Metagenomic Characterization of Gut Microbiota of Carriers of Extended-Spectrum Beta-Lactamase or Carbapenemase-Producing Enterobacteriaceae Following Treatment with Oral Antibiotics and Fecal Microbiota Transplantation: Results from a Multicenter Randomized Trial

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Study Sub-cohort
2.2. Whole Metagenome Shotgun Sequencing and Data Analyses
3. Results
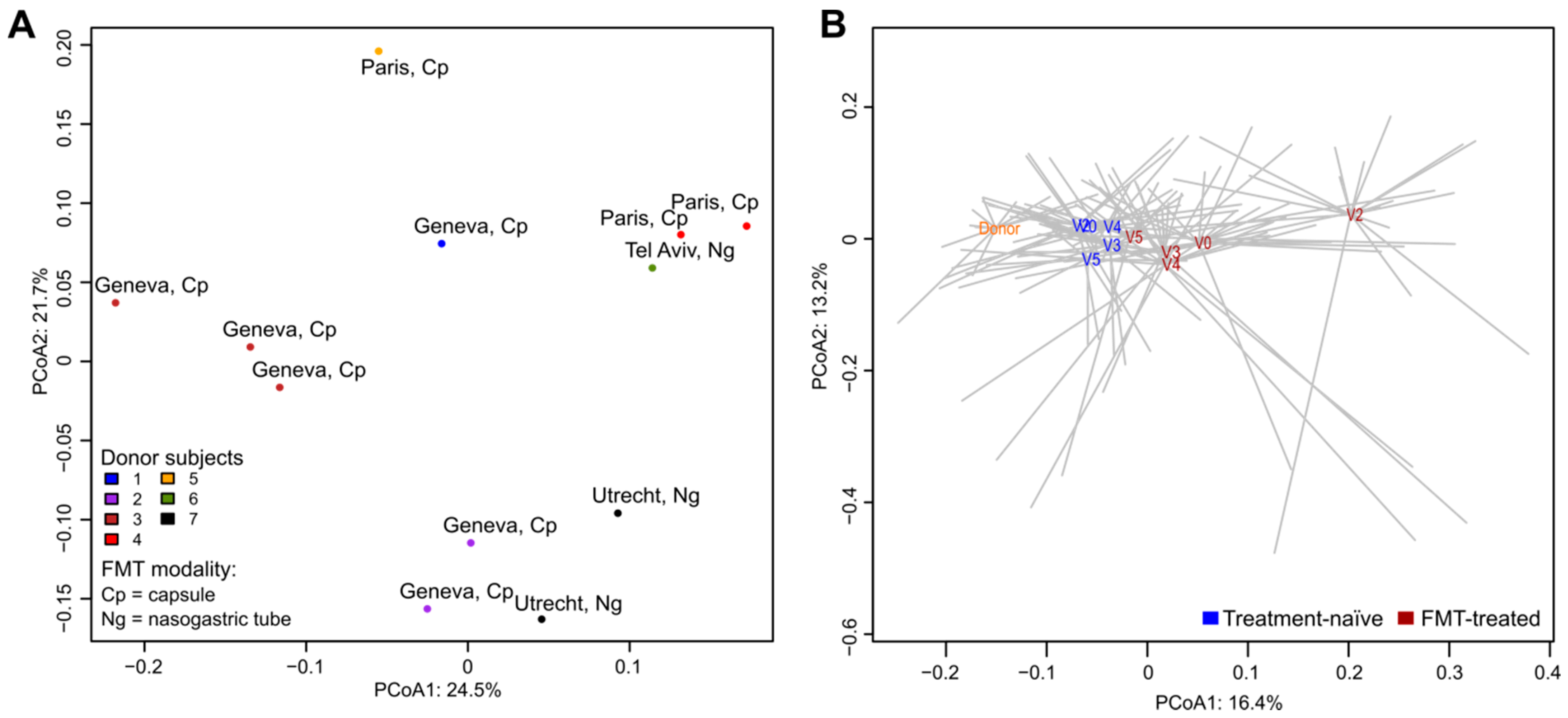
3.1. Donor Microbiota
3.2. Impact of Antibiotic Treatment on Microbiota Profiles and Resistome of FMT-treated Carriers
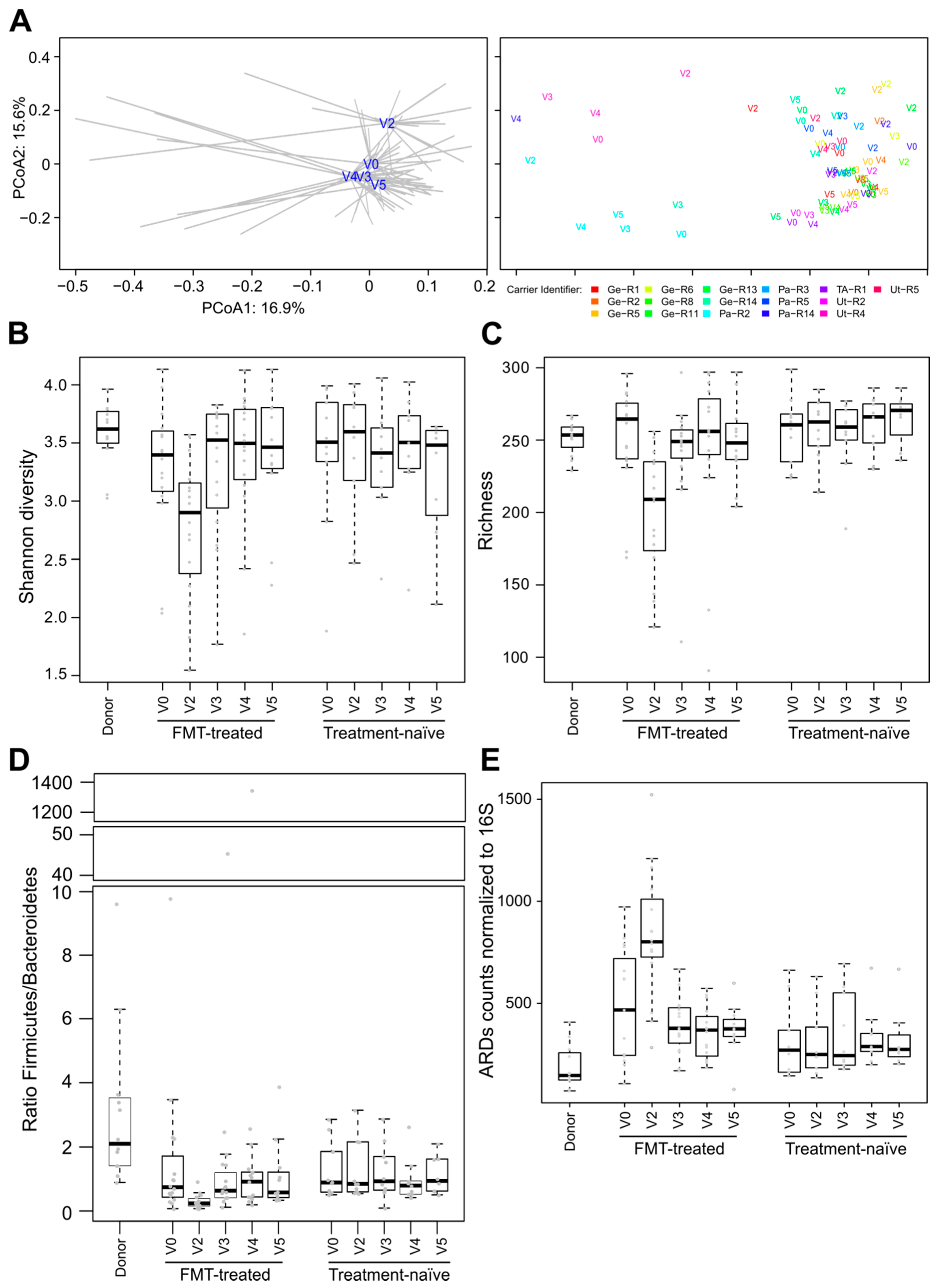
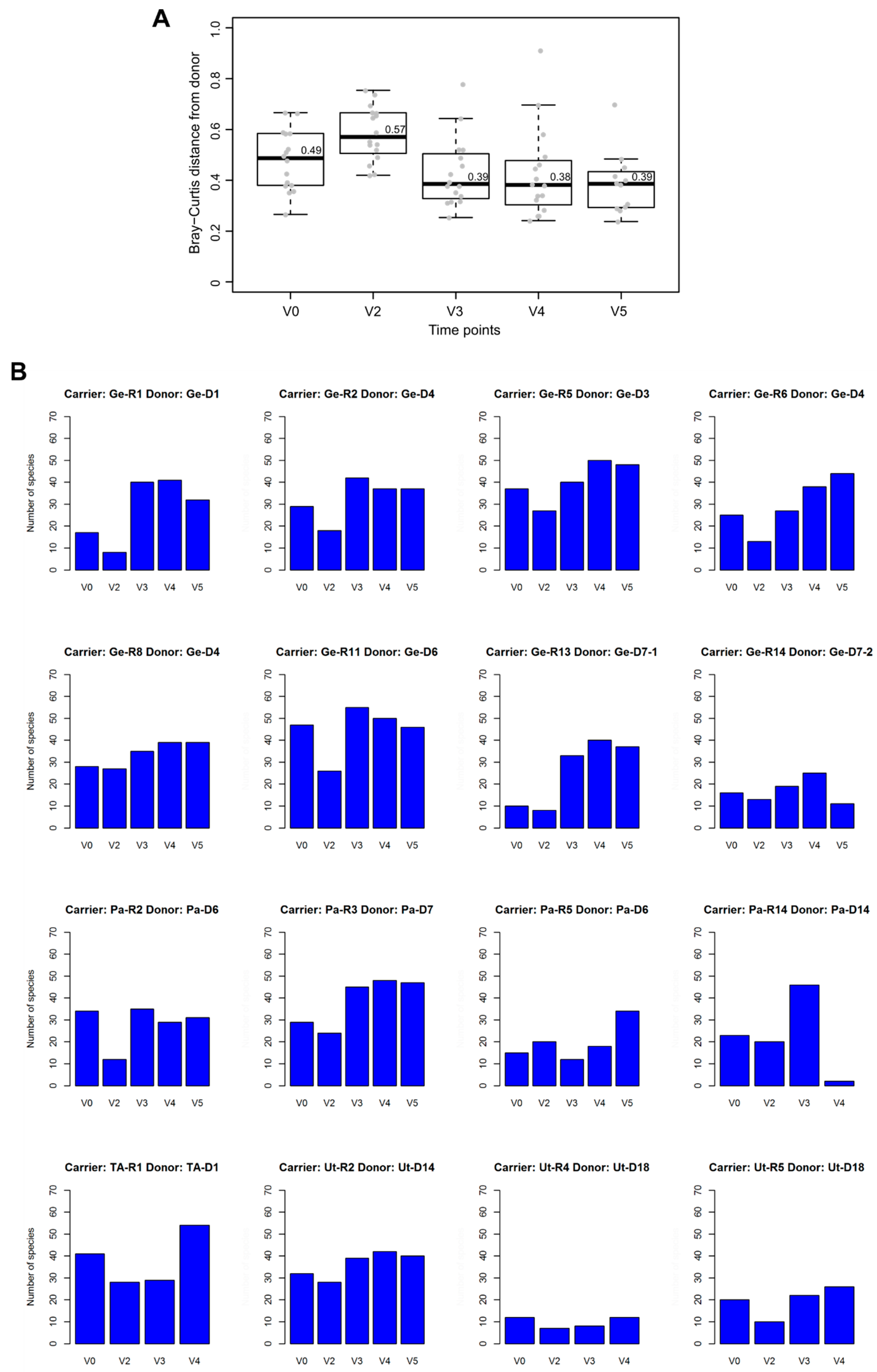
3.3. Post-FMT Microbiota and Its Association with Donor Microbiota
3.4. Comparisons of Microbiota of FMT-Treated and Treatment-Naïve Individuals
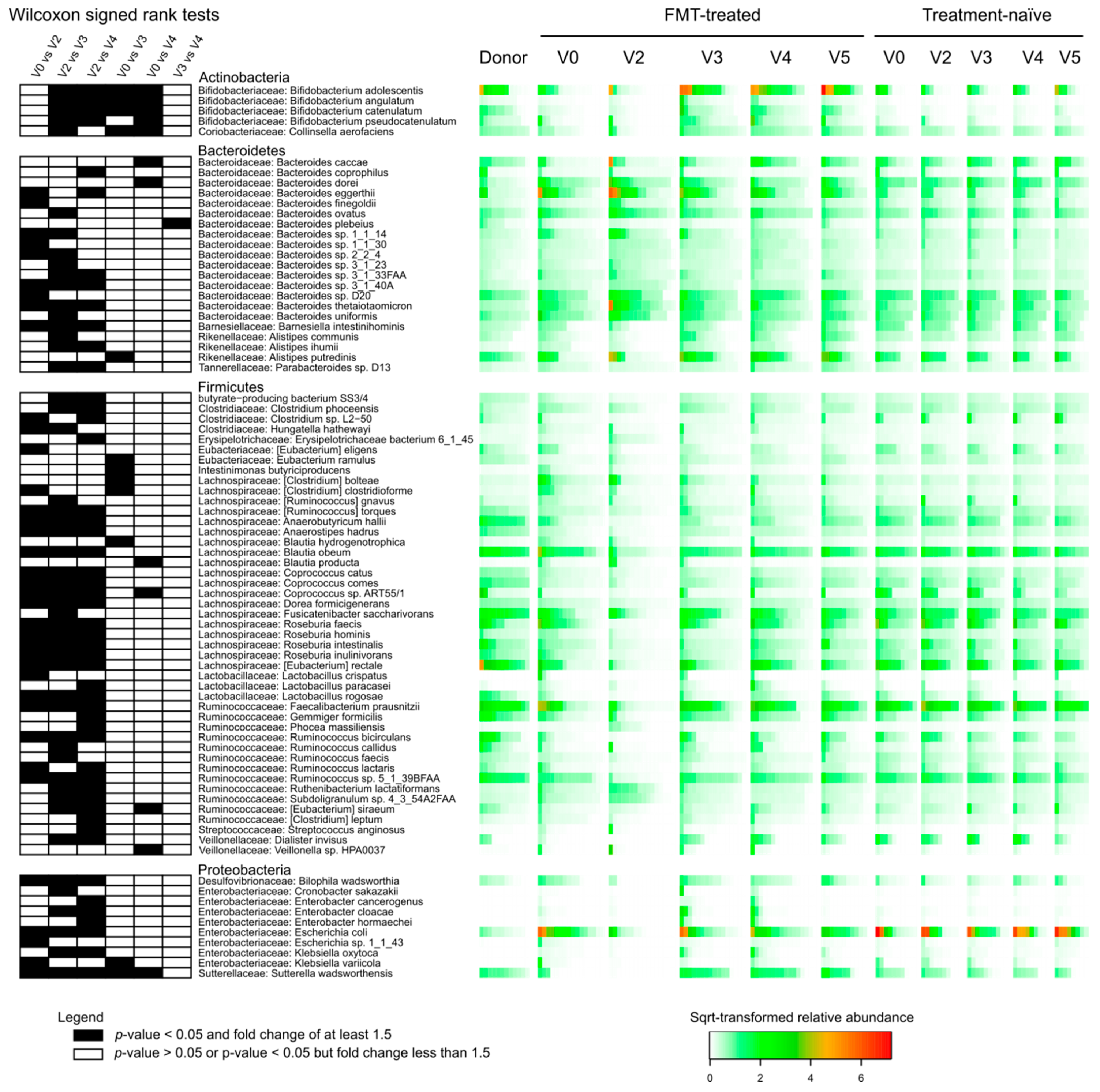
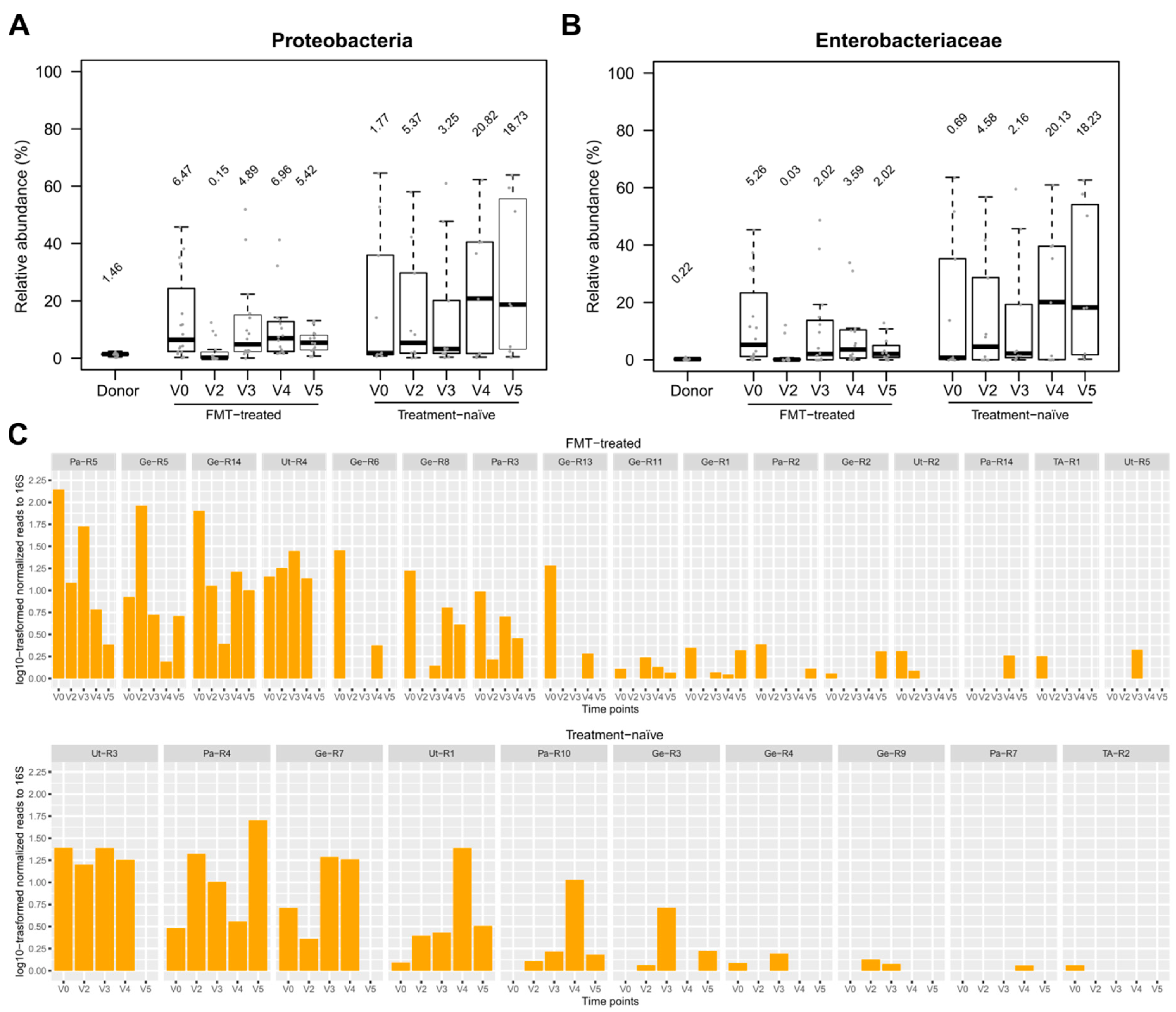
3.5. Effect of FMT/antibiotics on the Abundance of Proteobacteria, Enterobacteriaceae and of Selected Beta-lactam ARDs
3.6. Comparison of Metagenomic Results with R-GNOSIS ESBL-E/CPE Decolonization Outcomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Stewardson, A.J.; Marimuthu, K.; Sengupta, S.; Allignol, A.; El-Bouseary, M.; Carvalho, M.J.; Hassan, B.; Delgado-Ramirez, M.A.; Arora, A.; Bagga, R.; et al. Effect of carbapenem resistance on outcomes of bloodstream infection caused by Enterobacteriaceae in low-income and middle-income countries (PANORAMA): A multinational prospective cohort study. Lancet Infect. Dis. 2019, 19, 601–610. [Google Scholar] [CrossRef]
- Stewardson, A.; Fankhauser, C.; Augelis, G.D.; Rohner, P.; Safran, E.; Schrenzel, J.; Pittet, D.; Harbarth, S. Burden of bloodstream infection caused by extended-spectrum β-lactamase–producing Enterobacteriaceae determined using multistate modeling at a Swiss University Hospital and a nationwide predictive model. Infect. Control Hosp. Epidemiol. 2015, 34, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bert, F.; Larroque, B.; Paugam-Burtz, C.; Dondero, F.; Durand, F.; Marcon, E.; Belghiti, J.; Moreau, R.; Nicolas-Chanoine, M.-H. Pretransplant fecal carriage of extended-spectrum β-lactamase-producing Enterobacteriaceae and infection after liver transplant, France. Emerg. Infect. Dis. 2012, 18, 908–916. [Google Scholar] [CrossRef]
- Rieg, S.; Küpper, M.F.; de With, K.; Serr, A.; Bohnert, J.A.; Kern, W.V. Intestinal decolonization of Enterobacteriaceae producing extended-spectrum β-lactamases (ESBL): A retrospective observational study in patients at risk for infection and a brief review of the literature. BMC Infect. Dis. 2015, 15, 475. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Mazzaferri, F.; de Smet, A.M.; Bragantini, D.; Eggimann, P.; Huttner, B.D.; Kuijper, E.J.; Lucet, J.C.; Mutters, N.T.; Sanguinetti, M.; et al. ESCMID-EUCIC clinical guidelines on decolonization of multidrug-resistant Gram-negative bacteria carriers. Clin. Microbiol. Infect. 2019, 25, 807–817. [Google Scholar] [CrossRef] [Green Version]
- Drekonja, D.; Reich, J.; Gezahegn, S.; Greer, N.; Shaukat, A.; MacDonald, R.; Rutks, I.; Wilt, T.J. Fecal microbiota transplantation for Clostridium difficile infection: A systematic review. Ann. Intern. Med. 2015, 162, 630–638. [Google Scholar] [CrossRef]
- Caballero, S.; Carter, R.; Ke, X.; Sušac, B.; Leiner, I.M.; Kim, G.J.; Miller, L.; Ling, L.; Manova, K.; Pamer, E.G. Distinct but spatially overlapping intestinal niches for vancomycin-resistant Enterococcus faecium and carbapenem-resistant Klebsiella pneumoniae. PLoS Pathog. 2015, 11, e1005132. [Google Scholar] [CrossRef]
- Tavoukjian, V. Faecal microbiota transplantation for the decolonization of antibiotic-resistant bacteria in the gut: A systematic review and meta-analysis. J. Hosp. Infect. 2019, 102, 174–188. [Google Scholar] [CrossRef]
- Catho, G.; Huttner, B.D. Strategies for the eradication of extended-spectrum beta-lactamase or carbapenemase-producing Enterobacteriaceae intestinal carriage. Expert Rev. Anti-Infect. Ther. 2019, 17, 557–569. [Google Scholar] [CrossRef]
- Huttner, B.D.; de Lastours, V.; Wassenberg, M.; Maharshak, N.; Mauris, A.; Galperine, T.; Zanichelli, V.; Kapel, N.; Bellanger, A.; Olearo, F.; et al. A 5-day course of oral antibiotics followed by faecal transplantation to eradicate carriage of multidrug-resistant Enterobacteriaceae: A randomized clinical trial. Clin. Microbiol. Infect. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Wilson, H.; Török, M.E. Extended-spectrum β-lactamase-producing and carbapenemase-producing Enterobacteriaceae. Microb. Genom. 2018, 4, e000197. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Mevius, D.; Ceccarelli, D. A Review of SHV extended-spectrum β-lactamases: Neglected yet ubiquitous. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Galla, S.; Chakraborty, S.; Cheng, X.; Yeo, J.; Mell, B.; Zhang, H.; Mathew, A.V.; Vijay-Kumar, M.; Joe, B. Disparate effects of antibiotics on hypertension. Physiol. Genom. 2018, 50, 837–845. [Google Scholar] [CrossRef]
- Falagas, M.E.; Kasiakou, S.K.; Saravolatz, L.D. Colistin: The revival of polymyxins for the management of multidrug-resistant Gram-negative bacterial infections. Clin. Infect. Dis. 2005, 40, 1333–1341. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Yang, X.; Qin, J.; Lu, N.; Cheng, G.; Wu, N.; Pan, Y.; Li, J.; Zhu, L.; Wang, X.; et al. Metagenome-wide analysis of antibiotic resistance genes in a large cohort of human gut microbiota. Nat. Commun. 2013, 4, 2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forslund, K.; Sunagawa, S.; Kultima, J.R.; Mende, D.R.; Arumugam, M.; Typas, A.; Bork, P. Country-specific antibiotic use practices impact the human gut resistome. Genome Res. 2013, 23, 1163–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vital, M.; Karch, A.; Pieper, D.H. Colonic butyrate-producing communities in humans: An overview using omics data. mSystems 2017, 2, e00130-17. [Google Scholar] [CrossRef] [Green Version]
- Tedelind, S.; Westberg, F.; Kjerrulf, M.; Vidal, A. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: A study with relevance to inflammatory bowel disease. World J. Gastroenterol. 2007, 13, 2826–2832. [Google Scholar] [CrossRef] [PubMed]
- Khokhlova, E.V.; Smeianov, V.V.; Efimov, B.A.; Kafarskaia, L.I.; Pavlova, S.I.; Shkoporov, A.N. Anti-inflammatory properties of intestinal Bifidobacterium strains isolated from healthy infants. Microbiol. Immunol. 2012, 56, 27–39. [Google Scholar] [CrossRef]
- Delcaru, C.; Alexandru, I.; Podgoreanu, P.; Cristea, V.C.; Bleotu, C.; Chifiriuc, M.C.; Bezirtzoglou, E.; Lazar, V. Antagonistic activities of some Bifidobacterium sp. strains isolated from resident infant gastrointestinal microbiota on Gram-negative enteric pathogens. Anaerobe 2016, 39, 39–44. [Google Scholar] [CrossRef]
- Qin, P.; Zou, Y.; Dai, Y.; Luo, G.; Zhang, X.; Xiao, L. Characterization a novel butyric acid-producing bacterium Collinsella aerofaciens subsp. Shenzhenensis subsp. nov. Microorganisms 2019, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Bilinski, J.; Grzesiowski, P.; Sorensen, N.; Madry, K.; Muszynski, J.; Robak, K.; Wroblewska, M.; Dzieciatkowski, T.; Dulny, G.; Dwilewicz-Trojaczek, J.; et al. Fecal microbiota transplantation in patients with blood disorders inhibits gut colonization with antibiotic-resistant bacteria: Results of a prospective, single-center study. Clin. Infect. Dis. 2017, 65, 364–370. [Google Scholar] [CrossRef]
- Ubeda, C.; Bucci, V.; Caballero, S.; Djukovic, A.; Toussaint, N.C.; Equinda, M.; Lipuma, L.; Ling, L.; Gobourne, A.; No, D.; et al. Intestinal microbiota containing Barnesiella species cures vancomycin-resistant Enterococcus faecium colonization. Infect. Immun. 2013, 81, 965. [Google Scholar] [CrossRef] [Green Version]
- Hoyles, L.; McCartney, A.L.; Neve, H.; Gibson, G.R.; Sanderson, J.D.; Heller, K.J.; van Sinderen, D. Characterization of virus-like particles associated with the human faecal and caecal microbiota. Res. Microbiol. 2014, 165, 803–812. [Google Scholar] [CrossRef] [Green Version]
- Shkoporov, A.N.; Clooney, A.G.; Sutton, T.D.S.; Ryan, F.J.; Daly, K.M.; Nolan, J.A.; McDonnell, S.A.; Khokhlova, E.V.; Draper, L.A.; Forde, A.; et al. The human gut virome is highly diverse, stable, and individual specific. Cell Host Microbe 2019, 26, 527–541.e5. [Google Scholar] [CrossRef] [PubMed]
- Grall, N.; Lazarevic, V.; Gaïa, N.; Couffignal, C.; Laouénan, C.; Ilic-Habensus, E.; Wieder, I.; Plesiat, P.; Angebault, C.; Bougnoux, M.E.; et al. Unexpected persistence of extended-spectrum β-lactamase-producing Enterobacteriaceae in the faecal microbiota of hospitalised patients treated with imipenem. Int. J. Antimicrob. Agents 2017, 50, 81–87. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | V0 (n = 16) | V2 (n = 16) | V3 (n = 16) | V4 (n = 16) | V5 (n = 12) | Differentially Abundant |
|---|---|---|---|---|---|---|
| Alistipes putredinis | 4 | 3 | 9 | 9 | 7 | X |
| Alistipes shahii | 5 | 6 | 8 | 7 | 6 | |
| Anaerostipes hadrus | 6 | 1 | 9 | 9 | 7 | X |
| Bacteroides caccae | 4 | 4 | 7 | 11 | 9 | X |
| Bacteroides ovatus | 11 | 12 | 13 | 13 | 10 | X |
| Bacteroides sp. 4_1_36 | 10 | 11 | 11 | 11 | 8 | |
| Bacteroides sp. D20 | 12 | 13 | 14 | 13 | 10 | X |
| Bacteroides stercoris | 6 | 5 | 10 | 10 | 8 | |
| Bacteroides thetaiotaomicron | 9 | 10 | 11 | 10 | 7 | X |
| Bacteroides uniformis | 13 | 14 | 12 | 13 | 11 | X |
| Bacteroides vulgatus | 13 | 14 | 11 | 12 | 10 | |
| Bifidobacterium adolescentis | 4 | 1 | 8 | 9 | 8 | X |
| Bifidobacterium catenulatum | 2 | 0 | 8 | 7 | 5 | X |
| Bifidobacterium longum | 13 | 9 | 15 | 14 | 11 | |
| Blautia obeum | 14 | 9 | 15 | 15 | 11 | X |
| Collinsella aerofaciens | 4 | 2 | 14 | 13 | 10 | X |
| Coprococcus comes | 5 | 2 | 8 | 10 | 8 | X |
| Dorea formicigenerans | 7 | 3 | 11 | 11 | 8 | X |
| Dorea longicatena | 8 | 3 | 11 | 12 | 11 | |
| Escherichia coli | 9 | 3 | 6 | 9 | 8 | X |
| [Eubacterium] rectale | 7 | 4 | 10 | 11 | 10 | X |
| Faecalibacterium prausnitzii | 12 | 4 | 13 | 11 | 11 | X |
| Fusicatenibacter saccharivorans | 9 | 7 | 12 | 12 | 10 | X |
| Gemmiger formicilis | 6 | 2 | 12 | 12 | 11 | X |
| Lachnospiraceae bacterium 7_1_58FAA | 8 | 5 | 7 | 8 | 5 | |
| Odoribacter splanchnicus | 9 | 7 | 8 | 10 | 6 | |
| Roseburia faecis | 11 | 0 | 9 | 10 | 9 | X |
| Roseburia intestinalis | 12 | 1 | 7 | 12 | 9 | X |
| Roseburia inulinivorans | 9 | 0 | 7 | 10 | 8 | X |
| Ruminococcus sp. 5_1_39BFAA | 13 | 1 | 15 | 15 | 11 | X |
| Ruminococcus sp. SR1/5 | 11 | 6 | 12 | 13 | 10 | |
| Sutterella wadsworthensis | 3 | 0 | 11 | 11 | 7 | X |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leo, S.; Lazarevic, V.; Girard, M.; Gaïa, N.; Schrenzel, J.; de Lastours, V.; Fantin, B.; Bonten, M.; Carmeli, Y.; Rondinaud, E.; et al. Metagenomic Characterization of Gut Microbiota of Carriers of Extended-Spectrum Beta-Lactamase or Carbapenemase-Producing Enterobacteriaceae Following Treatment with Oral Antibiotics and Fecal Microbiota Transplantation: Results from a Multicenter Randomized Trial. Microorganisms 2020, 8, 941. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8060941
Leo S, Lazarevic V, Girard M, Gaïa N, Schrenzel J, de Lastours V, Fantin B, Bonten M, Carmeli Y, Rondinaud E, et al. Metagenomic Characterization of Gut Microbiota of Carriers of Extended-Spectrum Beta-Lactamase or Carbapenemase-Producing Enterobacteriaceae Following Treatment with Oral Antibiotics and Fecal Microbiota Transplantation: Results from a Multicenter Randomized Trial. Microorganisms. 2020; 8(6):941. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8060941
Chicago/Turabian StyleLeo, Stefano, Vladimir Lazarevic, Myriam Girard, Nadia Gaïa, Jacques Schrenzel, Victoire de Lastours, Bruno Fantin, Marc Bonten, Yehuda Carmeli, Emilie Rondinaud, and et al. 2020. "Metagenomic Characterization of Gut Microbiota of Carriers of Extended-Spectrum Beta-Lactamase or Carbapenemase-Producing Enterobacteriaceae Following Treatment with Oral Antibiotics and Fecal Microbiota Transplantation: Results from a Multicenter Randomized Trial" Microorganisms 8, no. 6: 941. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8060941