Effects of Different Stress Parameters on Growth and on Oleuropein-Degrading Abilities of Lactiplantibacillus plantarum Strains Selected as Tailored Starter Cultures for Naturally Table Olives


 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Culture Conditions
2.2. Growth Ability of L. plantarum Strains under Specific Stress Conditions
2.3. L. plantarum Beta-Glucosidase Gene Detection
2.4. Oleuropein-Degrading Ability Test
2.5. Statistical Analysis
3. Results and Discussion
3.1. Growth Performances Exhibited by the Tested Strains
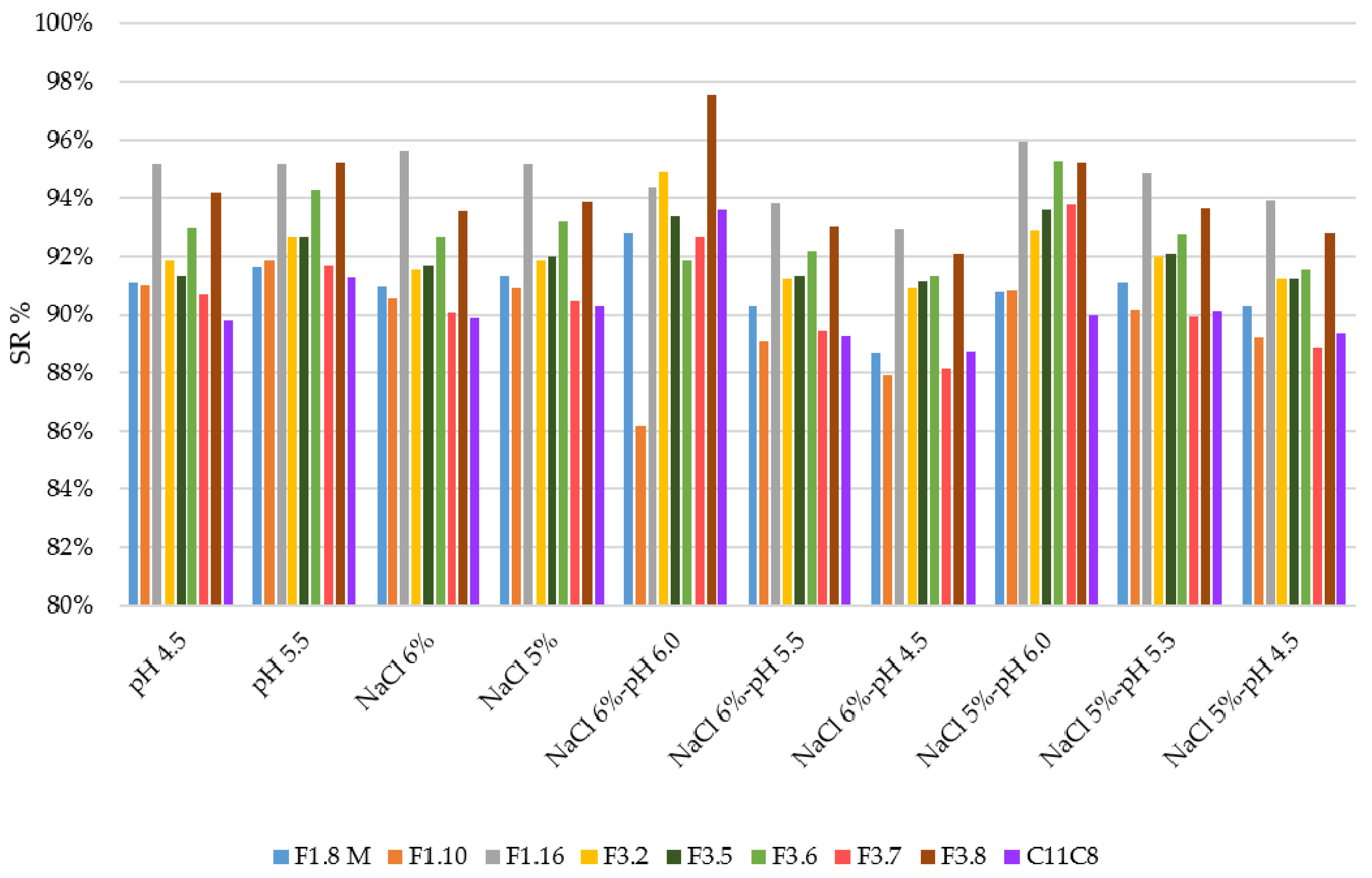
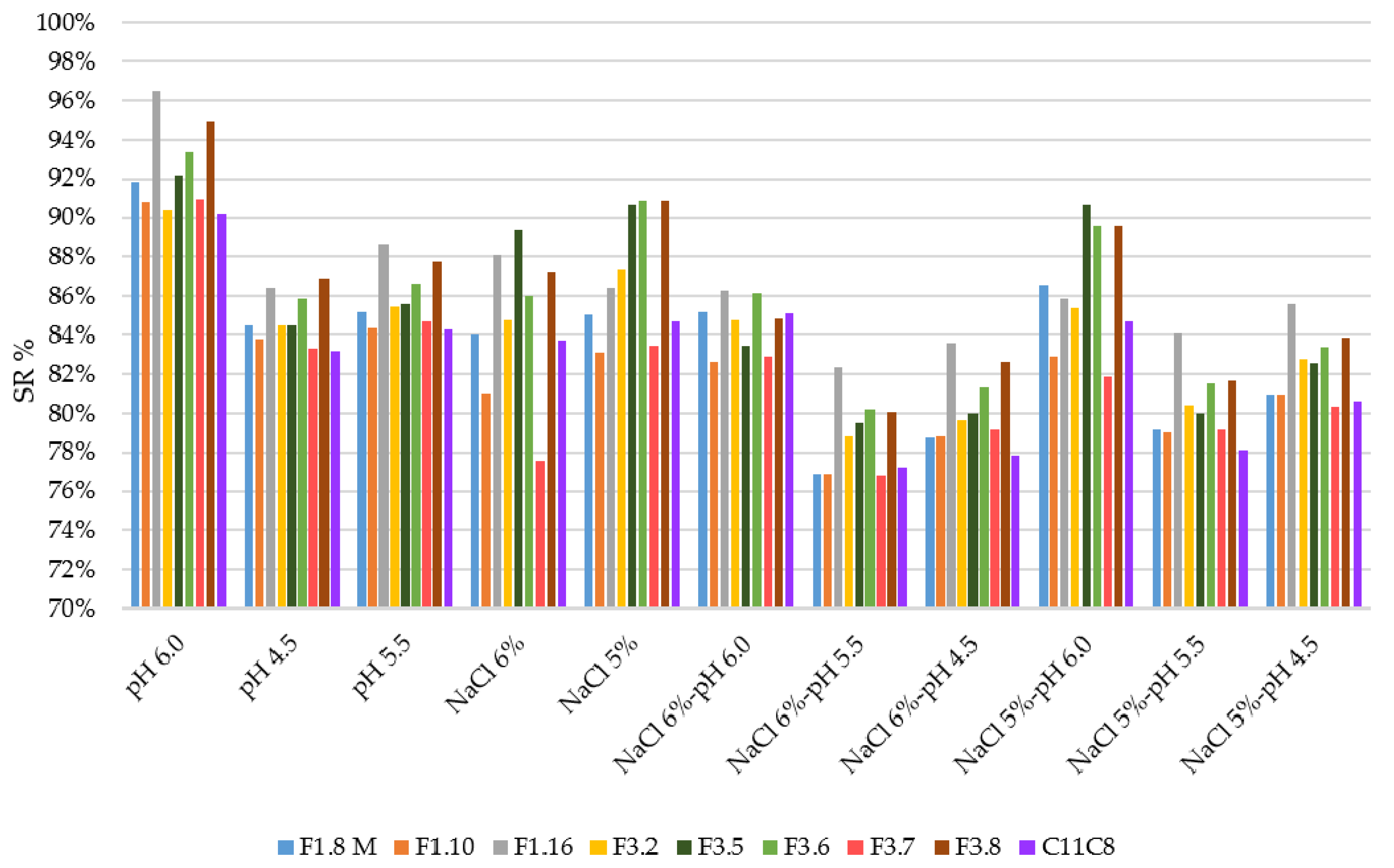
3.2. Growth Ability under Stress Conditions
3.3. Molecular Detection of Beta-Glucosidase Gene in L. plantarum Strains
3.4. Oleuropein Degrading Test
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lavermicocca, P.; Valerio, F.; Lonigro, S.L.; De Angelis, M.; Morelli, L.; Callegari, M.L.; Rizzello, C.G.; Visconti, A. Adhesion and survival of lactobacilli and bifidobacteria on table olives with the aim of formulating a new probiotic food. Appl. Environ. Microbiol. 2005, 71, 4233–4240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botta, C.; Cocolin, L. Microbial dynamics and biodiversity in table olive fermentation: Culture-dependent and-independent approaches. Front. Microbiol. 2012, 3, 245. [Google Scholar] [CrossRef] [Green Version]
- Othman, N.B.; Roblain, D.; Chammen, N.; Thonart, P.; Hamdi, M. Antioxidant phenolic compounds loss during the fermentation of Chétoui olives. Food Chem. 2009, 116, 662–669. [Google Scholar] [CrossRef]
- Perpetuini, G.; Caruso, G.; Urbani, S.; Schirone, M.; Esposto, S.; Ciarrocchi, A.; Prete, R.; Garcia-Gonzalez, N.; Battistelli, N.; Gucci, R.; et al. Changes in polyphenolic concentrations of table olives (cv. Itrana) produced under different irrigation regimes during spontaneous or inoculated fermentation. Front. Microbiol. 2018, 9, 1287. [Google Scholar] [CrossRef]
- de Castro, A.; Romero, C.; Brenes, M. A new polymer inhibitor of Lactobacillus growth in table olives. Eur. Food Res. Technol. 2005, 221, 192–196. [Google Scholar] [CrossRef]
- Medina, E.; Romero, C.; de Castro, A.; Brenes, M.; García, A. Inhibitors of lactic acid fermentation in Spanish-style green olive brines of the Manzanilla variety. Food Chem. 2008, 110, 932–937. [Google Scholar] [CrossRef]
- Medina, E.; García, A.; Romero, C.; de Castro, A.; Brenes, M. Study of the anti-lactic acid bacteria compounds in table olives. Int. J. Food Sci. Technol. 2009, 44, 1286–1291. [Google Scholar] [CrossRef]
- Vaccalluzzo, A.; Pino, A.; Russo, N.; De Angelis, M.; Caggia, C.; Randazzo, C.L. FoodOmics as a new frontier to reveal microbial community and metabolic processes occurring on table olives fermentation. Food Microbiol. 2020, 103606. [Google Scholar] [CrossRef]
- Bianchi, G. Lipids and phenols in table olives. Eur. J. Lipid Sci. Technol. 2003, 105, 229–242. [Google Scholar] [CrossRef]
- Tofalo, R.; Schirone, M.; Perpetuini, G.; Angelozzi, G.; Suzzi, G.; Corsetti, A. Microbiological and chemical profiles of naturally fermented table olives and brines from different Italian cultivars. Antonie Van Leeuwenhoek 2012, 102, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.; Gonçalves, T.; Quintas, C. Microbial quality and yeast population dynamics in cracked green table olives’ fermentations. Food Control 2012, 23, 363–368. [Google Scholar] [CrossRef]
- Bonatsou, S.; Tassou, C.C.; Panagou, E.Z.; Nychas, G.J.E. Table olive fermentation using starter cultures with multifunctional potential. Microorganisms 2017, 5, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsetti, A.; Perpetuini, G.; Schirone, M.; Tofalo, R.; Suzzi, G. Application of starter cultures to table olive fermentation: An overview on the experimental studies. Front. Microbiol. 2012, 3, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Lactic acid bacteria from fermented table olives. Food Microbiol. 2012, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rozes, N.; Peres, C. Effect of oleuropein and sodium chloride on viability and metabolism of Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 1996, 45, 839–843. [Google Scholar] [CrossRef]
- Marsilio, V.; Lanza, B.; Pozzi, N. Progress in table olives debittering: Degradation in vitro of oleuropein and its derivatives by L. plantarum. J. Am. Oil Chem. Soc. 1996, 73, 593–597. [Google Scholar] [CrossRef]
- Marsilio, V.; Lanza, B. Characterization of an oleuropein degrading strain of Lactobacillus plantarum. Combined effects of compounds present in olive fermenting brines (phenols, glucose and NaCl) on bacterial activity. J. Sci. Food Agric. 1998, 76, 520–524. [Google Scholar] [CrossRef]
- Landete, J.M.; Curiel, J.A.; Rodriguez, H.; de la Rivas, B.; Munoz, R. Study of the inhibitory activity of phenolic compounds found in olive products and their degradation by Lactobacillus plantarum strains. Food Chem. 2008, 107, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Ghabbour, N.; Lamzira, Z.; Thonart, P.; Cidalia, P.; Markaoui, M.; Asehraou, A. Selection of oleuropein-degrading lactic acid bacteria strains isolated from fermenting Moroccan green olives. Grasas Aceites 2011, 62, 84–89. [Google Scholar]
- Kaltsa, A.; Papaliaga, D.; Papaioannou, E.; Kotzekidou, P. Characteristics of oleuropeinolytic strains of Lactobacillus plantarum group and influence on phenolic compounds in table olives elaborated under reduced salt conditions. Food Microbiol. 2015, 48, 58–62. [Google Scholar] [CrossRef]
- Marasco, R.; Muscariello, L.; Varcamonti, M.; De Felice, M.; Sacco, M. Expression of the bglH gene of Lactobacillus plantarum is controlled by carbon catabolite repression. J. Bacteriol. 1998, 180, 3400–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gury, J.; Barthelmebs, L.; Tran, N.P.; Divi es, C.; Cavin, J.-F. Cloning, deletion, and characterization of PadR, the transcriptional repressor of the phenolic acid decarboxylase-encoding padA gene of Lactobacillus plantarum. Appl. Environ. Microbiol. 2004, 70, 2146–2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, N.; Esteban-Torres, M.; Mancheño, J.M.; de las Rivas, B.; Muñoz, R. Tannin degradation by a novel tannase enzyme present in some Lactobacillus plantarum strains. Appl. Environ. Microbiol. 2014, 80, 2991–2997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spano, G.; Rinaldi, A.; Ugliano, M.; Moio, L.; Beneduce, L.; Massa, S. A β-glucosidase gene isolated wine Lactobacillus plantarum is regulated by abiotic stresses. J. Appl. Microbiol. 2005, 98, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Pino, A.; Vaccalluzzo, A.; Solieri, L.; Romeo, F.V.; Todaro, A.; Caggia, C.; Arroyo-López, F.N.; Bautista-Gallego, J.; Randazzo, C.L. Effect of Sequential Inoculum of Beta-Glucosidase Positive and Probiotic Strains on Brine Fermentation to Obtain Low Salt Sicilian Table Olives. Front. Microbiol. 2019, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Zago, M.; Lanza, B.; Rossetti, L.; Muzzalupo, I.; Carminati, D.; Giraffa, G. Selection of Lactobacillus plantarum strains to use as starters in fermented table olives: Oleuropeinase activity and phage sensitivity. Food Microbiol. 2013, 34, 81–87. [Google Scholar] [CrossRef]
- Iorizzo, M.; Lombardi, S.J.; Macciola, V.; Testa, B.; Lustrato, G.; Lopez, F.; De Leonardis, A. Technological potential of Lactobacillus strains isolated from fermented green olives: In vitro studies with emphasis on oleuropein-degrading capability. Sci. World J. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yao, W.; Yang, L.; Shao, Z.; Xie, L.; Chen, L. Identification of salt tolerance-related genes of Lactobacillus plantarum D31 and T9 strains by genomic analysis. Ann. Microbiol. 2020, 70, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ghabbour, N.; Rokni, Y.; Abouloifa, H.; Bellaouchi, R.; Chihib, N.E.; Salah, R.B.; Lamzira, Z.; Saalaoui, E.; Asehraou, A. In vitro biodegradation of oleuropein by Lactobacillus plantarum FSO175 in stress conditions (pH, NaCl and glucose). J. Microbiol. Biotechnol. Food Sci. 2020, 9, 769–773. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Altieri, C.; Corbo, M.R.; Sinigaglia, M.; Ouoba, L.I.I. Characterization of lactic acid bacteria isolated from Italian Bella di Cerignola table olives: Selection of potential multifunctional starter cultures. J. Food Sci. 2010, 75, M536–M544. [Google Scholar] [CrossRef]
- Aponte, M.; Blaiotta, G.; La Croce, F.; Mazzaglia, A.; Farina, V.; Settanni, L.; Moschetti, G. Use of selected autochthonous lactic acid bacteria for Spanish-style table olive fermentation. Food Microbiol. 2012, 30, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Pino, A.; De Angelis, M.; Todaro, A.; Van Hoorde, K.; Randazzo, C.L.; Caggia, C. Fermentation of Nocellara Etnea table olives by functional starter cultures at different low salt concentrations. Front. Microbiol. 2018, 9, 1125. [Google Scholar] [CrossRef] [Green Version]
- Sorrentino, G.; Muzzalupo, I.; Muccilli, S.; Timpanaro, N.; Russo, M.P.; Guardo, M.; Rapisarda, P.; Romeo, F.V. New accessions of Italian table olives (Olea europaea): Characterization of genotypes and quality of brined products. Sci. Hortic. 2016, 213, 34–41. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Fava, G.; Tomaselli, F.; Romeo, F.V.; Pennino, G.; Vitello, E.; Caggia, C. Effect of kaolin and copper based products and of starter cultures on green table olive fermentation. Food Microbiol. 2011, 28, 910–919. [Google Scholar] [CrossRef]
- Carvalho, A.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X.; Gibbs, P. Relevant factors for the preparation of freeze-dried lactic acid bacteria. Int. Dairy J. 2004, 14, 835–847. [Google Scholar] [CrossRef]
- De Angelis, M.; Gobbetti, M. Stress responses of lactobacilli. In Stress Responses of Lactic Acid Bacteria; Papadimitriou, K., Tsakalidou, E., Eds.; Springer Science & Business Media LCC: New York, NY, USA, 2011; pp. 219–249. [Google Scholar]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; Van’t Riet, K.J.A.E.M. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Wang, W.; Sun, C.; Gu, L.; Liu, Z.; Yu, W.; Chen, L.; Jiang, Z.; Hou, J. Effects of environmental stresses on the physiological characteristics, adhesion ability and pathogen adhesion inhibition of Lactobacillus plantarum KLDS 1.0328. Process Biochem. 2020, 92, 426–436. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Restuccia, C.; Romano, A.D.; Caggia, C. Lactobacillus casei, dominant species in naturally fermented Sicilian green olives. Int. J. Food Microbiol. 2004, 90, 9–14. [Google Scholar] [CrossRef]
- Servili, M.; Settanni, L.; Veneziani, G.; Esposto, S.; Massitti, O.; Taticchi, A.; Urbani, S.; Montedoro, G.F.; Corsetti, A. The use of Lactobacillus pentosus 1MO to shorten the debittering process time of black table olives (Cv. Itrana and Leccino): A pilot-scale application. J. Agric. Food Chem. 2006, 54, 3869–3875. [Google Scholar] [CrossRef]
- Romeo, F.; Poiana, M. Ability of commercially available Lactobacillus strains as starter in brining and debittering of table olives. Acta Aliment. 2007, 36, 49–60. [Google Scholar] [CrossRef]
- Marasco, R.; Salatiello, I.; De Felice, M.; Sacco, M. A physical and functional analysis of the newly-identified bglGPT operon of Lactobacillus plantarum. FEMS Microbiol. Lett. 2000, 186, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Kara, H.E.; Sinan, S.; Turan, Y. Purification of beta-glucosidase from olive (Olea europaea L.) fruit tissue with specifically designed hydrophobic interaction chromatography and characterization of the purified enzyme. J. Chromatogr. B. 2011, 879, 1507–1512. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, J.A.; Lucena-Padrós, H.; Brenes, M.; Ruiz-Barba, J.L. Expression of genes involved in metabolism of phenolic compounds by Lactobacillus pentosus and its relevance for table-olive fermentations. Food Microbiol. 2018, 76, 382–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.J.; Wang, C.Z.; Ye, J.Z.; Tao, R.; Zhang, Y.S. Enzymatic hydrolysis of oleuropein from Olea europea (Olive) leaf extract and antioxidant activities. Molecules 2015, 20, 2903–2921. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| L. plantarum Strains | Control Condition (MRS Broth Medium pH 6.0) | Stress Condition (MRS Broth Medium pH 6.0 and NaCl 6%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | μmax | λ | A | μmax | λ | |||||||
| 32 °C | 16 °C | 32 °C | 16 °C | 32 °C | 16 °C | 32 °C | 16 °C | 32 °C | 16 °C | 32 °C | 16 °C | |
| F1.8M | 2.63 ± 0.02 | 0.43 ± 0.02 | 0.16 ± 0.02 | 0.01 ± 0.01 | 13.57 ± 0.08 | 20.02 ± 0.02 | 0.51 ± 0.04 | 0.46 ± 0.02 | 0.03 ± 0.02 | 0.03 ± 0.02 | 29.99 ± 0.07 | 32.91 ± 0.07 |
| F1.10 | 2.79 ± 0.02 | 0.38 ± 0.03 | 0.15 ± 0.03 | 0.02 ± 0.02 | 13.20 ± 0.05 | 20.80 ± 0.15 | 0.14 ± 0.02 | 0.12 ± 0.03 | 0.03 ± 0.02 | 0.03 ± 0.03 | 22.82 ± 0.06 | 24.15 ± 0.04 |
| F1.16 | 1.15 ± 0.03 | 0.45 ± 0.02 | 0.05 ± 0.02 | 0.02 ± 0.01 | 21.42 ± 0.04 | 19.96 ± 0.02 | 0.32 ± 0.03 | 0.29 ± 0.02 | 0.02 ± 0.02 | 0.02 ± 0.02 | 20.92 ± 0.06 | 22.77 ± 0.03 |
| F3.2 | 2.26 ± 0.02 | 0.26 ± 0.02 | 0.09 ± 0.02 | 0.02 ± 0.02 | 16.06 ± 0.03 | 20.23 ± 0.03 | 0.75 ± 0.03 | 0.59 ± 0.02 | 0.02 ± 0.01 | 0.01 ± 0.01 | 23.73 ± 0.05 | 26.22 ± 0.04 |
| F3.5 | 2.25 ± 0.02 | 0.50 ± 0.02 | 0.09 ± 0.02 | 0.01 ± 0.01 | 16.07 ± 0.02 | 18.76 ± 0.02 | 0.51 ± 0.03 | 0.49 ± 0.02 | 0.03 ± 0.03 | 0.07 ± 0.02 | 30.03 ± 0.03 | 31.48 ± 0.08 |
| F3.6 | 1.47 ± 0.05 | 0.38 ± 0.05 | 0.05 ± 0.04 | 0.02 ± 0.02 | 19.11 ± 0.02 | 24.29 ± 0.04 | 0.28 ± 0.02 | 0.25 ± 0.02 | 0.01 ± 0.01 | 0.01 ± 0.01 | 30.03 ± 0.05 | 24.16 ± 0.05 |
| F3.7 | 2.87 ± 0.02 | 0.44 ± 0.04 | 0.18 ± 0.02 | 0.08 ± 0.07 | 14.12 ± 0.03 | 15.50 ± 0.05 | 0.58 ± 0.02 | 0.49 ± 0.03 | 0.02 ± 0.01 | 0.02 ± 0.02 | 23.80 ± 0.06 | 25.29 ± 0.06 |
| F3.8 | 1.17 ± 0.04 | 0.43 ± 0.03 | 0.05 ± 0.04 | 0.02 ± 0.01 | 21.29 ± 0.03 | 19.83 ± 0.03 | 0.74 ± 0.04 | 0.52 ± 0.04 | 0.02 ± 0.02 | 0.02 ± 0.02 | 22.98 ± 0.11 | 24.47 ± 0.07 |
| C11C8 | 2.39 ± 0.03 | 0.38 ± 0.02 | 0.16 ± 0.04 | 0.02 ± 0.02 | 13.35 ± 0.04 | 24.26 ± 0.06 | 0.80 ± 0.04 | 0.60 ± 0.06 | 0.03 ± 0.01 | 0.02 ± 0.01 | 21.22 ± 0.03 | 22.89 ± 0.06 |
| MRS Broth, 0.1% (w/v) of OLE, NaCl 5.0% (w/v), pH 6.0 | MRS Broth, 0.1% (w/v) of OLE, NaCl 6.0% (w/v), pH 6.0 | |||||||
|---|---|---|---|---|---|---|---|---|
| 32 °C | 16 °C | 32 °C | 16 °C | |||||
| OLE (mg/L) | OLE (%) | OLE (mg/L) | OLE (%) | OLE (mg/L) | OLE (%) | OLE (mg/L) | OLE (%) | |
| Control | 975.0 ± 2.71 a | 00.0 | 985.0 ± 3.00 a | 00.0 | 978.0 ± 2.71 a | 00.0 | 983.0 ± 3.00 a | 00.0 |
| L. plantarum Strains | ||||||||
| F1.8M | 39.8 ± 0.04 h | 95.9 | 165.7 ± 11.98 d | 83.2 | 44.9 ± 2.83 g,h | 95.4 | 93.7 ± 6.47 d | 90.5 |
| F1.10 | 45.6 ± 1.60 h | 95.3 | 110.3 ± 13.60 f,g | 88.8 | 66.9 ± 2.56 e,f | 93.2 | 93.9 ± 13.31 d | 90.4 |
| F1.16 | 82.7 ± 0.25 g | 91.5 | 123.7 ± 1.05 e,f | 87.4 | 21.7 ± 0.51 i | 97.8 | 38.8 ± 0.91 g,h,i | 96.0 |
| F3.2 | 129.8 ± 0.64 e,f | 86.7 | 125.9 ± 0.55 e,f | 87.2 | 46.8 ± 1.19 g | 95.2 | 49.6 ± 1.57 f,g | 95.0 |
| F3.5 | 102.3 ± 0.18 f,g | 89.5 | 801.2 ± 10.52 b | 18.7 | 46.8 ± 0.77 g | 95.2 | 752.8 ± 2.54 b | 23.4 |
| F3.6 | 84.8 ± 2.66 g | 91.3 | 785.0 ± 7.72 b | 20.3 | 26.9 ± 1.57 i | 97.3 | 723.0 ± 3.28 c | 26.4 |
| F3.7 | 41.2 ± 3.57 h | 95.8 | 213.4 ± 21.63 c | 78.3 | 69.1 ± 0.26 e | 92.9 | 94.7 ± 0.11 d | 90.4 |
| F3.8 | 44.8 ± 1.85 h | 95.4 | 150.8 ± 1.51 d,e | 84.7 | 35.5 ± 4.11 g,h,i | 96.4 | 96.8 ± 8.34 d | 90.2 |
| C11C8 | 88.1 ± 0.13 g | 91.0 | 126.6 ± 2.26 e,f | 87.1 | 27.9 ± 0.61 h,i | 97.2 | 49.3 ± 1.37 g | 95.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaccalluzzo, A.; Pino, A.; De Angelis, M.; Bautista-Gallego, J.; Romeo, F.V.; Foti, P.; Caggia, C.; Randazzo, C.L. Effects of Different Stress Parameters on Growth and on Oleuropein-Degrading Abilities of Lactiplantibacillus plantarum Strains Selected as Tailored Starter Cultures for Naturally Table Olives. Microorganisms 2020, 8, 1607. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101607
Vaccalluzzo A, Pino A, De Angelis M, Bautista-Gallego J, Romeo FV, Foti P, Caggia C, Randazzo CL. Effects of Different Stress Parameters on Growth and on Oleuropein-Degrading Abilities of Lactiplantibacillus plantarum Strains Selected as Tailored Starter Cultures for Naturally Table Olives. Microorganisms. 2020; 8(10):1607. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101607
Chicago/Turabian StyleVaccalluzzo, Amanda, Alessandra Pino, Maria De Angelis, Joaquín Bautista-Gallego, Flora Valeria Romeo, Paola Foti, Cinzia Caggia, and Cinzia L Randazzo. 2020. "Effects of Different Stress Parameters on Growth and on Oleuropein-Degrading Abilities of Lactiplantibacillus plantarum Strains Selected as Tailored Starter Cultures for Naturally Table Olives" Microorganisms 8, no. 10: 1607. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8101607