Ultrasonic Modulation of the Technological and Functional Properties of Yeast Strains

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Ultrasound Treatment (US)
2.3. Growth Assays
- GI < 25%: complete inhibition;
- 25% < GI < 75%: partial inhibition;
- GI > 75%: no inhibition.
- GI > 120%: growth enhancement.
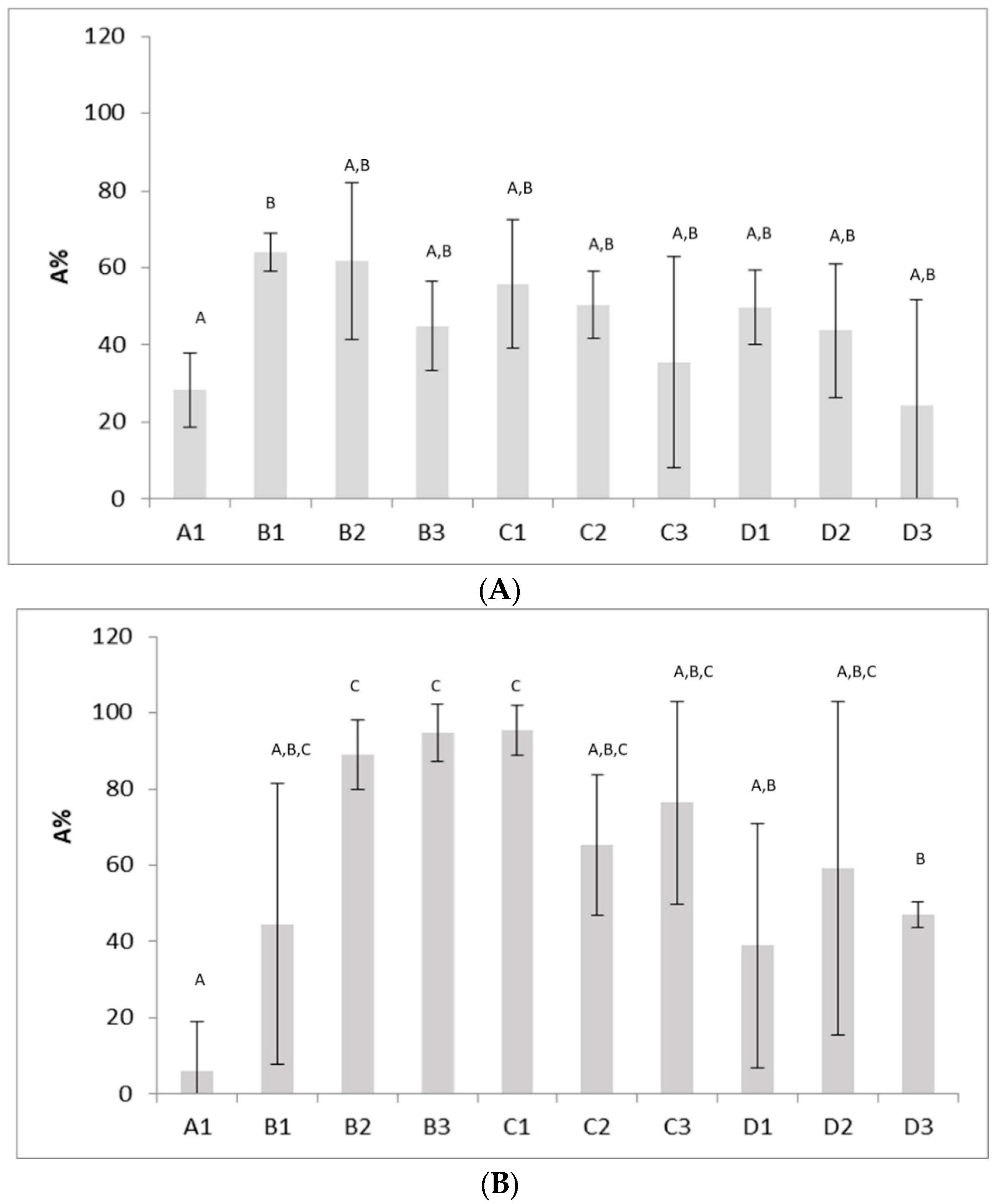
2.4. Autoaggregation Assays
2.5. Simulated Gastrointestinal Conditions
- SS, Salivary conditions, i.e., a sterile electrolyte solution (pH 6.5) containing 0.22 g/L CaCl2 (C. Erba, Milan, Italy), 6.5 g/L NaCl (C. Erba), 2.2 g/L KCl (J.T. Baker, Milan, Italy), 1.2 g/L NaHCO3 (Sigma-Aldrich), and 100 mg/L lysozyme (Sigma-Aldrich) [31].
- SGJ, Simulated Gastric Juice, i.e., a saline solution (0.9% NaCl) containing 3 g/L pepsin (porcine gastric mucosal, Sigma-Aldrich) and buffered to pH 2 [32].
- SIF, Simulated Intestinal Fluid, i.e., a fluid prepared by mixing 1 g/L of pancreatin (porcine pancreas, Sigma-Aldrich) and 3 g/L of bile extract (bile extract porcine, Sigma-Aldrich) in a solution (pH 8) containing 6.5 g/L NaCl, 0.835 g/L KCl, 0.22 g/L CaCl2, 1386 g/L NaHCO3) [32].
2.6. Statistical Analysis
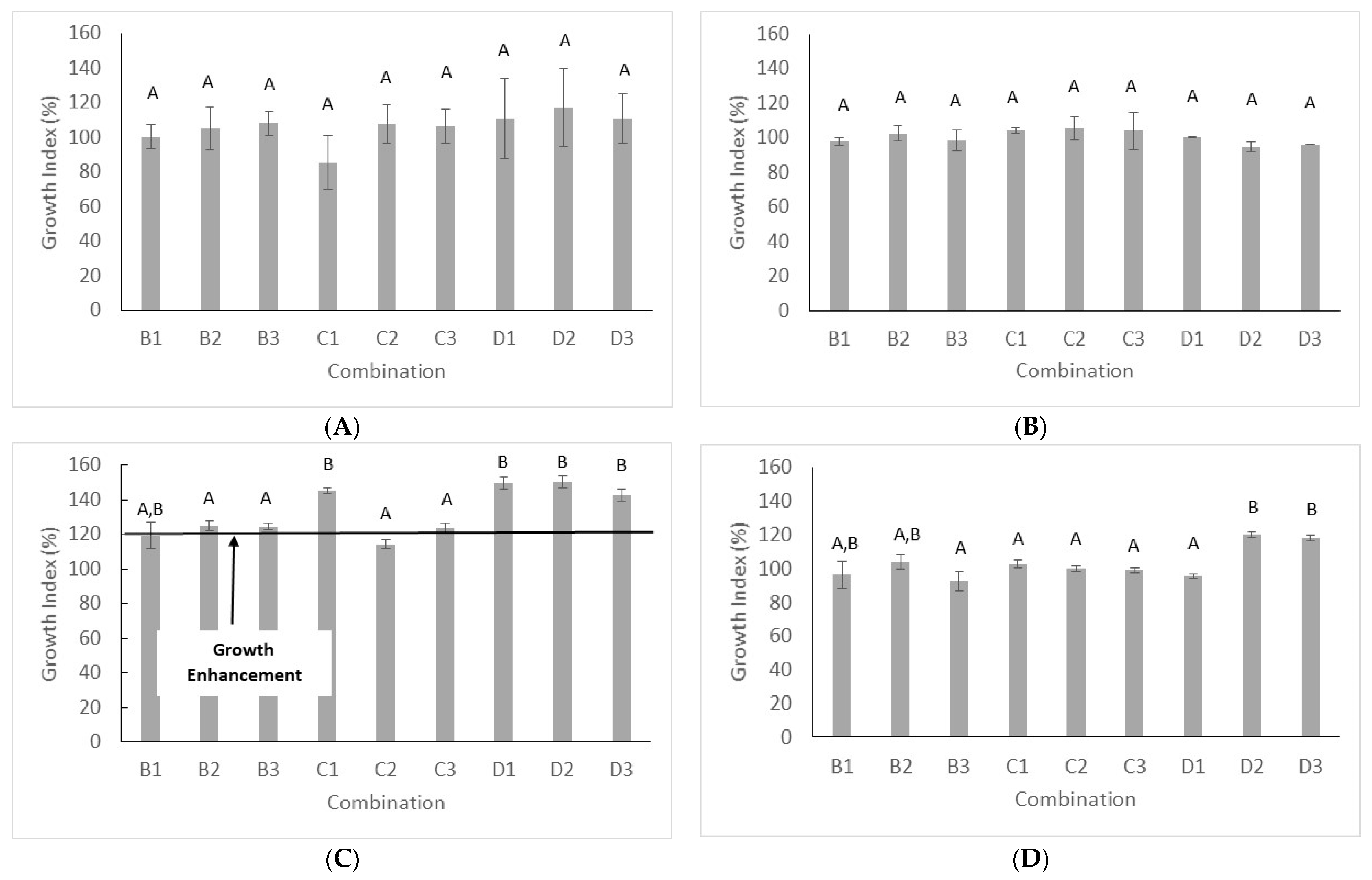
3. Results
- (a)
- Strain 17 in the combinations B2, B3 and C1 at 37 °C, with a GI ranging from 132.19 to 159.59%
- (b)
- Strain 41 at 37 °C in the combinations C1 (GI, 233%) and C2 (GI, 185.71%).
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Chisti, Y. Sonobioreactors: Using ultrasound for enhanced microbial productivity. Trends Biotechnol. 2003, 21, 89–93. [Google Scholar] [CrossRef]
- Zhao, Y.; Ang, W.T.; Xing, J.; Zhang, J.; Chen, J. Applications of ultrasound to enhance mycophenolic acid production. Ultrasound Med. Biol. 2012, 38, 1582–1588. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, A.; Campaniello, D.; Speranza, B.; Altieri, C.; Sinigaglia, M.; Corbo, M.R. Two Nonthermal Technologies for Food Safety and Quality—Ultrasound and High Pressure Homogenization: Effects on Microorganisms, Advances, and Possibilities: A Review. J. Food Prot. 2019, 82, 2049–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Li, L.; Zhou, L.; Li, B.; Xu, Z. Effect of ultrasound treatment conditions on Saccharomyces cerevisiae by response surface methodology. Microb. Pathog. 2017, 111, 497–502. [Google Scholar] [CrossRef]
- Gogate, P.R.; Kabadi, A.M. A review of applications of cavitation in biochemical engineering/biotechnology. Biochem. Eng. J. 2009, 44, 60–72. [Google Scholar] [CrossRef]
- Pagnossa, J.P.; Rocchetti, G.; Ribeiro, A.C.; Piccoli, R.H.; Lucini, L. Ultrasound: Beneficial biotechnological aspects on microorganisms-mediated processes. Curr. Opin. Food Sci. 2020, 31, 24–30. [Google Scholar] [CrossRef]
- Huang, G.; Chen, S.; Dai, C.; Sun, L.; Sun, W.; Tang, Y.; Xiong, F.; He, R.; Ma, H. Effects of ultrasound on microbial growth and enzyme activity. Ultrason. Sonochem. 2017, 37, 144–149. [Google Scholar] [CrossRef]
- Dai, C.; Xiong, F.; He, R.; Zhang, W.; Ma, H. Effects of low intensity ultrasound on the growth, cell membrane permeability and ethanol tolerance of Saccharomyces cerevisiae. Ultrason. Sonochem. 2017, 36, 191–197. [Google Scholar] [CrossRef]
- Awad, T.S.; Moharram, H.A.; Shaltout, O.E.; Asker, D.; Youssef, M.M. Applications of ultrasound in analysis, processing and quality control of food: A review. Food Res. Int. 2012, 48, 410–427. [Google Scholar] [CrossRef]
- Kwiatkowska, B.; Bennett, B.J.; Akunna, J.; Walker, G.M.; Bremner, D.H. Stimulation of bioprocesses by ultrasound. Biotechnol. Adv. 2011, 29, 768–780. [Google Scholar] [CrossRef] [Green Version]
- Ojha, K.S.; Mason, T.J.; O’Donnell, C.P.; Kerry, J.P.; Tiwari, B.K. Ultrasound technology for food fermentation applications. Ultrason. Sonochem. 2017, 34, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Huang, L.; Jia, J.; He, R.; Luo, L.; Zhu, W. Effect of energy-gathered ultrasound on Alcalase. Ultrason. Sonochem. 2011, 18, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Chao, L.; Li, H.; Zhao, L.; Xu, Y.; Long, Y.; Chen, Q.; Zhang, Z. Research progress of applications of ultrasonic technology in food industry. J. Food Saf. Qual. 2017, 8, 2670–2677. [Google Scholar]
- Polonorum, A.S.; Stadnik, J.; Stasiak, D. Applications of ultrasound in food technology. Acta Sci. Pol. Technol. Aliment. 2007, 6, 89–99. [Google Scholar]
- Vercet, A.; Oria, R.; Marquina, P.; Crelier, S.; Lopez-Buesa, P. Rheological properties of yoghurt made with milk submitted to manothermosonication. J. Agric. Food Chem. 2002, 50, 6165–6171. [Google Scholar] [CrossRef]
- Erriu, M.; Blus, C.; Szmukler-Moncler, S.; Buogo, S.; Levi, R.; Barbato, G.; Madonnaripa, D.; Denotti, G.; Piras, V.; Orrù, G. Microbial biofilm modulation by ultrasound: Current concepts and Controversies. Ultrason. Sonochem. 2014, 21, 15–22. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Racioppo, A.; Sinigaglia, M.; Speranza, B.; Campaniello, D.; Corbo, M.R. A low-power ultrasound attenuation improves the stability of biofilm and hydrophobicity of Propionibacterium freudenreichii subsp. freudenreichii DSM 20271 and Acidipropionibacterium jensenii DSM 20535. Food Microbiol. 2019, 78, 104–109. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Speranza, B.; Iorio, M.C.; Loi, M.; Sinigaglia, M.; Corbo, M.R. US-inactivation of foodborne bacteria: Screening in distilled water and combination with citrus extract in skim milk. LWT Food Sci. Technol. 2016, 70, 135–141. [Google Scholar] [CrossRef]
- Racioppo, A.; Corbo, M.R.; Piccoli, C.; Sinigaglia, M.; Speranza, B.; Bevilacqua, A. Ultrasound attenuation of lactobacilli and bifidobacteria: Effect on some technological and probiotic properties. Int. J. Food Microbiol. 2017, 243, 78–83. [Google Scholar] [CrossRef]
- Dahroud, B.D.; Mokarram, R.R.; Khiabani, M.S.; Hamishehkar, H.; Bialvaei, A.Z.; Yousefi, M.; Kafil, H.S. Low intensity ultrasound increases the fermentation efficiency of Lactobacillus casei subsp. casei ATTC 39392. Int. J. Biol. Macromol. 2016, 86, 462–467. [Google Scholar] [CrossRef]
- Nguyen, T.M.P.; Lee, Y.K.; Zhou, W. Stimulating fermentative activities of bifidobacteria in milk by high intensity ultrasound. Int. Dairy J. 2009, 19, 410–416. [Google Scholar] [CrossRef]
- Yeo, S.K.; Liong, M.T. Effect of ultrasound on the growth of probiotics and bioconversion of isoflavones in prebiotic-supplemented soymilk. J. Agric. Food Chem. 2011, 59, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Lanchun, S.; Bochu, W.; Zhiming, L.; Chuanren, D.; Dai, C.; Sakanishi, A. The research into the influence of low-intensity ultrasonic on the growth of S. cerevisiae. Colloids Surf. B Biointerfaces 2003, 30, 43–49. [Google Scholar] [CrossRef]
- Borah, A.J.; Agarwal, M.; Goyal, A.; Moholkar, V.S. Physical insights of ultrasound-assisted ethanol production from composite feedstock of invasive weeds. Ultrason. Sonochem. 2019, 51, 378–385. [Google Scholar] [CrossRef]
- Al Daccache, M.; Kouba, M.; Salameh, D.; Maroun, R.G.; Louka, N.; Vorobieva, E. Ultrasound-assisted fermentation for cider production from Lebanese apples. Ultrason. Sonochem. 2020, 63, 104952. [Google Scholar] [CrossRef]
- Perricone, M.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Technological characterization and probiotic traits of yeasts isolated from Altamura sourdough to select promising microorganisms as functional starter cultures for cereal-based products. Food Microbiol. 2014, 38, 26–35. [Google Scholar] [CrossRef]
- Petruzzi, L.; Bevilacqua, A.; Baiano, A.; Beneduce, L.; Corbo, M.R.; Sinigaglia, M. Study of Saccharomyces cerevisiae W13 as a functional starter for the removal of ochratoxin A. Food Control 2014, 35, 373–377. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Perricone, M.; Cannarsi, M.; Corbo, M.R.; Sinigaglia, M. Technological and spoiling characteristics of the yeast microflora isolated from Bella di Cerignola table olives. Int. J. Food Sci. Technol. 2009, 44, 2198–2207. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Arroyo-López, F.N.; Rantsiou, K.; Jiménez-Díaz, R.; Garrido- Fernández, A.; Cocolin, L. Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential. Food Res. Int. 2013, 50, 135–142. [Google Scholar] [CrossRef]
- Petruzzi, L.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Ochratoxin A Removal by Yeasts after Exposure to Simulated Human Gastrointestinal Conditions. J. Food Sci. 2016, 81, M2756–M2760. [Google Scholar] [CrossRef]
- Russo, P.; de Palencia, P.F.; Romano, A.; Fernandez, M.; Lucas, P.; Spano, G.; Lopez, P. Biogenic amine production by the wine Lactobacillus brevis IOEB 9809 in systems that partially mimic the gastrointestinal tract stress. BMC Microbiol. 2012, 12, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priya, A.J.; Vijayalakshmi, S.P.; Raichur, A.M. Enhanced survival of probiotic Lactobacillus acidophilus by encapsulation with nanostructured polyelectrolyte layers through layer-by-layer approach. J. Agric. Food Chem. 2011, 59, 11838–11845. [Google Scholar] [CrossRef] [PubMed]
- de Wouters, T.; Jans, C.; Niederberger, T.; Fischer, P.; Rühs, P.A. Adhesion potential of intestinal microbes predicted by physico-chemical characterization methods. PLoS ONE 2015, 10, e0136437. [Google Scholar] [CrossRef] [PubMed]
- Haddaji, N.; Mahdhi, A.K.; Krifi, B.; Ben Ismail, M.; Bakhrouf, A. Change in cell surface properties of Lactobacillus casei under heat shock treatment. FEMS Microbiol. Lett. 2015, 362, fnv047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Kotzamandis, C.; Kourelis, A.; Litopoilou-Tzanetaki, E.; Tzanetakis, N.; Yiangou, M. Evaluation of adhesion capacity, cell surface traits and immunomodulatory activity of presumptive probiotic Lactobacillus strains. Int. J. Food Microbiol. 2010, 140, 154–163. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Campaniello, D.; Speranza, B.; Racioppo, A.; Altieri, C.; Sinigaglia, M.; Corbo, M.R. Microencapsulation of Saccharomyces cerevisae into alginate beads: A focus of functional properties of released cells. Foods 2020, 9, 1051. [Google Scholar] [CrossRef]
- Ashokkumar, M. Applications of ultrasound in food and bioprocessing. Ultrason. Sonochem. 2015, 25, 17–23. [Google Scholar] [CrossRef]
- Majid, I.; Nayik, G.A.; Nanda, V. Ultrasonication and food technology: A review. Cogent Food Agric. 2015, 1, 1071022. [Google Scholar] [CrossRef]
- Jomdecha, C.; Prateepasen, A. Effects of pulse ultrasonic irradiation on the lag phase of Saccharomyces cerevisiae growth. Lett. Appl. Microbiol. 2011, 52, 62–69. [Google Scholar] [CrossRef]
- Singh, S.; Agarwal, M.; Sarma, S.; Goyal, A.; Moholkar, V.S. Mechanistic insight into ultrasound induced enhancement of simultaneous saccharification and fermentation of Parthenium hysterophorus for ethanol production. Ultrason. Sonochem. 2015, 26, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Huezo, L.; Shah, A.; Michel, F.C., Jr. Effects of ultrasound on fermentation of glucose to ethanol by Saccharomyces cerevisiae. Fermentation 2019, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Chuanyun, W.; Bochu, D.; Chuanren, A. Low ultrasonic stimulates fermentation of riboflavin producing strain Ecemothecium ashbyii. Colloids Surf. B Biointerfaces 2003, 30, 37–41. [Google Scholar] [CrossRef]
- Karshafian, R.; Samac, S.; Bevan, P.D.; Burns, P.N. Microbubble mediated sonoporation of cells in suspension: Clonogenic viability and influence of molecular size on uptake. Ultrasonics 2010, 50, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Herrán, N.S.; Casas, J.L.; López, J.A.; Sánchez Pérez, M.M.; Ballesteros Martín, B.; Chisti, Y. Ultrasound affects fungal morphology and broth rheology of Aspergillus terreus. J. Biotechnol. 2008, 136, S489–S490. [Google Scholar] [CrossRef]
- Mota, M.J.; Lopes, R.P.; Koubaa, M.; Shahin Roohinejad, S.; Barba, F.J.; Delgadillo, I.; Saraiva, J.A. Fermentation at non-conventional conditions in food- and bio-sciences by the application of advanced processing technologies. Crit. Rev. Biotechnol. 2017, 38, 122–140. [Google Scholar] [CrossRef]
- Wu, T.; Yu, X.; Hu, A.; Zhang, L.; Jin, Y.; Abid, M. Ultrasonic disruption of yeast cells: Underlying mechanism and effects of processing parameters. Innov. Food Sci. Emerg. Technol. 2015, 28, 59–65. [Google Scholar]
- Sango, D.M.; Abela, D.; McElhatton, A.; Valdramidis, V.P. Assisted ultrasound applications for the production of safe foods. J. Appl. Microbiol. 2014, 116, 1067–1083. [Google Scholar] [CrossRef]
- Ye, S.Y.; Qiu, Y.X.; Song, X.L.; Luo, S.C. Optimization of process parameters for the inactivation of Lactobacillus sporogenes in tomato paste with ultrasound 60 Cogamma irradiation using response surface methodology. Radiat. Phys. Chem. 2009, 78, 227–233. [Google Scholar] [CrossRef]
- Niemczewski, B. Maximisation of cavitation intensity in ultrasonic cleaning in aqueous solutions through selection of salt solution. Trans. Inst. Met. Finish. 2011, 89, 104–108. [Google Scholar] [CrossRef]
- García-Cayuela, T.; Korany, A.M.; Bustos, I.; Gómez de Cadiñanos, L.P.; Requena, T.; Peláez, P.; Martínez-Cuesta, M.C. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- Brückner, S.; Mösch, H.U. Choosing the right lifestyle: Adhesion and development in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2012, 36, 25–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabatabaie, F.; Mortazavi, A. Studying the effects of ultrasound shock on cell wall permeability and survival of some LAB in milk. World Appl. Sci. J. 2008, 3, 119–121. [Google Scholar]
- Touhami, A.; Hoffmann, B.; Vasella, A.; Denis, F.A.; Dufrene, Y.F. Aggregation of yeast cells: Direct measurement of discrete lectin-carbohydrate interactions. Microbiology 2003, 149, 2873–2878. [Google Scholar] [CrossRef] [Green Version]
- Powell, C.D.; Quain, D.E.; Smart, K.A. The impact of brewing yeast cell age on fermentation performance, attenuation and flocculation. FEMS Yeast Res. 2003, 3, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Zamora-Vega, R.; Montanez-Soto, J.L.; Martınez-Flores, H.E.; Flores-Magallon, R.; Munoz-Ruiz, C.V.; Venegas-Gonzalez, J.; Ariza, O.T. Effect of incorporating prebiotics in coating materials for the microencapsulation of Saccharomyces boulardii. Int. J. Food Sci. Nutr. 2012, 63, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Vega, R.; Martınez-Flores, E.; Montanez-Soto, J.L.; Rodiles-Lopez, J.O. Viabilidad de Saccharomyces boulardii en queso fresco bajo condiciones de acidez. Rev. Electron. Nova Sci. 2015, 15, 68–80. [Google Scholar]
- Rodriguez, E.T.; Flores, H.E.M.; Lopez, J.O.R.; Vega, R.Z.; Garciglia, R.S.; Sanchez, R.E.P. Survival rate of Saccharomyces boulardii adapted to a functional freeze-dried yogurt: Experimental study related to processing, storage and digestion by Wistar rats. Funct. Foods Health Dis. 2017, 7, 98–114. [Google Scholar]



{kind=link}
{kind=link}
| Power (%) | Time (min) | Pulse (s) | |
|---|---|---|---|
| A1 | 0 | 0 | 0 |
| B1 | 20 | 2 | 2 |
| B2 | 20 | 2 | 6 |
| B3 | 20 | 2 | 10 |
| C1 | 20 | 6 | 2 |
| C2 | 20 | 6 | 6 |
| C3 | 20 | 6 | 10 |
| D1 | 20 | 10 | 2 |
| D2 | 20 | 10 | 6 |
| D3 | 20 | 10 | 10 |
| Strains | Combinations | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | B1 | B2 | B3 | C1 | C2 | C3 | D1 | D2 | D3 | ||
| 2 | 5.55 ± 0.05 | 5.64 ± 0.02 | 5.67 ± 0.08 | 5.70 ± 0 | 5.68 ± 0 | 5.77 ± 0.10 | 5.76 ± 0.06 | 5.69 ± 0.17 | 5.65 ± 0.11 | 5.71 ± 0.01 | ns |
| 4 | 5 ± 0.30 | 4.98 ± 0.32 | 5 ± 0.35 | 4.98 ± 0.31 | 4.81 ± 0.30 | 4.95 ± 0.30 | 5.22 ± 0.34 | 5.19 ± 0.30 | 5 ± 0.32 | 5.16 ± 0.29 | ns |
| 17 | 5.68 ± 0.26 | 5.90 ± 0.20 | 5.79 ± 0 | 5.78 ± 0.06 | 5.82 ± 0.10 | 5.89 ± 0.27 | 5.76 ± 0.21 | 5.83 ± 0.07 | 5.78 ± 0.25 | 6.07 ± 0.36 | ns |
| 41 | 5.64 ± 0.15 | 6.02 ± 0.12 | 5.72 ± 0.38 | 5.48 ± 0.33 | 5.66 ± 0.30 | 5.55 ± 0.33 | 5.57 ± 0.35 | 5.63 ± 0.33 | 5.65 ± 0.34 | 5.73 ± 0.28 | ns |
| 4y | 5.54 ± 0.14 | 5.51 ± 0.30 | 5.42 ± 0.35 | 5.52 ± 0.32 | 5.49 ± 0.32 | 5.48 ± 0.31 | 5.39 ± 0.34 | 5.40 ± 0.31 | 5.44 ± 0.30 | 5.49 ± 0.29 | ns |
| WB | 5.73 ± 0.31 | 5.67 ± 0.33 | 5.68 ± 0.35 | 4.81 ± 0.33 | 4.81 ± 0.33 | 5.52 ± 0.30 | 5.53 ± 0.34 | 5.41 ± 0.35 | 5.43 ± 0.33 | 5.66 ± 0.35 | ns |
| W21 | 5.49 ± 0.21 | 5.49 ± 0.21 | 5.53 ± 0.28 | 5.28 ± 0.30 | 5.48 ± 0.29 | 5.64 ± 0.30 | 6.19 ± 0.31 | 5.58 ± 0.34 | 5.46 ± 0.35 | 5.56 ± 0.27 | ns |
| WL43 | 5.70 ± 0.21 | 5.48 ± 0.20 | 5.81 ± 0.25 | 5.59 ± 0.23 | 5.68 ± 0.27 | 5.50 ± 0.22 | 5.18 ± 0.33 | 5.58 ± 0.27 | 5.45 ± 0.35 | 5.56 ± 0.20 | ns |
| Combinations | B1 | B2 | B3 | C1 | C2 | C3 | D1 | D2 | D3 |
|---|---|---|---|---|---|---|---|---|---|
| Strain 2 | |||||||||
| 25 °C | 70.85 ± 13.82A | 65.26 ± 7.85A | 67.66 ± 26.79A | 64.21 ± 17.45A | 72.16 ± 23.98A | 84.91 ± 11.19A | 66.73 ± 34.87A | 69.31 ± 16.34A | 82.11 ± 30.71A |
| 37 °C | 87.30 ± 10.41A | 98.98 ± 1.27A | 101.31 ± 11.13A | 104.80 ± 10.06A | 94.18 ± 14.72A | 115.85 ± 7.59A | 108.61 ± 18.88A | 129.33 ± 13.97A,B | 90.30 ± 2.67A |
| 2% NaCl | 129.17 ± 0C | 129.76 ± 0C | 84.52 ± 0A | 106.55 ± 0B | 106.55 ± 0B | 80.65 ± 9.68A | 93.75 ± 8.84A,B | 109.82 ± 14.73A,B,C | 85.42 ± 2.10A |
| Strain 4 | |||||||||
| 25 °C | 89.64 ± 3.50A | 98.54 ± 8.50A | 94.31 ± 9.42A | 105.73 ± 2.01A | 101.95 ± 0.34A | 105.48 ± 5.92A | 103.66 ± 6.43A | 106.86 ± 1.44A | 108.16 ± 1.09A |
| 37 °C | 100.93 ± 7.45A | 104.65 ± 3.16A | 103.48 ± 1.50A | 97.20 ± 1.81A | 100.57 ± 10.89A | 99.34 ± 7.36A | 101.51 ± 0.57A | 97.28 ± 2.10A | 98.61 ± 2.36A |
| 2% NaCl | 106.29 ± 11.31A | 117.64 ± 12.51A | 124.72 ± 13.27A | 108.52 ± 11.54A | 120.06 ± 12.77A | 121.13 ± 12.88A | 117.54 ± 12.50A | 132.38 ± 14.08A | 131.70 ± 14.01A |
| Strain 17 | |||||||||
| 25 °C | 117.95 ± 24.52A | 113.54 ± 33.88A | 131.62 ± 20.05A | 123.28 ± 17.44A | 130.19 ± 14.21A | 119.73 ± 31.37A | 110.59 ± 44.45A | 104.93 ± 25.55A | 104.30 ± 13.38A |
| 37 °C | 103.81 ± 20.15 A,B | 159.59 ± 20.49B | 132.19 ± 9.47B | 133.44 ± 1.75B | 104.96 ± 9.63 A | 97.85 ± 11.63 A | 98.71 ± 11.71 A | 97.90 ± 16.48 A | 90.03 ± 11.61 A |
| 2% NaCl | 111.46 ± 16.20A | 137.59 ± 21.00A | 125.93 ± 28.08A | 136.73 ± 25.65A | 134.27 ± 16.30A | 113.90 ± 21.86A | 133.17 ± 3.46A | 106.61 ± 24.29A | 146.80 ± 12.41A |
| Strain 41 | |||||||||
| 25 °C | 91.48 ± 45.95A | 140.80 ± 21.78A | 138.89 ± 26.72A | 132.81 ± 30.71A | 110.69 ± 15.75A | 88.45 ± 201.64A | 111.94 ± 87.35A | 87.82 ± 0.48A | 149.82 ± 48.96A |
| 37 °C | 133.68 ± 12.76A | 127.11 ± 8.15A | 132.85 ± 1.97A | 233.01 ± 11.63C | 185.71 ± 25.08B | 112.99 ± 35.86A | 100.64 ± 33.34A | 137.91 ± 5.18A | 96.63 ± 35.94A |
| 2% NaCl | 92.04 ± 3.12C | 73.48 ± 9.64A,B | 72.37 ± 13.43A,B | 83.67 ± 12.23A,B | 67.58 ± 2.99A | 75.51 ± 3.93B | 90.73 ± 2.55C | 87.55 ± 13.31A,B | 102.76 ± 0.24D |
| Strain 4y | |||||||||
| 25 °C | 111.42 ± 31.77A | 116.27 ± 57.24A | 149.59 ± 17.92A | 108.11 ± 33.34A | 93.04 ± 38.29A | 91.97 ± 37.85A | 79.81 ± 32.84A | 142.75 ± 58.75A | 171.35 ± 70.52A |
| 37 °C | 94.15 ± 26.00A | 102.40 ± 7.63A | 94.84 ± 10.43A | 98.99 ± 3.55A | 96.69 ± 3.47A | 95.37 ± 3.42A | 105.73 ± 3.79A | 100.17 ± 3.59A | 97.11 ± 3.48A |
| 2% NaCl | 114.68 ± 18.47A | 110.90 ± 23.38A | 101.60 ± 4.89A | 118.40 ± 17.69A | 120.78 ± 18.05A | 113.91 ± 17.02A | 114.29 ± 17.08A | 117.61 ± 17.51A | 138.62 ± 20.71A |
| Strain WB | |||||||||
| 25 °C | 144.23 ± 66.79A | 134.73 ± 60.40A | 101.09 ± 28.64A | 155.38 ± 45.86A | 167.74 ± 49.50A | 123.34 ± 36.40A | 201.03 ± 59.33A | 215.41 ± 63.57A | 275.44 ± 81.29A |
| 37 °C | 90.46 ± 3.88A | 99.75 ± 2.72A | 98.31 ± 5.18A | 100.31 ± 3.59A | 96.57 ± 3.46A | 90.35 ± 3.23A | 90.90 ± 3.25A | 92.89 ± 3.33A | 89.99 ± 3.22A |
| 2% NaCl | 112.17 ± 1.45A | 105.95 ± 18.73A | 112.64 ± 11.17A | 121.05 ± 13.48A | 87.42 ± 9.73A | 122.17 ± 13.60A | 115.49 ± 12.86A | 128.00 ± 14.25A | 120.12 ± 14.38A |
| Strain W21 | |||||||||
| 25 °C | 81.72 ± 13.80A | 102.02 ± 32.70A | 59.60 ± 51.48A | 109.05 ± 33.59A | 93.83 ± 33.46A | 105.93 ± 23.20A | 106.78 ± 33.29A | 93.70 ± 33.33A | 102.57 ± 23.28A |
| 37 °C | 91.80 ± 13.68A | 82.02 ± 22.62A | 93.54 ± 25.11A | 60.34 ± 43.19A | 89.78 ± 23.36A | 86.23 ± 33.23A | 89.38 ± 43.20A | 86.13 ± 23.30A | 83.48 ± 43.12A |
| 2% NaCl | 97.30 ± 28.70A | 97.82 ± 25.20A | 101.58 ± 19.90A | 102.04 ± 23.14A | 98.95 ± 18.22A | 97.56 ± 35.00A | 95.45 ± 31.83A | 100 ± 38.70A | 98.95 ± 40.13A |
| Strain WL43 | |||||||||
| 25 °C | 101.38 ± 23.50A | 87.26 ± 12.90A | 86.30 ± 41.88A | 88.46 ± 30.50A | 95.07 ± 30.40A | 84.44 ± 33.25A | 92.25 ± 38.89A | 93.15 ± 33.35A | 92.97 ± 37.28A |
| 37 °C | 113.28 ± 18.60A | 93.33 ± 27.42A | 83.59 ± 35.19A | 86.30 ± 40.89A | 71.61 ± 28.33A | 86.25 ± 31.20A | 83.18 ± 23.20A | 75.74 ± 13.90A | 83.85 ± 40.10A |
| 2% NaCl | 106.75 ± 25.50A | 110.97 ± 28.80A | 102.27 ± 29.77A | 107.72 ± 33.15A | 108.96 ± 18.20A | 113.37 ± 30A | 99.68 ± 30.87A | 105.52 ± 28.50A | 104.35 ± 44.33A |
| Yeast | Treatment | Inoculum | Salivary Conditions | Gastric Conditions | Intestinal Conditions | Sequential Transit |
|---|---|---|---|---|---|---|
| S. cerevisiae 2 | A1 | 6.40 ± 0.30 | 6.54 ± 0.29 | 6.78 ± 0.10 | 6.78 ± 0.08 | 5.81 ± 0.50 |
| D3 | 6.78 ± 0.14 | 6.64 ± 0.30 | 6.34 ± 0.32 | 6.48 ± 0.33 | 6.32 ± 0.18 | |
| ns | ns | ns | ns | ns | ||
| S. cerevisiae 17 | A1 | 7.02 ± 0.35 | 7.16 ± 0.36 | 6.51 ± 0.33 | 6.50 ± 0.30 | 6.37 ± 0.25 |
| D3 | 6.54 ± 0.33 | 6.54 ± 0.35 | 6.45 ± 0.31 | 6.46 ± 0.36 | 6.23 ± 0.40 | |
| ns | ns | ns | ns | ns |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Speranza, B.; Campaniello, D.; Altieri, C.; Sinigaglia, M.; Bevilacqua, A.; Corbo, M.R. Ultrasonic Modulation of the Technological and Functional Properties of Yeast Strains. Microorganisms 2020, 8, 1399. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8091399
Speranza B, Campaniello D, Altieri C, Sinigaglia M, Bevilacqua A, Corbo MR. Ultrasonic Modulation of the Technological and Functional Properties of Yeast Strains. Microorganisms. 2020; 8(9):1399. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8091399
Chicago/Turabian StyleSperanza, Barbara, Daniela Campaniello, Clelia Altieri, Milena Sinigaglia, Antonio Bevilacqua, and Maria Rosaria Corbo. 2020. "Ultrasonic Modulation of the Technological and Functional Properties of Yeast Strains" Microorganisms 8, no. 9: 1399. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8091399