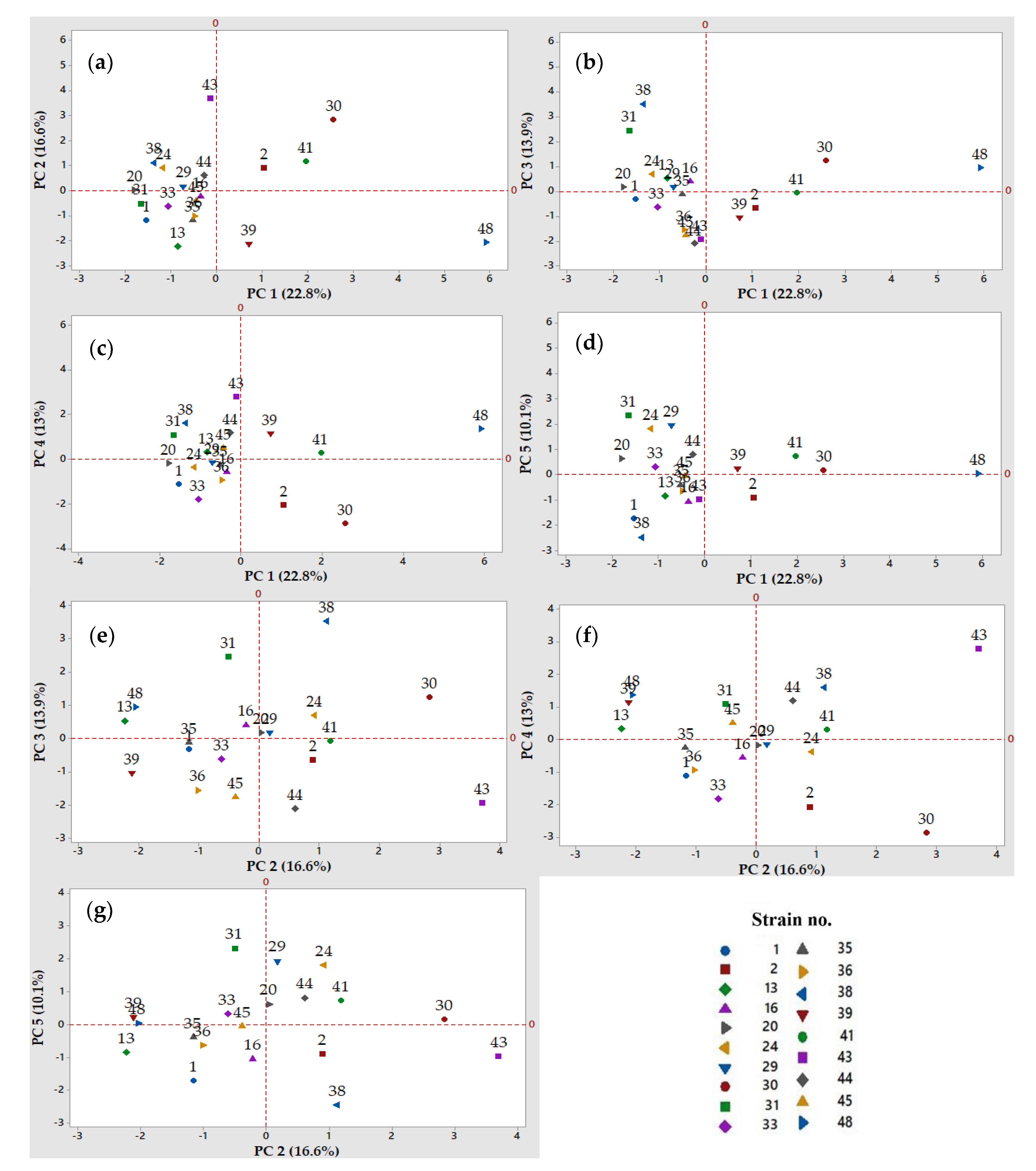
3.2.4. Principal Component Analysis to Discriminate the Most Relevant Factors and Presumptive Lactic Acid Bacteria strains for Further Application in Non-Gluten Sourdough Fermentations
3.2.5. Stability of the Antimicrobial Properties of Fermented Flour Extracts after Undergoing Thermal and Acidification Treatments
4. Discussion
4.1. Selection of Presumptive Lactic Acid Bacteria Strains Based on Their Fermentation Traits, Using Multivariate Analysis
4.2. Capacity of Presumptive Lactic Acid Bacteria to Produce Exopolysaccharides
4.3. Antimicrobial Properties of Fermented Flour Extracts with Presumptive Lactic Acid Bacteria against Typical Spoilage Microorganisms in Bakery Products
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Quigley, E.M.M.; Gajula, P.; Van De Wiele Belgium, T.; Val D’hebron, H. Open Peer Review Recent advances in modulating the microbiome [version 1; peer review: 2 approved]. F1000 Fac. Rev. 2020, 9, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimarães, J.T.; Balthazar, C.F.; Silva, R.; Rocha, R.S.; Graça, J.S.; Esmerino, E.A.; Silva, M.C.; Sant’Ana, A.S.; Duarte, M.C.K.H.; Freitas, M.Q.; et al. Impact of probiotics and prebiotics on food texture. Curr. Opin. Food Sci. 2020, 33, 38–44. [Google Scholar] [CrossRef]
- Yang, H.; Sun, Y.; Cai, R.; Chen, Y.; Gu, B. The impact of dietary fiber and probiotics in infectious diseases. Microb. Pathog. 2020, 140, 103931. [Google Scholar] [CrossRef] [PubMed]
- Xavier-Santos, D.; Bedani, R.; Lima, E.D.; Saad, S.M.I. Impact of probiotics and prebiotics targeting metabolic syndrome. J. Funct. Foods 2020, 64, 103666. [Google Scholar] [CrossRef]
- de Almada, C.N.; Almada, C.N.; Martinez, R.C.R.; Sant’Ana, A.S. Paraprobiotics: Evidences on their ability to modify biological responses, inactivation methods and perspectives on their application in foods. Trends Food Sci. Technol. 2016, 58, 96–114. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Zendeboodi, F.; Khorshidian, N.; Mortazavian, A.M.; da Cruz, A.G. Probiotic: Conceptualization from a new approach. Curr. Opin. Food Sci. 2020. [Google Scholar] [CrossRef]
- Pyclik, M.; Srutkova, D.; Schwarzer, M.; Górska, S. Bifidobacteria cell wall-derived exo-polysaccharides, lipoteichoic acids, peptidoglycans, polar lipids and proteins—Their chemical structure and biological attributes. Int. J. Biol. Macromol. 2020, 147, 333–349. [Google Scholar] [CrossRef]
- Barros, C.P.; Guimarães, J.T.; Esmerino, E.A.; Duarte, M.C.K.; Silva, M.C.; Silva, R.; Ferreira, B.M.; Sant’Ana, A.S.; Freitas, M.Q.; Cruz, A.G. Paraprobiotics and postbiotics: Concepts and potential applications in dairy products. Curr. Opin. Food Sci. 2020, 32, 1–8. [Google Scholar] [CrossRef]
- Shafipour Yordshahi, A.; Moradi, M.; Tajik, H.; Molaei, R. Design and preparation of antimicrobial meat wrapping nanopaper with bacterial cellulose and postbiotics of lactic acid bacteria. Int. J. Food Microbiol. 2020, 321, 108561. [Google Scholar] [CrossRef]
- Adefegha, S.A. Functional Foods and Nutraceuticals as Dietary Intervention in Chronic Diseases; Novel Perspectives for Health Promotion and Disease Prevention. J. Diet. Suppl. 2018, 15, 977–1009. [Google Scholar] [CrossRef] [PubMed]
- Al Mijan, M.; Lim, B.O. Diets, functional foods, and nutraceuticals as alternative therapies for inflammatory bowel disease: Present status and future trends. World J. Gastroenterol. 2018, 24, 2673–2685. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Olmedo, I.; Rubio, L.A. Dietary legumes, intestinal microbiota, inflammation and colorectal cancer. J. Funct. Foods 2020, 64, 103707. [Google Scholar] [CrossRef]
- Ricciuto, A.; Sherman, P.M.; Laxer, R.M. Gut microbiota in chronic inflammatory disorders: A focus on pediatric inflammatory bowel diseases and juvenile idiopathic arthritis. Clin. Immunol. 2020, 215, 108415. [Google Scholar] [CrossRef]
- Aggeletopoulou, I.; Konstantakis, C.; Assimakopoulos, S.F.; Triantos, C. The role of the gut microbiota in the treatment of inflammatory bowel diseases. Microb. Pathog. 2019, 137, 103774. [Google Scholar] [CrossRef]
- Gobbetti, M.; Pontonio, E.; Filannino, P.; Rizzello, C.G.; De Angelis, M.; Di Cagno, R. How to improve the gluten-free diet: The state of the art from a food science perspective. Food Res. Int. 2018, 110, 22–32. [Google Scholar] [CrossRef]
- Caio, G.; Ciccocioppo, R.; Zoli, G.; De Giorgio, R.; Volta, U. Therapeutic options for coeliac disease: What else beyond gluten-free diet? Dig. Liver Dis. 2020, 52, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, M.S.; Hsu, W.H.; Wang, J.W.; Wang, Y.K.; Hu, H.M.; Chang, W.K.; Chen, C.Y.; Wu, D.C.; Kuo, F.C.; Su, W.W. Nutritional and dietary strategy in the clinical care of inflammatory bowel disease. J. Formos. Med. Assoc. 2019. [Google Scholar] [CrossRef] [PubMed]
- Balestrieri, P.; Ribolsi, M.; Guarino, M.P.L.; Emerenziani, S.; Altomare, A.; Cicala, M. Nutritional Aspects in Inflammatory Bowel Diseases. Nutrients 2020, 12, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterworth, J.; Los, L. Coeliac disease. Medicine (Baltimore) 2019, 47, 314–319. [Google Scholar] [CrossRef]
- Juhász, A.; Colgrave, M.L.; Howitt, C.A. Developing gluten-free cereals and the role of proteomics in product safety. J. Cereal Sci. 2020, 93, 102932. [Google Scholar] [CrossRef]
- Martínez-Villaluenga, C.; Peñas, E.; Hernández-Ledesma, B. Pseudocereal grains: Nutritional value, health benefits and current applications for the development of gluten-free foods. Food Chem. Toxicol. 2020, 137, 111178. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.; Nunes, F.M.; Rodriguez-Quijano, M.; Carrillo, J.M.; Branlard, G.; Igrejas, G. Next-generation therapies for celiac disease: The gluten-targeted approaches. Trends Food Sci. Technol. 2018, 75, 56–71. [Google Scholar] [CrossRef]
- Scherf, K.A.; Wieser, H.; Koehler, P. Novel approaches for enzymatic gluten degradation to create high-quality gluten-free products. Food Res. Int. 2018, 110, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.M. Microbiological and Lipid Profiles of Broa: Contributions for the Characterization of a Traditional Portuguese Bread, Superior School of Agriculture; University of Lisbon (ISA-UL): Lisbon, Portugal, 2011. [Google Scholar]
- Novotni, D.; Gänzle, M.; Rocha, J.M. Chapter 15. Composition and activity of microbiota in sourdough and their effect on bread quality and safety. In Trends in Wheat and Bread Making; Elsevier-Academic Press: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Ortolan, F.; Steel, C.J. Protein Characteristics that Affect the Quality of Vital Wheat Gluten to be Used in Baking: A Review. Compr. Rev. Food Sci. Food Saf. 2017, 16, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Hardt, N.A.; Boom, R.M.; van der Goot, A.J. Starch facilitates enzymatic wheat gluten hydrolysis. LWT—Food Sci. Technol. 2015, 61, 557–563. [Google Scholar] [CrossRef]
- Sakandar, H.A.; Usman, K.; Imran, M. Isolation and characterization of gluten-degrading Enterococcus mundtii and Wickerhamomyces anomalus, potential probiotic strains from indigenously fermented sourdough (Khamir). LWT—Food Sci. Technol. 2018, 91, 271–277. [Google Scholar] [CrossRef]
- Nutter, J.; Saiz, A.I.; Iurlina, M.O. Microstructural and conformational changes of gluten proteins in wheat-rye sourdough. J. Cereal Sci. 2019, 87, 91–97. [Google Scholar] [CrossRef]
- Nikinmaa, M.; Mattila, O.; Holopainen-Mantila, U.; Heiniö, R.-L.; Nordlund, E. Impact of lactic acid bacteria starter cultures and hydrolytic enzymes on the characteristics of wholegrain crackers. J. Cereal Sci. 2019, 88, 1–8. [Google Scholar] [CrossRef]
- Mora-Villalobos, J.A.; Montero-Zamora, J.; Barboza, N.; Rojas-Garbanzo, C.; Usaga, J.; Redondo-Solano, M.; Schroedter, L.; Olszewska-Widdrat, A.; López-Gómez, J.P. Multi-product lactic acid bacteria fermentations: A review. Fermentation 2020, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Pinto, A.; Barbosa, J.; Albano, H.; Isidro, J.; Teixeira, P. Screening of Bacteriocinogenic Lactic Acid Bacteria and Their Characterization as Potential Probiotics. Microorganisms 2020, 8, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Alba, V.; Lazarte, C.E.; Perez-Rea, D.; Carlsson, N.; Almgren, A.; Bergenståhl, B.; Granfeldt, Y. Fermentation of pseudocereals quinoa, canihua, and amaranth to improve mineral accessibility through degradation of phytate. J. Sci. Food Agric. 2019, 99, 5239–5248. [Google Scholar] [CrossRef] [Green Version]
- Rai, S.; Kaur, A.; Chopra, C.S. Gluten-Free Products for Celiac Susceptible People. Front. Nutr. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Carrizo, S.L.; de Moreno de LeBlanc, A.; LeBlanc, J.G.; Rollán, G.C. Quinoa pasta fermented with lactic acid bacteria prevents nutritional deficiencies in mice. Food Res. Int. 2020, 127, 108735. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Angural, S.; Rana, M.; Puri, N.; Kondepudi, K.K.; Gupta, N. Phytase producing lactic acid bacteria: Cell factories for enhancing micronutrient bioavailability of phytate rich foods. Trends Food Sci. Technol. 2020, 96, 1–12. [Google Scholar] [CrossRef]
- Chiș, M.S.; Păucean, A.; Stan, L.; Suharoschi, R.; Socaci, S.-A.; Man, S.M.; Pop, C.R.; Muste, S. Impact of protein metabolic conversion and volatile derivatives on gluten-free muffins made with quinoa sourdough. CyTA—J. Food 2019, 17, 744–753. [Google Scholar] [CrossRef]
- Rocha, J.M.; Malcata, F.X. Microbial ecology dynamics in Portuguese Broa sourdough. J. Food Qual. 2016, 39, 634–648. [Google Scholar] [CrossRef]
- Rocha, J.M.; Malcata, F.X. Behavior of the complex micro-ecology in maize and rye flour and mother-dough for Broa throughout storage. J. Food Qual. 2016, 39, 218–233. [Google Scholar] [CrossRef] [Green Version]
- Arena, M.P.; Russo, P.; Spano, G.; Capozzi, V. From Microbial Ecology to Innovative Applications in Food Quality Improvements: The Case of Sourdough as a Model Matrix. J—Multidiscip. Sci. J. 2020, 3, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Fan, X.; Guo, H.; Yao, Y.; Ren, G.; Lv, X.; Yang, X. Overexpression of the bioactive lunasin peptide in soybean and evaluation of its anti-inflammatory and anti-cancer activities in vitro. J. Biosci. Bioeng. 2020, 129, 395–404. [Google Scholar] [CrossRef]
- Sadeghi, A.; Ebrahimi, M.; Mortazavi, S.A.; Abedfar, A. Application of the selected antifungal LAB isolate as a protective starter culture in pan whole-wheat sourdough bread. Food Control 2019, 95, 298–307. [Google Scholar] [CrossRef]
- Lynch, K.M.; Coffey, A.; Arendt, E.K. Exopolysaccharide producing lactic acid bacteria: Their techno-functional role and potential application in gluten-free bread products. Food Res. Int. 2018, 110, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Wolter, A.; Hager, A.S.; Zannini, E.; Galle, S.; Gänzle, M.G.; Waters, D.M.; Arendt, E.K. Evaluation of exopolysaccharide producing Weissella cibaria MG1 strain for the production of sourdough from various flours. Food Microbiol. 2014, 37, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Al Kassaa, I.; Rafei, R.; Moukhtar, M.; Zaylaa, M.; Gharsallaoui, A.; Asehraou, A.; El Omari, K.; Shahin, A.; Hamze, M.; Chihib, N.-E. LABiocin database: A new database designed specifically for Lactic Acid Bacteria bacteriocins. Int. J. Antimicrob. Agents 2019, 54, 771–779. [Google Scholar] [CrossRef]
- Birch, J.; Van Calsteren, M.R.; Pérez, S.; Svensson, B. The exopolysaccharide properties and structures database: EPS-DB. Application to bacterial exopolysaccharides. Carbohydr. Polym. 2019, 205, 565–570. [Google Scholar] [CrossRef]
- Paśko, P.; Tyszka-Czochara, M.; Namieśnik, J.; Jastrzębski, Z.; Leontowicz, H.; Drzewiecki, J.; Martinez-Ayala, A.L.; Nemirovski, A.; Barasch, D.; Gorinstein, S. Cytotoxic, antioxidant and binding properties of polyphenols from the selected gluten-free pseudocereals and their by-products: In vitro model. J. Cereal Sci. 2019, 87, 325–333. [Google Scholar] [CrossRef]
- Xing, Q.; Dekker, S.; Kyriakopoulou, K.; Boom, R.M.; Smid, E.J.; Schutyser, M.A.I. Enhanced nutritional value of chickpea protein concentrate by dry separation and solid state fermentation. Innov. Food Sci. Emerg. Technol. 2019, 102269. [Google Scholar] [CrossRef]
- Plessas, S.; Mantzourani, I.; Bekatorou, A. Evaluation of Pediococcus pentosaceus SP2 as Starter Culture on Sourdough Bread Making. Foods 2020, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Calasso, M.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2019, 302, 103–113. [Google Scholar] [CrossRef]
- United Nations. United Nations Transforming Our World: The 2030 Agenda for Sustainable Development, General Assembly Resolution A/RES/70/1; United Nations: New York, NY, USA, 2015. [Google Scholar]
- Collins, C.H.; Lyne, P.M. Collins and Lyne’s Microbiological Methods; Arnold: London, UK, 2004. [Google Scholar]
- Pyar, H.; Peh, K.K. Characterization and identification of Lactobacillus acidophilus using biolog rapid identification system. Int. J. Pharm. Pharm. Sci. 2014, 6, 189–193. [Google Scholar]
- Forouhandeh, H.; Zununi Vahed, S.; Hejazi, M.S.; Nahaei, M.R.; Akbari Dibavar, M. Isolation and phenotype characterization of Lactobacillus species from various dairy products. Curr. Res. Bacteriol. 2010, 3, 84–88. [Google Scholar]
- Thakur, M.; Deshpande, H.W.; Bhate, M.A. Isolation and Identification of Lactic Acid Bacteria and their Exploration in Non-Dairy Probiotic Drink. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1023–1030. [Google Scholar]
- Tashakor, A.; Hosseinzadehdehkordi, M.; Emruzi, Z.; Gholami, D. Isolation and identification of a novel bacterium, Lactobacillus sakei subsp. dgh strain 5, and optimization of growth condition for highest antagonistic activity. Microb. Pathog. 2017, 106, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Bendimerad, N.; Kihal, M.; Berthier, F. Isolation, identification, and technological characterization of wild leuconostocs and lactococci for traditional Raib type milk fermentation. Dairy Sci. Technol. 2012, 92, 249–264. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Kumai, S.; Ogawa, M.; Benno, Y.; Nakase, T. Characterization and identification of Pediococcus species isolated from forage crops and their application for silage preparation. Appl. Environ. Microbiol. 1999, 65, 2901–2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Toit, M.; Huch, M.; Cho, G.-S.; Franz, C.M.A.P. The genus Streptococcus. In Lactic Acid Bacteria; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 457–505. [Google Scholar]
- Sáez, G.D.; Saavedra, L.; Hebert, E.M.; Zárate, G. Identification and biotechnological characterization of lactic acid bacteria isolated from chickpea sourdough in northwestern Argentina. LWT—Food Sci. Technol. 2018, 93, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Salvucci, E.; LeBlanc, J.G.; Pérez, G. Technological properties of Lactic acid bacteria isolated from raw cereal material. LWT—Food Sci. Technol. 2016, 70, 185–191. [Google Scholar] [CrossRef]
- Lepczyńska, M.; Dzika, E. The influence of probiotic bacteria and human gut microorganisms causing opportunistic infections on Blastocystis ST3. Gut Pathog. 2019, 11, 6. [Google Scholar] [CrossRef]
- Iurlina, M.O.; Saiz, A.I.; Fuselli, S.R.; Fritz, R. Prevalence of Bacillus spp. in different food products collected in Argentina. LWT—Food Sci. Technol. 2006, 39, 105–110. [Google Scholar] [CrossRef]
- Ju, J.; Xu, X.; Xie, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Inhibitory effects of cinnamon and clove essential oils on mold growth on baked foods. Food Chem. 2018, 240, 850–855. [Google Scholar] [CrossRef]
- Pappier, U.; Fernández Pinto, V.; Larumbe, G.; Vaamonde, G. Effect of processing for saponin removal on fungal contamination of quinoa seeds (Chenopodium quinoa Willd.). Int. J. Food Microbiol. 2008, 125, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Brandt, M.J.; Schwab, C.; Gänzle, M.G. Propionic acid production by cofermentation of Lactobacillus buchneri and Lactobacillus diolivorans in sourdough. Food Microbiol. 2010, 27, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Marathe, S.K.; Vashistht, M.A.; Prashanth, A.; Parveen, N.; Chakraborty, S.; Nair, S.S. Isolation, partial purification, biochemical characterization and detergent compatibility of alkaline protease produced by Bacillus subtilis, Alcaligenes faecalis and Pseudomonas aeruginosa obtained from sea water samples. J. Genet. Eng. Biotechnol. 2018, 16, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Bachtarzi, N.; Kharroub, K.; Ruas-Madiedo, P. Exopolysaccharide-producing lactic acid bacteria isolated from traditional Algerian dairy products and their application for skim-milk fermentations. LWT—Food Sci. Technol. 2019, 107, 117–124. [Google Scholar] [CrossRef]
- Zhao, D.; Jiang, J.; Du, R.; Guo, S.; Ping, W.; Ling, H.; Ge, J. Purification and characterization of an exopolysaccharide from Leuconostoc lactis L2. Int. J. Biol. Macromol. 2019, 139, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Amao, J.A.; Omojasola, P.F.; Barooah, M.; Folakemi Omojasola, P.; Barooah, M. Isolation and characterization of some exopolysaccharide producing bacteria from cassava peel heaps. Sci. Afr. 2019, 4, 93. [Google Scholar] [CrossRef]
- Du, R.; Qiao, X.; Zhao, F.; Song, Q.; Zhou, Q.; Wang, Y.; Pan, L.; Han, Y.; Zhou, Z. Purification, characterization and antioxidant activity of dextran produced by Leuconostoc pseudomesenteroides from homemade wine. Carbohydr. Polym. 2018, 198, 529–536. [Google Scholar] [CrossRef]
- Merabti, R.; Madec, M.N.; Chuat, V.; Becila, F.Z.; Boussekine, R.; Bekhouche, F.; Valence, F. First Insight into the Technological Features of Lactic Acid Bacteria Isolated from Algerian Fermented Wheat Lemzeiet. Curr. Microbiol. 2019, 76, 1095–1104. [Google Scholar] [CrossRef] [Green Version]
- Fekri, A.; Torbati, M.; Yari Khosrowshahi, A.; Bagherpour Shamloo, H.; Azadmard-Damirchi, S.; Khosrowshahi, A.Y.; Shamloo, H.B.; Azadmard-Damirchi, S. Functional Effects of Phytate-Degrading, Probiotic Lactic Acid Bacteria and Yeast Strains Isolated from Iranian Traditional Sourdough on the Technological and Nutritional Properties of Whole Wheat Bread. Food Chem. 2019, 306, 125620. [Google Scholar] [CrossRef]
- Cizeikiene, D.; Juodeikiene, G.; Paskevicius, A.; Bartkiene, E. Antimicrobial activity of lactic acid bacteria against pathogenic and spoilage microorganism isolated from food and their control in wheat bread. Food Control 2013, 31, 539–545. [Google Scholar] [CrossRef]
- Garcia-Cela, E.; Gari Sanchez, F.J.; Sulyok, M.; Verheecke-Vaessen, C.; Medina, A.; Krska, R.; Magan, N. Carbon dioxide production as an indicator of Aspergillus flavus colonisation and aflatoxins/cyclopiazonic acid contamination in shelled peanuts stored under different interacting abiotic factors. Fungal Biol. 2020, 124, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Verón, H.E.; Di Risio, H.D.; Isla, M.I.; Torres, S. Isolation and selection of potential probiotic lactic acid bacteria from Opuntia ficus-indica fruits that grow in Northwest Argentina. LWT—Food Sci. Technol. 2017, 84, 231–240. [Google Scholar] [CrossRef]
- Ben Taheur, F.; Mansour, C.; Kouidhi, B.; Chaieb, K. Use of lactic acid bacteria for the inhibition of Aspergillus flavus and Aspergillus carbonarius growth and mycotoxin production. Toxicon 2019, 166, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Le Lay, C.; Coton, E.; Le Blay, G.; Chobert, J.M.; Haertlé, T.; Choiset, Y.; Van Long, N.N.; Meslet-Cladière, L.; Mounier, J. Identification and quantification of antifungal compounds produced by lactic acid bacteria and propionibacteria. Int. J. Food Microbiol. 2016, 239, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Todorov, S.D.; Stojanovski, S.; Iliev, I.; Moncheva, P.; Nero, L.A.; Ivanova, I.V. Technology and safety assessment for lactic acid bacteria isolated from traditional Bulgarian fermented meat product “lukanka”. Braz. J. Microbiol. 2017, 48, 576–586. [Google Scholar] [CrossRef]
- Omedi, J.O.; Huang, W.; Zheng, J. Effect of sourdough lactic acid bacteria fermentation on phenolic acid release and antifungal activity in pitaya fruit substrate. LWT—Food Sci. Technol. 2019, 111, 309–317. [Google Scholar] [CrossRef]
- Zannini, E.; Waters, D.M.; Coffey, A.; Arendt, E.K. Production, properties, and industrial food application of lactic acid bacteria-derived exopolysaccharides. Appl. Microbiol. Biotechnol. 2016, 100, 1121–1135. [Google Scholar] [CrossRef]
- Taroub, B.; Salma, L.; Manel, Z.; Ouzari, H.I.; Hamdi, Z.; Moktar, H. Isolation of lactic acid bacteria from grape fruit: Antifungal activities, probiotic properties, and in vitro detoxification of ochratoxin A. Ann. Microbiol. 2019, 69, 17–27. [Google Scholar] [CrossRef]
- Bartkiene, E.; Lele, V.; Ruzauskas, M.; Domig, K.J.; Starkute, V.; Zavistanaviciute, P.; Bartkevics, V.; Pugajeva, I.; Klupsaite, D.; Juodeikiene, G.; et al. Lactic Acid Bacteria Isolation from Spontaneous Sourdough and Their Characterization Including Antimicrobial and Antifungal Properties Evaluation. Microorganisms 2019, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Ogunsakin, A.O.; Vanajakshi, V.; Anu-Appaiah, K.A.; Vijayendra, S.V.N.; Walde, S.G.; Banwo, K.; Sanni, A.I.; Prabhasankar, P. Evaluation of functionally important lactic acid bacteria and yeasts from Nigerian sorghum as starter cultures for gluten-free sourdough preparation. LWT—Food Sci. Technol. 2017, 82, 326–334. [Google Scholar] [CrossRef]
- Liu, A.; Jia, Y.; Zhao, L.; Gao, Y.; Liu, G.; Chen, Y.; Zhao, G.; Xu, L.; Shen, L.; Liu, Y.; et al. Diversity of isolated lactic acid bacteria in Ya’an sourdoughs and evaluation of their exopolysaccharide production characteristics. LWT—Food Sci. Technol. 2018, 95, 17–22. [Google Scholar] [CrossRef]
- Rogalski, E.; Vogel, R.F.; Ehrmann, M.A. Monitoring of Lactobacillus sanfranciscensis strains during wheat and rye sourdough fermentations by CRISPR locus length polymorphism PCR. Int. J. Food Microbiol. 2020, 316, 108475. [Google Scholar] [CrossRef] [PubMed]
- Tkhruni, F.N.; Aghajanyan, A.E.; Balabekyan, T.R.; Khachatryan, T.V.; Karapetyan, K.J. Characteristic of Bacteriocins of Lactobacillus rhamnosus BTK 20-12 Potential Probiotic Strain. Probiotics Antimicrob. Proteins 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Domingos-Lopes, M.F.P.; Stanton, C.; Ross, P.R.; Dapkevicius, M.L.E.; Silva, C.C.G. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, M.; Jiaxin, C.; Luo, Y.; Ye, F.; Jiao, S.; Hu, X.; Zhang, J.; Lü, X. Bacterial diversity in traditional sourdough from different regions in China. LWT—Food Sci. Technol. 2018, 96, 251–259. [Google Scholar] [CrossRef]
- Fujimoto, A.; Ito, K.; Narushima, N.; Miyamoto, T. Identification of lactic acid bacteria and yeasts, and characterization of food components of sourdoughs used in Japanese bakeries. J. Biosci. Bioeng. 2019, 127, 575–581. [Google Scholar] [CrossRef]
- Adepehin, J.O.; Enujiugha, V.N.; Badejo, A.A.; Young, G.M.; Odeny, D.A.; Wu, F. Bacterial ecology and rheological parameters of multigrain gluten-free sourdoughs. LWT—Food Sci. Technol. 2018, 96, 344–349. [Google Scholar] [CrossRef]
- Gaglio, R.; Alfonzo, A.; Barbera, M.; Franciosi, E.; Francesca, N.; Moschetti, G.; Settanni, L. Persistence of a mixed lactic acid bacterial starter culture during lysine fortification of sourdough breads by addition of pistachio powder. Food Microbiol. 2019, 86, 103349. [Google Scholar] [CrossRef]
- Bottani, M.; Brasca, M.; Ferraretto, A.; Cardone, G.; Casiraghi, M.C.; Lombardi, G.; De Noni, I.; Cattaneo, S.; Silvetti, T. Chemical and nutritional properties of white bread leavened by lactic acid bacteria. J. Funct. Foods 2018, 45, 330–338. [Google Scholar] [CrossRef]
- Rocha, J.M.; Malcata, F.X. Microbiological profile of maize and rye flours, and sourdough used for the manufacture of traditional Portuguese bread. Food Microbiol. 2012, 31, 72–88. [Google Scholar] [CrossRef]
- Rocha, J.M.; Malcata, F.X. On the microbiological profile of traditional Portuguese sourdough. J. Food Prot. 1999, 62, 1416–1429. [Google Scholar] [CrossRef]
- Han, Q.; Kong, B.; Chen, Q.; Sun, F.; Zhang, H. In vitro comparison of probiotic properties of lactic acid bacteria isolated from Harbin dry sausages and selected probiotics. J. Funct. Foods 2017, 32, 391–400. [Google Scholar] [CrossRef]
- Corbo, M.R.; Bevilacqua, A.; Speranza, B.; Gallo, M.; Campaniello, D.; Sinigaglia, M. Selection of wild lactic acid bacteria for sausages: Design of a selection protocol combining statistic tools, technological and functional properties. LWT—Food Sci. Technol. 2017, 81, 144–152. [Google Scholar] [CrossRef]
- Ojha, K.S.; Harrison, S.M.; Brunton, N.P.; Kerry, J.P.; Tiwari, B.K. Statistical approaches to access the effect of Lactobacillus sakei culture and ultrasound frequency on fatty acid profile of beef jerky. J. Food Compos. Anal. 2017, 57, 1–7. [Google Scholar] [CrossRef]
- Belz, M.C.E.E.; Axel, C.; Arendt, E.K.; Lynch, K.M.; Brosnan, B.; Sheehan, E.M.; Coffey, A.; Zannini, E. Improvement of taste and shelf life of yeasted low-salt bread containing functional sourdoughs using Lactobacillus amylovorus DSM 19280 and Weisella cibaria MG1. Int. J. Food Microbiol. 2019, 302, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tao, Y.; Zhang, X.; Shao, S.; Han, Y.; Chu, D.T.; Xie, G.; Ye, X. Metabolic profile of ginkgo kernel juice fermented with lactic aicd bacteria: A potential way to degrade ginkgolic acids and enrich terpene lactones and phenolics. Process Biochem. 2019, 76, 25–33. [Google Scholar] [CrossRef]
- Wu, T.; Taylor, C.; Nebl, T.; Ng, K.; Bennett, L.E. Effects of chemical composition and baking on in vitro digestibility of proteins in breads made from selected gluten-containing and gluten-free flours. Food Chem. 2017, 233, 514–524. [Google Scholar] [CrossRef]
- Boukid, F.; Vittadini, E.; Lusuardi, F.; Ganino, T.; Carini, E.; Morreale, F.; Pellegrini, N. Does cell wall integrity in legumes flours modulate physiochemical quality and in vitro starch hydrolysis of gluten-free bread? J. Funct. Foods 2019, 59, 110–118. [Google Scholar] [CrossRef]
- Yang, H.; Liu, T.; Zhang, G.; He, G. Intraspecific diversity and fermentative properties of Saccharomyces cerevisiae from Chinese traditional sourdough. LWT—Food Sci. Technol. 2020, 124, 109195. [Google Scholar] [CrossRef]
- Liu, T.; Li, Y.; Yang, Y.; Yi, H.; Zhang, L.; He, G. The influence of different lactic acid bacteria on sourdough flavor and a deep insight into sourdough fermentation through RNA sequencing. Food Chem. 2019, 307, 125529. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Yong, S.; Lee, M.J.; Park, S.J.; Yun, Y.R.; Park, S.H.; Lee, M.A. Changes in volatile and non-volatile compounds of model kimchi through fermentation by lactic acid bacteria. LWT—Food Sci. Technol. 2019, 105, 118–126. [Google Scholar] [CrossRef]
- Francesca, N.; Gaglio, R.; Alfonzo, A.; Corona, O.; Moschetti, G.; Settanni, L. Characteristics of sourdoughs and baked pizzas as affected by starter culture inoculums. Int. J. Food Microbiol. 2019, 293, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Van Kerrebroeck, S.; Maes, D.; De Vuyst, L. Sourdoughs as a function of their species diversity and process conditions, a meta-analysis. Trends Food Sci. Technol. 2017, 68, 152–159. [Google Scholar] [CrossRef]
- Corona, O.; Alfonzo, A.; Ventimiglia, G.; Nasca, A.; Francesca, N.; Martorana, A.; Moschetti, G.; Settanni, L. Industrial application of selected lactic acid bacteria isolated from local semolinas for typical sourdough bread production. Food Microbiol. 2016, 59, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Cirrincione, S.; Breuer, Y.; Mangiapane, E.; Mazzoli, R.; Pessione, E. ‘Ropy’ phenotype, exopolysaccharides and metabolism: Study on food isolated potential probiotics LAB. Microbiol. Res. 2018, 214, 137–145. [Google Scholar] [CrossRef]
- Nachtigall, C.; Surber, G.; Herbi, F.; Wefers, D.; Jaros, D.; Rohm, H. Production and molecular structure of heteropolysaccharides from two lactic acid bacteria. Carbohydr. Polym. 2020, 236, 116019. [Google Scholar] [CrossRef]
- Nambiar, R.B.; Sellamuthu, P.S.; Perumal, A.B.; Sadiku, E.R.; Phiri, G.; Jayaramudu, J. Characterization of an exopolysaccharide produced by Lactobacillus plantarum HM47 isolated from human breast milk. Process Biochem. 2018, 73, 15–22. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Jin, M.; Haobin, Z.; Li, Q.; Shao, D.; Jiang, C.; Huang, Q.; Yang, H.; Shi, J.; Hussain, N. Functional characterization and biotechnological potential of exopolysaccharide produced by Lactobacillus rhamnosus strains isolated from human breast milk. LWT—Food Sci. Technol. 2018, 89, 638–647. [Google Scholar] [CrossRef]
- Abid, Y.; Casillo, A.; Gharsallah, H.; Joulak, I.; Lanzetta, R.; Corsaro, M.M.; Attia, H.; Azabou, S. Production and structural characterization of exopolysaccharides from newly isolated probiotic lactic acid bacteria. Int. J. Biol. Macromol. 2018, 108, 719–728. [Google Scholar] [CrossRef]
- Kansandee, W.; Moonmangmee, D.; Moonmangmee, S.; Itsaranuwat, P. Characterization and Bifidobacterium sp. growth stimulation of exopolysaccharide produced by Enterococcus faecalis EJRM152 isolated from human breast milk. Carbohydr. Polym. 2019, 206, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Adebayo-Tayo, B.; Ishola, R.; Oyewunmi, T. Characterization, antioxidant and immunomodulatory potential on exopolysaccharide produced by wild type and mutant Weissella confusa strains. Biotechnol. Rep. 2018, 19, e00271. [Google Scholar] [CrossRef] [PubMed]
- Adebayo-Tayo, B.; Fashogbon, R. In vitro antioxidant, antibacterial, in vivo immunomodulatory, antitumor and hematological potential of exopolysaccharide produced by wild type and mutant Lactobacillus delbureckii subsp. Bulgaricus. Heliyon 2020, 6, e03268. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Lv, S.; Shi, T.T.; Liu, K.; Li, Q.M.; Pan, L.H.; Zha, X.Q.; Luo, J.P. Exopolysaccharides from yoghurt fermented by Lactobacillus paracasei: Production, purification and its binding to sodium caseinate. Food Hydrocoll. 2020, 102, 105635. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, W.; Sun, L.; Sadiq, F.A.; Yang, Y.; Gao, J.; Sang, Y. Preparation screening, production optimization and characterization of exopolysaccharides produced by Lactobacillus sanfranciscensis Ls-1001 isolated from Chinese traditional sourdough. Int. J. Biol. Macromol. 2019, 139, 1295–1303. [Google Scholar] [CrossRef]
- Luz, C.; D’Opazo, V.; Mañes, J.; Meca, G. Antifungal activity and shelf life extension of loaf bread produced with sourdough fermented by Lactobacillus strains. J. Food Process. Preserv. 2019, 43. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.; Zhang, Y.; Yang, W.; Ma, G.; Ma, N.; Hu, Q.; Pei, F. A novel lactic acid bacterium for improving the quality and shelf life of whole wheat bread. Food Control 2020, 109, 106914. [Google Scholar] [CrossRef]
- Dentice Maidana, S.; Aristimuño Ficoseco, C.; Bassi, D.; Cocconcelli, P.S.; Puglisi, E.; Savoy, G.; Vignolo, G.; Fontana, C. Biodiversity and technological-functional potential of lactic acid bacteria isolated from spontaneously fermented chia sourdough. Int. J. Food Microbiol. 2020, 316, 108425. [Google Scholar] [CrossRef]
- Kaya, H.I.; Simsek, O. Characterization of pathogen-specific bacteriocins from lactic acid bacteria and their application within cocktail against pathogens in milk. LWT—Food Sci. Technol. 2019, 115, 108464. [Google Scholar] [CrossRef]
- Salomskiene, J.; Jonkuviene, D.; Macioniene, I.; Abraitiene, A.; Zeime, J.; Repeckiene, J.; Vaiciulyte-Funk, L. Differences in the occurence and efficiency of antimicrobial compounds produced by lactic acid bacteria. Eur. Food Res. Technol. 2019, 245, 569–579. [Google Scholar] [CrossRef]
- Varsha, K.K.; Priya, S.; Devendra, L.; Nampoothiri, K.M. Control of spoilage fungi by protective lactic acid bacteria displaying probiotic properties. Appl. Biochem. Biotechnol. 2014, 172, 3402–3413. [Google Scholar] [CrossRef] [PubMed]
- Gajbhiye, M.; Kapadnis, B. Bio-efficiency of Antifungal Lactic Acid Bacterial Isolates for Pomegranate Fruit Rot Management. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 1477–1488. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Algboory, H.L.; Kadum, H.; Mohammed, N.K.; Saari, N.; Hassan, Z.; Meor Hussin, A.S. Antifungal activity determination for the peptides generated by Lactobacillus plantarum TE10 against Aspergillus flavus in maize seeds. Food Control 2020, 109, 106898. [Google Scholar] [CrossRef]
- Missaoui, J.; Saidane, D.; Mzoughi, R.; Minervini, F. Fermented seeds (“Zgougou”) from aleppo pine as a novel source of potentially probiotic lactic acid bacteria. Microorganisms 2019, 7, 709. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain No. | MIUG Code * | Source of Isolation | Presumptive Genus |
|---|---|---|---|
| 1 | MIUG BL 68 | Buckwheat seeds | Lactobacillus spp. |
| 2 | MIUG BL 21 | Soil | |
| 13 | MIUG BL 57 | Millet seeds | |
| 16 | MIUG BL 53 | White beans | |
| 20 | MIUG BL 71 | Quinoa seeds | |
| 24 | MIUG BL 38 | Chickpea seeds | |
| 29 | MIUG BL 34 | Fermented wheat bran | |
| 30 | MIUG BL 24 | Whey | |
| 31 | MIUG BL 60 | Hemp seeds | |
| 33 | MIUG BL 25 | Wheat flour | Pediococcus spp. |
| 35 | MIUG BL 27 | Soil | Leuconostoc spp. |
| 36 | MIUG BL 39 | Whole wheat flour | Lactobacillus spp. |
| 38 | MIUG BL 59 | Red lentil seeds | |
| 39 | MIUG BL 7 | Soil | Pediococcus spp. |
| 41 | MIUG BL 66 | Chickpea seeds | Leuconostoc spp. |
| 43 | MIUG BL 69 | Buckwheat seeds | Lactobacillus spp. |
| 44 | MIUG BL 32 | Millet seeds | Streptococcus spp. |
| 45 | MIUG BL 19 | Emmental cheese | Leuconostoc spp. |
| 48 | MIUG BL 40 | Sesame seeds |
| Strain No. | Absorbance 1 (λ = 490 nm) | Exopolysaccharides Biosynthesized 2 (mg/mL) |
|---|---|---|
| 1 | 0.847 ± 0.12 | 11.307 |
| 13 | 0.934 ± 0.11 | 17.854 |
| 29 | 0.910 ± 0.51 | 16.092 |
| 33 | 0.939 ± 0.25 | 18.257 |
| 35 | 0.956 ± 0.08 | 19.516 |
| 36 | 1.004 ± 0.24 | 23.193 |
| 39 | 0.974 ± 0.25 | 21.027 |
| 45 | 0.933 ± 0.41 | 17.779 |
| 48 | 0.994 ± 0.36 | 22.387 |
| Strain No. | Chickpea Flour Extract (CFS) | Quinoa Flour Extract (CFS) | Buckwheat Flour Extract (CFS) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A. niger1 | A. flavus1 | Penicillium spp.1 | Bacillus spp.2 | A. niger1 | A. flavus1 | Penicillium spp.1 | Bacillus spp.2 | A. niger1 | A. flavus1 | Penicillium spp.1 | Bacillus spp.2 | |
| 1 | ++ | − | − | +++ | − | − | − | +++ | − | − | − | ++ |
| 2 | − | − | − | +++ | − | − | − | +++ | − | − | − | ++ |
| 13 | + | + | − | ++ | − | − | − | ++ | − | − | − | ++ |
| 16 | − | + | − | +++ | − | − | − | ++ | − | − | − | ++ |
| 20 | + | − | + | ++ | +++ | − | − | +++ | − | − | − | ++ |
| 24 | − | − | + | +++ | +++ | − | − | +++ | +++ | − | − | +++ |
| 29 | − | − | + | ++ | +++ | − | − | +++ | +++ | − | − | +++ |
| 30 | − | − | − | +++ | +++ | − | − | +++ | +++ | − | − | ++ |
| 31 | − | − | − | + | +++ | − | − | ++ | +++ | − | − | ++ |
| 33 | − | − | − | +++ | +++ | − | − | +++ | − | − | − | +++ |
| 35 | − | − | − | ++ | − | − | − | ++ | − | − | − | ++ |
| 36 | − | − | − | +++ | − | − | − | +++ | − | − | − | ++ |
| 38 | − | + | − | ++ | ++ | ++ | − | + | − | − | − | ++ |
| 39 | − | + | − | ++ | − | − | − | +++ | − | − | ++ | +++ |
| 41 | − | − | + | ++ | − | − | − | ++ | +++ | − | ++ | +++ |
| 43 | − | − | ++ | +++ | − | + | − | +++ | +++ | − | ++ | − |
| 44 | − | − | + | ++ | − | − | − | +++ | − | − | ++ | +++ |
| 45 | − | − | + | ++ | − | − | − | +++ | − | − | + | ++ |
| 48 | − | +++ | − | ++ | − | − | + | +++ | +++ | − | ++ | +++ |
| Strain No. | Treatment | Chickpea Flour Extract (a) | Quinoa Flour Extract (b) | Buckwheat Flour Extract (c) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| A. niger | A. flavus | Penicillium spp. | A. niger | A. flavus | Penicillium spp. | A. niger | A. flavus | Penicillium spp. | ||
| Control | 26.00 ± 1.41 | 37.00 ± 0.00 | 24.75 ± 1.06 | 30.50 ± 0.71 | 36.00 ± 0.00 | 25.25 ± 1.06 | 28.50 ± 0.71 | 35.50 ± 0.71 | 27.00 ± 1.41 | |
| 24 | Untreated | 30.00 ± 2.12 | 45.50 ± 0.71 | 23.5 ± 0.71 | 30.25 ± 3.18 | 52.25 ± 6.01 | 34.00 ± 0.00 | 34.00 ± 0.00 | 56.50 ± 12.02 | 30.00 ± 3.54 |
| I, % | n.d. | n.d. | 5.05 | 0.82 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| 48 | Untreated | 31.25 ± 0.35 | 54.00 ± 3.54 | 21.75 ± 5.30 | 32.75 ± 0.35 | 54.25 ± 1.06 | 23.50 ± 8.49 | 32.75 ± 4.60 | 42.25 ± 0.00 | 34.75 ± 3.89 |
| I, % | n.d. | n.d. | 12.12 | n.d. | n.d. | 6.93 | n.d. | n.d. | n.d. | |
| 24 | 60 °C | 27.50 ± 2.12 | 53.00 ± 2.12 | 31.25 ± 6.01 | 33.00 ± 1.41 | 43.75 ± 10.25 | 20,88 ± 0.53 | 32.00 ± 0.00 | 52.00 ± 3.54 | 35,75 ± 6.72 |
| I, % | n.d. | n.d. | n.d. | n.d. | n.d. | 17.33 | n.d. | n.d. | n.d. | |
| 48 | 60 °C | 31.00 ± 0.00 | 47.50 ± 2.12 | 25.00 ± 1.41 | 33.25 ± 1.06 | 51.00 ± 4.24 | 22.08 ± 10.49 | 32.25 ± 2.47 | 53.25 ± 5.30 | 31.75 ± 10.25 |
| I, % | n.d. | n.d. | n.d. | n.d. | n.d. | 12.54 | n.d. | n.d. | n.d. | |
| 24 | 80 °C | 36.08 ± 6.25 | 55.00 ± 0.00 | 38.50 ± 0.71 | 44.75 ± 3.89 | 52.50 ± 0.71 | 18.79 ± 5.96 | 44.50 ± 2.83 | 54.50 ± 0.00 | 24.33 ± 0.94 |
| I, % | n.d. | n.d. | n.d. | n.d. | n.d. | 25.60 | n.d. | n.d. | 9.88 | |
| 48 | 80 °C | 41.00 ± 2.12 | 51.25 ± 1.77 | 22.00 ± 1.41 | 44.50 ± 2.12 | 52.25 ± 1.06 | 31.50 ± 10.61 | 34.88 ± 9.37 | 53.75 ± 0.35 | 19.05 ± 7.71 |
| I, % | n.d. | n.d. | 11.11 | n.d. | n.d. | n.d. | n.d. | n.d. | 29.44 | |
| 24 | 121 °C | 42.75 ± 13.79 | 39.25 ± 7.42 | 26.53 ± 5.27 | 28.30 ± 7.70 | 41.00 ± 3.54 | 41.25 ± 3.18 | 35.00 ± 0.00 | 37.88 ± 9.37 | 36.50 ± 0.00 |
| I, % | n.d. | n.d. | n.d. | 7.20 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| 48 | 121 °C | 41.50 ± 0.00 | 43.50 ± 1.41 | 25.00 ± 0.00 | 40.00 ± 0.00 | 40.75 ± 6.72 | 55.00 ± 0.00 | 35.00 ± 0.00 | 45.50 ± 3.54 | 30.00 ± 0.00 |
| I, % | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| 24 | pH 3.5 | 25.25 ± 1.77 | 46.75 ± 0.35 | 32.25 ± 10.25 | 29.75 ± 0.35 | 39.50 ± 2.12 | 36.00 ± 7.07 | 25.75 ± 0.35 | 42.75 ± 1.77 | 30.00 ± 0.00 |
| I, % | 2.88 | n.d. | n.d. | 2.46 | n.d. | n.d. | 9.65 | n.d. | n.d. | |
| 48 | pH 3.5 | 27.50 ± 3.54 | 44.50 ± 0.71 | 25.00 ± 0.00 | 29.00 ± 0.71 | 46.00 ± 0.71 | 26.00 ± 0.00 | 24.75 ± 0.35 | 46.00 ± 0.71 | 30.00 ± 0.00 |
| I, % | n.d. | n.d. | n.d. | 4.92 | n.d. | n.d. | 13.16 | n.d. | n.d. | |
| 24 | pH 5.5 | 20.75 ± 1.77 | 39.00 ± 0.00 | 36.25 ± 2.47 | 32.00 ± 2.83 | 37.50 ± 0.71 | 32.13 ± 11.14 | 28.75 ± 0.35 | 36.25 ± 0.35 | 34.50 ± 2.12 |
| I, % | 20.19 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| 48 | pH 5.5 | 29.50 ± 9.90 | 40.25 ± 1.06 | 39.50 ± 0.00 | 38.75 ± 3.89 | 36.50 ± 2.12 | 38.00 ± 0.71 | 32.75 ± 1.77 | 36.75 ± 0.35 | 29.75 ± 15.91 |
| I, % | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| 24 | pH 7.5 | 26.75 ± 3.89 | 38.75 ± 2.47 | 29.25 ± 0.35 | 30.00 ± 0.00 | 37.00 ± 0.00 | 25.00 ± 0.00 | 26.50 ± 2.12 | 35.50 ± 0.71 | 33.00 ± 2.12 |
| I, % | n.d. | n.d. | n.d. | 1.64 | n.d. | 0.99 | 7.02 | n.d. | n.d. | |
| 48 | pH 7.5 | 26.00 ± 4.24 | 38.00 ± 2.12 | 30.50 ± 2.83 | 33.00 ± 0.00 | 37.50 ± 0.71 | 26.50 ± 2.12 | 25.50 ± 0.71 | 36.00 ± 1.41 | 34.00 ± 3.54 |
| I, % | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 10.53 | n.d. | n.d. | |
| Strain No. | Treatment | Chickpea Flour Extract | Quinoa Flour Extract | Buckwheat Flour Extract |
|---|---|---|---|---|
| 24 | Untreated | n.d. | 20.00 ± 0.00 | n.d. |
| 48 | Untreated | 16.67 ± 2.31 | 16.00 ± 4.00 | n.d. |
| 24 | 60 °C | n.d. | 17.33 ± 1.53 | n.d. |
| 48 | 60 °C | 25.67 ± 1.15 | 28.67 ± 2.31 | 23.67 ± 1.53 |
| 24 | 80 °C | 23.33 ± 1.53 | 24.00 ± 1.00 | 19.00 ± 1.73 |
| 48 | 80 °C | 17.67 ± 1.15 | 20.33 ± 0.58 | n.d. |
| 24 | 121 °C | 23.33 ± 2.89 | 26.67 ± 3.06 | n.d. |
| 48 | 121 °C | 19.00 + 1.00 | 20.00 ± 1.73 | n.d. |
| 24 | pH 3.5 | 22.33 ± 1.53 | 28.33 ± 2.89 | 25.33 ± 3.21 |
| 48 | pH 3.5 | n.d. | 20.33 ± 0.58 | n.d. |
| 24 | pH 5.5 | 23.67 ± 0.58 | 29.00 ± 1.73 | 21.67 ± 2.89 |
| 48 | pH 5.5 | 21.67 ± 1.53 | 26.33 ± 2.31 | n.d. |
| 24 | pH 7.5 | n.d. | 22.00 ± 1.73 | n.d. |
| 48 | pH 7.5 | n.d. | 21.33 ± 2.31 | n.d. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Păcularu-Burada, B.; Georgescu, L.A.; Vasile, M.A.; Rocha, J.M.; Bahrim, G.-E. Selection of Wild Lactic Acid Bacteria Strains as Promoters of Postbiotics in Gluten-Free Sourdoughs. Microorganisms 2020, 8, 643. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8050643
Păcularu-Burada B, Georgescu LA, Vasile MA, Rocha JM, Bahrim G-E. Selection of Wild Lactic Acid Bacteria Strains as Promoters of Postbiotics in Gluten-Free Sourdoughs. Microorganisms. 2020; 8(5):643. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8050643
Chicago/Turabian StylePăcularu-Burada, Bogdan, Luminița Anca Georgescu, Mihaela Aida Vasile, João Miguel Rocha, and Gabriela-Elena Bahrim. 2020. "Selection of Wild Lactic Acid Bacteria Strains as Promoters of Postbiotics in Gluten-Free Sourdoughs" Microorganisms 8, no. 5: 643. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8050643