Effect of Lactic Acid Bacteria on the Lipid Profile of Bean-Based Plant Substitute of Fermented Milk



Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Preparation of Bean-Based Plant Substitute of Fermented Milks
2.3. Microbiological Analysis
2.4. pH Determination
2.5. Lipid Extraction
2.6. Analysis of the Fatty Acid Profile
2.7. Analysis of the Positional Distribution of Fatty Acids in Triacylglycerols
2.8. Statistical Analysis
3. Results and Discussion
3.1. Lactic Acid Bacteria Population
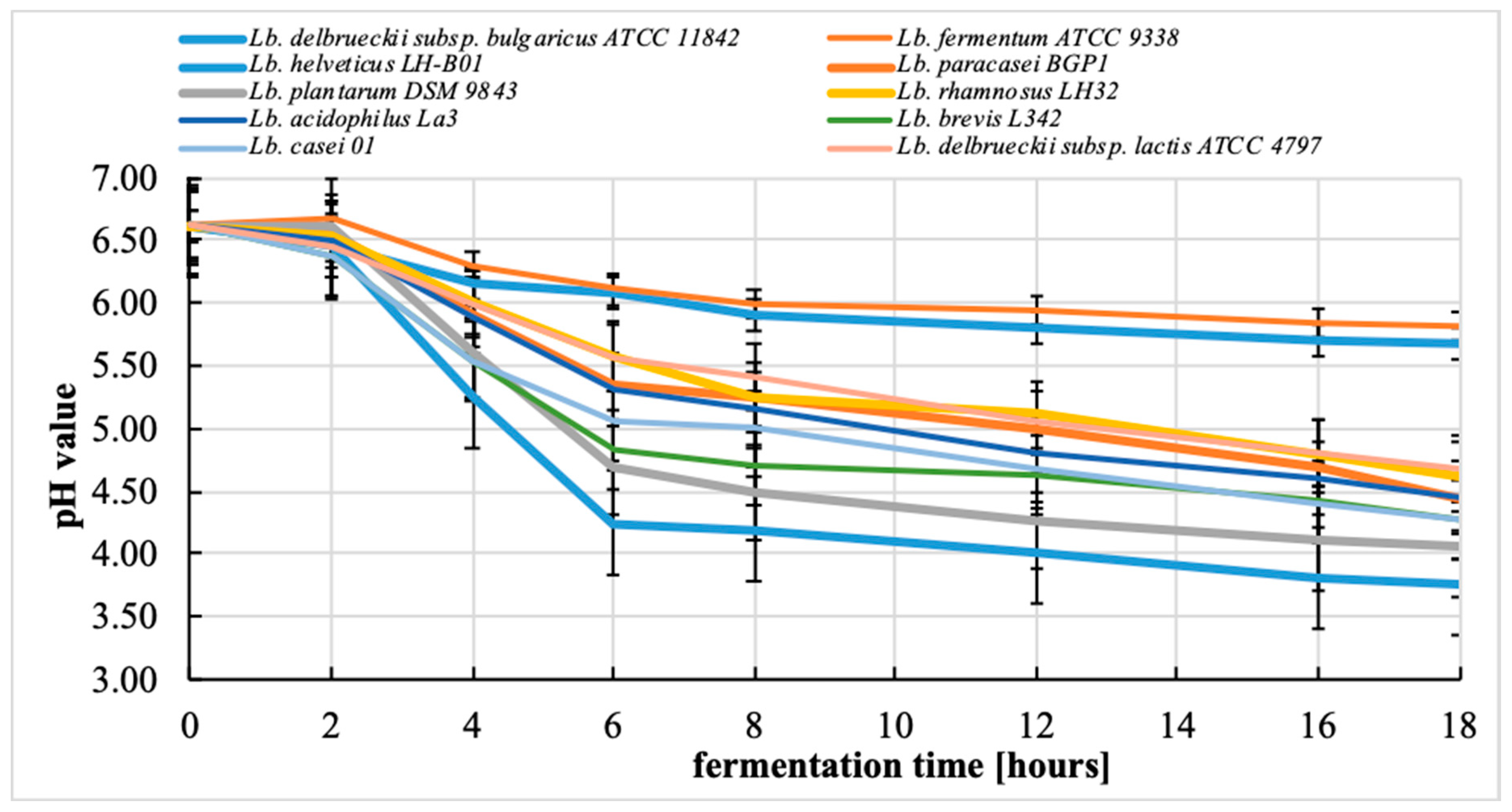
3.2. Changes in pH during the Fermentation Process
3.3. Determination of the Fatty Acid Profile
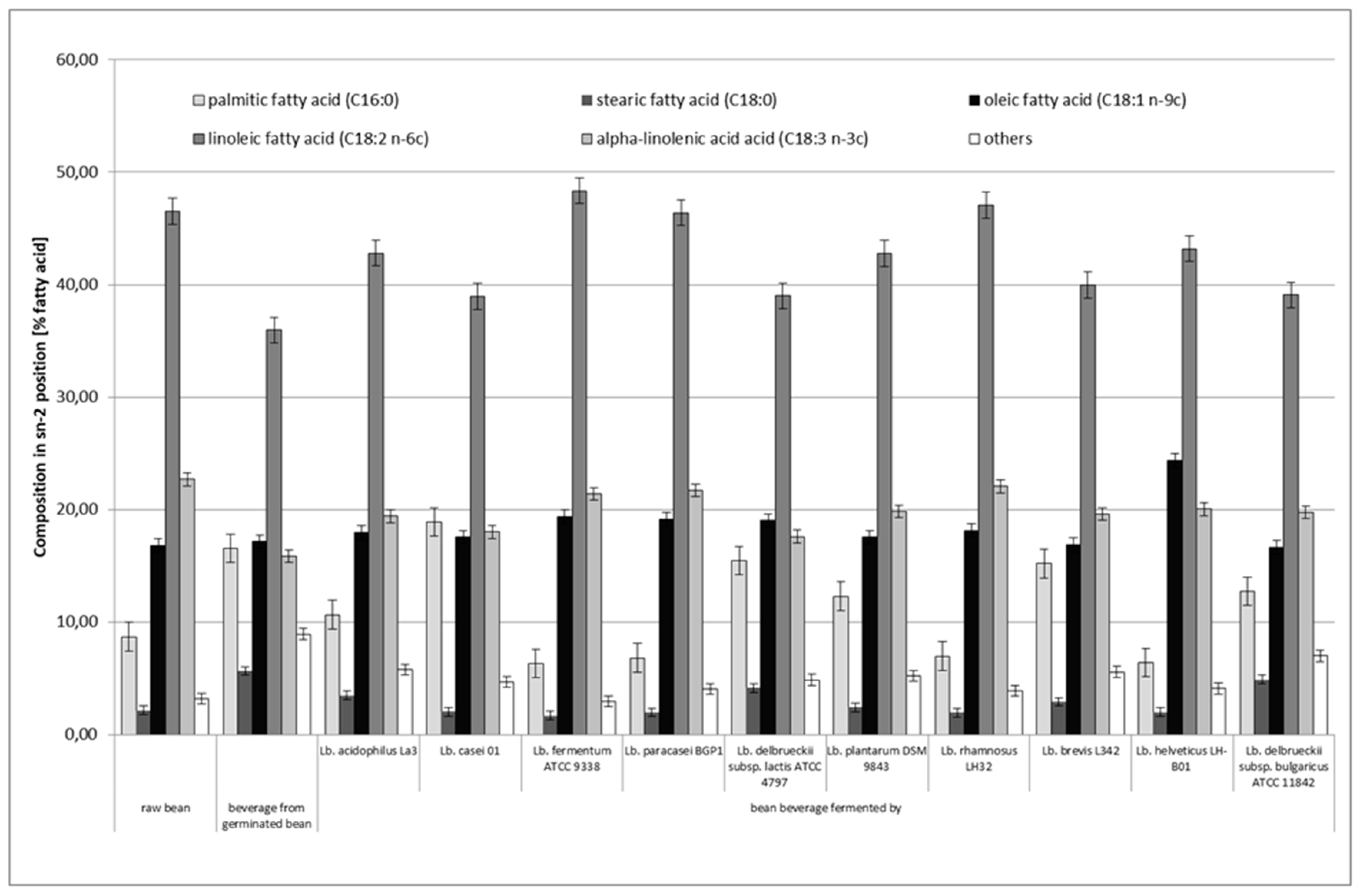
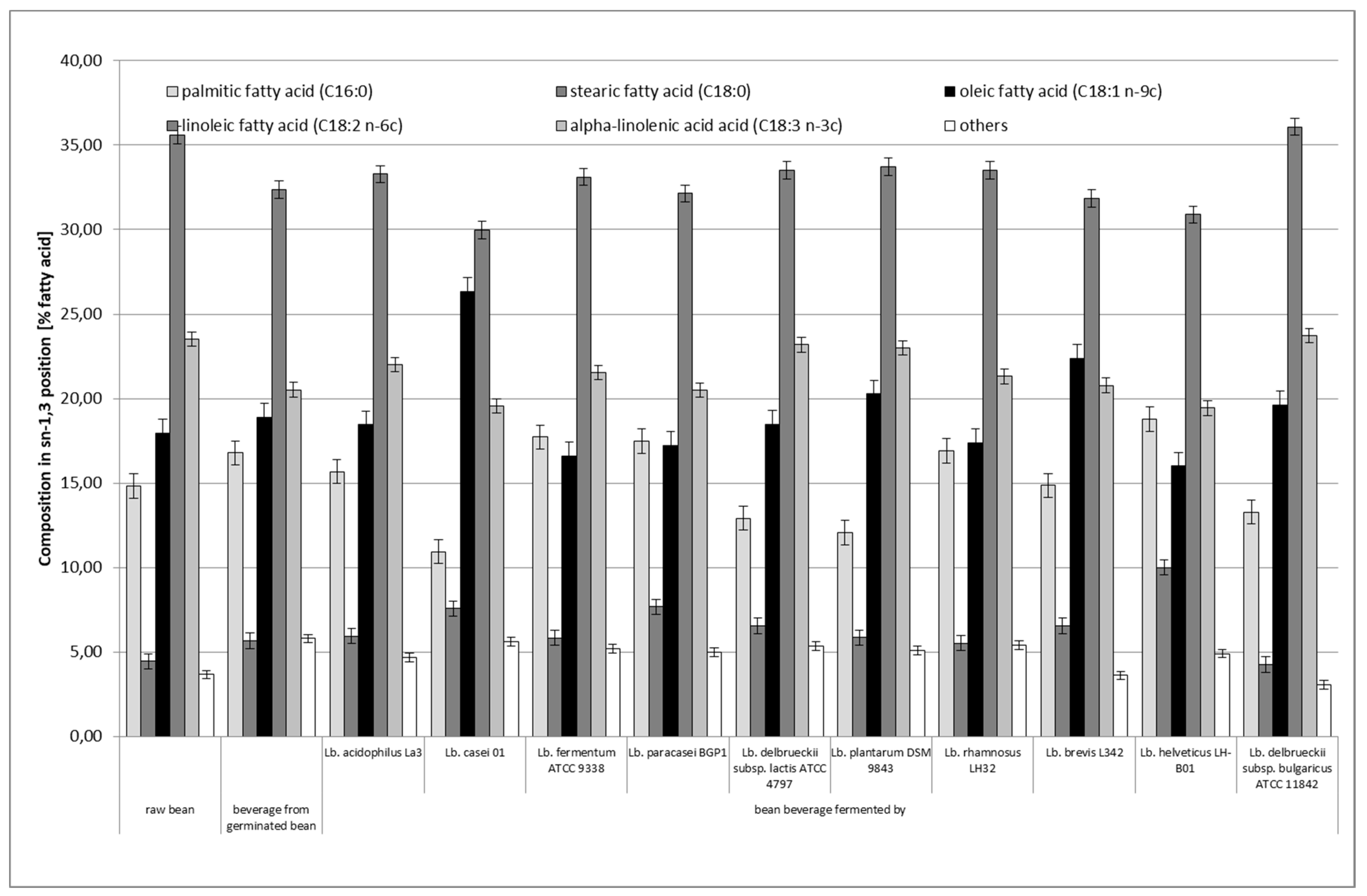
3.4. Distribution of Fatty Acids in sn–2 and sn–1,3 Positions of Triacylglycerol (TAG)
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yoshida, H.; Tomiyama, Y.; Yoshida, N.; Shibata, K.; Mizushina, Y. Regiospecific profiles of fatty acids in triacylglycerols and phospholipids from adzuki beans (Vigna angularis). Nutrients 2010, 2, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchenak, M.; Lamri–Senhadji, M. Nutritional quality of legumes, and their role in cardiometabolic risk prevention: A review. J. Med. Food 2013, 16, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Abdel–Rahman, E.A.; El–Fishawy, F.A.; El–Geddawy, M.A.; Kurz, T.; El–Rify, M.N. The changes in the lipid composition of mung bean seeds as affected by processing methods. Int. J. Food Eng. 2007, 3. [Google Scholar] [CrossRef]
- Anwar, F.; Latif, S.; Przybylski, R.; Sultana, B.; Ashraf, M. Chemical composition and antioxidant activity of seeds of different cultivars of mung bean. J. Food Sci. 2007, 72, 503–510. [Google Scholar] [CrossRef]
- Ryan, E.; Galvin, K.; O’Connor, T.P.; Maguire, A.R.; O’Brien, N.M. Phytosterol, squalene, tocopherol content and fatty acid profile of selected seeds, grains, and legumes. Plant Foods Hum. Nutr. 2007, 65, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Tomiyama, Y.; Tanaka, M.; Mizushina, Y. Distribution of fatty acids in triacylglycerols and phospholipids from peas (Pisum sativum L.). J. Sci. Food Agric. 2007, 87, 2709–2714. [Google Scholar] [CrossRef]
- Yoshida, H.; Saiki, M.; Yoshida, N.; Tomiyama, Y.; Mizushina, Y. Fatty acid distribution in triacylglycerols and phospholipids of broad beans (Vicia faba). Food Chem. 2009, 112, 924–928. [Google Scholar] [CrossRef]
- Megat–Rusydi, M.R.; Noraliza, C.W.; Azrina, A.; Zulkhairi, A. Nutritional changes in germinated legumes and rice varieties. Int. Food Res. J. 2011, 18, 705–713. [Google Scholar]
- Ziarno, M.; Zaręba, D.; Maciejak, M.; Veber, A.L. The impact of dairy starter cultures on selected qualitative properties of functional fermented beverage prepared from germinated White Kidney Beans. J. Food Nutr. Res. 2019, 2, 167–176. [Google Scholar]
- Eggers, L.F.; Schwudke, D. Liquid Extraction: Folch. In Encyclopedia of Lipidomics; Wenk, M., Ed.; Springer Science+Business Media: Dordrecht, The Netherlands, 2016. [Google Scholar]
- Yoshida, H.; Tomiyama, Y.; Mizushina, Y. Characterization in the fatty acid distributions of triacyloglicerols and phospholipids in kidney beans (Phaseolus vulgaris L.). J. Food Lipids 2005, 12, 169–180. [Google Scholar] [CrossRef]
- Zaręba, D.; Ziarno, M. The viability of yogurt bacteria in selected plant beverages. Zesz. Probl. Post. Nauk Roln. 2017, 591, 87–96. [Google Scholar] [CrossRef]
- Beasley, S.; Tuorila, H.; Saris, P.E.J. Fermented soymilk with a monoculture of Lactococcus lactis. Int. J. Food Microbiol. 2003, 81, 159–162. [Google Scholar] [CrossRef]
- Zaręba, D.; Ziarno, M.; Obiedziński, M. Viability of yoghurt bacteria and probiotic strains in models of fermented and non–fermented milk. Med. Weter. 2008, 64, 1007–1011, (In Polish, Abstract In English). [Google Scholar]
- Food and Agriculture Organization of the United Nations/World Health Organization (FAO/WHO). Health and Nutritional Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria. 2001. Available online: http://www.who.int/foodsafety/publications/fs_management/probiotics/en/ (accessed on 29 July 2020).
- Gan, R.Y.; Shah, N.P.; Wang, M.F.; Lui, W.Y.; Corke, H. Lactobacillus plantarum WCFS1 fermentation differentially affects antioxidant capacity and polyphenol content in mung bean (Vigna radiata) and soya bean (Glycine max) milks. J. Food Process. Preserv. 2016, 41, e12944. [Google Scholar] [CrossRef]
- Limon, R.I.; Penas, E.; Torino, M.I.; Martínez–Villaluenga, C.; Dueńas, M.; Frias, J. Fermentation enhances the content of bioactive compounds in kidney bean extracts. Food Chem. 2015, 172, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Boucher, I.; Vadeboncoeur, C.; Moineau, S. Characterization of genes involved in the metabolism of α–galactosides by Lactococcus raffinolactis. Appl. Environ. Microbiol. 2003, 69, 4049–4056. [Google Scholar] [CrossRef] [Green Version]
- Machielsen, R.; Siezen, R.J.; van Hijum, S.A.F.T.; van Hylckama Vlieg, J.E.T. Molecular description and industrial potential of Tn6098 conjugative transfer conferring α–galactoside metabolism in Lactococcus lactis. Appl. Environ. Microbiol. 2011, 77, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.L.; Kraidej, L.; Hesseltine, C.W. Lactic acid fermentation of soybean milk. J. Milk Food Technol. 1974, 37, 71–73. [Google Scholar] [CrossRef]
- Garro, M.S.; Valdez, G.F.; Giori, G.S. Temperature effect on the biological activity of Bifidobacterium longum CRL 849 and Lactobacillus fermentum CRL 251 in pure and mixed cultures grown in soymilk. Food Microbiol. 2004, 21, 511–518. [Google Scholar] [CrossRef]
- Santos, C.C.A.A.; Libeck, B.S.; Schwan, R.F. Co–culture fermentation of peanut–soy milk for the development of a novel functional beverage. Int. J. Food Microbiol. 2014, 186, 32–41. [Google Scholar] [CrossRef]
- Grela, E.R.; Gunter, K.D. Fatty acid composition and tocopherol content of some legume seeds. Anim. Feed Sci. Technol. 1995, 52, 325–331. [Google Scholar] [CrossRef]
- Zaręba, D. Fatty acid profile of soya milk fermented by various bacteria strains of lactic acid fermentation. Żywność Nauka Technol. Jakość 2009, 6, 59–71, (In Polish, Abstract In English). [Google Scholar]
- Collins, Y.F.; McSweeney, P.L.H.; Wilkinson, M.G. Lipolysis and catabolism of fatty acids in cheese: A review of current knowledge. Int. Dairy J. 2003, 13, 841–866. [Google Scholar] [CrossRef]
- Marilley, L.; Casey, M.G. Flavours of cheese products: Metabolic pathways, analytical tools and identification of producing strains. Int. J. Food Microbiol. 2004, 90, 139–159. [Google Scholar] [CrossRef]
- Holland, R.; Liu, S.-Q.; Crow, V.L.; Delabre, M.-L.; Lubbers, M.; Bennett, M.; Norris, G. Esterases of lactic acid bacteria and cheese flavour: Milk fat hydrolysis, alcoholysis and esterification. Int. Dairy J. 2005, 15, 711–718. [Google Scholar] [CrossRef]
- Treimo, J.; Vegarud, G.; Langsrud, T.; Rudi, K. Use of DNA quantification to measure growth and autolysis of Lactococcus and Propionibacterium spp. in mixed populations. Appl. Environ. Microbiol. 2006, 72, 6174–6182. [Google Scholar] [CrossRef] [Green Version]
- Lortal, S.; Chapot-Chartier, M.-P. Role, mechanisms and control of lactic acid bacteria lysis in cheese. Int. Dairy J. 2005, 15, 857–871. [Google Scholar] [CrossRef] [Green Version]
- Bzducha, A.; Obiedziński, M. Changes in CLA concentration in cheese models with the addition of Bifidobacterium animalis subsp. lactis and Lactobacillus acidophilus. Bromatol. Chem. Toxicol. 2009, 42, 241–246, (In Polish, Abstract In English). [Google Scholar]
- Akalin, A.S.; Tokugoglu, O.; Gönc, S.; Aycan, S. Occurrence of conjugated linoleic acid in probiotic yoghurts supplemented with fructooligosaccharide. Int. Dairy J. 2007, 17, 1089–1095. [Google Scholar] [CrossRef]
- Pérez Pulido, R.; Ben Ornar, N.; Abriouel, H.; Lucas López, R.; Martínez Cañamero, M.; Guyot, J.P.; Gálvez, A. Characterization of lactobacilli isolated from caper berry fermentations. J. Appl. Microbiol. 2007, 102, 583–590. [Google Scholar] [CrossRef]
- Angeles, A.G.; Marth, E.H. Growth and activity of lactic acid bacteria in soy milk. III. Lipolytic activity. J. Milk Food Technol. 1971, 34, 69–75. [Google Scholar] [CrossRef]
- Bryś, J.; Wirkowska–Wojdyła, M.; Górska, A.; Ostrowska–Ligęza, E.; Biardzka, M.; Ciemniewska-Żytkiewicz, H.; Bryś, A. Oils rich in Omega-3 fatty acids as a potential source of polyunsaturated fatty acids in the diet for infants and young children. Post. Tech. Przetw. Spoż. 2016, 1, 47–52, (In Polish, Abstract In English). [Google Scholar]
- Takagi, S.; Yoshida, H. Microwave heating influences on fatty acid distributions of triacylglycerols and phospholipids in hypocotyl of soybeans (Glycine max L.). Food Chem. 1999, 66, 345–351. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Bean–Based Beverages Fermented by: | Before Fermentation [log10 CFU/g] | After Fermentation [log10 CFU/g] |
|---|---|---|
| Lb. acidophilus La3 | 8.4 ± 0.3 | 8.3 ± 0.2 |
| Lb. brevis L342 | 8.6 ± 0.2 | 8.2 ± 0.3 |
| Lb. casei 01 | 7.6 ± 0.1 | 7.8 ± 0.4 |
| Lb. delbrueckii subsp. bulgaricus ATCC 11842 | 7.5 ± 0.2 | 7.6 ± 0.2 |
| Lb. delbrueckii subsp. lactis ATCC 4797 | 7.6 ± 0.3 | 7.5 ± 0.2 |
| Lb. fermentum ATCC 9338 | 7.8 ± 0.4 | 8.1 ± 0.2 |
| Lb. helveticus LH–B01 | 8.1 ± 0.2 | 8.0 ± 0.1 |
| Lb. paracasei BGP1 | 8.2 ± 0.3 | 8.2 ± 0.3 |
| Lb. plantarum DSM 9843 | 8.7 ± 0.2 | 8.5 ± 0.2 |
| Lb. rhamnosus LH32 | 8.1 ± 0.1 | 8.2 ± 0.2 |
| Fatty Acid Share [%] in the Total Pool of Fatty Acids in the Samples: | Palmitic C16:0 | Stearic C18:0 | Oleic C18:1 n–9c | Linoleic C 18:2 n–6c | α–Linolenic C18:3 n–3 |
|---|---|---|---|---|---|
| Raw white bean | 12.79 ± 0.57 a | 3.68 ± 0.16 a | 17.59 ±0.78 a | 39.26 ±1.74 d | 23.25 ±1.03 c |
| Bean–based beverages before fermentation | 16.72 ± 0.74 d | 5.66 ± 0.25 c | 18.30 ±0.81 a | 33.59 ±1.49 a | 18.97 ±0.84 a |
| Bean–based beverages fermented by: | |||||
| Lactobacillus delbrueckii subsp. bulgaricus ATCC 11842 | 13.10 ± 0.57 a.b | 4.46 ± 0.20 b | 18.62 ± 0.82 a | 37.12 ± 1.64 b.c | 22.42 ± 0.99 c |
| Lactobacillus fermentum ATCC 9338 | 13.93 ± 0.62 b.c | 4.45 ± 0.20 b | 17.52 ± 0.78 a | 38.23 ± 1.69 c.d | 21.51 ± 0.95 b |
| Lactobacillus plantarum DSM 9843 | 12.15 ± 0.54 a | 4.70 ± 0.21 b | 19.38 ± 0.86 b | 36.78 ± 1.63 b | 21.95 ± 0.97 b.c |
| Lactobacillus casei 01 | 13.59 ± 0.60 b | 5.73 ± 0.25 c | 23.41 ± 1.04 d | 32.99 ± 1.46 a | 19.05 ± 0.84 a |
| Lactobacillus delbrueckii subsp. lactis ATCC 4797 | 13.77 ± 0.61 b | 5.74 ± 0.25 c | 18.67 ± 0.83 a | 35.37 ± 1.56 a.b | 21.33 ± 0.94 b |
| Lactobacillus brevis L342 | 14.97 ± 0.66 c | 5.34 ± 0.24 c | 20.54 ± 0.91 c | 34.58 ± 1.53 a | 20.40 ± 0.90 a.b |
| Lactobacillus helveticus LH–B01 | 14.66 ± 0.65 c | 7.33 ± 0.32 d | 18.79 ± 0.83 a.b | 35.02 ± 1.55 a | 19.65 ± 0.87 a |
| Lactobacillus paracasei BGP1 | 13.92 ± 0.62 b | 5.77 ± 0.26 c | 17.86 ± 0.79 a | 36.93 ± 1.63 b | 20.91 ± 0.93 a.b |
| Lactobacillus acidophilus La3 | 14.00 ± 0.62 c | 5.11 ± 0.23 c | 18.28 ± 0.81 a | 36.49 ± 1.61 b | 21.16 ± 0.94 b |
| Lactobacillus rhamnosus LH32 | 13.59 ± 0.60 b | 4.32 ± 0.19 b | 17.63 ± 0.78 a | 38.06 ±1.68 c | 21.67 ± 0.96 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziarno, M.; Bryś, J.; Parzyszek, M.; Veber, A. Effect of Lactic Acid Bacteria on the Lipid Profile of Bean-Based Plant Substitute of Fermented Milk. Microorganisms 2020, 8, 1348. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8091348
Ziarno M, Bryś J, Parzyszek M, Veber A. Effect of Lactic Acid Bacteria on the Lipid Profile of Bean-Based Plant Substitute of Fermented Milk. Microorganisms. 2020; 8(9):1348. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8091348
Chicago/Turabian StyleZiarno, Małgorzata, Joanna Bryś, Mateusz Parzyszek, and Anna Veber. 2020. "Effect of Lactic Acid Bacteria on the Lipid Profile of Bean-Based Plant Substitute of Fermented Milk" Microorganisms 8, no. 9: 1348. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8091348