Potential of Bacteria from Alternative Fermented Foods as Starter Cultures for the Production of Wheat Sourdoughs

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Flour
2.2. Starter Culture Strains, Inoculum Build-up, and Enumeration
2.3. Sourdough Production
2.4. Sample Processing
2.4.1. Determination of pH and Total Titratable Acidity
2.4.2. Microbiological Analysis of Sourdough Fermentation Samples
2.4.3. Metabolite Target Analysis to Map Substrate Consumption and Metabolite Production
2.5. Bread Production and Evaluation
3. Results
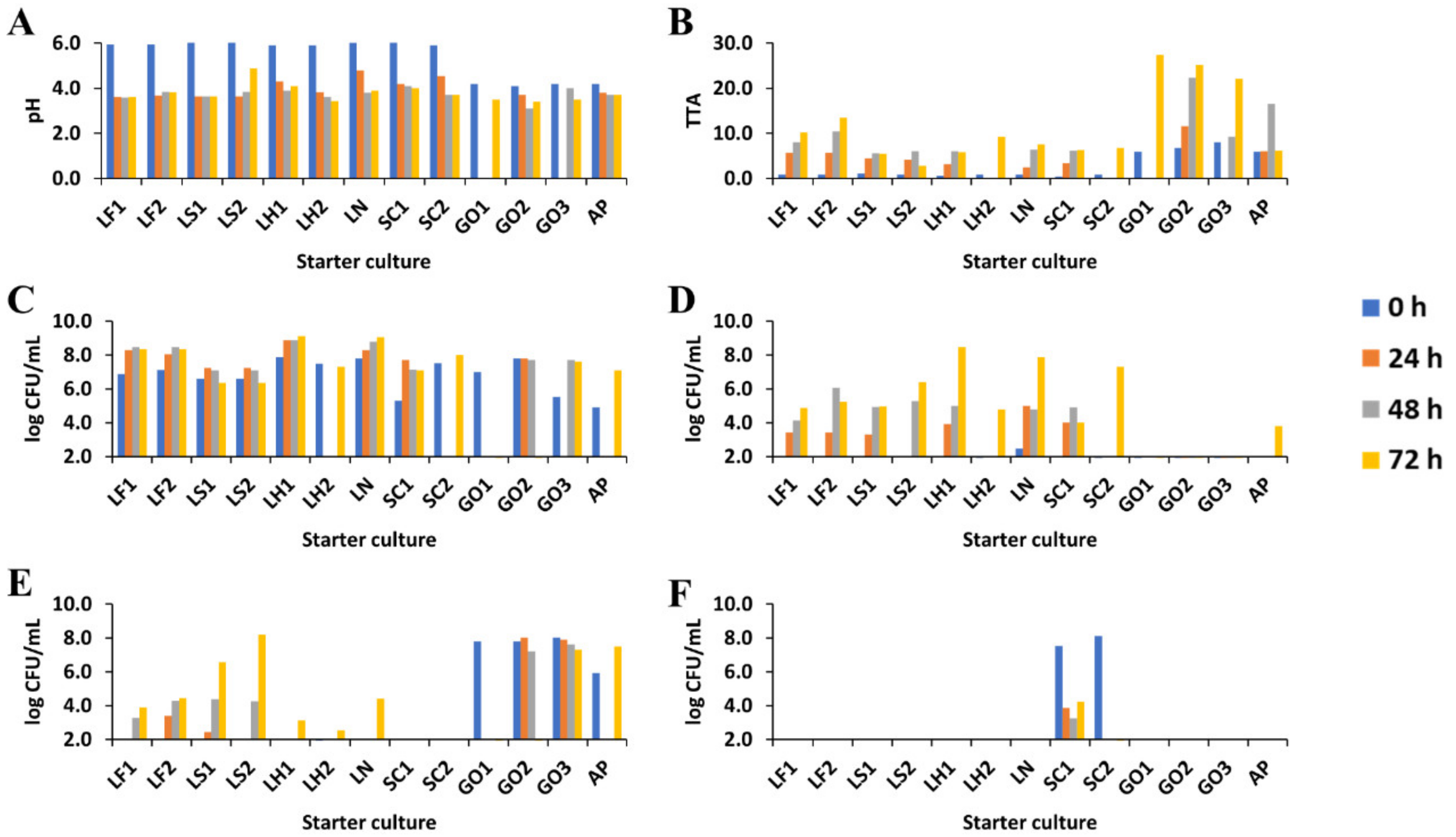
3.1. Sourdough pH and Total Titratable Acidity
3.2. Microbial Community Dynamics
3.2.1. Microbial Enumerations
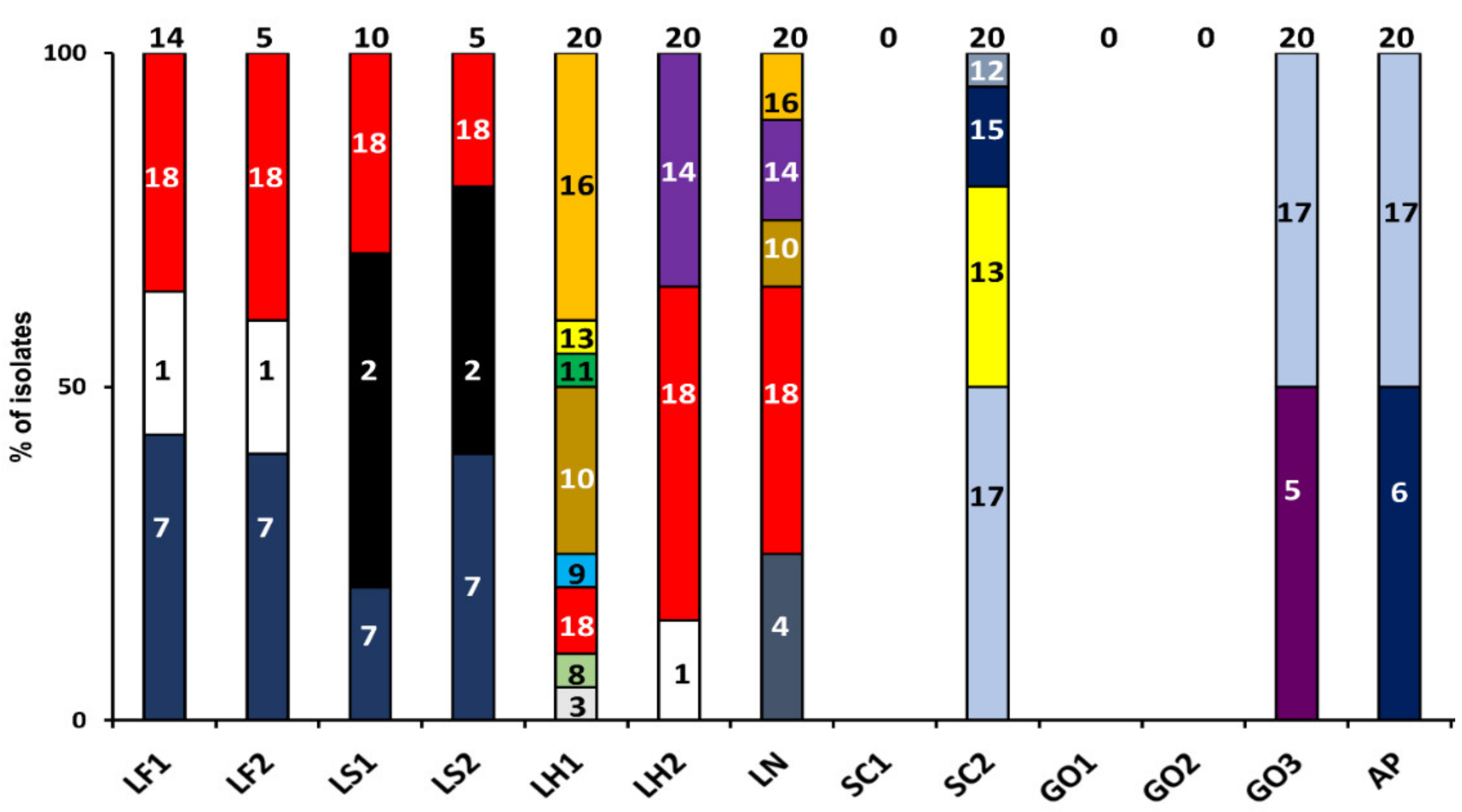
3.2.2. Microbial Identifications
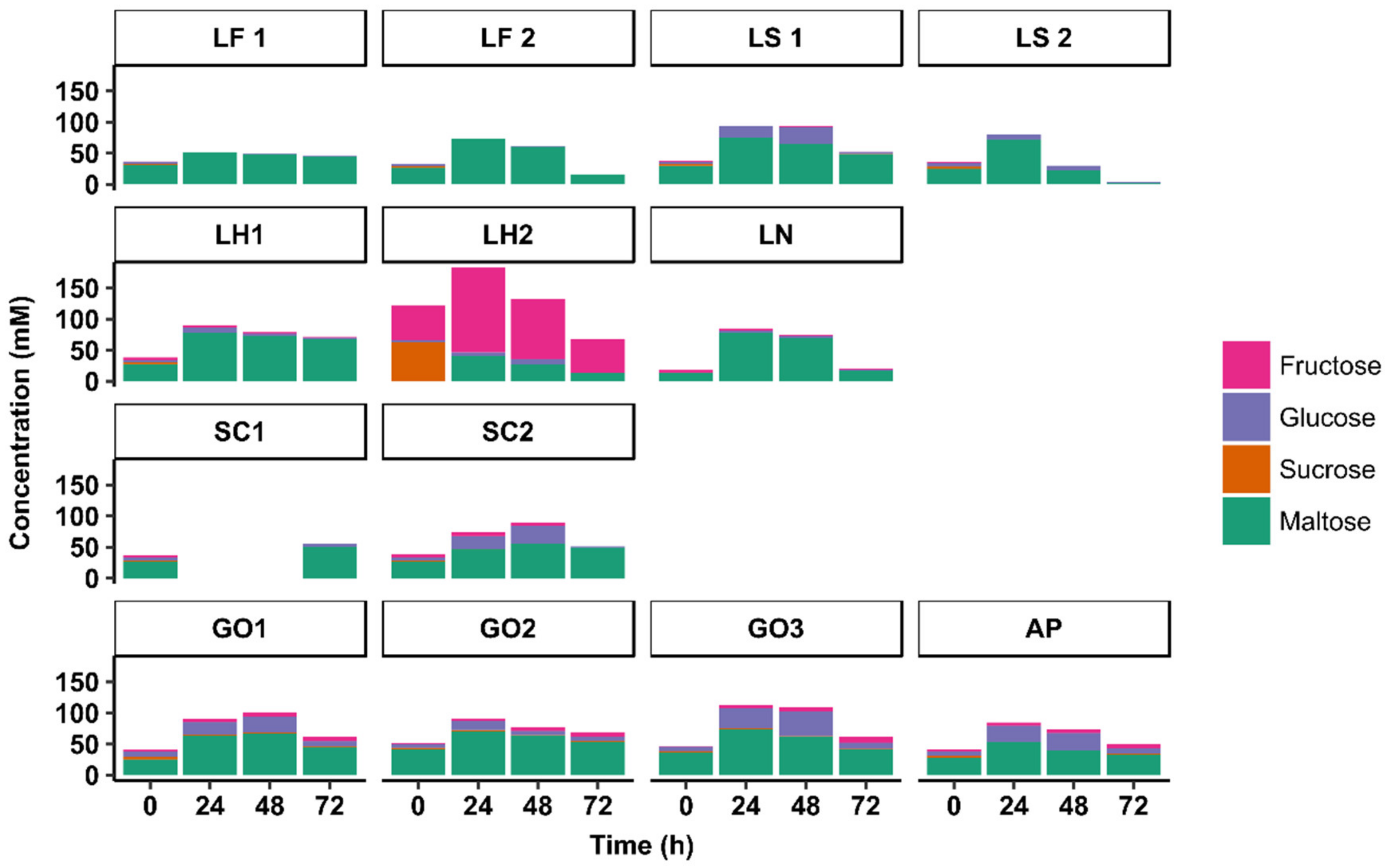
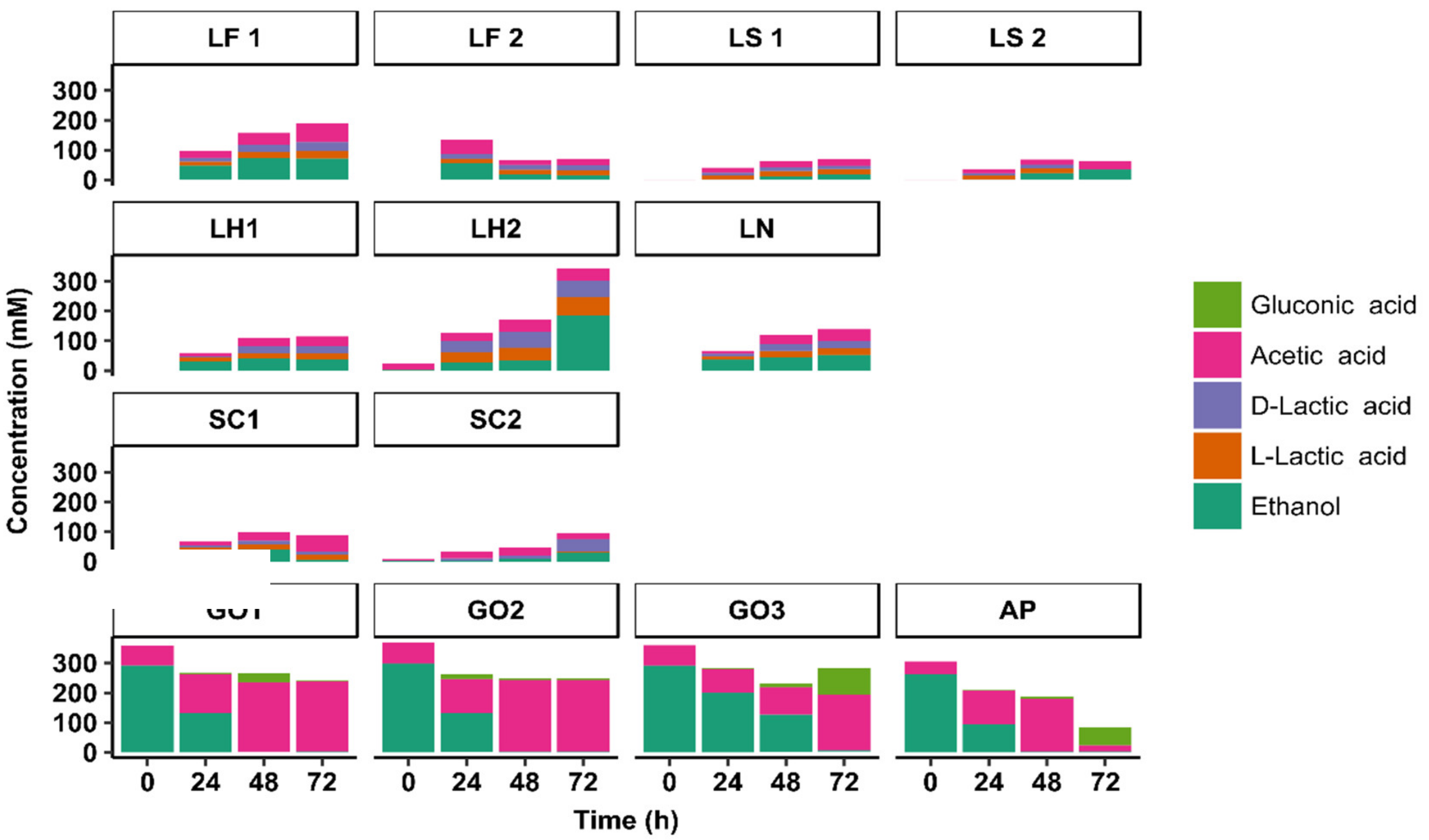
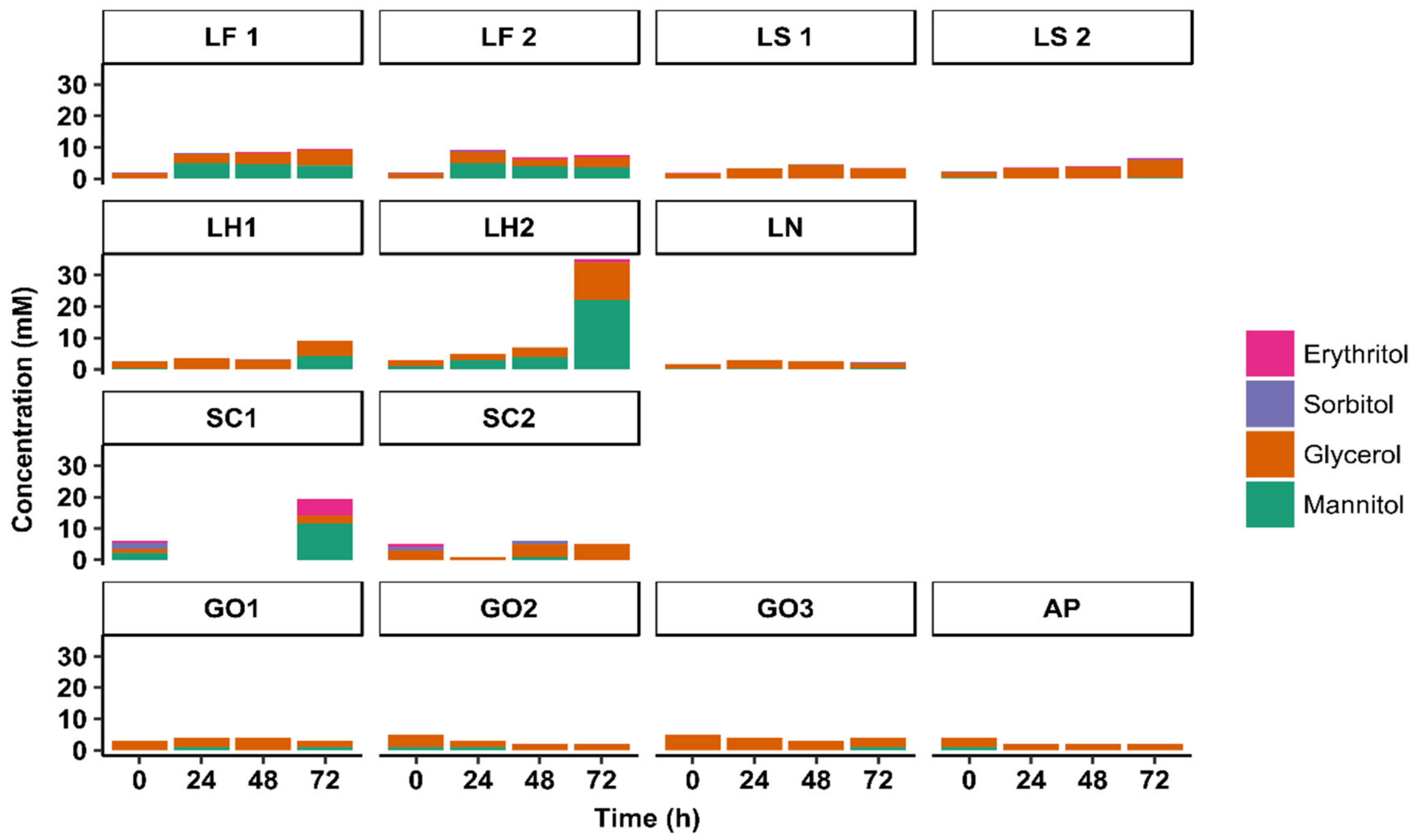
3.3. Substrate Consumption and Metabolite Production Kinetics
3.3.1. Substrate Consumption Kinetics
3.3.2. Metabolite Production Kinetics
3.4. Bread Production
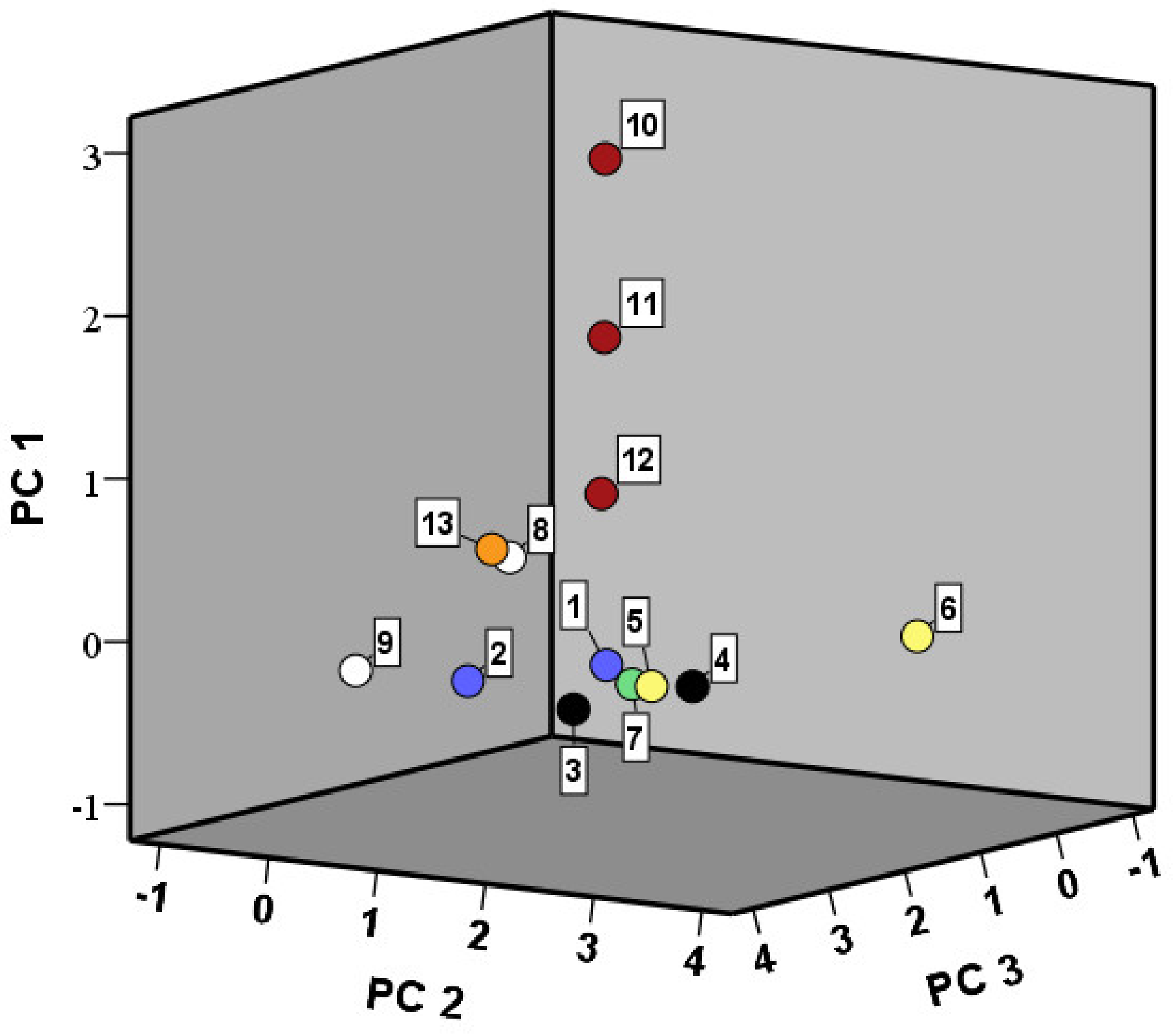
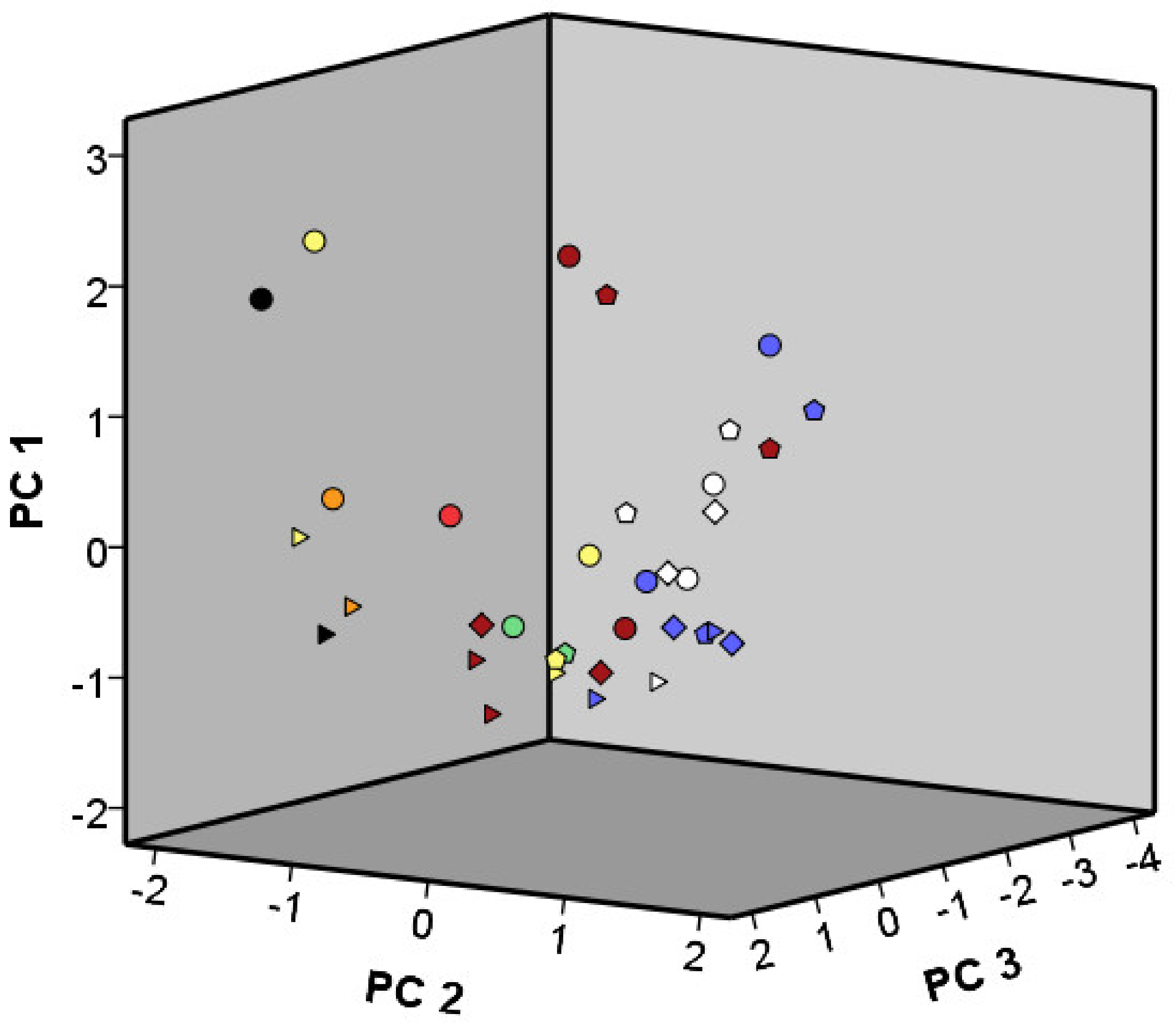
3.4.1. Volatile Organic Compounds
3.4.2. Color
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- De Vuyst, L.; Harth, H.; van Kerrebroeck, S.; Leroy, F. Yeast diversity of sourdoughs and associated metabolic properties and functionalities. Int. J. Food Microbiol. 2016, 239, 26–34. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; van Kerrebroeck, S.; Harth, H.; Huys, G.; Daniel, H.M.; Weckx, S. Microbial ecology of sourdough fermentations: Diverse or uniform? Food Microbiol. 2014, 37, 11–29. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; van Kerrebroeck, S.; Leroy, F. Microbial ecology and process technology of sourdough fermentation. Adv. Appl. Microbiol. 2017, 100, 49–160. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Vrancken, G.; Ravyts, F.; Rimaux, T.; Weckx, S. Biodiversity, ecological determinants, and metabolic exploitation of sourdough microbiota. Food Microbiol. 2009, 26, 666–675. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Ripari, V. Composition and function of sourdough microbiota: From ecological theory to bread quality. Int. J. Food Microbiol. 2016, 239, 19–25. [Google Scholar] [CrossRef]
- Gobbetti, M.; de Angelis, M.; Di Cagno, R.; Calasso, M.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2019, 302, 103–113. [Google Scholar] [CrossRef]
- Gobbetti, M.; Minervini, F.; Pontonio, E.; Di Cagno, R.; de Angelis, M. Drivers for the establishment and composition of the sourdough lactic acid bacteria biota. Int. J. Food Microbiol. 2016, 239, 3–18. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; de Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- Minervini, F.; de Angelis, M.; Di Cagno, R.; Gobbetti, M. Ecological parameters influencing microbial diversity and stability of traditional sourdough. Int. J. Food Microbiol. 2014, 171, 136–146. [Google Scholar] [CrossRef]
- Settanni, L. Sourdough and cereal-based foods: Traditional and innovative products. In Starter Cultures in Food Production, 1st ed.; Speranza, B., Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; John Wiley and Sons: Chichester, UK, 2017; pp. 199–230. [Google Scholar]
- Van Kerrebroeck, S.; Maes, D.; de Vuyst, L. Sourdoughs as a function of their species diversity and process conditions, a meta-analysis. Trends Food Sci. Technol. 2017, 68, 152–159. [Google Scholar] [CrossRef]
- Minervini, F.; Lattanzi, A.; de Angelis, M.; Celano, G.; Gobbetti, M. House microbiotas as sources of lactic acid bacteria and yeasts in traditional Italian sourdoughs. Food Microbiol. 2015, 52, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Ripari, V.; Gänzle, M.G.; Berardi, E. Evolution of sourdough microbiota in sponthaneous sourdoughs started with different plant materials. Int. J. Food Microbiol. 2016, 232, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Scheirlinck, I.; van der Meulen, R.; de Vuyst, L.; Vandamme, P.; Huys, G. Molecular source tracking of predominant lactic acid bacteria in traditional Belgian sourdoughs and their production environments. J. Appl. Microbiol. 2009, 106, 1081–1092. [Google Scholar] [CrossRef]
- Harth, H.; van Kerrebroeck, S.; de Vuyst, L. Community dynamics and metabolite target analysis of spontaneous, backslopped barley sourdough fermentations under laboratory and bakery conditions. Int. J. Food Microbiol. 2016, 228, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Van der Meulen, R.; Scheirlinck, I.; van Schoor, A.; Huys, G.; Vancanneyt, M.; Vandamme, P.; de Vuyst, L. Population dynamics and metabolite target analysis of lactic acid bacteria during laboratory fermentations of wheat and spelt sourdoughs. Appl. Environ. Microbiol. 2007, 73, 4741–4750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kerrebroeck, S.; Bastos, F.C.; Harth, H.; de Vuyst, L. A low pH does not determine the community dynamics of spontaneously developed backslopped liquid wheat sourdoughs but does influence their metabolite kinetics. Int. J. Food Microbiol. 2016, 239, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Weckx, S.; van der Meulen, R.; Allemeersch, J.; Huys, G.; Vandamme, P.; van Hummelen, P.; de Vuyst, L. Community dynamics of bacteria in sourdough fermentations as revealed by their metatranscriptome. Appl. Environ. Microbiol. 2010, 76, 5402–5408. [Google Scholar] [CrossRef] [Green Version]
- Weckx, S.; van der Meulen, R.; Maes, D.; Scheirlinck, I.; Huys, G.; Vandamme, P.; de Vuyst, L. Lactic acid bacteria community dynamics and metabolite production of rye sourdough fermentations share characteristics of wheat and spelt sourdough fermentations. Food Microbiol. 2010, 27, 1000–1008. [Google Scholar] [CrossRef]
- Corsetti, A. Technology of sourdough fermentation and sourdough applications. In Handbook on Sourdough Biotechnology, 1st ed.; Gobbetti, M., Gänzle, M.G., Eds.; Springer: Boston, MA, USA, 2013; pp. 85–103. [Google Scholar]
- Brandt, M.J. Starter cultures for cereal based foods. Food Microbiol. 2014, 37, 41–43. [Google Scholar] [CrossRef]
- Gaggiano, M.; Di Cagno, R.; de Angelis, M.; Arnault, P.; Tossut, P.; Fox, P.F.; Gobbetti, M. Defined multi-species semi-liquid ready-to-use sourdough starter. Food Microbiol. 2007, 24, 15–24. [Google Scholar] [CrossRef]
- Decock, P.; Cappelle, S. Bread technology and sourdough technology. Trends Food Sci. Technol. 2005, 16, 113–120. [Google Scholar] [CrossRef]
- Leroy, F.; de Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Comasio, A.; Harth, H.; Weckx, S.; de Vuyst, L. The addition of citrate stimulates the production of acetoin and diacetyl by a citrate-positive Lactobacillus crustorum strain during wheat sourdough fermentation. Int. J. Food Microbiol. 2019, 289, 88–105. [Google Scholar] [CrossRef] [PubMed]
- Perricone, M.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Technological characterization and probiotic traits of yeasts isolated from Altamura sourdough to select promising microorganisms as functional starter cultures for cereal-based products. Food Microbiol. 2014, 38, 26–35. [Google Scholar] [CrossRef]
- Pontonio, E.; Nionelli, L.; Curiel, J.A.; Sadeghi, A.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G. Iranian wheat flours from rural and industrial mills: Exploitation of the chemical and technology features, and selection of autochthonous sourdough starters for making breads. Food Microbiol. 2015, 47, 99–110. [Google Scholar] [CrossRef]
- Ravyts, F.; de Vuyst, L. Prevalence and impact of single-strain starter cultures of lactic acid bacteria on metabolite formation in sourdough. Food Microbiol. 2011, 28, 1129–1139. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Lorusso, A.; Montemurro, M.; Gobbetti, M. Use of sourdough made with quinoa (Chenopodium quinoa) flour and autochthonous selected lactic acid bacteria for enhancing the nutritional, textural and sensory features of white bread. Food Microbiol. 2016, 56, 1–13. [Google Scholar] [CrossRef]
- Ruiz Rodríguez, L.; Vera Pingitore, E.; Rollán, G.; Cocconcelli, P.S.; Fontana, C.; Saavedra, L.; Vignolo, G.; Hebert, E.M. Biodiversity and technological-functional potential of lactic acid bacteria isolated from spontaneously fermented quinoa sourdoughs. J. Appl. Microbiol. 2016, 120, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Sterr, Y.; Weiss, A.; Schmidt, H. Evaluation of lactic acid bacteria for sourdough fermentation of amaranth. Int. J. Food Microbiol. 2009, 136, 75–82. [Google Scholar] [CrossRef]
- Vogelmann, S.A.; Seitter, M.; Singer, U.; Brandt, M.J.; Hertel, C. Adaptability of lactic acid bacteria and yeasts to sourdoughs prepared from cereals, pseudocereals and cassava and use of competitive strains as starters. Int. J. Food Microbiol. 2009, 130, 205–212. [Google Scholar] [CrossRef]
- Palla, M.; Agnolucci, M.; Calzone, A.; Giovannetti, M.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G.; Pontonio, E. Exploitation of autochthonous Tuscan sourdough yeasts as potential starters. Int. J. Food Microbiol. 2019, 302, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhao, X.; Lin, X.B.; Gänzle, M.G. Comparative genomics Lactobacillus reuteri from sourdough reveals adaptation of an intestinal symbiont to food fermentations. Sci. Rep. 2015, 5, 18234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camu, N.; de Winter, T.; Verbrugghe, K.; Cleenwerck, I.; Vandamme, P.; Takrama, J.S.; Vancanneyt, M.; de Vuyst, L. Dynamics and biodiversity of populations of lactic acid bacteria and acetic acid bacteria involved in spontaneous heap fermentation of cocoa beans in Ghana. Appl. Environ. Microbiol. 2007, 73, 1809–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugas, M.; Garriga, M.; Aymerich, T.; Monfort, J.M. Biochemical characterization of lactobacilli from dry fermented sausages. Int. J. Food Microbiol. 1993, 18, 107–113. [Google Scholar] [CrossRef]
- Illeghems, K.; de Vuyst, L.; Weckx, S. Complete genome sequence and comparative analysis of Acetobacter pasteurianus 386B, a strain well-adapted to the cocoa bean fermentation ecosystem. BMC Genomics 2013, 14, 526. [Google Scholar] [CrossRef] [Green Version]
- Laureys, D.; Aerts, M.; Vandamme, P.; de Vuyst, L. The buffer capacity and calcium concentration of water influence the microbial species diversity, grain growth, and metabolite production during water kefir fermentation. Front. Microbiol. 2019, 10, 2876. [Google Scholar] [CrossRef] [Green Version]
- Laureys, D.; de Vuyst, L. Microbial species diversity, community dynamics, and metabolite kinetics of water kefir fermentation. Appl. Environ. Microbiol. 2014, 80, 2564–2572. [Google Scholar] [CrossRef] [Green Version]
- Moens, F.; Lefeber, T.; de Vuyst, L. Oxidation of metabolites highlights the microbial interactions and role of Acetobacter pasteurianus during cocoa bean fermentation. Appl. Environ. Microbiol. 2014, 80, 1848–1857. [Google Scholar] [CrossRef] [Green Version]
- Pelicaen, R.; Gonze, D.; Teusink, B.; de Vuyst, L.; Weckx, S. Genome-scale metabolic reconstruction of Acetobacter pasteurianus 386B, a candidate functional starter culture for cocoa bean fermentation. Front. Microbiol. 2020, 10, 2801. [Google Scholar] [CrossRef] [Green Version]
- Ravyts, F.; Barbuti, S.; Frustoli, M.A.; Parolari, G.; Saccani, G.; de Vuyst, L.; Leroy, F. Competitiveness and antibacterial potential of bacteriocin-producing starter cultures in different types of fermented sausages. J. Food Protect. 2008, 71, 1817–1827. [Google Scholar] [CrossRef]
- Sánchez Mainar, M.; Weckx, S.; Leroy, F. Coagulase-negative staphylococci favor conversion of arginine into ornithine despite a widespread genetic potential for nitric oxide synthase activity. Appl. Environ. Microbiol. 2014, 80, 7741–7751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verce, M.; de Vuyst, L.; Weckx, S. The metagenome-assembled genome of Candidatus Oenococcus aquikefiri from water kefir represents the species Oenococcus sicerae. Food Microbiol. 2020, 88, 103402. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Kim, Y.W.; Hwang, I.; Kim, J.; Yoon, S. Evaluation of Leuconostoc citreum HO12 and Weissella koreensis HO20 isolated from kimchi as a starter culture for whole wheat sourdough. Food Chem. 2012, 134, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Weckx, S. The cocoa bean fermentation process: From ecosystem analysis to starter culture development. J. Appl. Microbiol. 2016, 121, 1364–5072. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Functional role of yeasts, lactic acid bacteria, and acetic acid bacteria in cocoa fermentation processes. FEMS Microbiol. Rev. 2020, 44, 432–453. [Google Scholar] [CrossRef] [PubMed]
- Papalexandratou, Z.; Lefeber, T.; Bahrim, B.; Lee, O.; Daniel, H.M.; de Vuyst, L. Hanseniaspora opuntiae, Saccharomyces cerevisiae, Lactobacillus fermentum, and Acetobacter pasteurianus predominate during well-performed Malaysian cocoa bean box fermentations, underlining the importance of these microbial species for a successful cocoa bean fermentation process. Food Microbiol. 2013, 35, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Ravyts, F.; de Vuyst, L.; Leroy, F. Bacterial diversity and functionalities in food fermentations. Eng. Life Sci. 2012, 12, 356–367. [Google Scholar] [CrossRef]
- Laureys, D.; Aerts, M.; Vandamme, P.; de Vuyst, L. Oxygen and diverse nutrients influence the water kefir fermentation process. Food Microbiol. 2018, 73, 351–361. [Google Scholar] [CrossRef]
- Laureys, D.; de Vuyst, L. The water kefir grain inoculum determines the characteristics of the resulting water kefir fermentation process. J. Appl. Microbiol. 2017, 122, 719–732. [Google Scholar] [CrossRef]
- Li, Z.; Li, H.; Bian, K. Microbiological characterization of traditional dough fermentation starter (jiaozi) for steamed bread making by culture-dependent and culture-independent methods. Int. J. Food Microbiol. 2016, 234, 9–14. [Google Scholar] [CrossRef]
- Minervini, F.; Lattanzi, A.; de Angelis, M.; Di Cagno, R.; Gobbetti, M. Influence of artisan bakery- or laboratory-propagated sourdoughs on the diversity of lactic acid bacterium and yeast microbiotas. Appl. Environ. Microbiol. 2012, 78, 5328–5340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; He, G. Predominant bacteria diversity in Chinese traditional sourdough. J. Food Sci. 2013, 78, M1218–M1223. [Google Scholar] [CrossRef] [PubMed]
- Hermann, M.; Petermeier, H.; Vogel, R.F. Development of novel sourdoughs with in situ formed exopolysaccharides from acetic acid bacteria. Eur. Food Res. Technol. 2015, 241, 185–197. [Google Scholar] [CrossRef]
- Ua-Arak, T.; Jakob, F.; Vogel, R.F. Characterization of growth and exopolysaccharide production of selected acetic acid bacteria in buckwheat sourdoughs. Int. J. Food Microbiol. 2016, 239, 103–112. [Google Scholar] [CrossRef]
- Ua-Arak, T.; Jakob, F.; Vogel, R.F. Influence of levan-producing acetic acid bacteria on buckwheat-sourdough breads. Food Microbiol. 2017, 65, 95–104. [Google Scholar] [CrossRef]
- Ripari, V.; Cecchi, T.; Berardi, E. Microbiological characterisation and volatiles profile of model, ex-novo, and traditional Italian white wheat sourdoughs. Food Chem. 2016, 205, 297–307. [Google Scholar] [CrossRef]
- Ravyts, F.; Vrancken, G.; D’Hondt, K.; Vasilopoulos, C.; de Vuyst, L.; Leroy, F. Kinetics of growth and 3-methyl-1-butanol production by meat-borne, coagulase-negative staphylococci in view of sausage fermentation. Int. J. Food Microbiol. 2009, 134, 89–95. [Google Scholar] [CrossRef]
- Ravyts, F.; Steen, L.; Goemaere, O.; Paelinck, H.; de Vuyst, L.; Leroy, F. The application of staphylococci with flavour-generating potential is affected by acidification in fermented dry sausages. Food Microbiol. 2010, 27, 945–954. [Google Scholar] [CrossRef]
- Sánchez Mainar, M.; Stavropoulou, D.A.; Leroy, F. Exploring the metabolic heterogeneity of coagulase-negative staphylococci to improve the quality and safety of fermented meats: A review. Int. J. Food Microbiol. 2017, 247, 24–37. [Google Scholar] [CrossRef]
- Stavropoulou, D.A.; Borremans, W.; de Vuyst, L.; de Smet, S.; Leroy, F. Amino acid conversions by coagulase-negative staphylococci in a rich medium: Assessment of inter- and intraspecies heterogeneity. Int. J. Food Microbiol. 2015, 212, 34–40. [Google Scholar] [CrossRef]
- Verce, M.; de Vuyst, L.; Weckx, S. Comparative genomics of Lactobacillus fermentum suggests a free-living lifestyle of this lactic acid bacterial species. Food Microbiol. 2020, 89, 103488. [Google Scholar] [CrossRef] [PubMed]
- Meroth, C.B.; Walter, J.; Hertel, C.; Brandt, M.J.; Hammes, W.P. Monitoring the bacterial population dynamics in sourdough fermentation processes by using PCR-denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 2003, 69, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papalexandratou, Z.; Falony, G.; Romanens, E.; Jimenez, J.C.; Amores, F.; Daniel, H.M.; de Vuyst, L. Species diversity, community dynamics, and metabolite kinetics of the microbiota associated with traditional Ecuadorian spontaneous cocoa bean fermentations. Appl. Environ. Microbiol. 2011, 77, 7698–7714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Daniel, H.-M.; VAN Landschoot, A.; de Vuyst, L.; Vandamme, P. The microbial diversity of traditional spontaneously fermented lambic beer. PLoS ONE 2014, 9, e95384. [Google Scholar] [CrossRef]
- Ercolini, D.; Pontonio, E.; de Filippis, F.; Minervini, F.; La Storia, A.; Gobbetti, M.; Di Cagno, R. Microbial ecology dynamics during rye and wheat sourdough preparation. Appl. Environ. Microbiol. 2013, 79, 7827–7836. [Google Scholar] [CrossRef] [Green Version]
- Van Kerrebroeck, S.; Comasio, A.; Harth, H.; de Vuyst, L. Impact of starter culture, ingredients, and flour type on sourdough bread volatiles as monitored by selected ion flow tube-mass spectrometry. Food Res. Int. 2018, 106, 254–262. [Google Scholar] [CrossRef]
- Van Kerrebroeck, S.; Harth, H.; Comasio, A.; de Vuyst, L. Monitoring of starter culture-initiated liquid wheat and teff sourdough fermentations by selected ion flow tube-mass spectrometry. J. Sci. Food Agric. 2018, 98, 3501–3512. [Google Scholar] [CrossRef]
- Lefeber, T.; Papalexandratou, Z.; Gobert, W.; Camu, N.; de Vuyst, L. On-farm implementation of a starter culture for improved cocoa bean fermentation and its influence on the flavour of chocolates produced thereof. Food Microbiol. 2012, 30, 379–392. [Google Scholar] [CrossRef]
- Illeghems, K.; de Vuyst, L.; Weckx, S. Comparative genome analysis of the candidate functional starter culture strains Lactobacillus fermentum 222 and Lactobacillus plantarum 80 for controlled cocoa bean fermentation processes. BMC Genomics 2015, 16, 766. [Google Scholar] [CrossRef] [Green Version]
- Vrancken, G.; Rimaux, T.; de Vuyst, L.; Leroy, F. Kinetic analysis of growth and sugar consumption by Lactobacillus fermentum IMDO 130101 reveals adaptation to the acidic sourdough ecosystem. Int. J. Food Microbiol. 2008, 128, 58–66. [Google Scholar] [CrossRef]
- Rimaux, T.; Vrancken, G.; Pothakos, V.; Maes, D.; de Vuyst, L.; Leroy, F. The kinetics of the arginine deiminase pathway in the meat starter culture Lactobacillus sakei CTC 494 are pH-dependent. Food Microbiol. 2011, 28, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Rimaux, T.; Vrancken, G.; Vuylsteke, B.; de Vuyst, L.; Leroy, F. The pentose moiety of adenosine and inosine is an important energy source for the fermented-meat starter culture Lactobacillus sakei CTC 494. Appl. Environ. Microbiol. 2011, 77, 6539–6550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cousin, F.J.; Le Guellec, R.; Chagnot, C.; Goux, D.; Dalmasso, M.; Laplace, J.M.; Cretenet, M. Oenococcus sicerae sp. nov., isolated from French cider. Syst. Appl. Microbiol. 2019, 42, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Laureys, D.; Cnockaert, M.; de Vuyst, L.; Vandamme, P. Bifidobacterium aquikefiri sp. nov., isolated from water kefir. Int. J. Syst. Evol. Microbiol. 2016, 66, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Verce, M.; de Vuyst, L.; Weckx, S. Shotgun metagenomics of a water kefir fermentation ecosystem reveals a novel Oenococcus species. Front. Microbiol. 2019, 10, 479. [Google Scholar] [CrossRef]
- Zheng, J.; Ruan, L.; Sun, M.; Gänzle, M.G. A genomic view of lactobacilli and pediococci demonstrates that phylogeny matches ecology and physiology. Appl. Environ. Microbiol. 2015, 81, 7233–7243. [Google Scholar] [CrossRef] [Green Version]
- Edwards, C.G.; Collins, M.D.; Lawson, P.A.; Rodriguez, A.V. Lactobacillus nagelii sp.nov., an organism isolated from a partially fermented wine. Int. J. Syst. Evol. Microbiol. 2000, 50, 699–702. [Google Scholar] [CrossRef] [Green Version]
- Stavropoulou, D.A.; de Vuyst, L.; Leroy, F. Nonconventional starter cultures of coagoulase-negative staphylococci to produce animal-derived fermented foods, a SWOT analysis. J. App. Microbiol. 2018, 125, 1570–1586. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, K.; Toyama, H.; Tonouchi, N.; Okamoto-Kainuma, A. Acetic Acid Bacteria: Ecology and Physiology, 1st ed.; Springer: Tokyo, Japan, 2016. [Google Scholar]
- Prust, C.; Hoffmeister, M.; Liesegang, H.; Wiezer, A.; Fricke, W.F.; Ehrenreich, A.; Gottschalk, G.; Deppenmeier, U. Complete genome sequence of the acetic acid bacterium Gluconobacter oxydans. Nat. Biotechnol. 2005, 23, 195–200. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, V.K.; Qazi, G.N.; Kumar, A. Gluconobacter oxydans: Its biotechnological applications. J. Mol. Microbiol. Biotechnol. 2001, 3, 445–456. [Google Scholar]
- Claret, C.; Salmon, J.M.; Romieu, C.; Bories, A. Physiology of Gluconobacter oxydans during dihydroxyacetone production from glycerol. Appl. Microbiol. Biotechnol. 1994, 41, 359–365. [Google Scholar] [CrossRef]
- Hu, Z.C.; Zheng, Y.G.; Schen, Y.C. Dissolved-oxygen-stat fed-batch fermentation of 1,3-dihydroxyacetone from glycerol by Gluconobacter oxydans ZJB09112. Biotechnol. Bioprocess Eng. 2010, 15, 651–656. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Starter Culture Strain (References) | Source of Isolation | Sourdough Production Code | Ingredient Supplementation of the Flour–Water Mixture and Specific Fermentation Conditions |
|---|---|---|---|
| Lactic acid bacteria (LAB) | |||
| Limosilactobacillus fermentum IMDO 222 [35,63] | Fermenting cocoa pulp-bean mass | LF1, LF2 | None |
| Latilactobacillus sakei CTC 494 [36,42] | Fermented sausage | LS1, LS2 | None |
| Lentilactobacillus hilgardii IMDO 2M0G15 [38,39] | Water kefir | LH1, LH2 | None (LH1) or sucrose (40 g/L, LH2) a |
| Liquorilactobacillus nagelii IMDO 2M48G6 [38,39] | Water kefir | LN | None |
| Coagulase-negative staphylococci (CNS) | |||
| Staphylococcus carnosus IMDO F4P1 [43] | Fermented sausage | SC1, SC2 | None (SC1) or NaCl (40 g/L, SC2) b |
| Acetic acid bacteria (AAB) | |||
| Gluconobacter oxydans IMDO A845 [35,47] | Fermenting cocoa pulp-bean mass | GO1, GO2, GO3 | Acetic acid (3 g/L), ethanol (10 g/L), and aeration (5 L/min) through the wheat flour-water mixture (GO1, GO2) or the fermenter headspace (GO3) c |
| Acetobacter pasteurianus IMDO 386B [35,37,40,41] | Fermenting cocoa pulp-bean mass | AP | Acetic acid (3 g/L), ethanol (10 g/L), and aeration (5 L/min) through the wheat flour-water mixture c |
| PC1 | PC2 | PC3 |
|---|---|---|
| 1,3-Octadiene | 2,4-Heptadienal (E,E) | 1-Decanol |
| 1-Nitro-hexane | 2-Ethylhexanol | 1-Ethyl-1-methylcyclopentane |
| 1-Undecanol | 2-Octanol | 1-Heptanol |
| 2,4-Decadienal (E,E) | 3-Nonen-1-ol | 1-Nonanol |
| 2,4-Nonadienal (E,E) | 3-Nonenyl acetate | 1-Octanol |
| 2,5-Octanedione | 4-Methyl-2-pentanol | 2-Nonanone |
| 2-Cyclopentene-1,4-dione | Ethyl acetate | 2-Nonen-1-ol |
| 2-Decenal, (E) | Ethyldecanoate | 2-Octanone |
| 2-Ethyl-3-methyl-pyrazine | Heptyl acetate | 3,5-Octadien-2-ol |
| 2-Furanmethanol | Hexyl acetate | 3-Methylbutanoic acid |
| 2-Methoxyphenol | Hexyl-2-methoxyacetate | 3-Octanone |
| 2-Methyl-5-(1-methylethenyl)-cyclohexanol | Isobutyloctanoate | Butanoic acid |
| 2-Nonenal (E) | Isoledene | Dodecane |
| 2-Octenal (E) | Methylpyrazine | Ethyl heptanoate |
| 2-Pentylfuran | Octyl acetate | |
| 2-Propylphenol | Pentyl acetate | |
| 2-Undecenal | Vinylhexanoate | |
| 3-Methyl-6-propyl-phenol | ||
| 4-Oxononanal | ||
| 5-Ethylcyclopent-1-enecarboxaldehyde | ||
| 5-Methyl-2-furancarboxaldehyde | ||
| 6-Methyl-5-hepten-2-one | ||
| Acetic acid | ||
| Decanal | ||
| Ethylpyrazine | ||
| Hexanal | ||
| Nonanal | ||
| Octanal | ||
| Pentanal | ||
| Precocene 1 | ||
| Tetradecane | ||
| Toluene | ||
| Trimethylsilyl-di(trimethylsiloxy)-silane |
| PC1 | PC2 | PC3 |
|---|---|---|
| 1-Decanol | 1-Octen-3-one | 2,4-Decadien-1-ol (E,Z) |
| 1-Heptanol | 2,3-Pentanedione | 3,5-Octadien-2-one |
| 1-Octanol | 2,3-Butanedione | 3-Methyl-1-butanol |
| 1-Octen-3-ol | 2-Methylnaphthalene | 3-Methylthiopropanol |
| 2,3-Octanedione | Butanoic acid | 3-Nonen-1-ol (Z) |
| 2,5-Octanedione | Dodecane | 3-Nonenyl acetate |
| 2-Heptanal (Z) | Ethanol | Ethyl decanoate |
| 2-Octanone | Nonanoic acid | Hexyl acetate |
| 2-Octenal (E) | Naphthalene | Vinyl caproate |
| 2-Pentylfuran | Octanoic acid | |
| 3-Methyl-6-propyl-phenol | ||
| 5-Methyl-2-furancarboxaldehyde | ||
| Benzaldehyde | ||
| Ethyl octanoate | ||
| Hexanal | ||
| Octanal |
| Sourdough Used for Wheat Bread Production | Crumb Color | Crust Color | ||||
|---|---|---|---|---|---|---|
| L * | a * | b * | L * | a * | b * | |
| Reference | 73.1 ± 5.3 a | 0.3 ± 0.3 a | 16.6 ± 1.1 | 60.0±4.3 a,d | 11.7 ± 2.1 a | 32.5 ± 2.2 a,b |
| Sparged, 0 h | 72.5 ± 4.2 a | 0.1 ± 0.1 a | 15.9 ± 1.2 | 60.8±5.8 a,d | 11.5 ± 3.4 a | 33.1 ± 1.5 a,b |
| Sparged, 24 h | 72.3 ± 4.2 a | 0.2 ± 0.2 a | 16.5 ± 2.1 | 59.8±3.0 a,c,d | 12.3 ± 2.6 a | 35.2 ± 1.6 a,b |
| Sparged, 48 h | 45.5 ± 3.6 b | 9.1 ± 0.6 b | 17.6 ± 1.3 | 44.9±3.5 b,d | 14.1 ± 0.7 a | 24.4 ± 0.4 c,d |
| Sparged, 72 h | 51.3 ± 1.2 b | 8.4 ± 0.3 b | 17.6 ± 0.8 | 44.6±2.0 b,d | 14.0 ± 1.4 a | 23.2 ± 1.9 c |
| Aerated, 72 h | 72.9 ± 1.5 a | 0.7 ± 1.1 a,c | 16.4 ± 2.5 | 62.8±2.7 a,d | 12.1 ± 1.8 a | 36.9 ± 2.7 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Comasio, A.; Van Kerrebroeck, S.; Harth, H.; Verté, F.; De Vuyst, L. Potential of Bacteria from Alternative Fermented Foods as Starter Cultures for the Production of Wheat Sourdoughs. Microorganisms 2020, 8, 1534. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101534
Comasio A, Van Kerrebroeck S, Harth H, Verté F, De Vuyst L. Potential of Bacteria from Alternative Fermented Foods as Starter Cultures for the Production of Wheat Sourdoughs. Microorganisms. 2020; 8(10):1534. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101534
Chicago/Turabian StyleComasio, Andrea, Simon Van Kerrebroeck, Henning Harth, Fabienne Verté, and Luc De Vuyst. 2020. "Potential of Bacteria from Alternative Fermented Foods as Starter Cultures for the Production of Wheat Sourdoughs" Microorganisms 8, no. 10: 1534. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101534