Middle East Respiratory Syndrome Coronavirus (MERS-CoV): State of the Science

 ,
,  , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
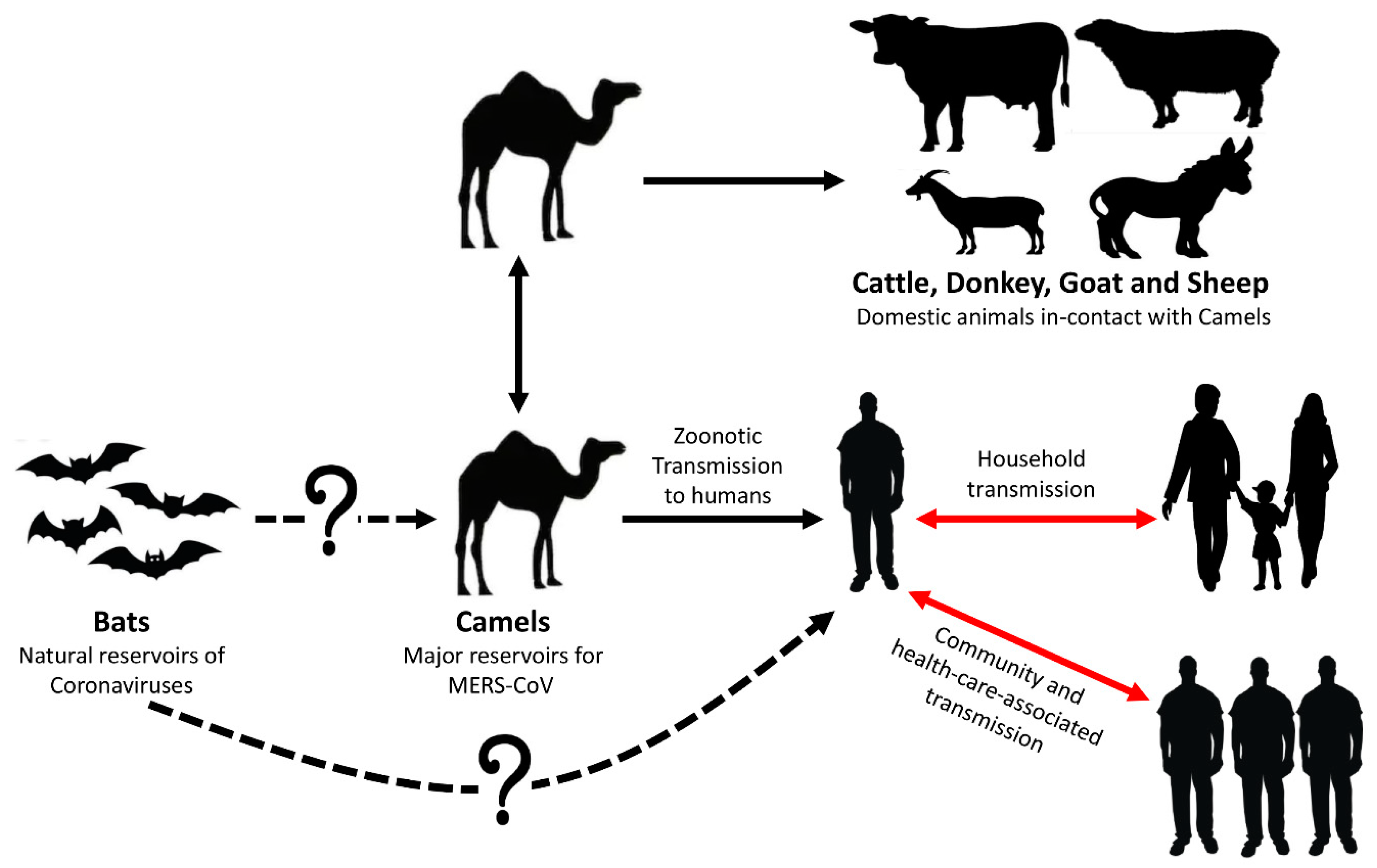
2. MERS-CoV Origin of Transmission
3. Epidemiology of MERS-CoV in Animals
3.1. MERS-CoV Infection in Dromedary Camels
3.2. Sero-Prevalence of MERS-CoV in Domestic Animals
4. Epidemiology of MERS-CoV in Humans
4.1. Distribution Pattern and Human Infections in Middle East
4.2. Distribution Pattern and Human Infections in Africa
4.3. Viral Invasion to Asia, Europe, and America
4.4. Clusters of MERS-CoV Infections
5. Impact of Comorbidities on MERS-CoV Infections in Humans
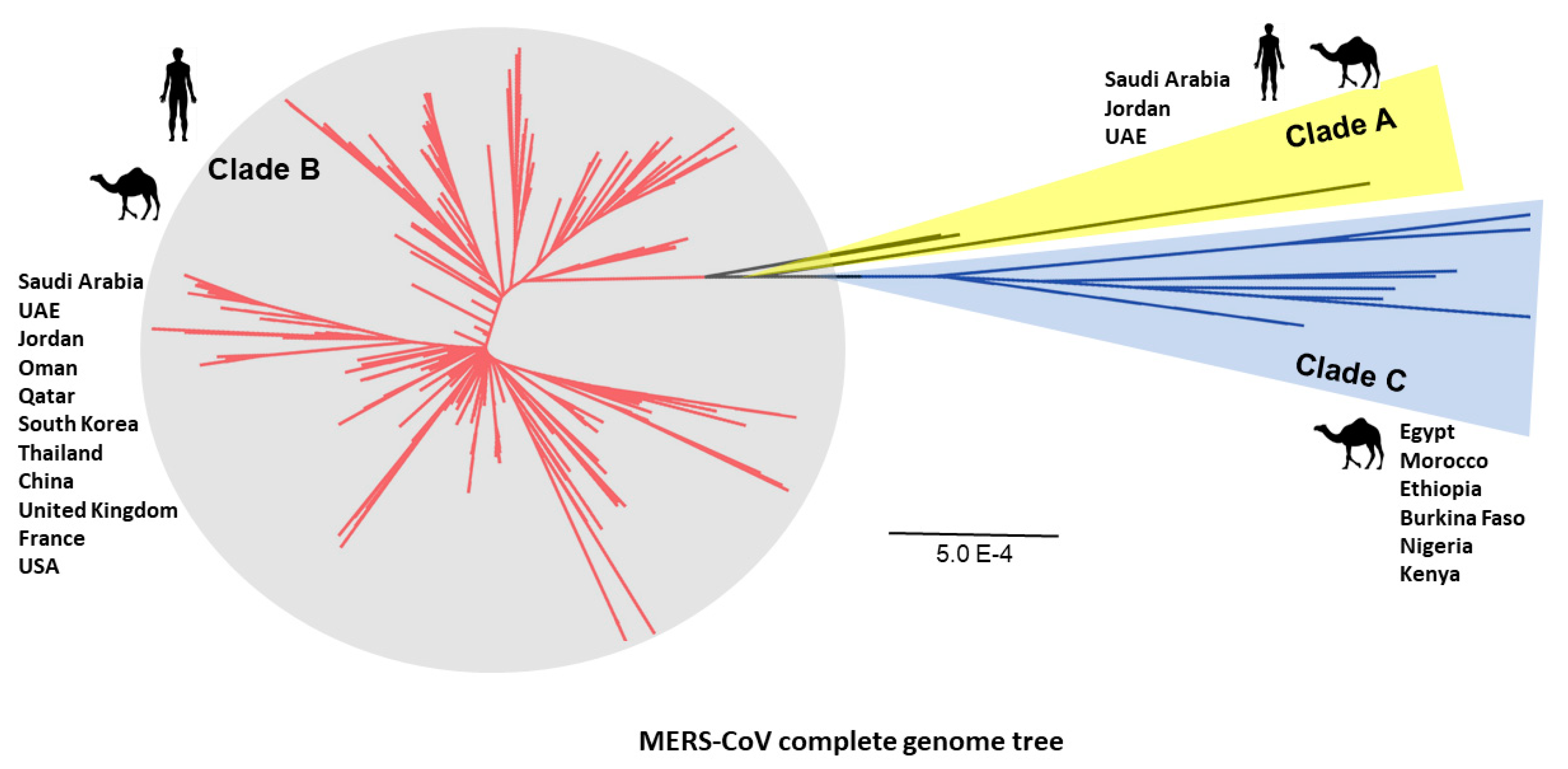
6. Evolution of MERS-CoV
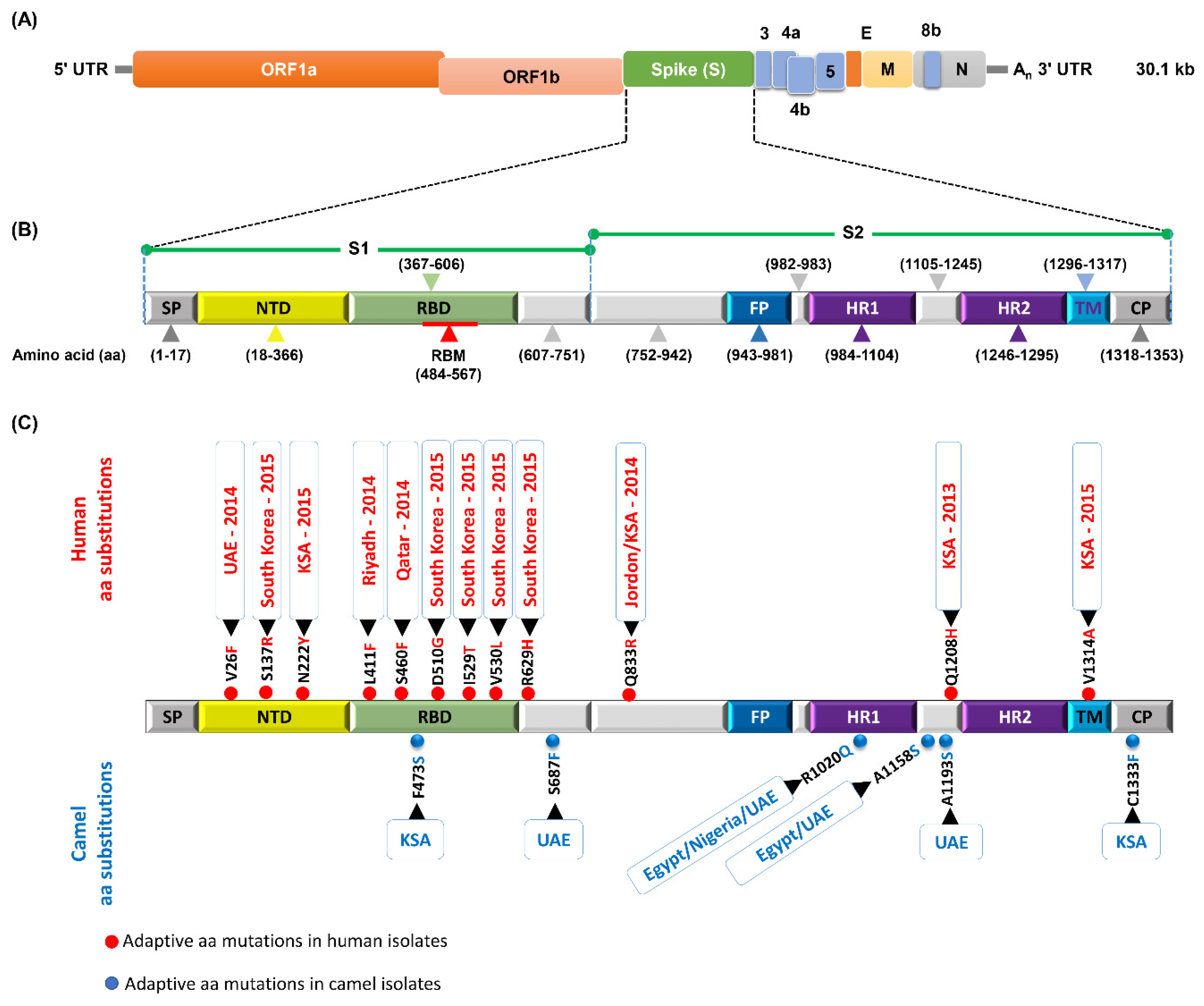
7. Mutation Patterns in Spike Protein of MERS-CoV
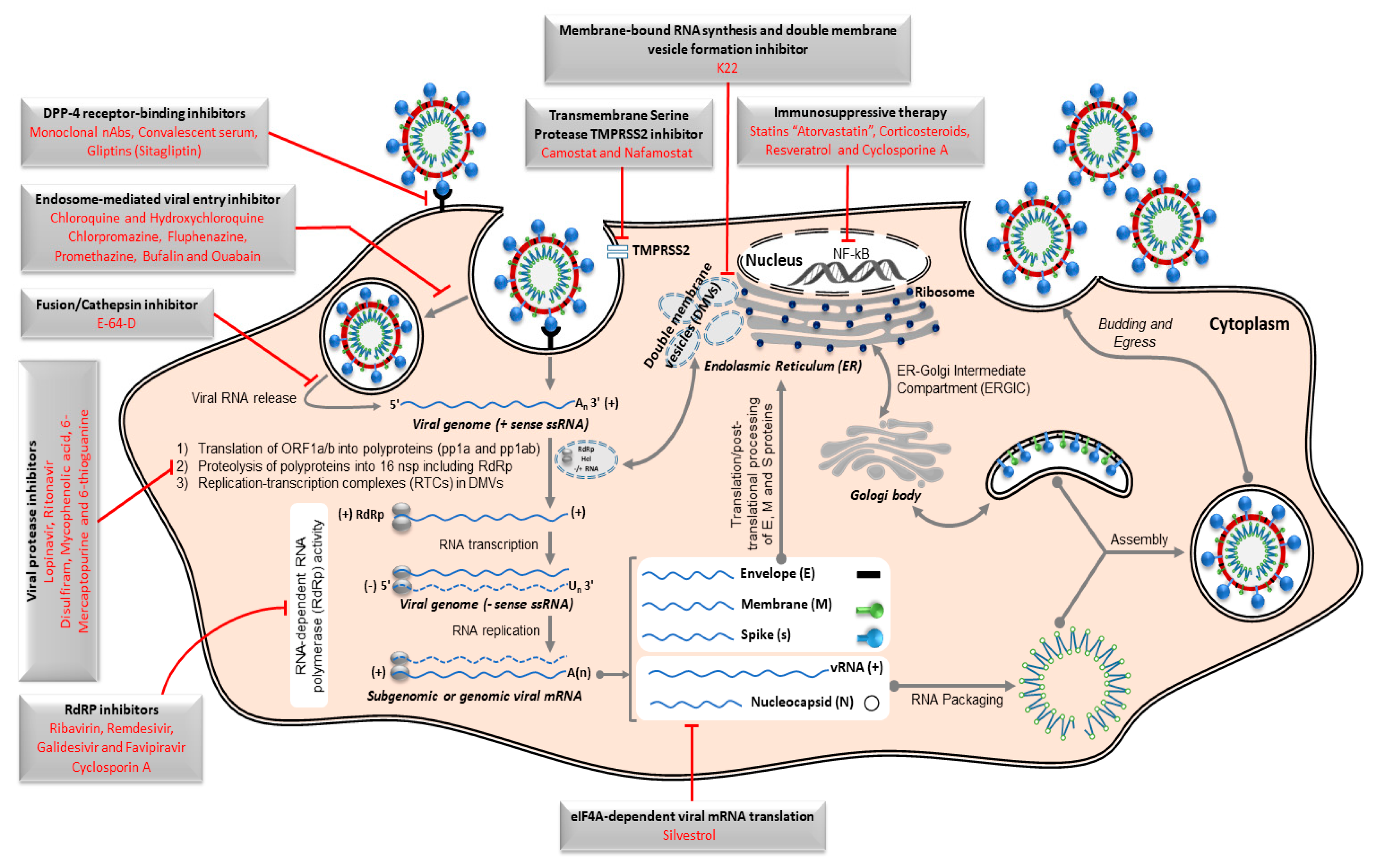
8. Control Approaches for MERS-CoV
8.1. Antiviral Remedy
8.2. Vaccination
8.2.1. Inactivated Vaccine
8.2.2. Live-Attenuated Vaccine
8.2.3. Subunit Vaccine
8.2.4. Virus-Like Particle (VLP) Vaccine
8.2.5. Viral Vector-Recombinant Vaccines
8.2.6. DNA Vaccine
9. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gonzalez, J.M.; Gomez-Puertas, P.; Cavanagh, D.; Gorbalenya, A.E.; Enjuanes, L. A comparative sequence analysis to revise the current taxonomy of the family Coronaviridae. Arch. Virol. 2003, 148, 2207–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ICTV. ICTV Taxonomy History for Coronavirinae Virus Taxonomy: 2011 Release. Available online: http://www.ictvonline.org/virusTaxonomy.asp?taxnode_id=20110624 (accessed on 20 August 2014).
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- FAO. MERS-CoV Situation Update. Available online: http://www.fao.org/ag/againfo/programmes/en/empres/mers/situation_update.html (accessed on 30 June 2020).
- WHO. Middle East respiratory Syndrome Coronavirus (MERS-CoV): MERS Monthly Summary, September 2019. Available online: https://www.who.int/emergencies/mers-cov/en/ (accessed on 30 June 2020).
- Gulland, A. WHO voices concern over rising numbers of MERS-CoV cases. BMJ 2014, 348, g2968. [Google Scholar] [CrossRef] [PubMed]
- Memish, Z.A.; Assiri, A.; Alhakeem, R.; Yezli, S.; Almasri, M.; Zumla, A.; Al-Tawfiq, J.A.; Drosten, C.; Albarrak, A.; Petersen, E. Middle East respiratory syndrome corona virus, MERS-CoV. Conclusions from the 2nd scientific advisory board meeting of the WHO collaborating center for mass gathering medicine, Riyadh. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2014, 24, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Drosten, C.; Gunther, S.; Preiser, W.; van der Werf, S.; Brodt, H.R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.; et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.B.; Opal, S.M. Middle East respiratory syndrome (MERS): A new zoonotic viral pneumonia. Virulence 2014, 5, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Shehata, M.M.; Gomaa, M.R.; Ali, M.A.; Kayali, G. Middle East respiratory syndrome coronavirus: A comprehensive review. Front. Med. 2016, 10, 120–136. [Google Scholar] [CrossRef] [Green Version]
- Cotten, M.; Watson, S.J.; Kellam, P.; Al-Rabeeah, A.A.; Makhdoom, H.Q.; Assiri, A.; Al-Tawfiq, J.A.; Alhakeem, R.F.; Madani, H.; AlRabiah, F.A.; et al. Transmission and evolution of the Middle East respiratory syndrome coronavirus in Saudi Arabia: A descriptive genomic study. Lancet 2013, 382, 1993–2002. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.K.; Poon, L.L.; Gomaa, M.M.; Shehata, M.M.; Perera, R.A.; Abu Zeid, D.; El Rifay, A.S.; Siu, L.Y.; Guan, Y.; Webby, R.J.; et al. MERS coronaviruses in dromedary camels, Egypt. Emerg. Infect. Dis. 2014, 20, 1049–1053. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.K.W.; Hui, K.P.Y.; Perera, R.; Miguel, E.; Niemeyer, D.; Zhao, J.; Channappanavar, R.; Dudas, G.; Oladipo, J.O.; Traore, A.; et al. MERS coronaviruses from camels in Africa exhibit region-dependent genetic diversity. Proc. Natl. Acad. Sci. USA 2018, 115, 3144–3149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckerle, I.; Ehlen, L.; Kallies, R.; Wollny, R.; Corman, V.M.; Cottontail, V.M.; Tschapka, M.; Oppong, S.; Drosten, C.; Muller, M.A. Bat airway epithelial cells: A novel tool for the study of zoonotic viruses. PLoS ONE 2014, 9, e84679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memish, Z.A.; Mishra, N.; Olival, K.J.; Fagbo, S.F.; Kapoor, V.; Epstein, J.H.; Alhakeem, R.; Durosinloun, A.; Al Asmari, M.; Islam, A.; et al. Middle East respiratory syndrome coronavirus in bats, Saudi Arabia. Emerg. Infect. Dis. 2013, 19, 1819–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annan, A.; Baldwin, H.J.; Corman, V.M.; Klose, S.M.; Owusu, M.; Nkrumah, E.E.; Badu, E.K.; Anti, P.; Agbenyega, O.; Meyer, B.; et al. Human betacoronavirus 2c EMC/2012-related viruses in bats, Ghana and Europe. Emerg. Infect. Dis. 2013, 19, 456–459. [Google Scholar] [CrossRef]
- Anthony, S.J.; Ojeda-Flores, R.; Rico-Chavez, O.; Navarrete-Macias, I.; Zambrana-Torrelio, C.M.; Rostal, M.K.; Epstein, J.H.; Tipps, T.; Liang, E.; Sanchez-Leon, M.; et al. Coronaviruses in bats from Mexico. J. Gen. Virol. 2013, 94, 1028–1038. [Google Scholar] [CrossRef]
- Yang, L.; Wu, Z.; Ren, X.; Yang, F.; Zhang, J.; He, G.; Dong, J.; Sun, L.; Zhu, Y.; Zhang, S.; et al. MERS-related betacoronavirus in Vespertilio superans bats, China. Emerg. Infect. Dis. 2014, 20. [Google Scholar] [CrossRef] [Green Version]
- Azhar, E.I.; Hashem, A.M.; El-Kafrawy, S.A.; Sohrab, S.S.; Aburizaiza, A.S.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Jamjoom, G.A.; Madani, T.A. Detection of the middle East respiratory syndrome coronavirus genome in an air sample originating from a camel barn owned by an infected patient. mBio 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Corman, V.M.; Jores, J.; Meyer, B.; Younan, M.; Liljander, A.; Said, M.Y.; Gluecks, I.; Lattwein, E.; Bosch, B.J.; Drexler, J.F.; et al. Antibodies against MERS coronavirus in dromedary camels, Kenya, 1992–2013. Emerg. Infect. Dis. 2014, 20. [Google Scholar] [CrossRef]
- Hemida, M.G.; Perera, R.A.; Wang, P.; Alhammadi, M.A.; Siu, L.Y.; Li, M.; Poon, L.L.; Saif, L.; Alnaeem, A.; Peiris, M. Middle East Respiratory Syndrome (MERS) coronavirus seroprevalence in domestic livestock in Saudi Arabia, 2010 to 2013. Eurosurveillance 2013, 18, 20659. [Google Scholar] [CrossRef]
- Reusken, C.B.; Messadi, L.; Feyisa, A.; Ularamu, H.; Godeke, G.J.; Danmarwa, A.; Dawo, F.; Jemli, M.; Melaku, S.; Shamaki, D.; et al. Geographic distribution of MERS coronavirus among dromedary camels, Africa. Emerg. Infect. Dis. 2014, 20, 1370–1374. [Google Scholar] [CrossRef] [Green Version]
- Nowotny, N.; Kolodziejek, J. Middle East respiratory syndrome coronavirus (MERS-CoV) in dromedary camels, Oman, 2013. Eurosurveillance 2014, 19, 20781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, B.; Muller, M.A.; Corman, V.M.; Reusken, C.B.; Ritz, D.; Godeke, G.J.; Lattwein, E.; Kallies, S.; Siemens, A.; van Beek, J.; et al. Antibodies against MERS coronavirus in dromedary camels, United Arab Emirates, 2003 and 2013. Emerg. Infect. Dis. 2014, 20, 552–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemida, M.; Perera, R.; Al Jassim, R.; Kayali, G.; Siu, L.; Wang, P.; Chu, K.; Perlman, S.; Ali, M.; Alnaeem, A.; et al. Seroepidemiology of Middle East respiratory syndrome (MERS) coronavirus in Saudi Arabia (1993) and Australia (2014) and characterisation of assay specificity. Eurosurveillance 2014, 19, 20828. [Google Scholar] [CrossRef] [PubMed]
- Raj, V.S.; Farag, E.A.; Reusken, C.B.; Lamers, M.M.; Pas, S.D.; Voermans, J.; Smits, S.L.; Osterhaus, A.D.; Al-Mawlawi, N.; Al-Romaihi, H.E.; et al. Isolation of MERS coronavirus from a dromedary camel, Qatar, 2014. Emerg. Infect. Dis. 2014, 20. [Google Scholar] [CrossRef]
- Hemida, M.G.; Chu, D.K.; Poon, L.L.; Perera, R.A.; Alhammadi, M.A.; Ng, H.Y.; Siu, L.Y.; Guan, Y.; Alnaeem, A.; Peiris, M. MERS coronavirus in dromedary camel herd, Saudi Arabia. Emerg. Infect. Dis. 2014, 20. [Google Scholar] [CrossRef] [Green Version]
- Perera, R.A.; Wang, P.; Gomaa, M.R.; El-Shesheny, R.; Kandeil, A.; Bagato, O.; Siu, L.Y.; Shehata, M.M.; Kayed, A.S.; Moatasim, Y.; et al. Seroepidemiology for MERS coronavirus using microneutralisation and pseudoparticle virus neutralisation assays reveal a high prevalence of antibody in dromedary camels in Egypt, June 2013. Eurosurveillance 2013, 18, 20574. [Google Scholar] [CrossRef]
- Reusken, C.B.; Ababneh, M.; Raj, V.S.; Meyer, B.; Eljarah, A.; Abutarbush, S.; Godeke, G.J.; Bestebroer, T.M.; Zutt, I.; Muller, M.A.; et al. Middle East Respiratory Syndrome coronavirus (MERS-CoV) serology in major livestock species in an affected region in Jordan, June to September 2013. Eurosurveillance 2013, 18, 20662. [Google Scholar] [CrossRef]
- Harcourt, J.L.; Rudoler, N.; Tamin, A.; Leshem, E.; Rasis, M.; Giladi, M.; Haynes, L.M. The prevalence of Middle East respiratory syndrome coronavirus (MERS-CoV) antibodies in dromedary camels in Israel. Zoonoses Public Health 2018. [Google Scholar] [CrossRef]
- Gardner, E.G.; Kiambi, S.; Sitawa, R.; Kelton, D.; Kimutai, J.; Poljak, Z.; Tadesse, Z.; Von Dobschuetz, S.; Wiersma, L.; Greer, A.L. Force of infection of Middle East respiratory syndrome in dromedary camels in Kenya. Epidemiol. Infect. 2019, 147, e275. [Google Scholar] [CrossRef] [Green Version]
- Cha, R.-H.; Joh, J.-S.; Jeong, I.; Lee, J.Y.; Shin, H.-S.; Kim, G.; Kim, Y.; Critical Care Team of National Medical Center. Renal complications and their prognosis in Korean patients with Middle East respiratory syndrome-coronavirus from the central MERS-CoV designated hospital. J. Korean Med. Sci. 2015, 30, 1807–1814. [Google Scholar] [CrossRef]
- Corman, V.M.; Ithete, N.L.; Richards, L.R.; Schoeman, M.C.; Preiser, W.; Drosten, C.; Drexler, J.F. Rooting the phylogenetic tree of middle East respiratory syndrome coronavirus by characterization of a conspecific virus from an African bat. J. Virol. 2014, 88, 11297–11303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memish, Z.A.; Zumla, A.I.; Assiri, A. Middle East respiratory syndrome coronavirus infections in health care workers. N. Engl. J. Med. 2013, 369, 884–886. [Google Scholar] [CrossRef] [PubMed]
- van Boheemen, S.; de Graaf, M.; Lauber, C.; Bestebroer, T.M.; Raj, V.S.; Zaki, A.M.; Osterhaus, A.D.; Haagmans, B.L.; Gorbalenya, A.E.; Snijder, E.J.; et al. Genomic characterization of a newly discovered coronavirus associated with acute respiratory distress syndrome in humans. mBio 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.C.; Lau, S.K.; Lam, C.S.; Lau, C.C.; Tsang, A.K.; Lau, J.H.; Bai, R.; Teng, J.L.; Tsang, C.C.; Wang, M.; et al. Discovery of seven novel Mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gierer, S.; Hofmann-Winkler, H.; Albuali, W.H.; Bertram, S.; Al-Rubaish, A.M.; Yousef, A.A.; Al-Nafaie, A.N.; Al-Ali, A.K.; Obeid, O.E.; Alkharsah, K.R.; et al. Lack of MERS coronavirus neutralizing antibodies in humans, eastern province, Saudi Arabia. Emerg. Infect. Dis. 2013, 19, 2034–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aburizaiza, A.S.; Mattes, F.M.; Azhar, E.I.; Hassan, A.M.; Memish, Z.A.; Muth, D.; Meyer, B.; Lattwein, E.; Muller, M.A.; Drosten, C. Investigation of anti-middle East respiratory syndrome antibodies in blood donors and slaughterhouse workers in Jeddah and Makkah, Saudi Arabia, fall 2012. J. Infect. Dis. 2014, 209, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.A.; Meyer, B.; Corman, V.M.; Al-Masri, M.; Turkestani, A.; Ritz, D.; Sieberg, A.; Aldabbagh, S.; Bosch, B.J.; Lattwein, E.; et al. Presence of Middle East respiratory syndrome coronavirus antibodies in Saudi Arabia: A nationwide, cross-sectional, serological study. Lancet Infect. Dis. 2015, 15, 629. [Google Scholar] [CrossRef] [Green Version]
- Reusken, C.B.; Farag, E.A.; Haagmans, B.L.; Mohran, K.A.; Godeke, G.J.t.; Raj, S.; Alhajri, F.; Al-Marri, S.A.; Al-Romaihi, H.E.; Al-Thani, M.; et al. Occupational exposure to dromedaries and risk for MERS-CoV infection, Qatar, 2013–2014. Emerg. Infect. Dis. 2015, 21, 1422–1425. [Google Scholar] [CrossRef]
- Memish, Z.A.; Almasri, M.; Turkestani, A.; Al-Shangiti, A.M.; Yezli, S. Etiology of severe community-acquired pneumonia during the 2013 Hajj-part of the MERS-CoV surveillance program. Int. J. Infect. Dis. Ijid Off. Publ. Int. Soc. Infect. Dis. 2014, 25, 186–190. [Google Scholar] [CrossRef] [Green Version]
- Kandeil, A.; Gomaa, M.; Shehata, M.; El-Taweel, A.; Kayed, A.E.; Abiadh, A.; Jrijer, J.; Moatasim, Y.; Kutkat, O.; Bagato, O.; et al. Middle East respiratory syndrome coronavirus infection in non-camelid domestic mammals. Emerg. Microbes Infect. 2019, 8, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Gikonyo, S.; Kimani, T.; Matere, J.; Kimutai, J.; Kiambi, S.G.; Bitek, A.O.; Juma Ngeiywa, K.J.Z.; Makonnen, Y.J.; Tripodi, A.; Morzaria, S.; et al. Mapping potential amplification and transmission hotspots for MERS-CoV, Kenya. EcoHealth 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.A.; Shehata, M.M.; Gomaa, M.R.; Kandeil, A.; El-Shesheny, R.; Kayed, A.S.; El-Taweel, A.N.; Atea, M.; Hassan, N.; Bagato, O.; et al. Systematic, active surveillance for Middle East respiratory syndrome coronavirus in camels in Egypt. Emerg. Microbes Infect. 2017, 6, e1. [Google Scholar] [CrossRef] [Green Version]
- Reusken, C.B.; Haagmans, B.L.; Muller, M.A.; Gutierrez, C.; Godeke, G.J.; Meyer, B.; Muth, D.; Raj, V.S.; Smits-De Vries, L.; Corman, V.M.; et al. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: A comparative serological study. Lancet Infect. Dis. 2013, 13, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Reusken, C.; Farag, E.; Jonges, M.; Godeke, G.; El-Sayed, A.; Pas, S.; Raj, V.; Mohran, K.; Moussa, H.; Ghobashy, H.; et al. Middle East respiratory syndrome coronavirus (MERS-CoV) RNA and neutralising antibodies in milk collected according to local customs from dromedary camels, Qatar, April 2014. Eurosurveillance 2014, 19, 20829. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.A.; Corman, V.M.; Jores, J.; Meyer, B.; Younan, M.; Liljander, A.; Bosch, B.J.; Lattwein, E.; Hilali, M.; Musa, B.E.; et al. MERS coronavirus neutralizing antibodies in camels, Eastern Africa, 1983–1997. Emerg. Infect. Dis. 2014, 20, 2093–2095. [Google Scholar] [CrossRef] [PubMed]
- Azhar, E.I.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for camel-to-human transmission of MERS coronavirus. N. Engl. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef] [PubMed]
- Haagmans, B.L.; Al Dhahiry, S.H.; Reusken, C.B.; Raj, V.S.; Galiano, M.; Myers, R.; Godeke, G.J.; Jonges, M.; Farag, E.; Diab, A.; et al. Middle East respiratory syndrome coronavirus in dromedary camels: An outbreak investigation. Lancet Infect. Dis. 2014, 14, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Alagaili, A.N.; Briese, T.; Mishra, N.; Kapoor, V.; Sameroff, S.C.; Burbelo, P.D.; de Wit, E.; Munster, V.J.; Hensley, L.E.; Zalmout, I.S.; et al. Middle East respiratory syndrome coronavirus infection in dromedary camels in Saudi Arabia. mBio 2014, 5, e00884-14. [Google Scholar] [CrossRef] [Green Version]
- Adney, D.R.; van Doremalen, N.; Brown, V.R.; Bushmaker, T.; Scott, D.; de Wit, E.; Bowen, R.A.; Munster, V.J. Replication and shedding of MERS-CoV in upper respiratory tract of inoculated dromedary camels. Emerg. Infect. Dis. 2014, 20, 1999–2005. [Google Scholar] [CrossRef] [Green Version]
- Widagdo, W.; Raj, V.S.; Schipper, D.; Kolijn, K.; van Leenders, G.J.L.H.; Bosch, B.J.; Bensaid, A.; Segalés, J.; Baumgärtner, W.; Osterhaus, A.D.M.E.; et al. Differential expression of the Middle East respiratory syndrome coronavirus receptor in the upper respiratory tracts of humans and dromedary camels. J. Virol. 2016, 90, 4838–4842. [Google Scholar] [CrossRef] [Green Version]
- Kandeil, A.; Gomaa, M.; Nageh, A.; Shehata, M.M.; Kayed, A.E.; Sabir, J.S.M.; Abiadh, A.; Jrijer, J.; Amr, Z.; Said, M.A.; et al. Middle East respiratory syndrome coronavirus (MERS-CoV) in dromedary camels in Africa and Middle East. Viruses 2019, 11, 717. [Google Scholar] [CrossRef] [Green Version]
- Reusken, C.B.; Raj, V.S.; Koopmans, M.P.; Haagmans, B.L. Cross host transmission in the emergence of MERS coronavirus. Curr. Opin. Virol. 2016, 16, 55–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalafalla, A.I.; Lu, X.; Al-Mubarak, A.I.A.; Dalab, A.H.S.; Al-Busadah, K.A.S.; Erdman, D.D. MERS-CoV in upper respiratory tract and lungs of dromedary camels, Saudi Arabia, 2013–2014. Emerg. Infect. Dis. 2015, 21, 1153–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.M.; Damdinjav, B.; Perera, R.A.; Chu, D.K.; Khishgee, B.; Enkhbold, B.; Poon, L.L.; Peiris, M. Absence of MERS-Coronavirus in bactrian camels, Southern Mongolia, November 2014. Emerg. Infect. Dis. 2015, 21, 1269–1271. [Google Scholar] [CrossRef]
- Ali, M.; El-Shesheny, R.; Kandeil, A.; Shehata, M.; Elsokary, B.; Gomaa, M.; Hassan, N.; El Sayed, A.; El-Taweel, A.; Sobhy, H.; et al. Cross-sectional surveillance of Middle East respiratory syndrome coronavirus (MERS-CoV) in dromedary camels and other mammals in Egypt, August 2015 to January 2016. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Epstein, J.H.; Rostal, M.K.; Islam, S.; Rahman, M.Z.; Hossain, M.E.; Uzzaman, M.S.; Munster, V.J.; Peiris, M.; Flora, M.S.; et al. Middle East Respiratory Syndrome Coronavirus Antibodies in Dromedary Camels, Bangladesh, 2015. Emerg. Infect. Dis. 2018, 24, 926–928. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, C.; Tejedor-Junco, M.T.; González, M.; Lattwein, E.; Renneker, S. Presence of antibodies but no evidence for circulation of MERS-CoV in dromedaries on the Canary Islands, 2015. Eurosurveillance 2015, 20, 30019. [Google Scholar] [CrossRef]
- David, D.; Rotenberg, D.; Khinich, E.; Erster, O.; Bardenstein, S.; van Straten, M.; Okba, N.M.A.; Raj, S.V.; Haagmans, B.L.; Miculitzki, M.; et al. Middle East respiratory syndrome coronavirus specific antibodies in naturally exposed Israeli llamas, alpacas and camels. One Health 2018, 5, 65–68. [Google Scholar] [CrossRef]
- Falzarano, D.; Kamissoko, B.; de Wit, E.; Maiga, O.; Cronin, J.; Samake, K.; Traore, A.; Milne-Price, S.; Munster, V.J.; Sogoba, N.; et al. Dromedary camels in northern Mali have high seropositivity to MERS-CoV. One Health 2017, 3, 41–43. [Google Scholar] [CrossRef]
- Memish, Z.A.; Al-Tawfiq, J.A.; Assiri, A.; AlRabiah, F.A.; Al Hajjar, S.; Albarrak, A.; Flemban, H.; Alhakeem, R.F.; Makhdoom, H.Q.; Alsubaie, S.; et al. Middle East respiratory syndrome coronavirus disease in children. Pediatric Infect. Dis. J. 2014, 33, 904–906. [Google Scholar] [CrossRef]
- Harrath, R.; Abu Duhier, F.M. Sero-prevalence of Middle East respiratory syndrome coronavirus (MERS-CoV) specific antibodies in dromedary camels in Tabuk, Saudi Arabia. J. Med. Virol. 2018, 90, 1285–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandersen, S.; Kobinger, G.P.; Soule, G.; Wernery, U. Middle East respiratory syndrome coronavirus antibody reactors among camels in Dubai, United Arab Emirates, in 2005. Transbound. Emerg. Dis. 2014, 61, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miguel, E.; Chevalier, V.; Ayelet, G.; Ben Bencheikh, M.N.; Boussini, H.; Chu, D.K.; El Berbri, I.; Fassi-Fihri, O.; Faye, B.; Fekadu, G.; et al. Risk factors for MERS coronavirus infection in dromedary camels in Burkina Faso, Ethiopia, and Morocco, 2015. Eurosurveillance 2017, 22, 30498. [Google Scholar] [CrossRef] [PubMed]
- Zohaib, A.; Saqib, M.; Athar, M.A.; Chen, J.; Sial, A.U.; Khan, S.; Taj, Z.; Sadia, H.; Tahir, U.; Tayyab, M.H.; et al. Countrywide survey for MERS-coronavirus antibodies in dromedaries and humans in Pakistan. Virol. Sin. 2018, 33, 410–417. [Google Scholar] [CrossRef] [Green Version]
- Saqib, M.; Sieberg, A.; Hussain, M.H.; Mansoor, M.K.; Zohaib, A.; Lattwein, E.; Muller, M.A.; Drosten, C.; Corman, V.M. Serologic evidence for MERS-CoV infection in dromedary camels, Punjab, Pakistan, 2012–2015. Emerg. Infect. Dis. 2017, 23, 550–551. [Google Scholar] [CrossRef] [Green Version]
- Reusken, C.B.; Schilp, C.; Raj, V.S.; De Bruin, E.; Kohl, R.H.; Farag, E.A.; Haagmans, B.L.; Al-Romaihi, H.; Le Grange, F.; Bosch, B.J.; et al. MERS-CoV infection of alpaca in a region where MERS-CoV is endemic. Emerg. Infect. Dis. 2016, 22, 1129–1131. [Google Scholar] [CrossRef] [PubMed]
- Rha, B.; Rudd, J.; Feikin, D.; Watson, J.; Curns, A.T.; Swerdlow, D.L.; Pallansch, M.A.; Gerber, S.I.; Centers for Disease Control and Prevention. Update on the epidemiology of Middle East respiratory syndrome coronavirus (MERS-CoV) infection, and guidance for the public, clinicians, and public health authorities—January 2015. MMWR Morb. Mortal. Wkly. Rep. 2015, 64, 61–62. [Google Scholar]
- Zumla, A.; Hui, D.S.; Perlman, S. Middle East respiratory syndrome. Lancet 2015. [Google Scholar] [CrossRef] [Green Version]
- WHO. Middle East Respiratory Syndrome Coronavirus (MERS-CoV): Summary of Current Situation, Literature Update and Risk Assessment–as of 5 February 2015; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Fanoy, E.B.; van der Sande, M.A.; Kraaij-Dirkzwager, M.; Dirksen, K.; Jonges, M.; van der Hoek, W.; Koopmans, M.P.; van der Werf, D.; Sonder, G.; van der Weijden, C.; et al. Travel-related MERS-CoV cases: An assessment of exposures and risk factors in a group of Dutch travellers returning from the Kingdom of Saudi Arabia, May 2014. Emerg. Themes Epidemiol. 2014, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- WHO. MERS-CoV in the Republic of Korea at a Glance; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Cowling, B.J.; Park, M.; Fang, V.J.; Wu, P.; Leung, G.M.; Wu, J.T. Preliminary epidemiological assessment of MERS-CoV outbreak in South Korea, May to June 2015. Eurosurveillance 2015, 20, 21163. [Google Scholar] [CrossRef] [Green Version]
- Wise, J. Patient with new strain of coronavirus is treated in intensive care at London hospital. BMJ 2012, 345, e6455. [Google Scholar] [CrossRef] [PubMed]
- WHO. Novel Coronavirus Infection in the United Kingdom; WHO: Geneva, Switzerland, 2012. [Google Scholar]
- WHO. Middle East Respiratory Syndrome Coronavirus (MERS-CoV), Report August 2019. Available online: http://www.emro.who.int/health-topics/mers-cov/mers-outbreaks.html (accessed on 30 June 2020).
- Sikkema, R.S.; Farag, E.; Islam, M.; Atta, M.; Reusken, C.; Al-Hajri, M.M.; Koopmans, M.P.G. Global status of Middle East respiratory syndrome coronavirus in dromedary camels: A systematic review—CORRIGENDUM. Epidemiol. Infect. 2019, 147, e198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, T.; Samy, A.M.; Peterson, A.T. MERS-CoV geography and ecology in the Middle East: Analyses of reported camel exposures and a preliminary risk map. BMC Res. Notes 2015, 8, 801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Abdallat, M.M.; Payne, D.C.; Alqasrawi, S.; Rha, B.; Tohme, R.A.; Abedi, G.R.; Al Nsour, M.; Iblan, I.; Jarour, N.; Farag, N.H.; et al. Hospital-associated outbreak of Middle East respiratory syndrome coronavirus: A serologic, epidemiologic, and clinical description. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2014, 59, 1225–1233. [Google Scholar] [CrossRef] [Green Version]
- Memish, Z.A.; Perlman, S.; Van Kerkhove, M.D.; Zumla, A. Middle east respiratory syndrome. Lancet 2020, 395, 1063–1077. [Google Scholar] [CrossRef]
- Corman, V.M.; Eckerle, I.; Bleicker, T.; Zaki, A.; Landt, O.; Eschbach-Bludau, M.; van Boheemen, S.; Gopal, R.; Ballhause, M.; Bestebroer, T.M.; et al. Detection of a novel human coronavirus by real-time reverse-transcription polymerase chain reaction. Eurosurveillance 2012, 17, 20285. [Google Scholar] [CrossRef] [Green Version]
- CDC. Severe respiratory illness associated with a novel coronavirus—Saudi Arabia and Qatar. MMWR Morb. Mortal. Wkly. Rep. 2012, 61, 820. [Google Scholar]
- Assiri, A.; McGeer, A.; Perl, T.M.; Price, C.S.; Al Rabeeah, A.A.; Cummings, D.A.; Alabdullatif, Z.N.; Assad, M.; Almulhim, A.; Makhdoom, H.; et al. Hospital outbreak of Middle East respiratory syndrome coronavirus. N. Engl. J. Med. 2013, 369, 407–416. [Google Scholar] [CrossRef]
- Guery, B.; Poissy, J.; el Mansouf, L.; Sejourne, C.; Ettahar, N.; Lemaire, X.; Vuotto, F.; Goffard, A.; Behillil, S.; Enouf, V.; et al. Clinical features and viral diagnosis of two cases of infection with Middle East Respiratory Syndrome coronavirus: A report of nosocomial transmission. Lancet 2013, 381, 2265–2272. [Google Scholar] [CrossRef] [Green Version]
- Oboho, I.K.; Tomczyk, S.M.; Al-Asmari, A.M.; Banjar, A.A.; Al-Mugti, H.; Aloraini, M.S.; Alkhaldi, K.Z.; Almohammadi, E.L.; Alraddadi, B.M.; Gerber, S.I.; et al. 2014 MERS-CoV outbreak in Jeddah—A link to health care facilities. N. Engl. J. Med. 2015, 372, 846–854. [Google Scholar] [CrossRef]
- Bialek, S.R.; Allen, D.; Alvarado-Ramy, F.; Arthur, R.; Balajee, A.; Bell, D.; Best, S.; Blackmore, C.; Breakwell, L.; Cannons, A.; et al. First confirmed cases of Middle East respiratory syndrome coronavirus (MERS-CoV) infection in the United States, updated information on the epidemiology of MERS-CoV infection, and guidance for the public, clinicians, and public health authorities—May 2014. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 431–436. [Google Scholar] [PubMed]
- Centers for Disease Control and Precention. Interim Guidelines for Collecting, Handling, and Testing Clinical Specimens from Patients under Investigation (PUIs) for Middle East Respiratory Syndrome Coronavirus (MERS-CoV); CDC: Atlanta, GA, USA, 2014; Version 2.1.
- Park, J.; Lee, D.S.; Christakis, N.A.; Barabasi, A.L. The impact of cellular networks on disease comorbidity. Mol. Syst. Biol. 2009, 5, 262. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.S.; Azhar, E.I.; Kim, Y.J.; Memish, Z.A.; Oh, M.D.; Zumla, A. Middle East respiratory syndrome coronavirus: Risk factors and determinants of primary, household, and nosocomial transmission. Lancet Infect. Dis. 2018, 18, e217–e227. [Google Scholar] [CrossRef] [Green Version]
- Badawi, A.; Ryoo, S.G. Prevalence of comorbidities in the Middle East respiratory syndrome coronavirus (MERS-CoV): A systematic review and meta-analysis. Int. J. Infect. Dis. 2016, 49, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Alqahtani, F.Y.; Aleanizy, F.S.; Ali El Hadi Mohamed, R.; Alanazi, M.S.; Mohamed, N.; Alrasheed, M.M.; Abanmy, N.; Alhawassi, T. Prevalence of comorbidities in cases of Middle East respiratory syndrome coronavirus: A retrospective study. Epidemiol. Infect. 2018, 147, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-M.; Hsu, C.-Y.; Lai, C.-C.; Yen, M.-F.; Wikramaratna, P.S.; Chen, H.-H.; Wang, T.-H. Impact of comorbidity on fatality rate of patients with Middle East respiratory syndrome. Sci. Rep. 2017, 7, 11307. [Google Scholar] [CrossRef] [Green Version]
- Seys, L.J.M.; Widagdo, W.; Verhamme, F.M.; Kleinjan, A.; Janssens, W.; Joos, G.F.; Bracke, K.R.; Haagmans, B.L.; Brusselle, G.G. DPP4, the Middle East respiratory syndrome coronavirus receptor, is upregulated in lungs of smokers and chronic obstructive pulmonary disease patients. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2018, 66, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.; Li, K.S.; Tsang, A.K.; Lam, C.S.; Ahmed, S.; Chen, H.; Chan, K.H.; Woo, P.C.; Yuen, K.Y. Genetic characterization of Betacoronavirus lineage C viruses in bats reveals marked sequence divergence in the spike protein of pipistrellus bat coronavirus HKU5 in Japanese pipistrelle: Implications for the origin of the novel Middle East respiratory syndrome coronavirus. J. Virol. 2013, 87, 8638–8650. [Google Scholar] [CrossRef] [Green Version]
- Ithete, N.L.; Stoffberg, S.; Corman, V.M.; Cottontail, V.M.; Richards, L.R.; Schoeman, M.C.; Drosten, C.; Drexler, J.F.; Preiser, W. Close relative of human Middle East respiratory syndrome coronavirus in bat, South Africa. Emerg. Infect. Dis. 2013, 19, 1697–1699. [Google Scholar] [CrossRef]
- Hatcher, E.L.; Zhdanov, S.A.; Bao, Y.; Blinkova, O.; Nawrocki, E.P.; Ostapchuck, Y.; Schaffer, A.A.; Brister, J.R. Virus Variation Resource—Improved response to emergent viral outbreaks. Nucleic Acids Res. 2017, 45, D482–D490. [Google Scholar] [CrossRef]
- Lau, S.K.; Wernery, R.; Wong, E.Y.; Joseph, S.; Tsang, A.K.; Patteril, N.A.; Elizabeth, S.K.; Chan, K.H.; Muhammed, R.; Kinne, J.; et al. Polyphyletic origin of MERS coronaviruses and isolation of a novel clade A strain from dromedary camels in the United Arab Emirates. Emerg. Microbes Infect. 2016, 5, e128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, K.G.; Redden, C.L.; Bishop-Lilly, K.A.; Johnson, R.; Hensley, L.E.; Raviprakash, K.; Luke, T.; Kochel, T.; Mokashi, V.P.; Defang, G.N. Full-genome sequence of human betacoronavirus 2c jordan-n3/2012 after serial passage in Mammalian cells. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandeil, A.; Shehata, M.M.; El Shesheny, R.; Gomaa, M.R.; Ali, M.A.; Kayali, G. Complete genome sequence of Middle East respiratory syndrome coronavirus isolated from a dromedary camel in Egypt. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleine-Weber, H.; Elzayat, M.T.; Wang, L.; Graham, B.S.; Muller, M.A.; Drosten, C.; Pohlmann, S.; Hoffmann, M. Mutations in the spike protein of Middle East respiratory syndrome coronavirus transmitted in Korea increase resistance to antibody-mediated neutralization. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Cheon, S.; Min, C.K.; Sohn, K.M.; Kang, Y.J.; Cha, Y.J.; Kang, J.I.; Han, S.K.; Ha, N.Y.; Kim, G.; et al. Spread of mutant Middle East respiratory syndrome coronavirus with reduced affinity to human CD26 during the South Korean outbreak. mBio 2016, 7, e00019. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Shen, L.; Gu, X. Evolutionary dynamics of MERS-CoV: Potential recombination, positive selection and transmission. Sci. Rep. 2016, 6, 25049. [Google Scholar] [CrossRef] [Green Version]
- Drosten, C.; Muth, D.; Corman, V.M.; Hussain, R.; Al Masri, M.; HajOmar, W.; Landt, O.; Assiri, A.; Eckerle, I.; Al Shangiti, A.; et al. An observational, laboratory-based study of outbreaks of middle East respiratory syndrome coronavirus in Jeddah and Riyadh, kingdom of Saudi Arabia, 2014. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2015, 60, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Fagbo, S.F.; Skakni, L.; Chu, D.K.; Garbati, M.A.; Joseph, M.; Peiris, M.; Hakawi, A.M. Molecular epidemiology of hospital outbreak of Middle East respiratory syndrome, Riyadh, Saudi Arabia, 2014. Emerg. Infect. Dis. 2015, 21, 1981–1988. [Google Scholar] [CrossRef] [Green Version]
- Forni, D.; Filippi, G.; Cagliani, R.; De Gioia, L.; Pozzoli, U.; Al-Daghri, N.; Clerici, M.; Sironi, M. The heptad repeat region is a major selection target in MERS-CoV and related coronaviruses. Sci. Rep. 2015, 5, 14480. [Google Scholar] [CrossRef] [Green Version]
- Cotten, M.; Watson, S.J.; Zumla, A.I.; Makhdoom, H.Q.; Palser, A.L.; Ong, S.H.; Al Rabeeah, A.A.; Alhakeem, R.F.; Assiri, A.; Al-Tawfiq, J.A.; et al. Spread, circulation, and evolution of the Middle East respiratory syndrome coronavirus. mBio 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Hunter, J.C.; Nguyen, D.; Aden, B.; Al Bandar, Z.; Al Dhaheri, W.; Abu Elkheir, K.; Khudair, A.; Al Mulla, M.; El Saleh, F.; Imambaccus, H.; et al. Transmission of Middle East respiratory syndrome coronavirus infections in healthcare settings, Abu Dhabi. Emerg. Infect. Dis. 2016, 22, 647–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Liu, C.; Du, L.; Jiang, S.; Shi, Z.; Baric, R.S.; Li, F. Two mutations were critical for bat-to-human transmission of Middle East respiratory syndrome coronavirus. J. Virol. 2015, 89, 9119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Wohlford-Lenane, C.L.; Channappanavar, R.; Park, J.E.; Earnest, J.T.; Bair, T.B.; Bates, A.M.; Brogden, K.A.; Flaherty, H.A.; Gallagher, T.; et al. Mouse-adapted MERS coronavirus causes lethal lung disease in human DPP4 knockin mice. Proc. Natl. Acad. Sci. USA 2017, 114, E3119–E3128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Wit, E.; Feldmann, F.; Cronin, J.; Jordan, R.; Okumura, A.; Thomas, T.; Scott, D.; Cihlar, T.; Feldmann, H. Prophylactic and therapeutic remdesivir (GS-5734) treatment in the rhesus macaque model of MERS-CoV infection. Proc. Natl. Acad. Sci. USA 2020, 117, 6771–6776. [Google Scholar] [CrossRef] [Green Version]
- Cong, Y.; Hart, B.J.; Gross, R.; Zhou, H.; Frieman, M.; Bollinger, L.; Wada, J.; Hensley, L.E.; Jahrling, P.B.; Dyall, J.; et al. MERS-CoV pathogenesis and antiviral efficacy of licensed drugs in human monocyte-derived antigen-presenting cells. PLoS ONE 2018, 13, e0194868. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Leist, S.R.; Schäfer, A.; Won, J.; Brown, A.J.; Montgomery, S.A.; Hogg, A.; Babusis, D.; Clarke, M.O.; et al. Comparative therapeutic efficacy of remdesivir and combination lopinavir, ritonavir, and interferon beta against MERS-CoV. Nat. Commun. 2020, 11, 222. [Google Scholar] [CrossRef] [Green Version]
- Dyall, J.; Coleman, C.M.; Hart, B.J.; Venkataraman, T.; Holbrook, M.R.; Kindrachuk, J.; Johnson, R.F.; Olinger, G.G., Jr.; Jahrling, P.B.; Laidlaw, M.; et al. Repurposing of clinically developed drugs for treatment of Middle East respiratory syndrome coronavirus infection. Antimicrob. Agents Chemother. 2014, 58, 4885–4893. [Google Scholar] [CrossRef] [Green Version]
- Burkard, C.; Verheije, M.H.; Haagmans, B.L.; van Kuppeveld, F.J.; Rottier, P.J.; Bosch, B.J.; de Haan, C.A. ATP1A1-mediated Src signaling inhibits coronavirus entry into host cells. J. Virol. 2015, 89, 4434–4448. [Google Scholar] [CrossRef] [Green Version]
- Shirato, K.; Kawase, M.; Matsuyama, S. Middle East respiratory syndrome coronavirus infection mediated by the transmembrane serine protease TMPRSS2. J. Virol. 2013, 87, 12552–12561. [Google Scholar] [CrossRef] [Green Version]
- Muller, C.; Schulte, F.W.; Lange-Grunweller, K.; Obermann, W.; Madhugiri, R.; Pleschka, S.; Ziebuhr, J.; Hartmann, R.K.; Grunweller, A. Broad-spectrum antiviral activity of the eIF4A inhibitor silvestrol against corona- and picornaviruses. Antivir. Res. 2018, 150, 123–129. [Google Scholar] [CrossRef]
- Agostini, M.L.; Andres, E.L.; Sims, A.C.; Graham, R.L.; Sheahan, T.P.; Lu, X.; Smith, E.C.; Case, J.B.; Feng, J.Y.; Jordan, R.; et al. Coronavirus susceptibility to the antiviral remdesivir (GS-5734) is mediated by the viral polymerase and the proofreading exoribonuclease. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.S.; Jung, E.; Kim, M.; Baric, R.S.; Go, Y.Y. Saracatinib inhibits Middle East respiratory syndrome-coronavirus replication in vitro. Viruses 2018, 10, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Wilde, A.H.; Raj, V.S.; Oudshoorn, D.; Bestebroer, T.M.; van Nieuwkoop, S.; Limpens, R.; Posthuma, C.C.; van der Meer, Y.; Bárcena, M.; Haagmans, B.L.; et al. MERS-coronavirus replication induces severe in vitro cytopathology and is strongly inhibited by cyclosporin A or interferon-α treatment. J. Gen. Virol. 2013, 94, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Kindrachuk, J.; Ork, B.; Hart, B.J.; Mazur, S.; Holbrook, M.R.; Frieman, M.B.; Traynor, D.; Johnson, R.F.; Dyall, J.; Kuhn, J.H.; et al. Antiviral potential of ERK/MAPK and PI3K/AKT/mTOR signaling modulation for Middle East respiratory syndrome coronavirus infection as identified by temporal kinome analysis. Antimicrob. Agents Chemother. 2015, 59, 1088–1099. [Google Scholar] [CrossRef] [Green Version]
- de Wilde, A.H.; Jochmans, D.; Posthuma, C.C.; Zevenhoven-Dobbe, J.C.; van Nieuwkoop, S.; Bestebroer, T.M.; van den Hoogen, B.G.; Neyts, J.; Snijder, E.J. Screening of an FDA-approved compound library identifies four small-molecule inhibitors of Middle East respiratory syndrome coronavirus replication in cell culture. Antimicrob. Agents Chemother. 2014, 58, 4875–4884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Chu, H.; Li, C.; Wong, B.H.; Cheng, Z.S.; Poon, V.K.; Sun, T.; Lau, C.C.; Wong, K.K.; Chan, J.Y.; et al. Active replication of Middle East respiratory syndrome coronavirus and aberrant induction of inflammatory cytokines and chemokines in human macrophages: Implications for pathogenesis. J. Infect. Dis. 2014, 209, 1331–1342. [Google Scholar] [CrossRef]
- Lin, S.C.; Ho, C.T.; Chuo, W.H.; Li, S.; Wang, T.T.; Lin, C.C. Effective inhibition of MERS-CoV infection by resveratrol. BMC Infect. Dis. 2017, 17, 144. [Google Scholar] [CrossRef] [Green Version]
- DeZure, A.D.; Berkowitz, N.M.; Graham, B.S.; Ledgerwood, J.E. Whole-inactivated and virus-like particle vaccine strategies for chikungunya virus. J. Infect. Dis. 2016, 214, S497–S499. [Google Scholar] [CrossRef]
- Shehata, M.M.; Mostafa, A.; Teubner, L.; Mahmoud, S.H.; Kandeil, A.; Elshesheny, R.; Frantz, R.; La Pietra, L.; Pleschka, S.; Osman, A.; et al. Bacterial Outer Membrane Vesicles (OMVs)-based dual vaccine for influenza A H1N1 virus and MERS-CoV. Vaccines 2019, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Lan, J.; Bao, L.; Huang, B.; Ye, F.; Chen, Y.; Yao, Y.; Wang, W.; Qin, C.; Tan, W. Enhanced protection in mice induced by immunization with inactivated whole viruses compare to spike protein of middle east respiratory syndrome coronavirus. Emerg. Microbes Infect. 2018, 7, 60. [Google Scholar] [CrossRef] [Green Version]
- Wirblich, C.; Coleman, C.M.; Kurup, D.; Abraham, T.S.; Bernbaum, J.G.; Jahrling, P.B.; Hensley, L.E.; Johnson, R.F.; Frieman, M.B.; Schnell, M.J. One-Health: A safe, efficient, dual-use vaccine for humans and animals against Middle East respiratory syndrome coronavirus and rabies virus. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, A.S.; Tao, X.; Algaissi, A.; Garron, T.; Narayanan, K.; Peng, B.H.; Couch, R.B.; Tseng, C.T. Immunization with inactivated Middle East Respiratory Syndrome coronavirus vaccine leads to lung immunopathology on challenge with live virus. Hum. Vaccines Immunother. 2016, 12, 2351–2356. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.T.; Sbrana, E.; Iwata-Yoshikawa, N.; Newman, P.C.; Garron, T.; Atmar, R.L.; Peters, C.J.; Couch, R.B. Immunization with SARS coronavirus vaccines leads to pulmonary immunopathology on challenge with the SARS virus. PLoS ONE 2012, 7, e35421. [Google Scholar] [CrossRef]
- Shehata, M.M.; Kandeil, A.; Mostafa, A.; Mahmoud, S.H.; Gomaa, M.R.; El-Shesheny, R.; Webby, R.; Kayali, G.; Ali, M.A. A Recombinant Influenza A/H1N1 Carrying A Short Immunogenic Peptide of MERS-CoV as Bivalent Vaccine in BALB/c Mice. Pathogens 2019, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Almazan, F.; DeDiego, M.L.; Sola, I.; Zuniga, S.; Nieto-Torres, J.L.; Marquez-Jurado, S.; Andres, G.; Enjuanes, L. Engineering a replication-competent, propagation-defective Middle East respiratory syndrome coronavirus as a vaccine candidate. mBio 2013, 4, e00650-13. [Google Scholar] [CrossRef] [Green Version]
- Malczyk, A.H.; Kupke, A.; Prufer, S.; Scheuplein, V.A.; Hutzler, S.; Kreuz, D.; Beissert, T.; Bauer, S.; Hubich-Rau, S.; Tondera, C.; et al. A highly immunogenic and protective Middle East respiratory syndrome coronavirus vaccine based on a recombinant measles virus vaccine platform. J. Virol. 2015, 89, 11654–11667. [Google Scholar] [CrossRef] [Green Version]
- Menachery, V.D.; Gralinski, L.E.; Mitchell, H.D.; Dinnon, K.H., 3rd; Leist, S.R.; Yount, B.L., Jr.; Graham, R.L.; McAnarney, E.T.; Stratton, K.G.; Cockrell, A.S.; et al. Middle East respiratory syndrome coronavirus nonstructural protein 16 is necessary for interferon resistance and viral pathogenesis. mSphere 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Bodmer, B.S.; Fiedler, A.H.; Hanauer, J.R.H.; Prufer, S.; Muhlebach, M.D. Live-attenuated bivalent measles virus-derived vaccines targeting Middle East respiratory syndrome coronavirus induce robust and multifunctional T cell responses against both viruses in an appropriate mouse model. Virology 2018, 521, 99–107. [Google Scholar] [CrossRef]
- Liu, R.; Wang, J.; Shao, Y.; Wang, X.; Zhang, H.; Shuai, L.; Ge, J.; Wen, Z.; Bu, Z. A recombinant VSV-vectored MERS-CoV vaccine induces neutralizing antibody and T cell responses in rhesus monkeys after single dose immunization. Antivir. Res. 2018, 150, 30–38. [Google Scholar] [CrossRef]
- Kato, H.; Takayama-Ito, M.; Iizuka-Shiota, I.; Fukushi, S.; Posadas-Herrera, G.; Horiya, M.; Satoh, M.; Yoshikawa, T.; Yamada, S.; Harada, S.; et al. Development of a recombinant replication-deficient rabies virus-based bivalent-vaccine against MERS-CoV and rabies virus and its humoral immunogenicity in mice. PLoS ONE 2019, 14, e0223684. [Google Scholar] [CrossRef]
- Du, L.; Tai, W.; Zhou, Y.; Jiang, S. Vaccines for the prevention against the threat of MERS-CoV. Expert Rev. Vaccines 2016, 15, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Kou, Z.; Ma, C.; Tao, X.; Wang, L.; Zhao, G.; Chen, Y.; Yu, F.; Tseng, C.T.; Zhou, Y.; et al. A truncated receptor-binding domain of MERS-CoV spike protein potently inhibits MERS-CoV infection and induces strong neutralizing antibody responses: Implication for developing therapeutics and vaccines. PLoS ONE 2013, 8, e81587. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Wang, L.; Tao, X.; Zhang, N.; Yang, Y.; Tseng, C.T.; Li, F.; Zhou, Y.; Jiang, S.; Du, L. Searching for an ideal vaccine candidate among different MERS coronavirus receptor-binding fragments--the importance of immunofocusing in subunit vaccine design. Vaccine 2014, 32, 6170–6176. [Google Scholar] [CrossRef] [PubMed]
- Mou, H.; Raj, V.S.; van Kuppeveld, F.J.; Rottier, P.J.; Haagmans, B.L.; Bosch, B.J. The receptor binding domain of the new Middle East respiratory syndrome coronavirus maps to a 231-residue region in the spike protein that efficiently elicits neutralizing antibodies. J. Virol. 2013, 87, 9379–9383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Du, L.; Ma, C.; Li, Y.; Li, L.; Poon, V.K.; Wang, L.; Yu, F.; Zheng, B.J.; Jiang, S.; et al. A safe and convenient pseudovirus-based inhibition assay to detect neutralizing antibodies and screen for viral entry inhibitors against the novel human coronavirus MERS-CoV. Virol. J. 2013, 10, 266. [Google Scholar] [CrossRef] [Green Version]
- Lan, J.; Deng, Y.; Chen, H.; Lu, G.; Wang, W.; Guo, X.; Lu, Z.; Gao, G.F.; Tan, W. Tailoring subunit vaccine immunity with adjuvant combinations and delivery routes using the Middle East respiratory coronavirus (MERS-CoV) receptor-binding domain as an antigen. PLoS ONE 2014, 9, e112602. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Channappanavar, R.; Ma, C.; Wang, L.; Tang, J.; Garron, T.; Tao, X.; Tasneem, S.; Lu, L.; Tseng, C.T.; et al. Identification of an ideal adjuvant for receptor-binding domain-based subunit vaccines against Middle East respiratory syndrome coronavirus. Cell. Mol. Immunol. 2016, 13, 180–190. [Google Scholar] [CrossRef] [Green Version]
- Lan, J.; Deng, Y.; Song, J.; Huang, B.; Wang, W.; Tan, W. Significant spike-specific IgG and neutralizing antibodies in mice induced by a novel chimeric virus-like particle vaccine candidate for Middle East respiratory syndrome coronavirus. Virol. Sin. 2018, 33, 453–455. [Google Scholar] [CrossRef] [Green Version]
- Tai, W.; Zhao, G.; Sun, S.; Guo, Y.; Wang, Y.; Tao, X.; Tseng, C.K.; Li, F.; Jiang, S.; Du, L.; et al. A recombinant receptor-binding domain of MERS-CoV in trimeric form protects human dipeptidyl peptidase 4 (hDPP4) transgenic mice from MERS-CoV infection. Virology 2016, 499, 375–382. [Google Scholar] [CrossRef]
- Pallesen, J.; Wang, N.; Corbett, K.S.; Wrapp, D.; Kirchdoerfer, R.N.; Turner, H.L.; Cottrell, C.A.; Becker, M.M.; Wang, L.; Shi, W.; et al. Immunogenicity and structures of a rationally designed prefusion MERS-CoV spike antigen. Proc. Natl. Acad. Sci. USA 2017, 114, E7348–E7357. [Google Scholar] [CrossRef] [Green Version]
- Coleman, C.M.; Liu, Y.V.; Mu, H.; Taylor, J.K.; Massare, M.; Flyer, D.C.; Glenn, G.M.; Smith, G.E.; Frieman, M.B. Purified coronavirus spike protein nanoparticles induce coronavirus neutralizing antibodies in mice. Vaccine 2014, 32, 3169–3174. [Google Scholar] [CrossRef] [PubMed]
- Jiaming, L.; Yanfeng, Y.; Yao, D.; Yawei, H.; Linlin, B.; Baoying, H.; Jinghua, Y.; Gao, G.F.; Chuan, Q.; Wenjie, T. The recombinant N-terminal domain of spike proteins is a potential vaccine against Middle East respiratory syndrome coronavirus (MERS-CoV) infection. Vaccine 2017, 35, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shi, W.; Joyce, M.G.; Modjarrad, K.; Zhang, Y.; Leung, K.; Lees, C.R.; Zhou, T.; Yassine, H.M.; Kanekiyo, M.; et al. Evaluation of candidate vaccine approaches for MERS-CoV. Nat. Commun. 2015, 6, 7712. [Google Scholar] [CrossRef] [PubMed]
- Adney, D.R.; Wang, L.; van Doremalen, N.; Shi, W.; Zhang, Y.; Kong, W.P.; Miller, M.R.; Bushmaker, T.; Scott, D.; de Wit, E.; et al. Efficacy of an adjuvanted Middle East respiratory syndrome coronavirus spike protein vaccine in dromedary camels and alpacas. Viruses 2019, 11, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zheng, X.; Gai, W.; Wong, G.; Wang, H.; Jin, H.; Feng, N.; Zhao, Y.; Zhang, W.; Li, N.; et al. Novel chimeric virus-like particles vaccine displaying MERS-CoV receptor-binding domain induce specific humoral and cellular immune response in mice. Antivir. Res. 2017, 140, 55–61. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, X.; Gai, W.; Zhao, Y.; Wang, H.; Wang, H.; Feng, N.; Chi, H.; Qiu, B.; Li, N.; et al. MERS-CoV virus-like particles produced in insect cells induce specific humoural and cellular imminity in rhesus macaques. Oncotarget 2017, 8, 12686–12694. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Deng, Y.; Chen, H.; Lan, J.; Wang, W.; Zou, X.; Hung, T.; Lu, Z.; Tan, W. Systemic and mucosal immunity in mice elicited by a single immunization with human adenovirus type 5 or 41 vector-based vaccines carrying the spike protein of Middle East respiratory syndrome coronavirus. Immunology 2015, 145, 476–484. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Okada, K.; Kenniston, T.; Raj, V.S.; AlHajri, M.M.; Farag, E.A.; AlHajri, F.; Osterhaus, A.D.; Haagmans, B.L.; Gambotto, A. Immunogenicity of an adenoviral-based Middle East Respiratory Syndrome coronavirus vaccine in BALB/c mice. Vaccine 2014, 32, 5975–5982. [Google Scholar] [CrossRef]
- Hashem, A.M.; Algaissi, A.; Agrawal, A.; Al-Amri, S.S.; Alhabbab, R.Y.; Sohrab, S.S.; Almasoud, A.; Alharbi, N.K.; Peng, B.H.; Russell, M.; et al. A highly immunogenic, protective and safe adenovirus-based vaccine expressing MERS-CoV S1-CD40L fusion protein in transgenic human DPP4 mouse model. J. Infect. Dis. 2019. [Google Scholar] [CrossRef]
- Alharbi, N.K.; Padron-Regalado, E.; Thompson, C.P.; Kupke, A.; Wells, D.; Sloan, M.A.; Grehan, K.; Temperton, N.; Lambe, T.; Warimwe, G.; et al. ChAdOx1 and MVA based vaccine candidates against MERS-CoV elicit neutralising antibodies and cellular immune responses in mice. Vaccine 2017, 35, 3780–3788. [Google Scholar] [CrossRef]
- Munster, V.J.; Wells, D.; Lambe, T.; Wright, D.; Fischer, R.J.; Bushmaker, T.; Saturday, G.; van Doremalen, N.; Gilbert, S.C.; de Wit, E.; et al. Protective efficacy of a novel simian adenovirus vaccine against lethal MERS-CoV challenge in a transgenic human DPP4 mouse model. NPJ Vaccines 2017, 2, 28. [Google Scholar] [CrossRef] [Green Version]
- Song, F.; Fux, R.; Provacia, L.B.; Volz, A.; Eickmann, M.; Becker, S.; Osterhaus, A.D.; Haagmans, B.L.; Sutter, G. Middle East respiratory syndrome coronavirus spike protein delivered by modified vaccinia virus Ankara efficiently induces virus-neutralizing antibodies. J. Virol. 2013, 87, 11950–11954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volz, A.; Kupke, A.; Song, F.; Jany, S.; Fux, R.; Shams-Eldin, H.; Schmidt, J.; Becker, C.; Eickmann, M.; Becker, S.; et al. Protective efficacy of recombinant Modified Vaccinia virus Ankara (MVA) delivering Middle East Respiratory Syndrome coronavirus spike glycoprotein. J. Virol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Li, K.; Wohlford-Lenane, C.; Agnihothram, S.S.; Fett, C.; Zhao, J.; Gale, M.J., Jr.; Baric, R.S.; Enjuanes, L.; Gallagher, T.; et al. Rapid generation of a mouse model for Middle East respiratory syndrome. Proc. Natl. Acad. Sci. USA 2014, 111, 4970–4975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.Q.; Ge, J.-Y.; Wang, J.-L.; Shao, Y.; Zhang, H.-L.; Wang, J.-L.; Wen, Z.-Y.; Bu, Z.-G. Newcastle disease virus-based mers-cov candidate vaccine elicits high-level and lasting neutralizing antibodies in bactrian camels. J. Integr. Agric. 2017, 16. [Google Scholar] [CrossRef]
- Muthumani, K.; Falzarano, D.; Reuschel, E.L.; Tingey, C.; Flingai, S.; Villarreal, D.O.; Wise, M.; Patel, A.; Izmirly, A.; Aljuaid, A.; et al. A synthetic consensus anti-spike protein DNA vaccine induces protective immunity against Middle East respiratory syndrome coronavirus in nonhuman primates. Sci. Transl. Med. 2015, 7, 301ra132. [Google Scholar] [CrossRef] [Green Version]
- Chi, H.; Zheng, X.; Wang, X.; Wang, C.; Wang, H.; Gai, W.; Perlman, S.; Yang, S.; Zhao, J.; Xia, X. DNA vaccine encoding Middle East respiratory syndrome coronavirus S1 protein induces protective immune responses in mice. Vaccine 2017, 35, 2069–2075. [Google Scholar] [CrossRef]
- Al-Amri, S.S.; Abbas, A.T.; Siddiq, L.A.; Alghamdi, A.; Sanki, M.A.; Al-Muhanna, M.K.; Alhabbab, R.Y.; Azhar, E.I.; Li, X.; Hashem, A.M. Immunogenicity of candidate MERS-CoV DNA vaccines based on the spike protein. Sci. Rep. 2017, 7, 44875. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Country | Sampling Year(s) | Sample Size | Assay Applied | Host | Sero-Positivity (%) | Age | Ref. |
|---|---|---|---|---|---|---|---|
| Australia | 2013–14 | 25 | PNA | Dromedary camel | 0 | Ad/Ju | [26] |
| Bangladesh | 2015 | 55 | PNA | Dromedary camel | 31 | Ad/Ju | [59] |
| Bangladesh | 2015 | 18 | PNA, ELISA | Sheep | 0 | Ad | [59] |
| Chile | 2012–13 | 2 | PM, NA | Bactrian camel | 0 | UK | [46] |
| Chile | 2012–13 | 25 | PM, NA | Alpaca, llama | 0 | UK | [46] |
| Netherlands | 2012–13 | 2 | PM, NA | Bactrian camel | 0 | UK | [46] |
| Netherlands | 2012–13 | 8 | PM, NA | Alpaca, llama | 0 | UK | [46] |
| Spain “Canary Islands” | 2012–13 | 195 | PM, NA | Dromedary camel | 14 | Ad/Ju | [46] |
| Spain “Canary Islands” | 2015 | 170 | ELISA | Dromedary camel | 4.1 | UK | [60] |
| Egypt | 2013 | 110 | PNA | Dromedary camel | 94 | Ad | [29] |
| Egypt | 2013 | 52 | PNA | Dromedary camel | 92.3 | Ad | [13] |
| Egypt | 1997 | 43 | ELISA, NA | Dromedary camel | 81.4 | UK | [48] |
| Egypt | 2015–16 | 1031 | NA | Dromedary camel | 84.5 | Ad/Ju | [58] |
| Egypt | 2014–16 | 2541 | NA | Dromedary camel | 71 | Ad/Ju | [45] |
| Egypt | 2015–16 | 145 | NA | Sheep, donkey, goat, cattle, buffalo | 0.7 | Ad | [58] |
| Egypt | 2015–17 | 409 | NA, PNA | Cattle, sheep, goat, donkey, buffalo, and horse | 1.7 | Ad/Ju | [43] |
| Egypt | 2016–18 | 2033 | NA | Dromedary camel | 68.9 | Ad/Ju | [54] |
| Ethiopia | 2010–11 | 188 | PM | Dromedary camel | 96 | Ad | [23] |
| Iraq | 2017 | 32 | NA | Dromedary camel | 43.7 | Ad/Ju | [54] |
| Israeli | 2013 | 71 | NA, IFA | Dromedary camel | 71.8 | Ad/Ju | [31] |
| Israeli | 2012–17 | 411 | ELISA, NA | Dromedary camel | 62 | Ad/Ju | [61] |
| Israeli | 2016 | 121 | ELISA, NA | Alpaca, llama | 23.5 | UK | [61] |
| Jordan | 2013 | 11 | PM, NA | Dromedary camel | 100 | Ju | [30] |
| Jordan | 2015–16 | 304 | NA | Dromedary camel | 81 | Ad/Ju | [54] |
| Kenya | 1993–2013 | 774 | ELISA | Dromedary camel | 27.5 | UK | [21] |
| Kenya | 2016–2017 | 1421 | ELISA | Dromedary camel | 63.7 | Ad/Ju | [32] |
| Mali | 2009–10 | 262 | NA, ELISA | Dromedary camel | 90 | Ad/Ju | [62] |
| Mongolia | 2015 | 200 | PNA | Bactrian camel | 0 | Ad/Ju | [57] |
| Nigeria | 2010–11 | 358 | PM | Dromedary camel | 94 | Ad | [23] |
| Oman | 2013 | 50 | PM, NA | Dromedary camel | 100 | Ad | [46] |
| Qatar | 2013 | 14 | NA | Dromedary camel | 100 | UK | [50] |
| KSA | 2010–13 | 310 | PNA | Dromedary camel | 90 | Ad/Ju | [22] |
| KSA | 2014 | 9 | IFA | Dromedary camel | 22.2 | Ad/Ju | [63] |
| KSA | 2016 | 171 | ELISA | Dromedary camel | 84.21 | Ad/Ju | [64] |
| KSA | 2017 | 222 | NA | Dromedary camel | 81.3 | Ad/Ju | [54] |
| Senegal | 2017 | 72 | NA, PNA | Sheep and goat | 51.3 | Ad/Ju | [43] |
| Senegal | 2017 | 198 | NA | Dromedary camel | 65.1 | Ad/Ju | [54] |
| Somalia | 1983, 1984 | 25, 61 | ELISA, NA | Dromedary camel | 80, 85.2 | Ad | [48] |
| Sudan | 1983 | 60 | ELISA, NA | Dromedary camel | 86.7 | Ad/Ju | [48] |
| Tunisia | 2010–11 | 204 | PM | Dromedary camel | 48.5 | Ad/Ju | [23] |
| Tunisia | 2016–17 | 342 | NA, PNA | Cattle, sheep, goats, donkey, mule, and horse | 1.7 | Ad/Ju | [43] |
| Tunisia | 2015–18 | 782 | NA | Dromedary camel | 87.3 | Ad/Ju | [54] |
| UAE | 2005 | 151, 500 | SPM | Dromedary camel | 81.8 | Ad | [25] |
| Uganda | 2017 | 500 | NA | Dromedary camel | 61.6 | Ad/Ju | [54] |
| USA/Canada | 2000–01 | 6 | NA | Dromedary camel | 0 | Ad | [65] |
| Ethiopia | 2015 | 632 | NA, PNA | Dromedary camel | 93.8 | Ad/Ju | [66] |
| Burkina Faso | 2015 | 525 | NA, PNA | Dromedary camel | 77.1 | Ad/Ju | [66] |
| Morocco | 2015 | 343 | NA, PNA | Dromedary camel | 74.2 | Ad/Ju | [66] |
| Pakistan | 2015–2018 | 1050 | ELISA | Dromedary camel | 75 | Ad | [67] |
| Pakistan | 2012–2015 | 565 | ELISA | Dromedary camel | 39.5 | Ad/Ju | [68] |
| Qatar | 2015 | 15 | DPM, PNA | Dromedary camel | 100 | Ad/Ju | [69] |
| Qatar | 2015 | 10 | DPM, PNA | Alpaca | 90 | UK | [69] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mostafa, A.; Kandeil, A.; Shehata, M.; El Shesheny, R.; Samy, A.M.; Kayali, G.; Ali, M.A. Middle East Respiratory Syndrome Coronavirus (MERS-CoV): State of the Science. Microorganisms 2020, 8, 991. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8070991
Mostafa A, Kandeil A, Shehata M, El Shesheny R, Samy AM, Kayali G, Ali MA. Middle East Respiratory Syndrome Coronavirus (MERS-CoV): State of the Science. Microorganisms. 2020; 8(7):991. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8070991
Chicago/Turabian StyleMostafa, Ahmed, Ahmed Kandeil, Mahmoud Shehata, Rabeh El Shesheny, Abdallah M. Samy, Ghazi Kayali, and Mohamed A. Ali. 2020. "Middle East Respiratory Syndrome Coronavirus (MERS-CoV): State of the Science" Microorganisms 8, no. 7: 991. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8070991