Functional Biodiversity of Yeasts Isolated from Colombian Fermented and Dry Cocoa Beans

 ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Raw Materials
2.2. Yeasts Enumeration and Isolation
2.3. Molecular Identification
2.4. Screening of the Yeast Strains for the Enzyme Activities
2.5. Growth at 37 °C and 46 °C in Cocoa Pulp Simulation Medium
2.6. Statistical Analysis
3. Results and Discussion
3.1. Yeast Counts and Water Activity
3.2. Identification of the Yeast Isolates
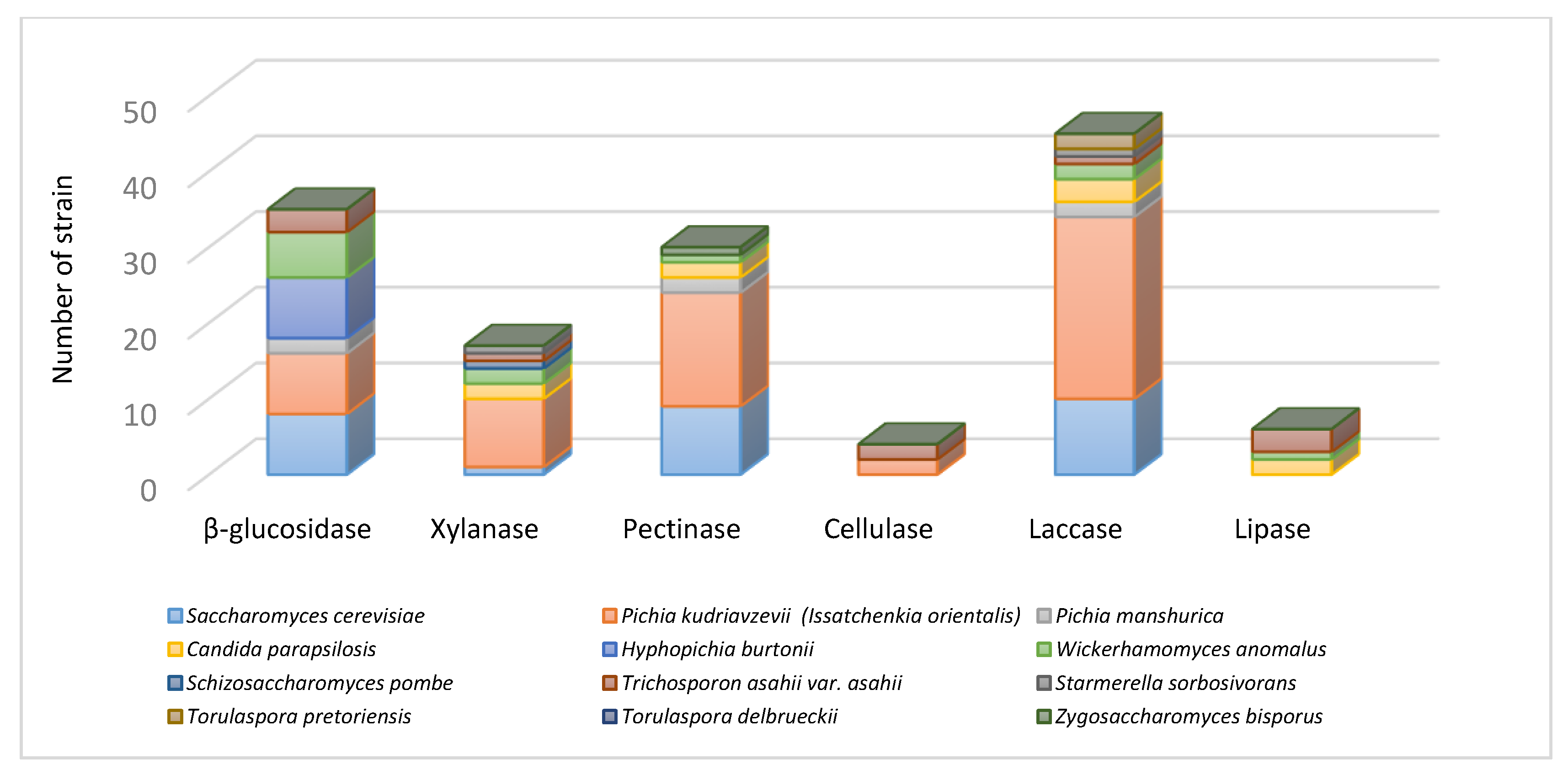
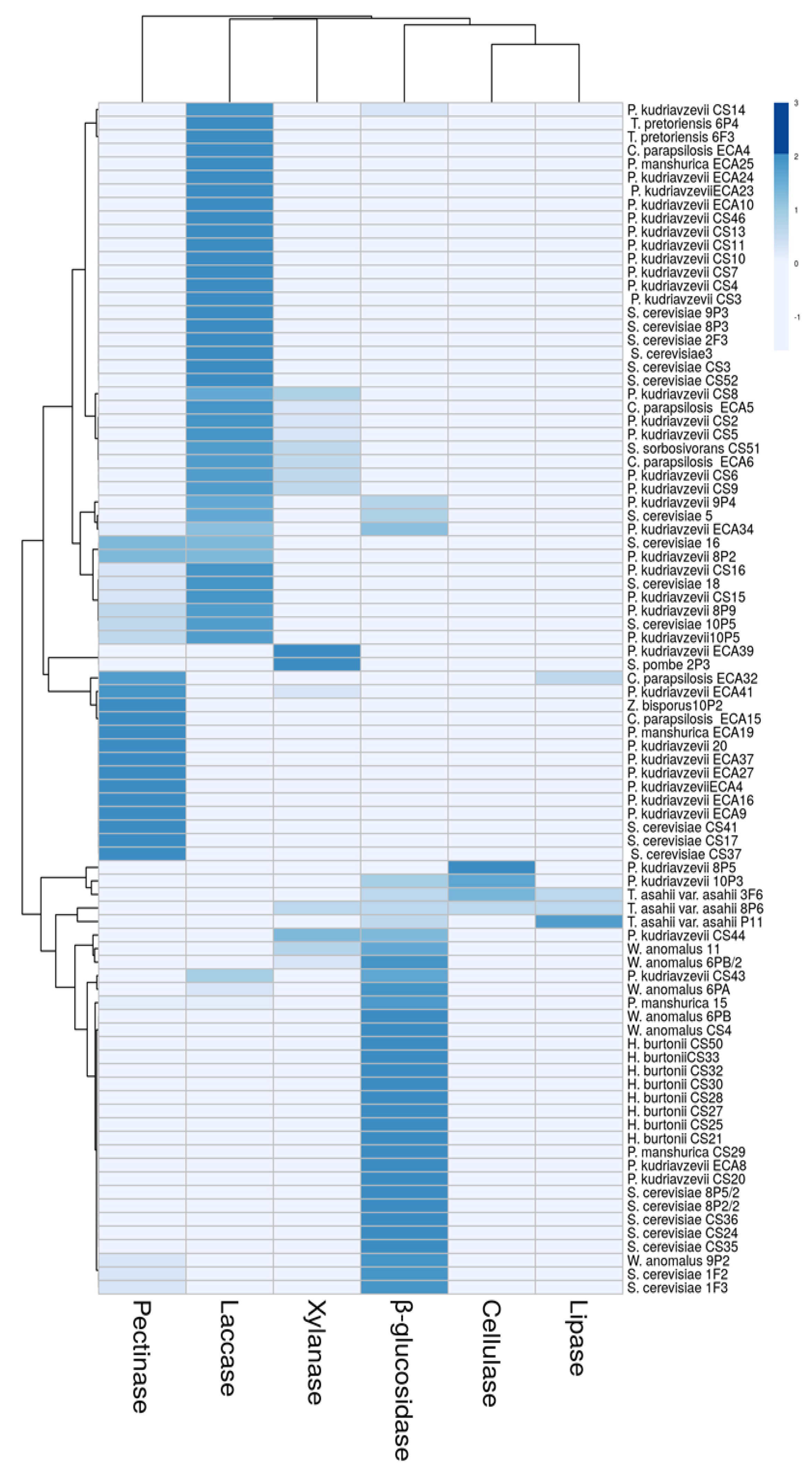
3.3. Extracellular Enzyme Activity Profile Differences among the Yeasts
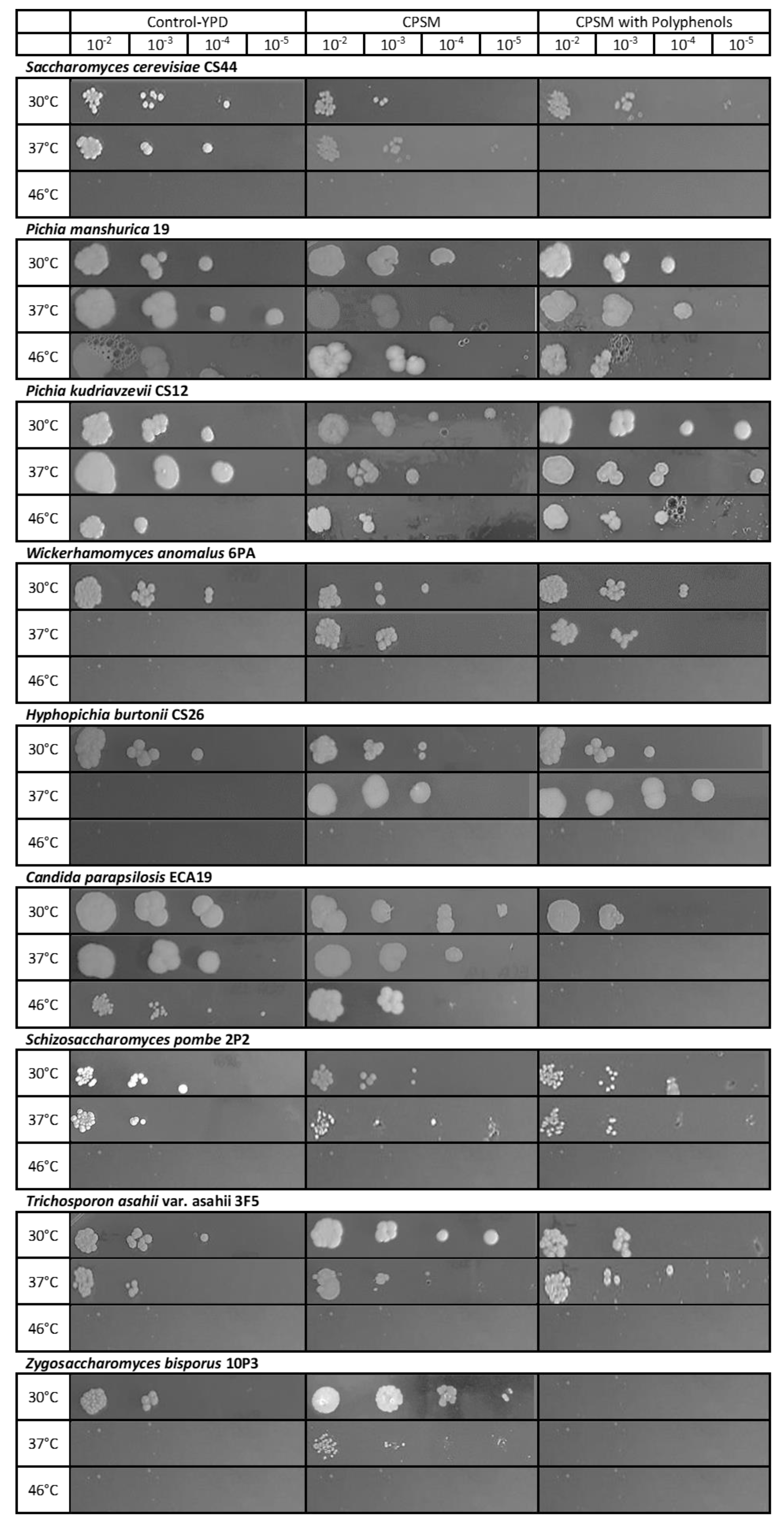
3.4. Yeast Thermotolerance Is Modulated by Polyphenols Content
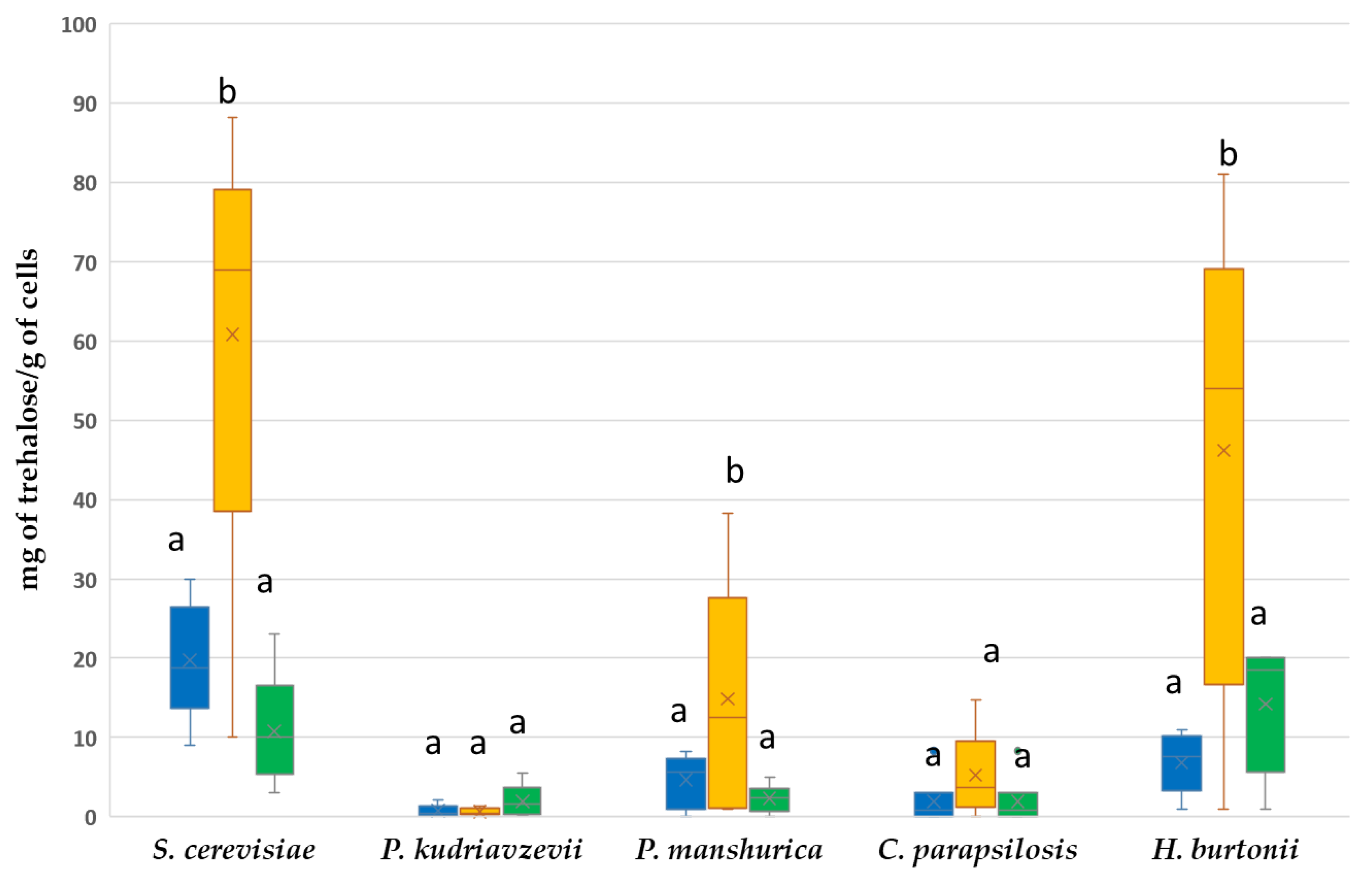
3.5. Differences in Trehalose Accumulation as a Response to Mild Heat Stress
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Afoakwa, E.O.; Quao, J.; Takrama, J.; Budu, A.S.; Saalia, F.K. Chemical composition and physical quality characteristics of Ghanaian cocoa beans as affected by pulp pre-conditioning and fermentation. J. Food Sci. Technol. 2011, 50, 1097–1105. [Google Scholar] [CrossRef] [Green Version]
- International Cocoa Organization. Quarterly Bulletin of Cocoa Statistics. Available online: http:www.icco.org (accessed on 12 December 2019).
- Ardhana, M. The microbial ecology of cocoa bean fermentations in Indonesia. Int. J. Food Microbiol. 2003, 86, 87–99. [Google Scholar] [CrossRef]
- Nigam, P.; Singh, A. Cocoa and Coffee Fermentations. In Encyclopedia of Microbiology; Elsevier BV: Amsterdam, The Netherlands, 2014; pp. 485–492. [Google Scholar]
- Pereira, G.V.D.M.; Soccol, V.T.; Soccol, C.R. Current state of research on cocoa and coffee fermentations. Curr. Opin. Food Sci. 2016, 7, 50–57. [Google Scholar] [CrossRef]
- Hansen, A.P.; Welty, R.E. Microflora of raw cacao beans. Mycopathology 1971, 44, 309–316. [Google Scholar] [CrossRef]
- Mota-Gutierrez, J.; Botta, C.; Ferrocino, I.; Giordano, M.; Bertolino, M.; Dolci, P.; Cannoni, M.; Cocolin, L. Dynamics and Biodiversity of Bacterial and Yeast Communities during Fermentation of Cocoa Beans. Appl. Environ. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maura, Y.F.; Balzarini, T.; Borges, P.C.; Evrard, P.; De Vuyst, L.; Daniel, H.-M. The environmental and intrinsic yeast diversity of Cuban cocoa bean heap fermentations. Int. J. Food Microbiol. 2016, 233, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, D.S.; Hønholt, S.; Tano-Debrah, K.; Jespersen, L. Yeast populations associated with Ghanaian cocoa fermentations analysed using denaturing gradient gel electrophoresis (DGGE). Yeast 2005, 22, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Hamdouche, Y.; Guehi, T.; Durand, N.; Kedjebo, K.B.D.; Montet, D.; Meile, J.C. Dynamics of microbial ecology during cocoa fermentation and drying: Towards the identification of molecular markers. Food Control 2015, 48, 117–122. [Google Scholar] [CrossRef]
- Ludlow, C.L.; Cromie, G.A.; Garmendia-Torres, C.; Sirr, A.; Hays, M.; Field, C.; Jeffery, E.W.; Fay, J.C.; Dudley, A.M. Independent Origins of Yeast Associated with Coffee and Cacao Fermentation. Curr. Boil. 2016, 26, 965–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrigan, W.F.; McCance, M.E. Laboratory Methods in Food and Dairy Microbiology, 1st ed.; Academic Press Inc. (London) Ltd.: London, UK, 1976; 25p. [Google Scholar]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Evol. Microbiol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Elsevier BV: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar]
- Granchi, L.; Bosco, M.; Messini, A.; Vincenzini, M. Rapid detection and quantification of yeast species during spontaneous wine fermentation by PCR-RFLP analysis of the rDNA ITS region. J. Appl. Microbiol. 1999, 87, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Sabate, J.; Cano-Lira, J.F.; Esteve-Zarzoso, B.; Guillamón, J.M. Isolation and identification of yeasts associated with vineyard and winery by RFLP analysis of ribosomal genes and mitochondrial DNA. Microbiol. Res. 2002, 157, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Guillamón, J.M.; Sabaté, J.; Barrio, E.; Cano-Lira, J.F.; Querol, A. Rapid identification of wine yeast species based on RFLP analysis of the ribosomal internal transcribed spacer (ITS) region. Arch. Microbiol. 1998, 169, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Identification of clinically important ascomycetous yeasts based on nucleotide divergence in the 5′ end of the large-subunit (26S) ribosomal DNA gene. J. Clin. Microbiol. 1997, 35, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- Chaves-Lopez, C.; Serio, A.; Ospina, J.D.; Rossi, C.; Tovar, C.D.G.; Paparella, A. Exploring the Bacterial Microbiota of Colombian Fermented Maize Dough “Masa Agria” (Maiz Añejo). Front. Microbiol. 2016, 7, 1168. [Google Scholar] [CrossRef]
- Chaves-López, C.; De Angelis, M.; Martuscelli, M.; Serio, A.; Paparella, A.; Suzzi, G. Characterization of the Enterobacteriaceae isolated from an artisanal Italian ewe’s cheese (Pecorino Abruzzese). J. Appl. Microbiol. 2006, 101, 353–360. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Mangieri, N.; Maghradze, D.; Foschino, R.; Valdetara, F.; Cantoral, J.M.; Vigentini, I. Wild Grape-Associated Yeasts as Promising Biocontrol Agents against Vitis vinifera Fungal Pathogens. Front. Microbiol. 2017, 8, 2025. [Google Scholar] [CrossRef] [Green Version]
- Pointing, S.B.; Jones, E.B.G.; Vrijmoed, L.L.P. Optimization of Laccase Production by Pycnoporus sanguineus in Submerged Liquid Culture. Mycologia 2000, 92, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Buzzini, P.; Martini, A. Extracellular enzymatic activity profiles in yeast and yeast-like strains isolated from tropical environments. J. Appl. Microbiol. 2002, 93, 1020–1025. [Google Scholar] [CrossRef]
- Bavaro, S.L.; Susca, A.; Frisvad, J.C.; Tufariello, M.; Chytiri, A.; Perrone, G.; Mita, G.; Logrieco, A.F.; Bleve, G. Isolation, Characterization, and Selection of Molds Associated to Fermented Black Table Olives. Front. Microbiol. 2017, 8, 1356. [Google Scholar] [CrossRef] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Portillo, E.; Labarca, M.; Grazziani, L.; Cros, E.; Assemat, S.; Davrieux, F.; Boulager, R. Influencia de las condiciones del tratamiento poscosecha sobre la temperatura y acidez en granos de cacao criollo (Theobroma Cacao L.). Rev. Fac. Agron. 2011, 28, 646–660. [Google Scholar]
- Ho, V.T.T.; Zhao, J.; Fleet, G.H. Yeasts are essential for cocoa bean fermentation. Int. J. Food Microbiol. 2014, 174, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.V.D.M.; Miguel, M.G.D.C.P.; Ramos, C.L.; Schwan, R.F. Microbiological and physicochemical characterization of small-scale cocoa fermentations and screening of yeast and bacterial strains to develop a defined starter culture. Appl. Environ. Microbiol. 2012, 78, 5395–5405. [Google Scholar] [CrossRef] [Green Version]
- Machuca-Guevara, J.I.; Suárez-Peña, E.A.; Darricau, E.M.; Mialhe-Matonnier, E.L. Caracterización molecular de los microorganismos presentes durante el proceso fermentativo de los granos de cacao (Theobroma cacao). Rev. Peru. Biol. 2019, 26, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Daniel, H.-M.; Vrancken, G.; Takrama, J.F.; Camu, N.; De Vos, P.; De Vuyst, L. Yeast diversity of Ghanaian cocoa bean heap fermentations. FEMS Yeast Res. 2009, 9, 774–783. [Google Scholar] [CrossRef] [Green Version]
- Almeida, S.D.F.O.D.; Silva, L.R.C.; Junior, G.C.A.C.; Oliveira, G.; Da Silva, S.H.M.; Vasconcelos, S.; Lopes, A.S. Diversity of yeasts during fermentation of cocoa from two sites in the Brazilian Amazon. Acta Amaz. 2019, 49, 64–70. [Google Scholar] [CrossRef]
- Samagaci, L.; Ouattara, H.G.; Niamké, S.; Lemaire, M. Pichia kudrazevii and Candida nitrativorans are the most well-adapted and relevant yeast species fermenting cocoa in Agneby-Tiassa, a local Ivorian cocoa producing region. Food Res. Int. 2016, 89, 773–780. [Google Scholar] [CrossRef]
- Visintin, S.; Alessandria, V.; Valente, A.; Dolci, P.; Cocolin, L. Molecular identification and physiological characterization of yeasts, lactic acid bacteria and acetic acid bacteria isolated from heap and box cocoa bean fermentations in West Africa. Int. J. Food Microbiol. 2016, 216, 69–78. [Google Scholar] [CrossRef]
- Jespersen, L.; Nielsen, D.S.; Hønholt, S.; Jakobsen, M. Occurrence and diversity of yeasts involved in fermentation of West African cocoa beans. FEMS Yeast Res. 2005, 5, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Papalexandratou, Z.; De Vuyst, L. Assessment of the yeast species composition of cocoa bean fermentations in different cocoa-producing regions using denaturing gradient gel electrophoresis. FEMS Yeast Res. 2011, 11, 564–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwenninger, S.M.; Leischtfeld, S.F.; Gantenbein-Demarchi, C. High-throughput identification of the microbial biodiversity of cocoa bean fermentation by MALDI-TOF MS. Lett. Appl. Microbiol. 2016, 63, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Koff, O.; Samagaci, L.; Goualie, B.; Niamke, S.; Koffi, O. Diversity of Yeasts Involved in Cocoa Fermentation of Six Major Cocoa-Producing Regions in Ivory Coast. Eur. Sci. J. ESJ 2017, 13, 496–516. [Google Scholar] [CrossRef] [Green Version]
- Ooi, T.; Ting, A.; Siow, L. Influence of selected native yeast starter cultures on the antioxidant activities, fermentation index and total soluble solids of Malaysia cocoa beans: A simulation study. Lebensm.-Wiss. Technol. 2020, 122, 108977. [Google Scholar] [CrossRef]
- Koné, M.K.; Guehi, T.S.; Durand, N.; Ban-Koffi, L.; Berthiot, L.; Tachon, A.F.; Brou, K.; Boulanger, R.; Montet, D. Contribution of predominant yeasts to the occurrence of aroma compounds during cocoa bean fermentation. Food Res. Int. 2016, 89, 910–917. [Google Scholar] [CrossRef]
- Chaves-Lopez, C.; Serio, A.; Osorio-Cadavid, E.; Paparella, A.; Suzzi, G. Volatile compounds produced in wine by Colombian wild Saccharomyces cerevisiae strains. Ann. Microbiol. 2009, 59, 733–740. [Google Scholar] [CrossRef]
- Schwan, R.F. Cocoa Fermentations Conducted with a Defined Microbial Cocktail Inoculum. Appl. Environ. Microbiol. 1998, 64, 1477–1483. [Google Scholar] [CrossRef] [Green Version]
- Schwan, R.F.; Wheals, A.E. The Microbiology of Cocoa Fermentation and its Role in Chocolate Quality. Crit. Rev. Food Sci. Nutr. 2004, 44, 205–221. [Google Scholar] [CrossRef]
- Masoud, W.; Jespersen, L. Pectin degrading enzymes in yeasts involved in fermentation of Coffea arabica in East Africa. Int. J. Food Microbiol. 2006, 110, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Du Plessis, H.; Hoff, J.; Hart, R.; Ndimba, B.; Jolly, N.; Du Toit, M. Characterisation of Non-Saccharomyces Yeasts Using Different Methodologies and Evaluation of their Compatibility with Malolactic Fermentation. South Afr. J. Enol. Vitic. 2017, 38, 46–63. [Google Scholar] [CrossRef]
- Quijada-Morín, N.; Garcia, F.; Lambert, K.; Walker, A.-S.; Tiers, L.; Viaud, M.; Sauvage, F.-X.; Hirtz, C.; Saucier, C. Strain effect on extracellular laccase activities from Botrytis cinerea. Aust. J. Grape Wine Res. 2017, 24, 241–251. [Google Scholar] [CrossRef]
- De Souza, J.P.; Prado, C.D.D.; Eleutherio, E.C.; Bonatto, R.C.; Malavazi, I.; Cunha, A.F.; Paulino, J.D.S. Improvement of Brazilian bioethanol production—Challenges and perspectives on the identification and genetic modification of new strains of Saccharomyces cerevisiae yeasts isolated during ethanol process. Fungal Boil. 2018, 122, 583–591. [Google Scholar] [CrossRef]
- Pongcharoen, P.; Chawneua, J.; Tawong, W. High temperature alcoholic fermentation by new thermotolerant yeast strains Pichia kudriavzevii isolated from sugarcane field soil. Agric. Nat. Resour. 2018, 52, 511–518. [Google Scholar] [CrossRef]
- Chamnipa, N.; Thanonkeo, S.; Klanrit, P.; Thanonkeo, P. The potential of the newly isolated thermotolerant yeast Pichia kudriavzevii RZ8-1 for high-temperature ethanol production. Braz. J. Microbiol. 2017, 49, 378–391. [Google Scholar] [CrossRef]
- Phong, H.X.; Klanrit, P.; Dung, N.T.P.; Yamada, M.; Thanonkeo, P. Isolation and characterization of thermotolerant yeasts for the production of second-generation bioethanol. Ann. Microbiol. 2019, 69, 765–776. [Google Scholar] [CrossRef]
- El-Gamal, M.S.; Desouky, S.E.-S.; Abdel-Rahman, M.A.; Khattab, A.-R.M. High-Temperature Citric Acid Production from Sugar Cane Molasses using A Newly Isolated Thermotolerant Yeast Strain, Candida parapsilosis NH-3. Int. J. Adv. Res. Biol. Sci. 2018, 5, 187–211. [Google Scholar] [CrossRef]
- Deák, T. Handbook of food spoilage yeasts, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2007; p. 352. [Google Scholar]
- Golden, D.; Beuchat, L. Interactive effects of solutes, potassium sorbate and incubation temperature on growth, heat resistance and tolerance to freezing of Zygosaccharomyces rouxii. J. Appl. Bacteriol. 1992, 73, 524–530. [Google Scholar] [CrossRef]
- Peláez-Soto, A.; Roig, P.; Martínez-Culebras, P.V.; Fernández-Espinar, M.T.; Gil, J.V. Proteomic Analysis of Saccharomyces cerevisiae Response to Oxidative Stress Mediated by Cocoa Polyphenols Extract. Molecules 2020, 25, 452. [Google Scholar] [CrossRef] [Green Version]
- Todorović, V.; Milenkovic, M.; Vidovic, B.; Todorovic, Z.; Sobajic, S. Correlation between Antimicrobial, Antioxidant Activity, and Polyphenols of Alkalized/Nonalkalized Cocoa Powders. J. Food Sci. 2017, 82, 1020–1027. [Google Scholar] [CrossRef]
- Vilaça, R.; Mendes, V.; Mendes, M.V.; Carreto, L.; Amorim, M.A.; De Freitas, V.; Moradas-Ferreira, P.; Mateus, N.; Costa, V. Quercetin Protects Saccharomyces cerevisiae against Oxidative Stress by Inducing Trehalose Biosynthesis and the Cell Wall Integrity Pathway. PLoS ONE 2012, 7, e45494. [Google Scholar] [CrossRef]
- Granek, J.; Magwene, P.M. Environmental and Genetic Determinants of Colony Morphology in Yeast. PLoS Genet. 2010, 6, e1000823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, S. Osmotic Stress Signaling and Osmoadaptation in Yeasts. Microbiol. Mol. Boil. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomé, P.E. Cell wall involvement in the glycerol response to high osmolarity in the halotolerant yeast Debaryomyces hansenii. Antonie van Leeuwenhoek 2006, 91, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, S.A.; Hirasawa, T.; Shimizu, H. Differential importance of trehalose accumulation in Saccharomyces cerevisiae in response to various environmental stresses. J. Biosci. Bioeng. 2010, 109, 262–266. [Google Scholar] [CrossRef]
- Pan, D.; Wiedemann, N.; Kammerer, B. Heat stress-induced metabolic remodeling in Saccharomyces cerevisiae. Metabolites 2019, 9, 266. [Google Scholar] [CrossRef] [Green Version]
- Walker, G.M.; Van Dijck, P. Physiological and Molecular Responses of Yeasts to the Environment; Springer Science and Business Media LLC: Berlin, Germany, 2006; pp. 111–152. [Google Scholar]
- Mensonides, F.I.C.; Hellingwerf, K.J.; De Mattos, M.J.T.; Brul, S. Multiphasic adaptation of the transcriptome of Saccharomyces cerevisiae to heat stress. Food Res. Int. 2013, 54, 1103–1112. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Production Stage | Yeast Counts Log CFU/g | pH | aw | Moisture (%) |
|---|---|---|---|---|
| Before drying | 6.59 ± 0.46 a | 5.3 ± 0.37 a | 0.98 ± 0.21 a | 42.6 ± 3.2 a |
| After drying | 2.73 ± 1.71 b | 5.1 ± 0.66 a | 0.52 ± 0.11 b | 3.4 ± 1.3 a |
| Strains | Closest Relative | Identity (%) | Accession Number |
|---|---|---|---|
| ECA30, ECA34, ECA32, ECA29, ECA31, ECA4, ECA3, ECA5, ECA19, ECA14. | Candida parapsilosis | 99.83% | MH911007.1 |
| CS27, CS31, CS26, CS24, CS20, CS29, CS32, CS49. | Hyphopichia burtonii | 100% | CP024760.1 |
| CS2, CS3, CS42, CS43, 7, ECA20, 3F3, 8P1, ECA6, 9P3, 1P8, CS1, CS4, CS19, CS18, CS17, CS6, CS 9, CS10, CS7, CS 8, CS 14, CS 15, 3F4, 10P2, 10P4, CS13, CS12, CS45, ECA33, ECA22, ECA23, ECA26, ECA38, ECA37, ECA39, O5?, ECA21, 19, ECA40 ECA41, ECA8, ECA7, ECA6, ECA10, ECA9, ECA11, ECA13, ECA12, ECA15, ECA16, ECA36, 8P8, 8P4, 8P9, 8P10. | Pichia kudriavzevii | 100% | AY529502.1 |
| 3F ECA25, ECA24, 1, 14, ECA35, ECA18, ECA17, 2F1, ECA28, 16, 12, 19, CS28/3. | Pichia manshurica | 100% | MK034750.1 |
| 6P2, 6F1. | Torulaspora delbrueckii | 99.83% | MT255021.1 |
| 6F2, 6P3. | Torulaspora pretoriensis | 99.43% | KY109883.1 |
| P10, 8P5, 3F5. | Trichosporon asahii var. asahii | 99.55% | KY109934.1 |
| 1P6, 2F3, 2F2, 1F1, 2F1, 1F2, 1P4, 9P2, CS39, CS44, 3, 5, 4, 8P7, 2, CS36, CS35, CS34, CS37, CS40, 10P4, CS1, CS23, ECA1, ECA2, CS25, 3F2, 8P2/1, 3P2, 3P1, 3P3, 15, 8P2, 8P6, CS41, 17, CS16, 8P5/1, CS11. | Saccharomyces cerevisiae | 99.66% | JX141382.1 |
| CS50, 8. | Starmerella sorbosivorans | 99.55% | NG_060827.1 |
| 6PA, 6PB, 6P1/b, 9P1, 6PB/1, 10, CS2/1. | Wickerhamomyces anomalus | 99.66% | MT255020.1 |
| 10P1, 10P3. | Zygosaccharomyces bisporus | 99.49% | NG058447.1 |
| CS48, CS47, CS46, 2P2. | Schizosaccharomyces pombe | 99.84% | KY109602.1 |
| Yeast Species | Frequency a | Abundance b (%) |
|---|---|---|
| Candida parapsilosis | 5/19 | 6.6 |
| Hyphopichia burtonii | 2/19 | 5.18 |
| Pichia kudriavzevii | 15/19 | 40.00 |
| Pichia manshurica | 7/19 | 8.88 |
| Saccharomyces cerevisiae | 15/19 | 28.14 |
| Schizosaccharomyces pombe | 2/19 | 2.77 |
| Starmerella sorbosivorans | 2/19 | 1.48 |
| Torulaspora delbrueckii | 2/19 | 1.38 |
| Torulaspora pretoriensis | 1/19 | 1.48 |
| Trichosporon asahii var. asahii | 2/19 | 2.01 |
| Wickerhamomyces anomalus | 4/19 | 4.4 |
| Zygosaccharomyces bisporus | 2/19 | 1.48 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado-Ospina, J.; Triboletti, S.; Alessandria, V.; Serio, A.; Sergi, M.; Paparella, A.; Rantsiou, K.; Chaves-López, C. Functional Biodiversity of Yeasts Isolated from Colombian Fermented and Dry Cocoa Beans. Microorganisms 2020, 8, 1086. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8071086
Delgado-Ospina J, Triboletti S, Alessandria V, Serio A, Sergi M, Paparella A, Rantsiou K, Chaves-López C. Functional Biodiversity of Yeasts Isolated from Colombian Fermented and Dry Cocoa Beans. Microorganisms. 2020; 8(7):1086. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8071086
Chicago/Turabian StyleDelgado-Ospina, Johannes, Samantha Triboletti, Valentina Alessandria, Annalisa Serio, Manuel Sergi, Antonello Paparella, Kalliopi Rantsiou, and Clemencia Chaves-López. 2020. "Functional Biodiversity of Yeasts Isolated from Colombian Fermented and Dry Cocoa Beans" Microorganisms 8, no. 7: 1086. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8071086