Both Soil Bacteria and Soil Chemical Property Affected the Micropredator Myxobacterial Community: Evidence from Natural Forest Soil and Greenhouse Rhizosphere Soil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Field Sample Collection
2.2. Measurement of Soil pH and Organic Matter Content
2.3. Soil DNA Extraction and HiSeq Sequencing
2.4. Bioinformatic and Statistical Analysis
2.5. Myxobacterial Sequence Collection and Data Analysis from the Mesocosm Experiment under Greenhouse Conditions
3. Results
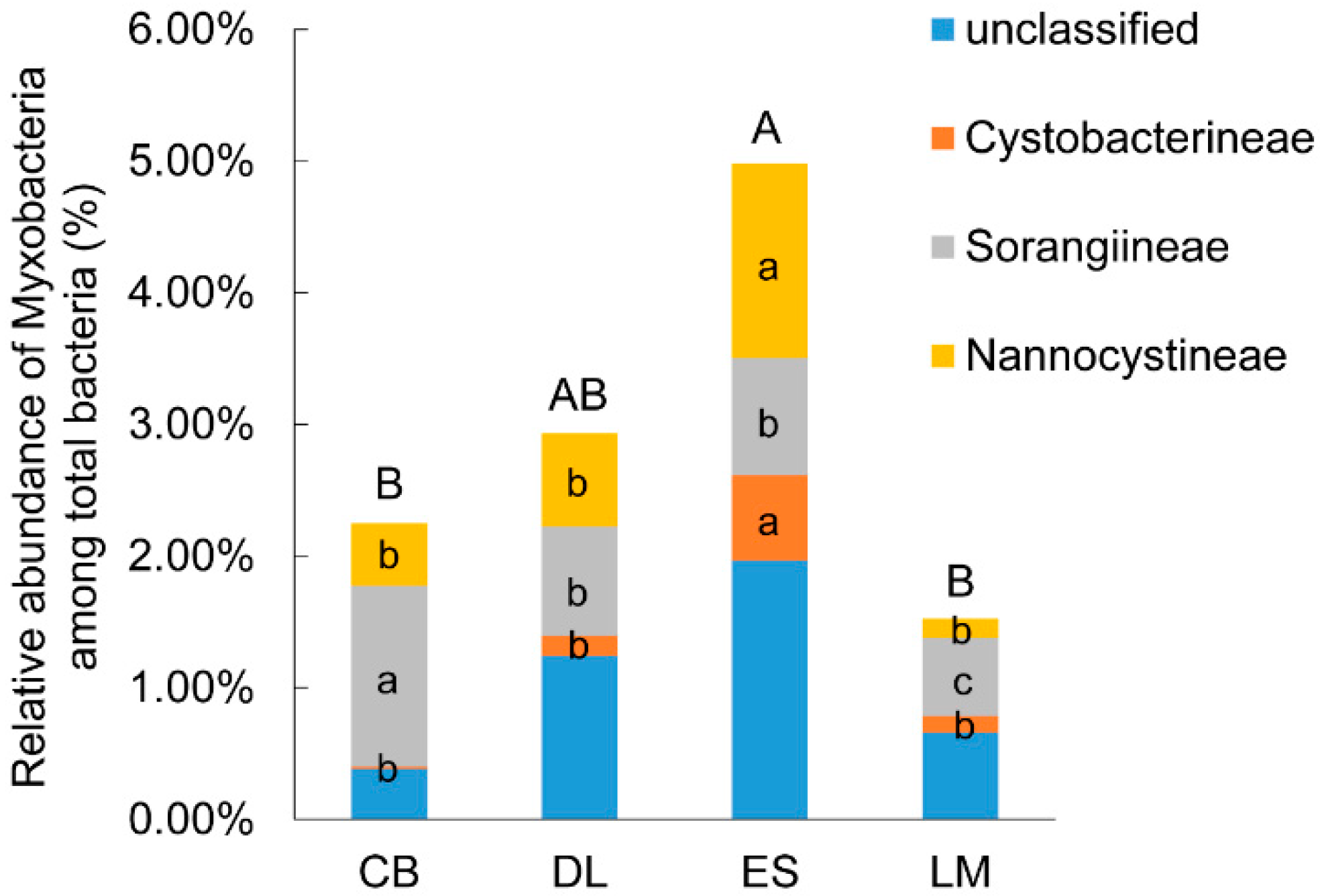
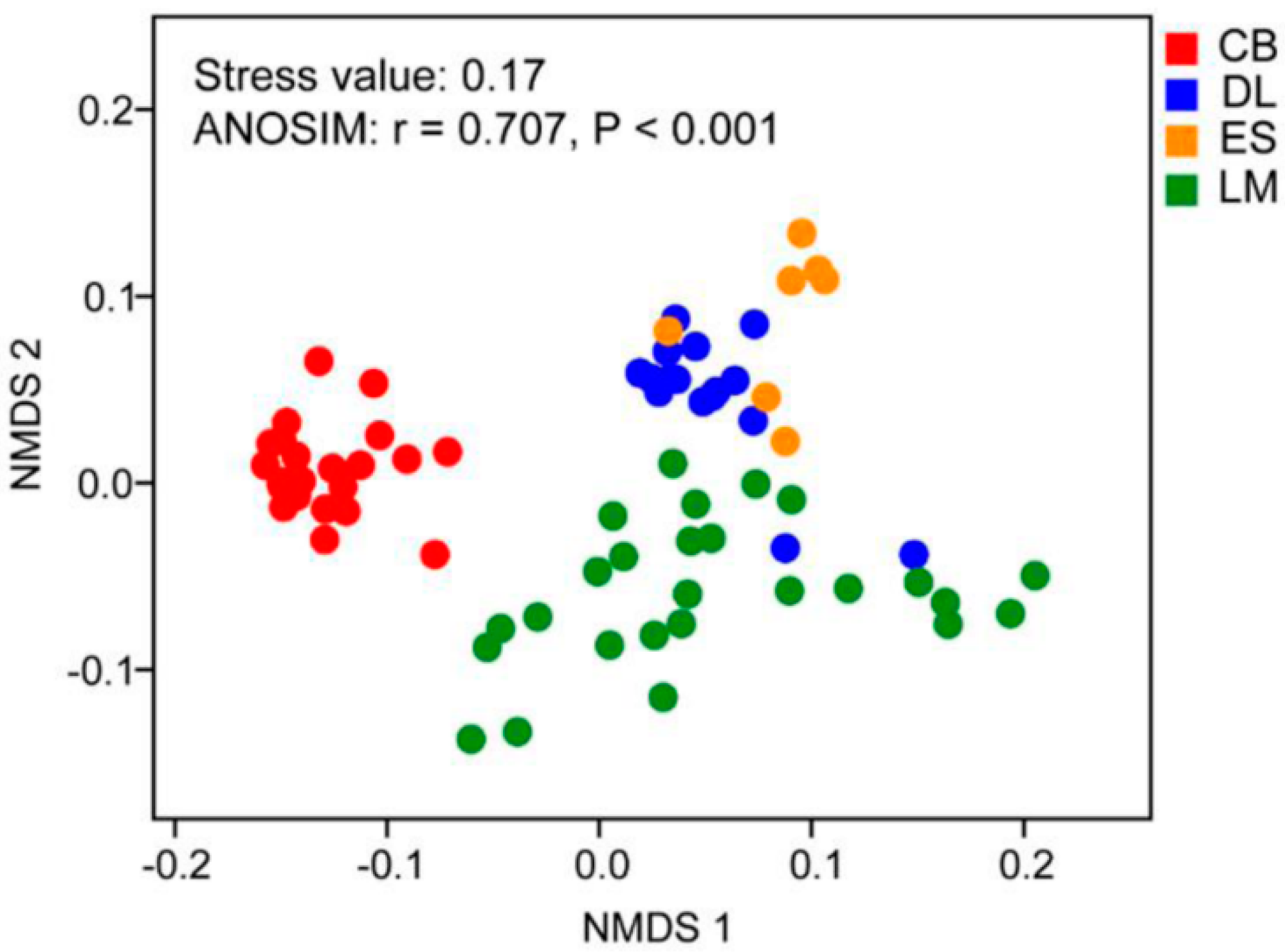
3.1. Myxobacterial Abundance and Community Composition in Subtropical and Tropical Forest Soil
3.2. Forest Soil Bacteria as a Biotic Factor Correlated with Myxobacteria
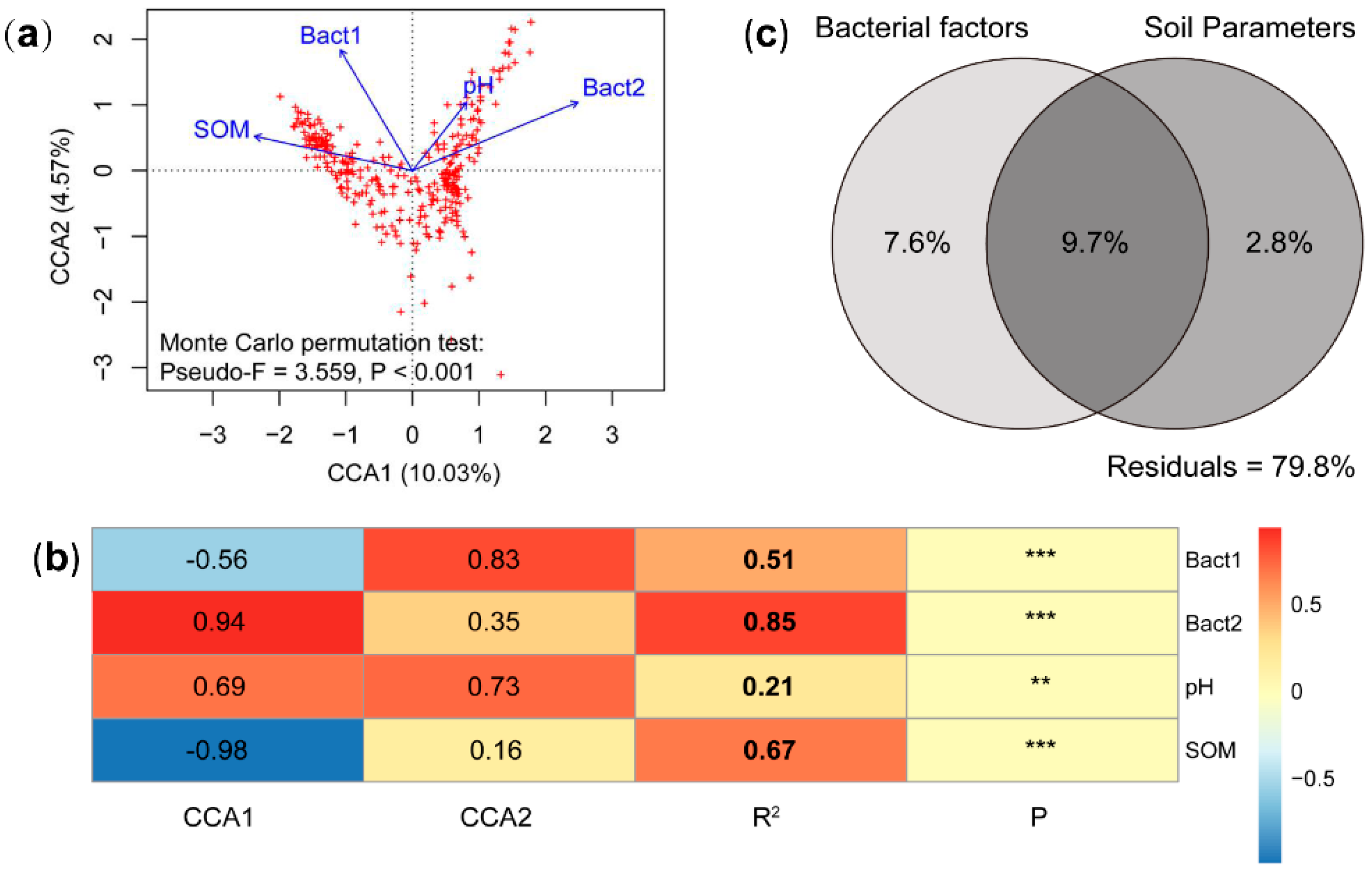
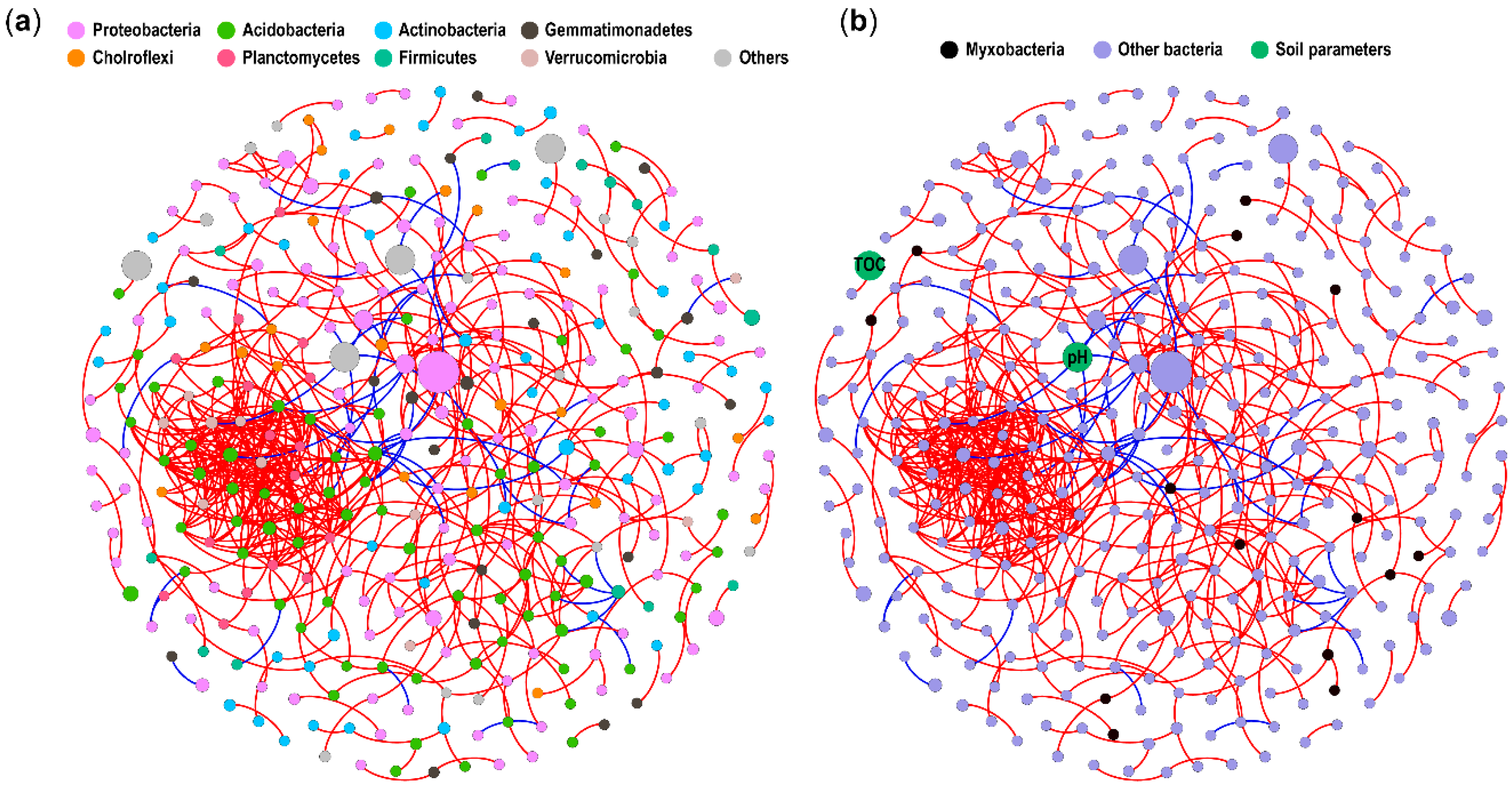
3.3. The Contribution of Biotic and Abiotic Factors in Relation to Forest Myxobacterial Community
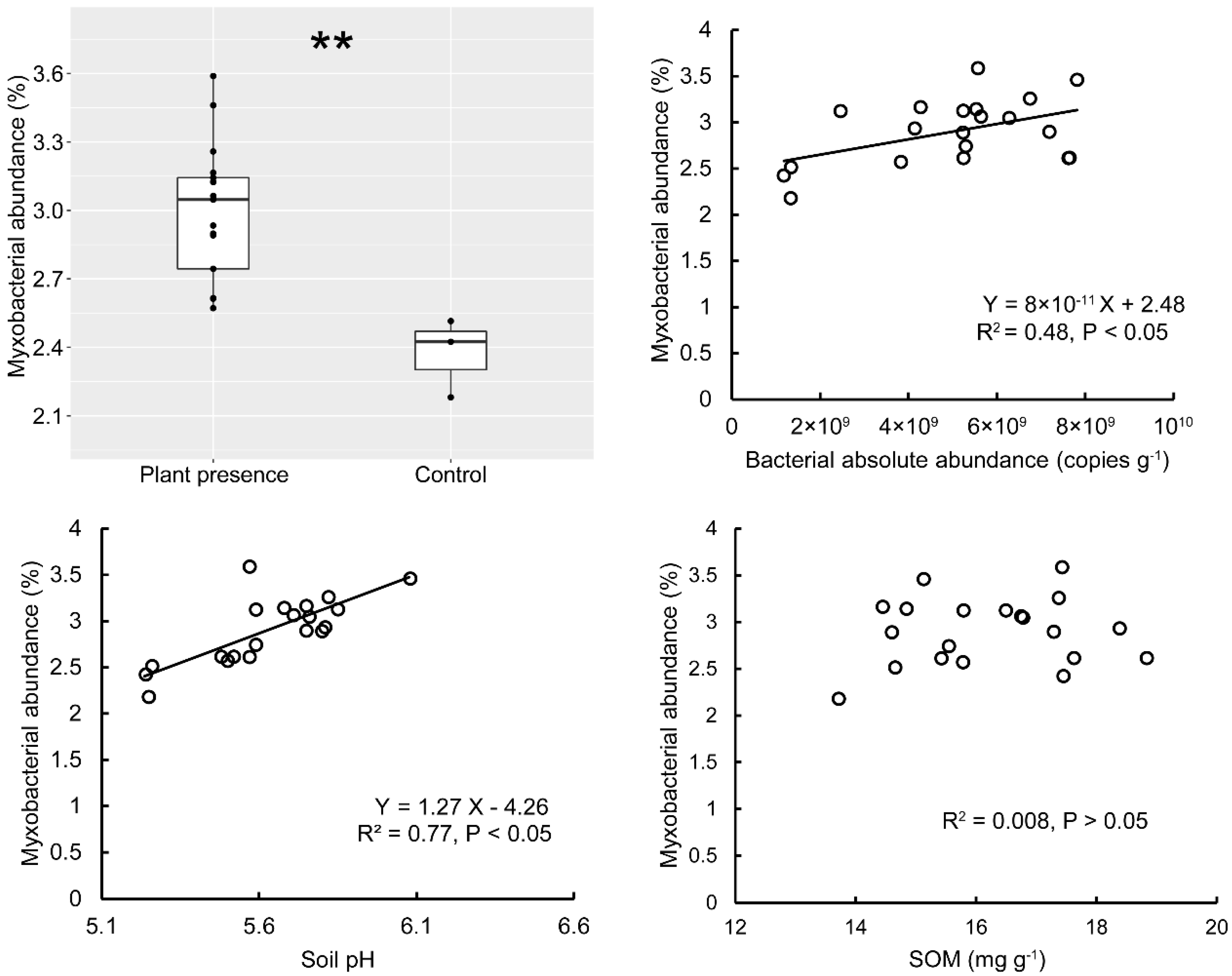
3.4. The Influence of Bacterial Abundance and Interactions on Myxobacteria in the Greenhouse Mesocosm Experiment
4. Discussion
4.1. Myxobacteria Relative Abundance and Community Composition in Subtropical and Tropical Forest Soil from South China
4.2. Effect of Soil Bacteria on Myxobacterial Community
4.3. Effect of Soil Abiotic Factors on Myxobacterial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Muñoz-Dorado, J.; Marcos-Torres, F.J.; García-Bravo, E.; Moraleda-Muñoz, A.; Pérez, J. Myxobacteria: moving, killing, feeding, and surviving together. Front. Microbiol. 2016, 7, 781. [Google Scholar] [CrossRef] [Green Version]
- Pérez, J.; Moraleda-Muñoz, A.; Marcos-Torres, F.J.; Muñoz-Dorado, J. Bacterial predation: 75 years and counting! Environ. Microbiol. 2016, 18, 766–779. [Google Scholar] [CrossRef] [PubMed]
- Dawid, W. Biology and global distribution of myxobacteria in soils. FEMS Microbiol. Rev. 2000, 24, 403–427. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yao, Q.; Zhu, H. Meta-16S rRNA gene phylogenetic reconstruction reveals the astonishing diversity of cosmopolitan myxobacteria. Microorganisms 2019, 7, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohr, K. Diversity of myxobacteria-we only see the tip of the iceberg. Microorganisms 2018, 6, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.; Kato, C.; Zhou, X.; Wu, Z.; Sato, T.; Li, Y. Phylogeographic separation of marine and soil myxobacteria at high levels of classification. ISME J. 2010, 4, 1520–1530. [Google Scholar] [CrossRef] [Green Version]
- Mohr, K.I.; Stechling, M.; Wink, J.; Wilharm, E.; Stadler, M. Comparison of myxobacterial diversity and evaluation of isolation success in two niches: Kiritimati Island and German compost. MicrobiologyOpen 2016, 5, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Mohr, K.I.; Zindler, T.; Wink, J.; Wilharm, E.; Stadler, M. Myxobacteria in high moor and fen: an astonishing diversity in a neglected extreme habitat. Microbiologyopen 2017, 6, e464. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Bailey, M.; Whiteley, A.S. The bacterial biogeography of British soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, G.T.; Bates, S.T.; Eilers, K.G.; Lauber, C.L.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. The under-recognized dominance of Verrucomicrobia in soil bacterial communities. Soil Biol. Biochem. 2011, 43, 1450–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naether, A.; Foesel, B.U.; Naegele, V.; Wüst, P.K.; Weinert, J.; Bonkowski, M.; Alt, F.; Oelmann, Y.; Polle, A.; Lohaus, G.; et al. Environmental factors affect acidobacterial communities below the subgroup level in grassland and forest soils. Appl. Environ. Microbiol. 2012, 78, 7398–7406. [Google Scholar] [CrossRef] [Green Version]
- Walsh, C.M.; Gebert, M.J.; Delgado-Baquerizo, M.; Maestre, F.T.; Fierer, N. A global survey of mycobacterial diversity in soil. Appl. Environ. Microbiol. 2019, 85, e01180-19. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Li, S.; Li, W.; Jiang, D.; Han, K.; Wu, Z.; Li, Y. Myxobacterial community is a predominant and highly diverse bacterial group in soil niches. Environ. Microbiol. Rep. 2014, 6, 45–56. [Google Scholar] [CrossRef]
- Huntley, S.; Hamann, N.; Wegener-Feldbrugge, S.; Treuner-Lange, A.; Kube, M.; Reinhardt, R.; Klages, S.; Muller, R.; Ronning, C.M.; Nierman, W.C.; et al. Comparative genomic analysis of fruiting body formation in Myxococcales. Mol. Biol. Evol. 2011, 28, 1083–1097. [Google Scholar] [CrossRef] [Green Version]
- Lueders, T.; Kindler, R.; Miltner, A.; Friedrich, M.W.; Kaestner, M. Identification of bacterial micropredators distinctively active in a soil microbial food web. Appl. Environ. Microbiol. 2006, 72, 5342–5348. [Google Scholar] [CrossRef] [Green Version]
- Murase, J.; Frenzel, P. A methane-driven microbial food web in a wetland rice soil. Environ. Microbiol. 2007, 9, 3025–3034. [Google Scholar] [CrossRef]
- Morgan, A.D.; MacLean, R.C.; Hillesland, K.L.; Velicer, G.J. Comparative analysis of Myxococcus predation on soil bacteria. Appl. Environ. Microbiol. 2010, 76, 6920–6927. [Google Scholar] [CrossRef] [Green Version]
- Müller, S.; Strack, S.N.; Hoefler, B.C.; Straight, P.D.; Kearns, D.B.; Kirby, J.R. Bacillaene and sporulation protect Bacillus subtilis from predation by Myxococcus xanthus. Appl. Environ. Microbiol. 2014, 80, 5603–5610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, J.; Jiménez-Zurdo, J.I.; Martínez-Abarca, F.; Millán, V.; Shimkets, L.J.; Muñoz-Dorado, J. Rhizobial galactoglucan determines the predatory pattern of Myxococcus xanthus and protects Sinorhizobium meliloti from predation. Environ. Microbiol. 2014, 16, 2341–2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Athar, R.; Zheng, G.; Williams, H.N. Prey bacteria shape the community structure of their predators. ISME J. 2011, 5, 1314–1322. [Google Scholar] [CrossRef] [Green Version]
- Cordero, O.X.; Datta, M.S. Microbial interactions and community assembly at microscales. Curr. Opin. Microbiol. 2016, 31, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, A.; Angel, R.; Veraart, A.J.; Daebeler, A.; Jia, Z.; Kim, S.Y.; Kerckhof, F.; Boon, N.; Bodelier, P.L.E. Biotic interactions in microbial communities as modulators of biogeochemical processes: methanotrophy as a model system. Front. Microbiol. 2016, 7, 1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zengler, K.; Zaramela, L.S. The social network of microorganisms-how auxotrophies shape complex communities. Nat. Rev. Microbiol. 2018, 16, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhou, G.; Liu, S.; Zhang, D.; Liu, S.; Li, J.; Zhou, C. Dependence of soil respiration on soil temperature and soil moisture in successional forests in Southern China. J. Integr. Plant Biol. 2006, 48, 654–663. [Google Scholar] [CrossRef]
- Yao, W.; Ma, Y.; Chen, F.; Xiao, Z.; Shu, Z.; Chen, L.; Xiao, W.; Liu, J.; Jiang, L.; Zhang, S. Analysis of Ice Storm Impact on and Post-Disaster Recovery of Typical Subtropical Forests in Southeast China. Remote Sens. 2020, 12, 164. [Google Scholar] [CrossRef] [Green Version]
- Fellowes, J.R.; Chan, B.P.L.; Ssi-Chit, N.; Lau, M.W.N.; Siu, G.L.P. Report of rapid biodiversity assessments at Diaoluoshan National Forest Park, Southeast Hainan, China, 23–28 May 1999. South China For. Biodivers. Surv. Rep. Ser. (Online Simpl. Version) 2002, 23, 1–27. [Google Scholar]
- Zheng, X.; Wei, J.; Sun, W.; Li, R.; Liu, S.; Dai, H. Ethnobotanical study on medicinal plants around Limu Mountains of Hainan Island, China. J. Ethnopharmacol. 2013, 148, 964–974. [Google Scholar] [CrossRef]
- Qiu, P.; Xu, S.; Xie, G.; Tang, B.; Hua, B.; Yu, L. Analysis of the ecological vulnerability of the western Hainan Island based on its landscape pattern and ecosystem sensitivity. Acta Ecol. Sin. 2007, 27, 1257–1264. [Google Scholar]
- Tiessen, H.; Moir, J.O. Total and organic carbon. In Soil Sampling and Methods of Analysis; Carter, M.R., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 187–199. [Google Scholar]
- Zhou, Y.; Qin, Y.; Liu, X.; Feng, Z.; Zhu, H.; Yao, Q. Soil bacterial function associated with stylo (legume) and bahiagrass (grass) is affected more strongly by soil chemical property than by bacterial community composition. Front. Microbiol. 2019, 10, 798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhu, H.; Fu, S.; Yao, Q. Variation in soil microbial community structure associated with different legume species is greater than that associated with different grass species. Front. Microbiol. 2017, 8, 1007. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; Xu, Z.Z.; Kightley, E.P.; Thompson, L.R.; Hyde, E.R.; Gonzalez, A.; et al. Deblur rapidly resolves single-nucleotide community sequence patterns. MSystems 2017, 2, e00191-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Zhang, X.; Johnston, E.R.; Barberán, A.; Ren, Y.; Lü, X.; Han, X. Decreased plant productivity resulting from plant group removal experiment constrains soil microbial functional diversity. Glob. Chang. Biol. 2017, 23, 4318–4332. [Google Scholar] [CrossRef]
- Oksane, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “Vegan”: Community Ecology Package. R Package Version 2.5-4. 2017. Available online: https://CRAN.R-project.org/package=vegan/ (accessed on 25 July 2019).
- Fuhrman, J.A. Microbial community structure and its functional implications. Nature 2009, 459, 193–199. [Google Scholar] [CrossRef]
- Barberan, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Jiang, Y.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. ICWSM 2009, 8, 361–362. [Google Scholar]
- Rønn, R.; McCaig, A.E.; Griffiths, B.S.; Prosser, J.I. Impact of protozoan grazing on bacterial community structure in soil microcosms. Appl. Environ. Microbiol. 2002, 68, 6094–6105. [Google Scholar] [CrossRef] [Green Version]
- Embree, M.; Liu, J.K.; Al-Bassam, M.M.; Zengler, K. Networks of energetic and metabolic interactions define dynamics in microbial communities. Proc. Natl. Acad. Sci. USA 2015, 12, 15450–15455. [Google Scholar] [CrossRef] [Green Version]
- Petters, S.; Soellinger, A.; Bengtsson, M.M.; Urich, T. The soil microbial food web revisited with metatranscriptomics-predatory myxobacteria as keystone taxon? bioRxiv 2018, 373365. [Google Scholar] [CrossRef] [Green Version]
- Garcia, R.; Müller, R. The family Myxococcaceae. In The Prokaryotes: Deltaproteobacteria and Epsilonproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer-Verlag: Berlin, Germany, 2014; pp. 191–212. [Google Scholar]
- Wu, Z.; Jiang, D.; Li, P.; Li, Y. Exploring the diversity of myxobacteria in a soil niche by myxobacteria-specific primers and probes. Environ. Microbiol. 2005, 7, 1602–1610. [Google Scholar] [CrossRef]
- Pasternak, Z.; Pietrokovski, S.; Rotem, O.; Gophna, U.; Lurie-Weinberger, M.N.; Jurkevitch, E. By their genes ye shall know them: genomic signatures of predatory bacteria. ISME J. 2013, 7, 756–769. [Google Scholar] [CrossRef] [Green Version]
- Kramer, S.; Dibbern, D.; Moll, J.; Huenninghaus, M.; Koller, R.; Krueger, D.; Marhan, S.; Urich, T.; Wubet, T.; Bonkowski, M.; et al. Resource partitioning between bacteria, fungi, and protists in the detritusphere of an agricultural soil. Front. Microbiol. 2016, 7, 1524. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lueders, T. Micropredator niche differentiation between bulk soil and rhizosphere of an agricultural soil depends on bacterial prey. FEMS Microbiol. Ecol. 2017, 9, fix103. [Google Scholar] [CrossRef]
- Garcia, R.; La Clair, J.; Müller, R. Future directions of marine myxobacterial natural product discovery inferred from metagenomics. Mar. Drugs 2018, 16, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabe, S.; Sakai, Y.; Abe, K.; Yokota, A. Diversity of Ktedonobacteria with Actinomycetes-like morphology in terrestrial environments. Microbes Environ. 2017, 32, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Kang, S.; Schadt, C.W.; Garten, C.T. Spatial scaling of functional gene diversity across various microbial taxa. Proc. Natl. Acad. Sci. USA 2008, 105, 7768–7773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierer, N. Embracing the unknown: disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Shi, Y.; Grogan, P.; Sun, H.; Xiong, J.; Yang, Y.; Zhou, J.; Chu, H. Multi-scale variability analysis reveals the importance of spatial distance in shaping Arctic soil microbial functional communities. Soil Biol. Biochem. 2015, 86, 126–134. [Google Scholar] [CrossRef]
- Shi, Y.; Li, Y.; Xiang, X.; Sun, R.; Yang, T.; He, D.; Zhang, K.; Ni, Y.; Zhu, Y.; Adams, J.M.; et al. Spatial scale affects the relative role of stochasticity versus determinism in soil bacterial communities in wheat fields across the North China Plain. Microbiome 2018, 6, 1–12. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Zhang, P.; Xue, K.; Liang, Y.; Van Nostrand, J.D.; Yang, Y.; He, Z.; Wu, L.; Stahl, D.A.; et al. Stochasticity, succession, and environmental perturbations in a fluidic ecosystem. Proc. Natl. Acad. Sci. USA 2014, 111, E836–E845. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Myxobacterial Abundance | Myxobacterial Community Composition | |||
|---|---|---|---|---|
| r | p | r | p | |
| Proteobacteria | −0.021 | 0.865 | 0.297 | <0.001 |
| Acidobacteria | −0.495 | <0.001 | 0.216 | <0.001 |
| Actinobacteria | −0.112 | 0.351 | 0.169 | <0.001 |
| Bacteroidetes | 0.316 | 0.007 | 0.185 | <0.001 |
| Chloroflexi | 0.255 | 0.032 | 0.242 | <0.001 |
| Firmicutes | 0.415 | <0.001 | 0.156 | <0.001 |
| Gemmatimonadetes | 0.694 | <0.001 | 0.106 | 0.005 |
| Verrucomicrobia | −0.063 | 0.602 | 0.05 | 0.121 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Zhang, X.; Yao, Q.; Zhu, H. Both Soil Bacteria and Soil Chemical Property Affected the Micropredator Myxobacterial Community: Evidence from Natural Forest Soil and Greenhouse Rhizosphere Soil. Microorganisms 2020, 8, 1387. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8091387
Zhou Y, Zhang X, Yao Q, Zhu H. Both Soil Bacteria and Soil Chemical Property Affected the Micropredator Myxobacterial Community: Evidence from Natural Forest Soil and Greenhouse Rhizosphere Soil. Microorganisms. 2020; 8(9):1387. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8091387
Chicago/Turabian StyleZhou, Yang, Xianjiao Zhang, Qing Yao, and Honghui Zhu. 2020. "Both Soil Bacteria and Soil Chemical Property Affected the Micropredator Myxobacterial Community: Evidence from Natural Forest Soil and Greenhouse Rhizosphere Soil" Microorganisms 8, no. 9: 1387. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8091387