Impact of Inoculum Type on the Microbial Community and Power Performance of Urine-Fed Microbial Fuel Cells

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
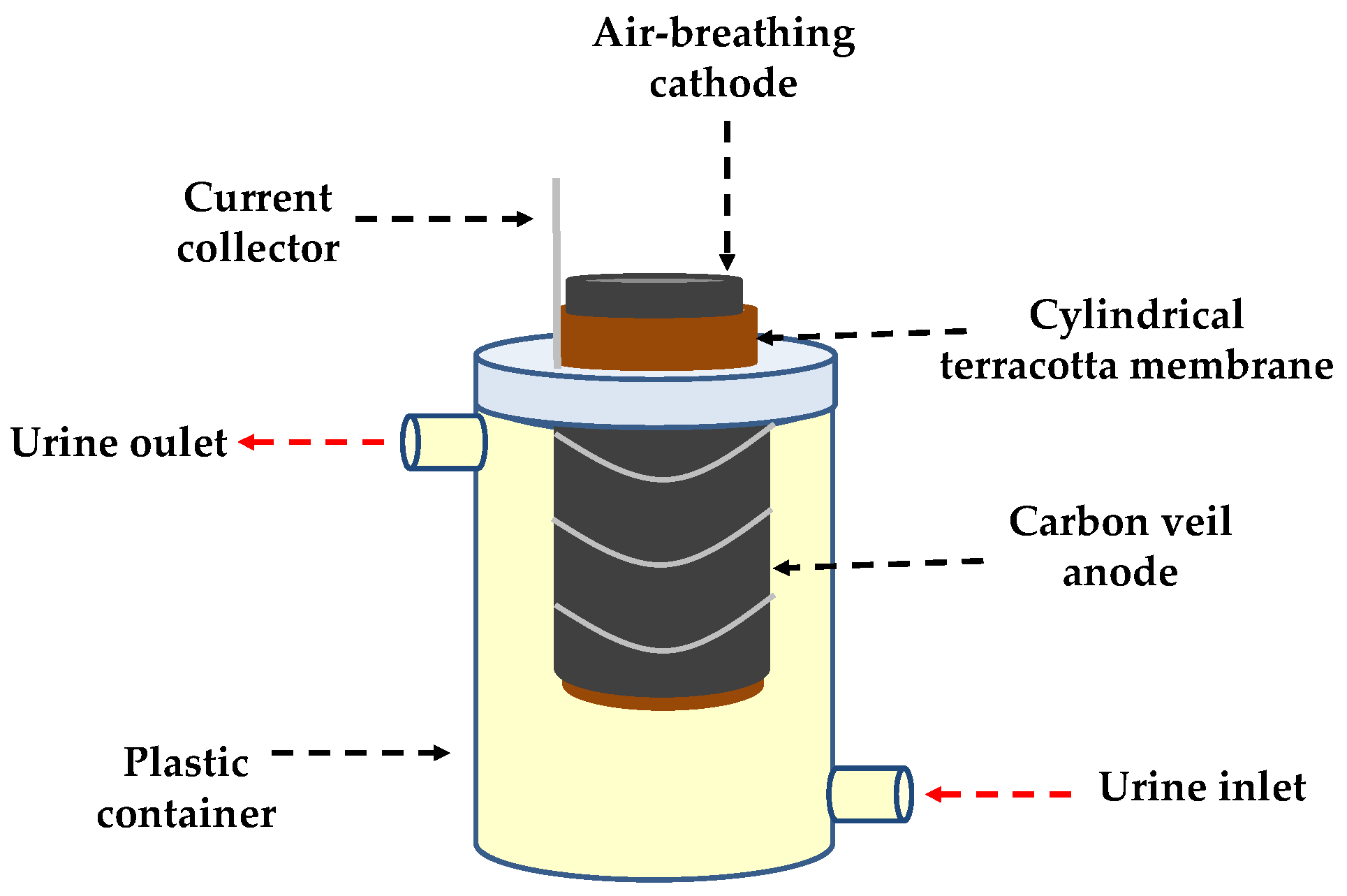
2.1. Microbial Fuel Cell (MFC) Set-Up
2.2. Inoculation Methods
- Method I: The MFCs were inoculated with naturally hydrolysed human urine. Before collection, urine was stored in a tank for at least 48 h, which allowed the hydrolase enzymes to precipitate struvite, increasing the pH to 9.3. The MFC were batch-fed daily for 4 days and then continuously fed with stored urine at a flow rate of 200 mL·day−1.
- Method II: In this case, the systems were inoculated with a mixture containing 1/1V/V anaerobic sludge collected from a Wessex Water treatment plant (Saltford, UK) and stored urine (pH: 9.28), which was replenished with fresh solution daily for 4 days. After this period, the MFCs were continuously fed with stored urine at a flow rate of 200 mL·day−1.
- Method III: The last inoculum consisted of using a solution containing 1/1V/V effluent from a long-term working MFC and stored urine (pH: 9.29). The solution was replenished daily with a fresh mixture for 4 days, after which period, the systems were continuously fed with stored urine at a flow rate of 200 mL·day−1.
2.3. Electrochemical Characterisation
2.4. DNA Isolation, Next-Generation 16S rRNA Amplicon Sequencing and Sequence Data Analysis
3. Results
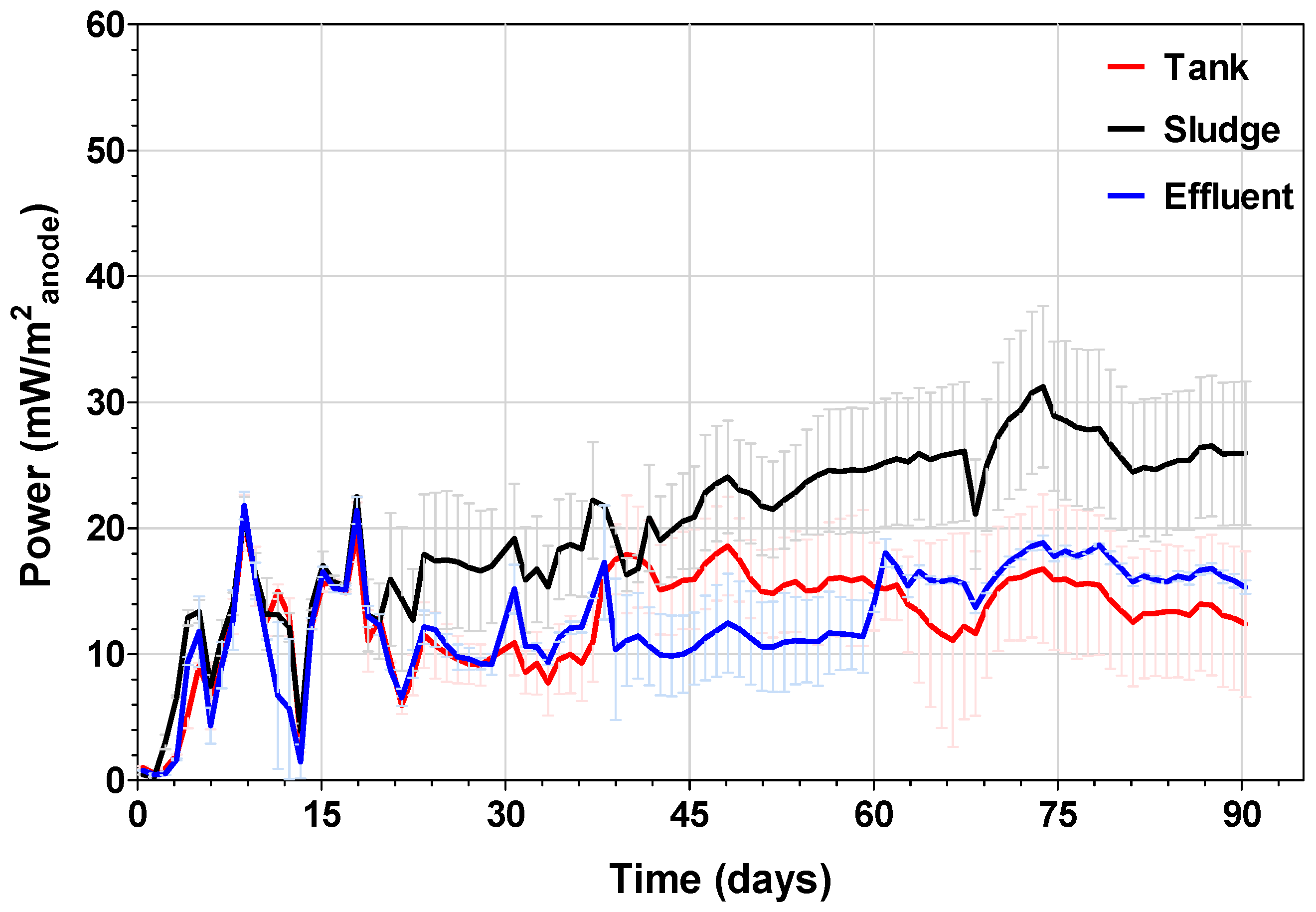
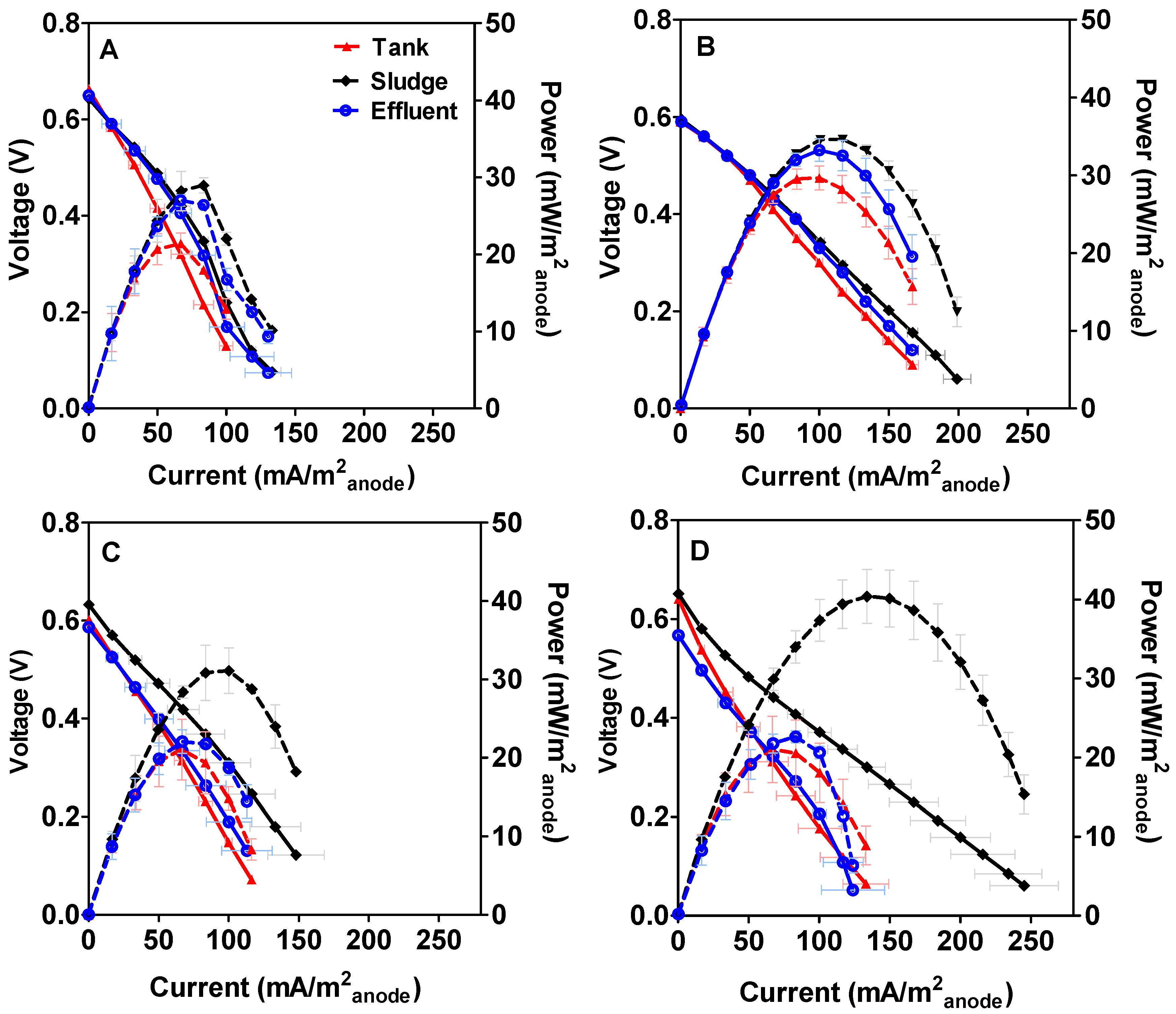
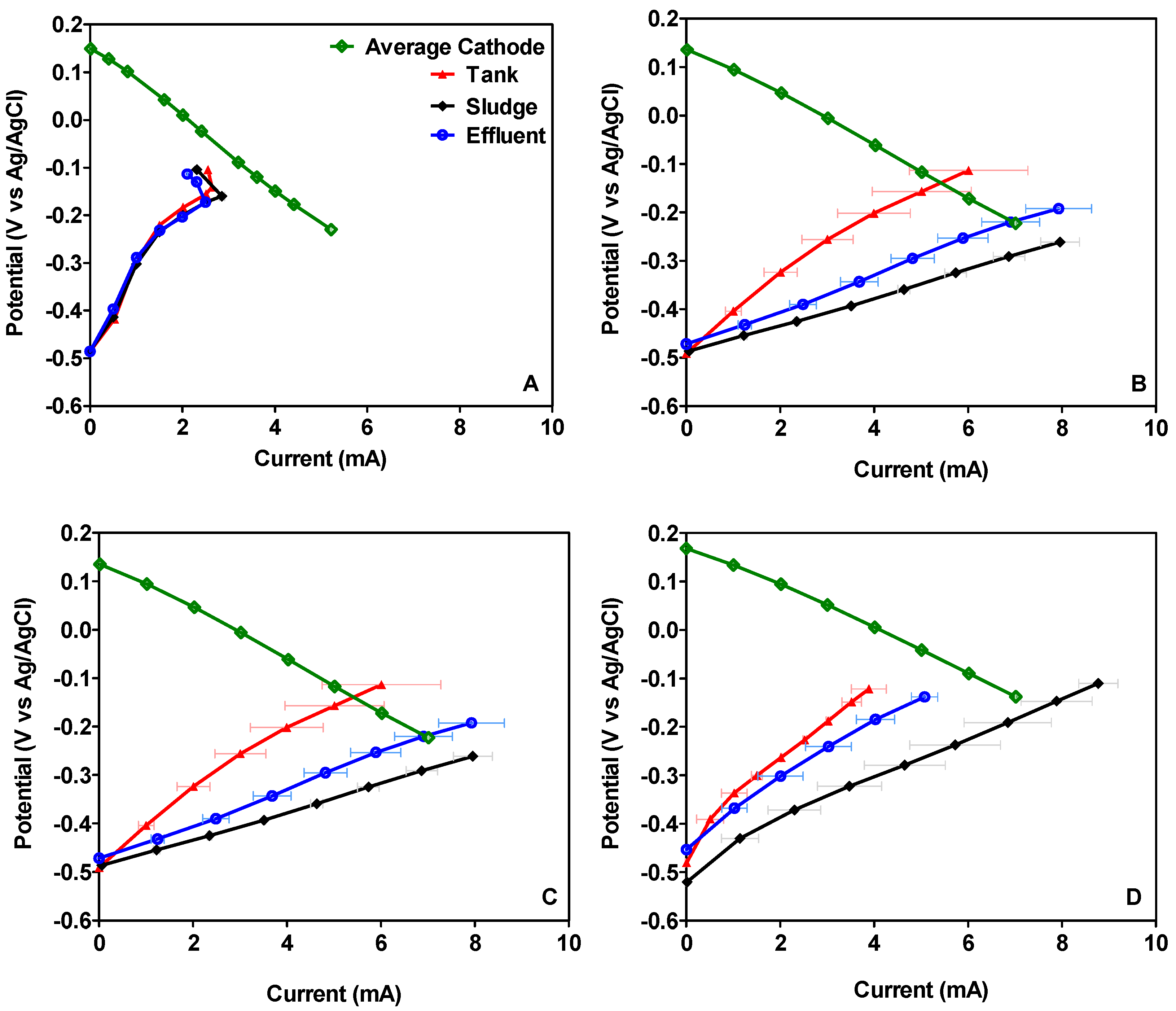
3.1. Microbial Fuel Cell Performance
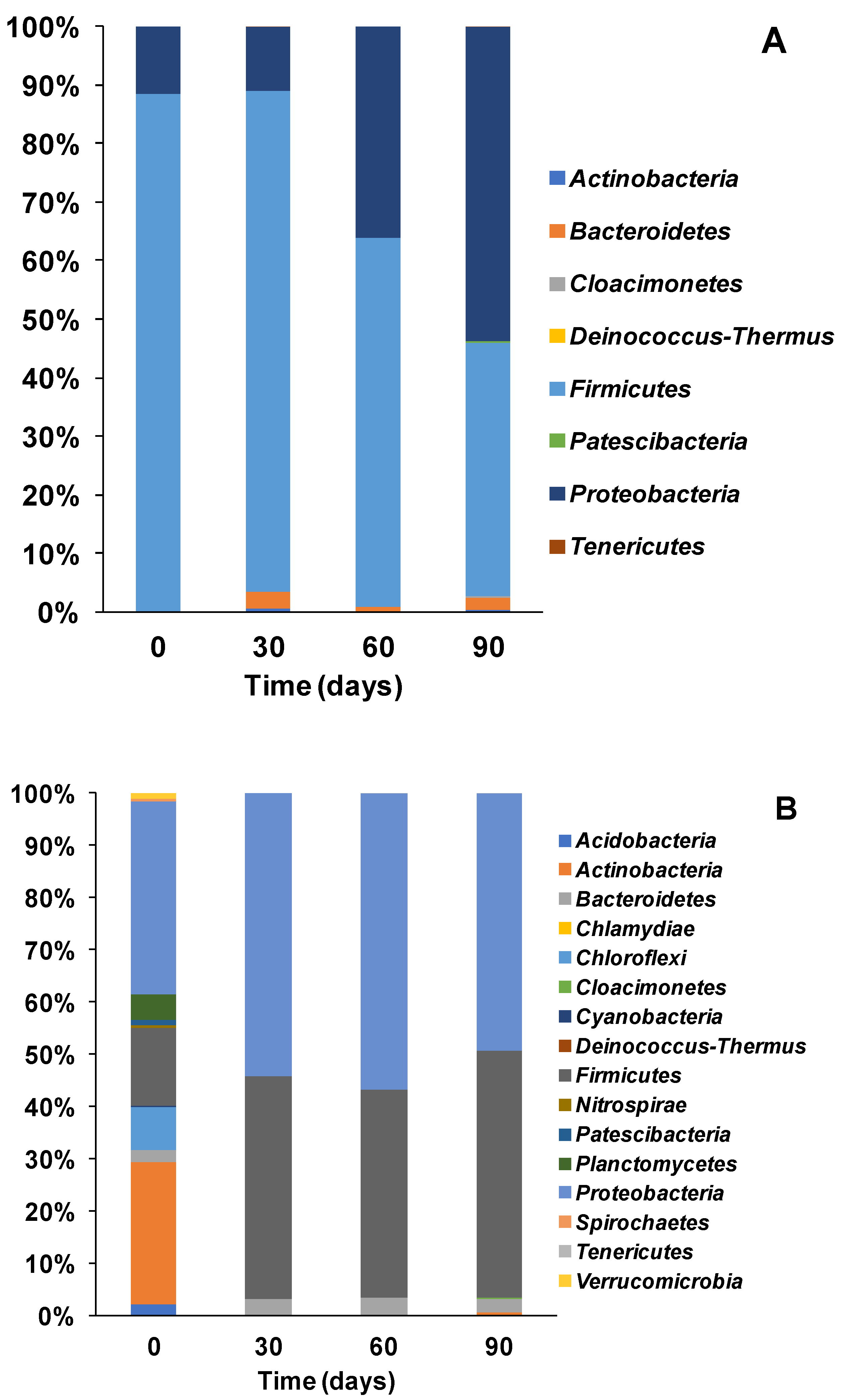
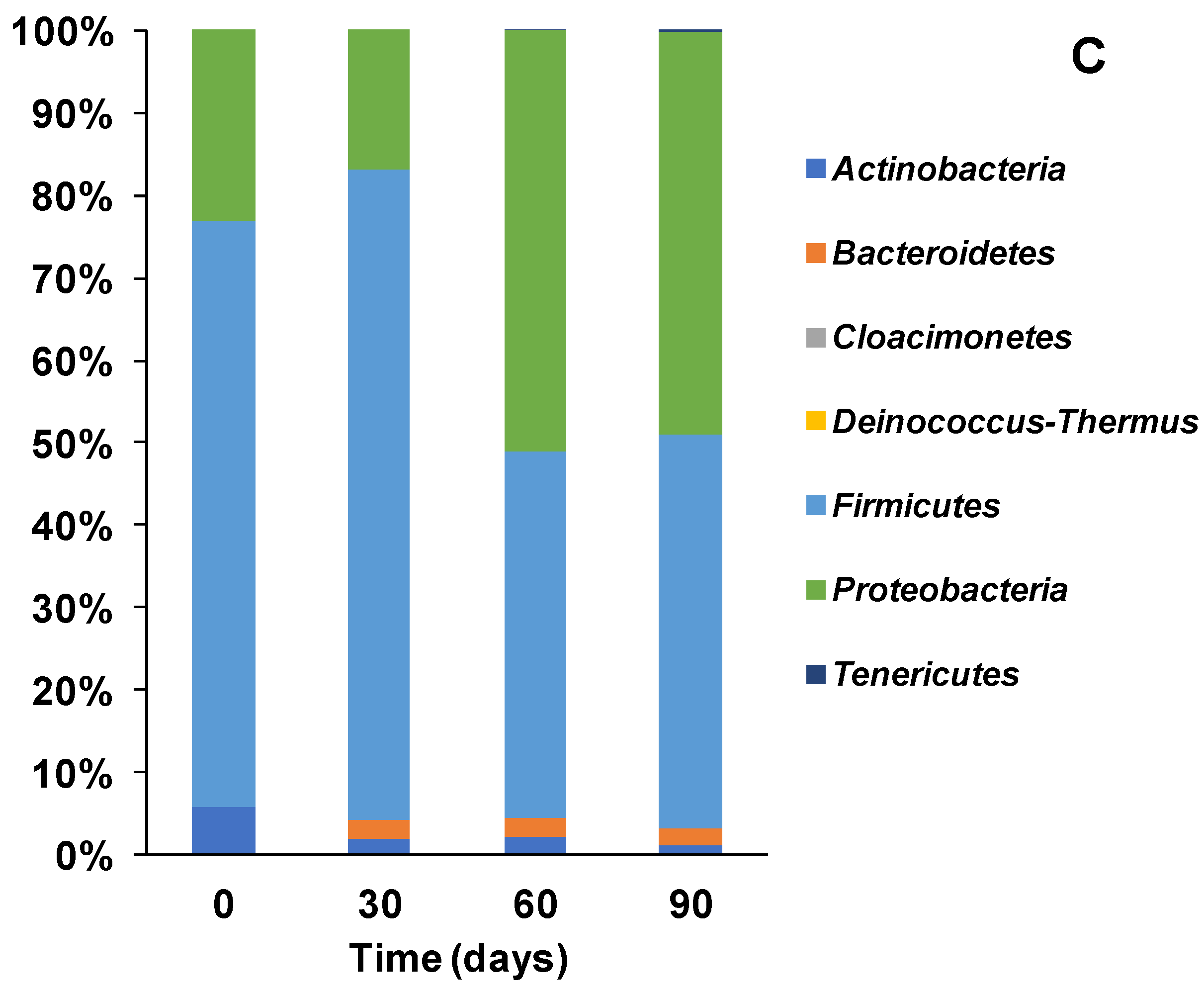
3.2. Microbial Ecology and Phyla Distribution
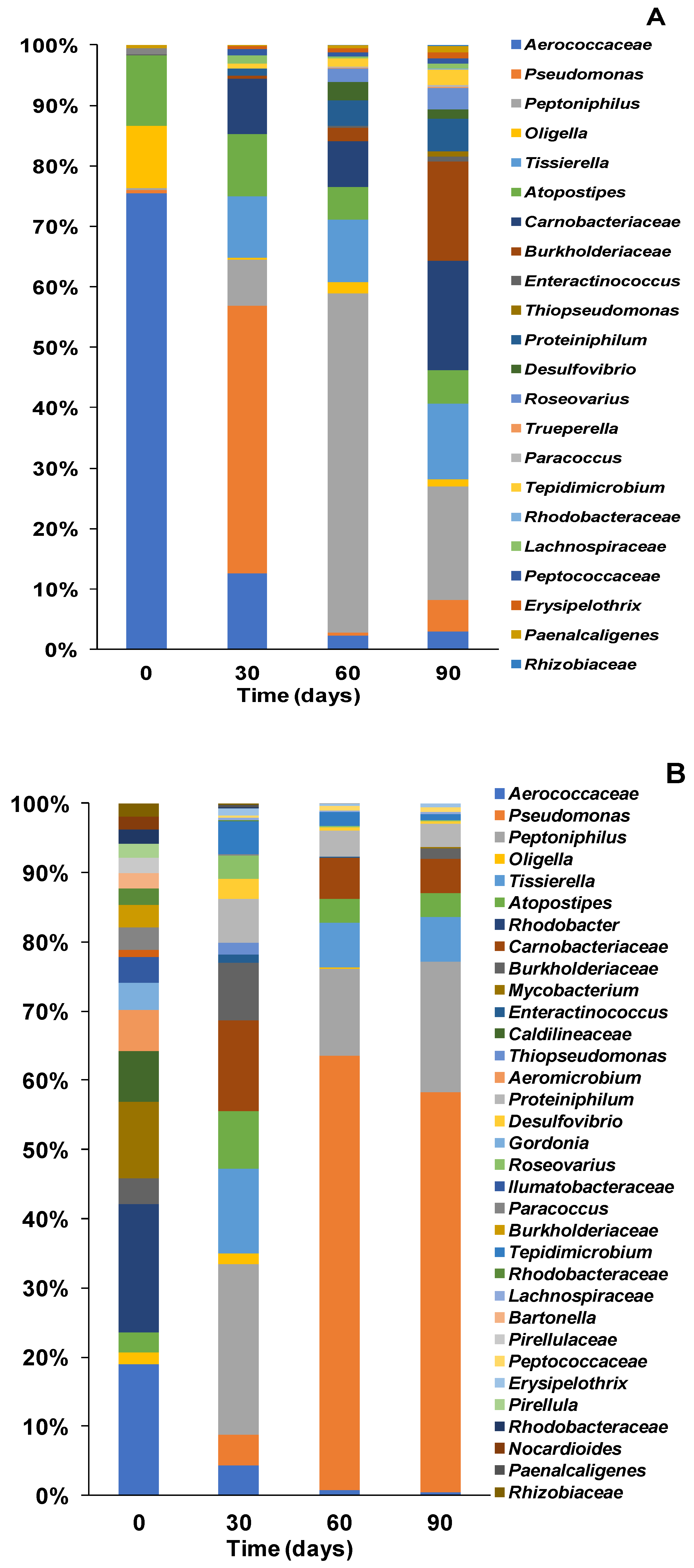
3.3. Bacterial Composition of Community at Genera Level with Different Inoculations
3.3.1. Bacterial Community Structure and Genus Distribution in Stored Urine Inoculated MFC Reactors
3.3.2. Bacteria Community Structure and Genus Distribution in Sludge Inoculated MFC Reactors
3.3.3. Bacteria Community Structure and Genus Distribution in Effluent Inoculated MFC Reactors
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stirling, J.L.; Bennetto, H.P.; Delaney, G.M.; Mason, J.R.; Roller, S.D.; Tanaka, K.; Thurston, C.F. Microbial fuel cells. Biochem. Soc. Trans. 1983, 11, 451–453. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.C. Electrical Effects Accompanying the Decomposition of Organic Compounds. Proc. R. Soc. B Biol. Sci. 1911, 84, 260–276. [Google Scholar] [CrossRef]
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial fuel cells: Methodology and technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Fernández, F.J.; Pérez De Los Ríos, A.; Salar-García, M.J.; Ortiz-Martínez, V.M.; Lozano-Blanco, L.J.; Godínez, C.; Tomás-Alonso, F.; Quesada-Medina, J. Recent progress and perspectives in microbial fuel cells for bioenergy generation and wastewater treatment. Fuel Process. Technol. 2015, 138, 284–297. [Google Scholar] [CrossRef]
- Yahia, S.A.A.; Hamadou, L.; Salar-García, M.J.; Kadri, A.; Ortiz-Martínez, V.M.; Hernández-Fernández, F.J.; de los Rios, A.P.; Benbrahim, N. TiO2 nanotubes as alternative cathode in microbial fuel cells: Effect of annealing treatment on its performance. Appl. Surf. Sci. 2016, 387, 1037–1045. [Google Scholar] [CrossRef]
- Touach, N.; Ortiz-Martínez, V.M.; Salar-García, M.J.; Benzaouak, A.; Hernández-Fernández, F.; de Ríos, A.P.; El Mahi, M.; Lotfi, E.M. On the use of ferroelectric material LiNbO3 as novel photocatalyst in wastewater-fed microbial fuel cells. Particuology 2017, 34, 147–155. [Google Scholar] [CrossRef]
- Haoyu, E.; Cheng, S.; Scott, K.; Logan, B. Microbial fuel cell performance with non-Pt cathode catalysts. J. Power Sources 2007, 171, 275–281. [Google Scholar] [CrossRef]
- Yuan, H.; Hou, Y.; Abu-Reesh, I.M.; Chen, J.; He, Z. Oxygen reduction reaction catalysts used in microbial fuel cells for energy-efficient wastewater treatment: A review. Mater. Horiz. 2016, 3, 382–401. [Google Scholar] [CrossRef] [Green Version]
- Guendouz, S.; Salar-García, M.J.; Ortiz-Martínez, V.M.; Zerrouki, A.; Houcine, I.; Ríos, A.P.; Hernández-Fernández, F.J. Electrocodeposition method to synthesize low-cost cathodes based on inert carriers for bioenergy production and wastewater treatment in microbial fuel cells. Environ. Prog. Sustain. Energy 2019, 38, 13083. [Google Scholar] [CrossRef]
- Yang, W.; He, W.; Zhang, F.; Hickner, M.A.; Logan, B.E. Single-Step Fabrication Using a Phase Inversion Method of Poly(vinylidene fluoride) (PVDF) Activated Carbon Air Cathodes for Microbial Fuel Cells. Environ. Sci. Technol. Lett. 2014. [Google Scholar] [CrossRef] [Green Version]
- Salar Garcia, M.J.; Santoro, C.; Kodali, M.; Serov, A.; Artyushkova, K.; Atanassov, P.; Ieropoulos, I. Iron-streptomycin derived catalyst for efficient oxygen reduction reaction in ceramic microbial fuel cells operating with urine. J. Power Sources 2019, 425, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Gajda, I.; You, J.; Santoro, C.; Greenman, J.; Ieropoulos, I.A. A new method for urine electrofiltration and long term power enhancement using surface modified anodes with activated carbon in ceramic microbial fuel cells. Electrochim. Acta 2020, 353, 136388. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wen, Q.; Chen, Y.; Zheng, H.; Wang, S. Enhanced performance of microbial fuel cell with polyaniline/sodium alginate/carbon brush hydrogel bioanode and removal of COD. Energy 2020, 202, 117780. [Google Scholar] [CrossRef]
- Wang, Y.; Wen, Q.; Chen, Y.; Li, W. Conductive polypyrrole-carboxymethyl cellulose-titanium nitride/carbon brush hydrogels as bioanodes for enhanced energy output in microbial fuel cells. Energy 2020, 204, 117942. [Google Scholar] [CrossRef]
- Salar-Garcia, M.J.; Montilla, F.; Quijada, C.; Morallon, E.; Ieropoulos, I. Improving the power performance of urine-fed microbial fuel cells using PEDOT-PSS modified anodes. Appl. Energy 2020, 278, 115528. [Google Scholar] [CrossRef]
- Li, W.-W.; Sheng, G.-P.; Liu, X.-W.; Yu, H.-Q. Recent advances in the separators for microbial fuel cells. Bioresour. Technol. 2011, 102, 244–252. [Google Scholar] [CrossRef]
- Yousefi, V.; Mohebbi-Kalhori, D.; Samimi, A. Ceramic-based microbial fuel cells (MFCs): A review. Int. J. Hydrog. Energy 2017, 42, 1672–1690. [Google Scholar] [CrossRef]
- Winfield, J.; Gajda, I.; Greenman, J.; Ieropoulos, I. A review into the use of ceramics in microbial fuel cells. Bioresour. Technol. 2016, 215, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Mačiulaitis, R.; Malaiškienė, J. Possibilities to control ceramics properties by changing firing cycles. Constr. Build. Mater. 2009, 23, 226–232. [Google Scholar] [CrossRef]
- Ghadge, A.N.; Ghangrekar, M.M. Development of low cost ceramic separator using mineral cation exchanger to enhance performance of microbial fuel cells. Electrochim. Acta 2015, 166, 320–328. [Google Scholar] [CrossRef]
- Salar-García, M.J.; Ieropoulos, I. Optimisation of the internal structure of ceramic membranes for electricity production in urine-fed microbial fuel cells. J. Power Sources 2020, 451, 227741. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Shinde, V.N.; Deopurkar, R.L.; Kale, S.P.; Patil, S.A.; Pant, D. Recent advances in the use of different substrates in microbial fuel cells toward wastewater treatment and simultaneous energy recovery. Appl. Energy 2016, 168, 706–723. [Google Scholar] [CrossRef]
- Zhang, E.; Wang, F.; Zhai, W.; Scott, K.; Wang, X.; Diao, G. Efficient removal of nitrobenzene and concomitant electricity production by single-chamber microbial fuel cells with activated carbon air-cathode. Bioresour. Technol. 2017, 229, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pant, D.; Van Bogaert, G.; Diels, L.; Vanbroekhoven, K. A review of the substrates used in microbial fuel cells (MFCs) for sustainable energy production. Bioresour. Technol. 2010, 101, 1533–1543. [Google Scholar] [CrossRef]
- Addi, H.; Mateo-Ramírez, F.; Ortiz-Martínez, V.M.; Salar-García, M.J.; Hernández-Fernández, F.J.; de los Ríos, A.P.; Godínez, C.; Lotfi, E.M.; El Mahi, M.; Blanco, L.J.L. Treatment of mineral oil refinery wastewater in microbial fuel cells using ionic liquid based separators. Appl. Sci. 2018, 8, 438. [Google Scholar] [CrossRef] [Green Version]
- Addi, H.; Mateo-Ramírez, F.; Salar-García, M.J.; Ortiz-Martínez, V.M.; Hernández-Fernández, F.J.; de los Ríos, A.P.; Godínez, C.; Lotfi, E.M.; El Mahi, M.; Lozano-Blanco, L.J. Ammonium-Based Polymer Ionic Liquid Membrane for Wastewater Treatment and Bioenergy Production. Chem. Eng. Technol. 2018, 41, 379–384. [Google Scholar] [CrossRef]
- Ieropoulos, I.; Greenman, J.; Melhuish, C. Urine utilisation by microbial fuel cells; Energy fuel for the future. Phys. Chem. Chem. Phys. 2012, 14, 94–98. [Google Scholar] [CrossRef]
- Ledezma, P.; Kuntke, P.; Buisman, C.J.N.; Keller, J.; Freguia, S. Source-separated urine opens golden opportunities for microbial electrochemical technologies. Trends Biotechnol. 2015, 33, 214–220. [Google Scholar] [CrossRef]
- Santoro, C.; Garcia, M.J.S.; Walter, X.A.; You, J.; Theodosiou, P.; Gajda, I.; Obata, O.; Winfield, J.; Greenman, J.; Ieropoulos, I. Urine in Bioelectrochemical Systems: An Overall Review. ChemElectroChem 2020, 7, 1312–1331. [Google Scholar] [CrossRef]
- Kuntke, P.; Śmiech, K.M.; Bruning, H.; Zeeman, G.; Saakes, M.; Sleutels, T.H.J.A.; Hamelers, H.V.M.; Buisman, C.J.N. Ammonium recovery and energy production from urine by a microbial fuel cell. Water Res. 2012. [Google Scholar] [CrossRef]
- You, J.; Staddon, C.; Cook, A.; Walker, J.; Boulton, J.; Powell, W.; Ieropoulos, I. Multidimensional Benefits of Improved Sanitation: Evaluating ‘PEE POWER®’ in Kisoro, Uganda. Int. J. Environ. Res. Public Health 2020, 17, 2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, V.B.; Simões, M.; Melo, L.F.; Pinto, A.M.F.R. Overview on the developments of microbial fuel cells. Biochem. Eng. J. 2013, 73, 53–64. [Google Scholar] [CrossRef]
- Heidrich, E.S.; Dolfing, J.; Wade, M.J.; Sloan, W.T.; Quince, C.; Curtis, T.P. Temperature, inocula and substrate: Contrasting electroactive consortia, diversity and performance in microbial fuel cells. Bioelectrochemistry 2018, 119, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Rabaey, K.; Rozendal, R.A. Microbial electrosynthesis—Revisiting the electrical route for microbial production. Nat. Rev. Microbiol. 2010, 8, 706–716. [Google Scholar] [CrossRef]
- Zhang, Y.; Min, B.; Huang, L.; Angelidaki, I. Electricity generation and microbial community response to substrate changes in microbial fuel cell. Bioresour. Technol. 2011. [Google Scholar] [CrossRef]
- Obata, O.; Salar-Garcia, M.J.; Greenman, J.; Kurt, H.; Chandran, K.; Ieropoulos, I. Development of efficient electroactive biofilm in urine-fed microbial fuel cell cascades for bioelectricity generation. J. Environ. Manag. 2020, 258, 109992. [Google Scholar] [CrossRef]
- Rismani-Yazdi, H.; Christy, A.D.; Carver, S.M.; Yu, Z.; Dehority, B.A.; Tuovinen, O.H. Effect of external resistance on bacterial diversity and metabolism in cellulose-fed microbial fuel cells. Bioresour. Technol. 2011. [Google Scholar] [CrossRef]
- Sun, Y.; Wei, J.; Liang, P.; Huang, X. Electricity generation and microbial community changes in microbial fuel cells packed with different anodic materials. Bioresour. Technol. 2011. [Google Scholar] [CrossRef]
- Gajda, I.; Greenman, J.; Melhuish, C.; Ieropoulos, I. Simultaneous electricity generation and microbially-assisted electrosynthesis in ceramic MFCs. Bioelectrochemistry 2015, 104, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Ferris, M.J.; Muyzer, G. Denaturing Gradient Gel Electrophoresis Profiles of 16S rRNA-Defined Populations Inhabiting a Hot Spring Microbial Mat Community. Appl. Environ. Microbiol. 1996, 62, 340–346. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Park, H.; Chandran, K. Molecular and Kinetic Characterization of Planktonic Nitrospira spp. Selectively Enriched from Activated Sludge. Environ. Sci. Technol. 2017, 51, 2720–2728. [Google Scholar] [CrossRef] [PubMed]
- Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Gabriel, A.; Ghalith, A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. QIIME 2: Reproducible, interactive, scalable, and extensible microbiome data science. PeerJ Prepr. 2018, 6, 1–53. [Google Scholar]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Glo, F.O.; Yarza, P. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Tursun, H.; Hou, X.; Odey, F.; Li, Y.; Wang, X.; Xie, T. Microbial community dynamics in a pilot-scale MFC-AA/O system treating domestic sewage. Bioresour. Technol. 2017, 241, 439–447. [Google Scholar] [CrossRef]
- Mei, X.; Xing, D.; Yang, Y.; Liu, Q.; Zhou, H.; Guo, C.; Ren, N. Adaptation of microbial community of the anode biofilm in microbial fuel cells to temperature. Bioelectrochemistry 2017, 117, 29–33. [Google Scholar] [CrossRef]
- Jung, S.; Regan, J.M. Comparison of anode bacterial communities and performance in microbial fuel cells with different electron donors. Appl. Microbiol. Biotechnol. 2007, 77, 393–402. [Google Scholar] [CrossRef]
- Park, Y.; Cho, H.; Yu, J.; Min, B.; Kim, H.S.; Kim, B.G.; Lee, T. Response of microbial community structure to pre-acclimation strategies in microbial fuel cells for domestic wastewater treatment. Bioresour. Technol. 2017, 233, 176–183. [Google Scholar] [CrossRef]
- Cid, C.A.; Stinchcombe, A.; Ieropoulos, I.; Ho, M.R. Urine microbial fuel cells in a semi-controlled environment for onsite urine pre-treatment and electricity production. J. Power Sources 2018, 400, 441–448. [Google Scholar] [CrossRef]
- Chae, K.J.; Choi, M.J.; Lee, J.W.; Kim, K.Y.; Kim, I.S. Effect of different substrates on the performance, bacterial diversity, and bacterial viability in microbial fuel cells. Bioresour. Technol. 2009, 100, 3518–3525. [Google Scholar] [CrossRef]
- Suzuki, K.; Kato, Y.; Yui, A.; Yamamoto, S.; Ando, S.; Rubaba, O.; Tashiro, Y.; Futamata, H. Bacterial communities adapted to higher external resistance can reduce the onset potential of anode in microbial fuel cells. J. Biosci. Bioeng. 2018, 125, 565–571. [Google Scholar] [CrossRef]
- Paitier, A.; Godain, A.; Lyon, D.; Haddour, N.; Vogel, T.M.; Monier, J.M. Microbial fuel cell anodic microbial population dynamics during MFC start-up. Biosens. Bioelectron. 2017, 92, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, G.; Greenman, J.; Ieropoulos, I. Comprehensive Study on Ceramic Membranes for Low-Cost Microbial Fuel Cells. ChemSusChem 2016, 9, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Higgins, A.; Garg, T. Inflammation and Infection Aerococcus urinae: An Emerging Cause of Urinary Tract Infection in Older Adults with Multimorbidity and Urologic Cancer. Urol. Case Rep. 2017, 13, 24–25. [Google Scholar] [CrossRef] [PubMed]
- Rabaey, K.; Verstraete, W. Microbial fuel cells: Novel biotechnology for energy generation. Trends Biotechnol. 2005, 23, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Sydow, A.; Krieg, T.; Mayer, F.; Schrader, J.; Holtmann, D. Electroactive bacteria—Molecular mechanisms and genetic tools. Appl. Microbiol. Biotechnol. 2014, 98, 8481–8495. [Google Scholar] [CrossRef]
- Harrns, C.; Schleicher, A.; Collins, M.D.; Andreesen, J.R. Tissierella creatinophila sp. nov., a Gram- positive, anaerobic, non-spore-f orming, creatinine-f errnent ing organism. Int. J. Syst. Bacteriol. 1998, 48, 983–993. [Google Scholar] [CrossRef] [Green Version]
- Rabaey, K.; Rodríguez, J.; Blackall, L.L.; Keller, J.; Gross, P.; Batstone, D.; Verstraete, W.; Nealson, K.H. Microbial ecology meets electrochemistry: Electricity-driven and driving communities. ISME J. 2007, 1, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, P.; Uday, U.S.P.; Mahata, N.; Nath Tiwari, O.; Narayan Ray, R.; Kanti Bandyopadhyay, T.; Bhunia, B. Performance improvement of microbial fuel cells for waste water treatment along with value addition: A review on past achievements and recent perspectives. Renew. Sustain. Energy Rev. 2017, 79, 372–389. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
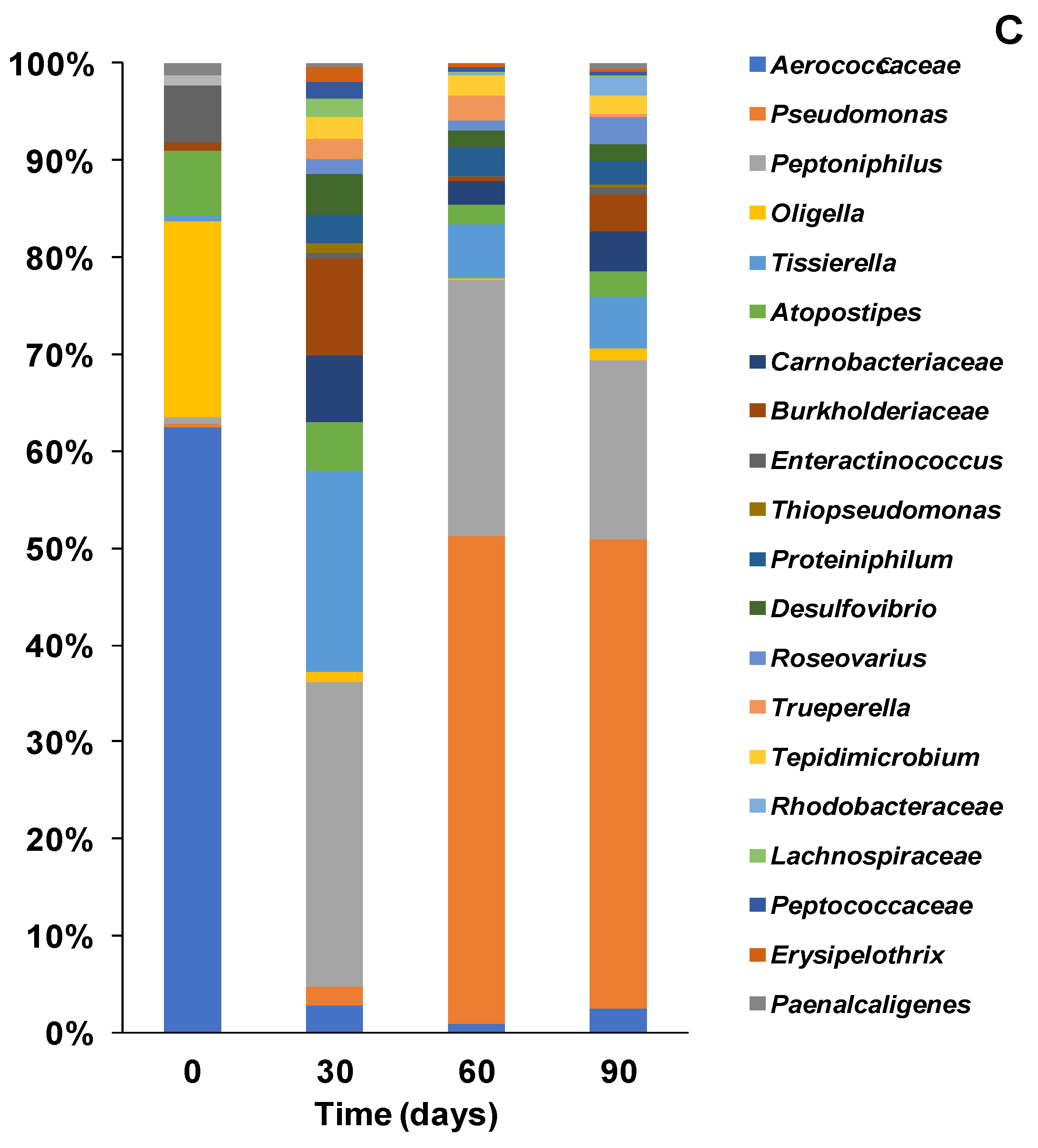{kind=link}
| Maximum Power Output (mW/m2anode) | |||
|---|---|---|---|
| Time (Days) | Stored Urine | Sludge | Effluent |
| 15 | 21.40 ± 2.5 | 28.95 ± 2.8 | 27.00 ± 3.5 |
| 30 | 29.64 ± 2.3 | 34.63 ± 1.9 | 33.25 ± 2.8 |
| 60 | 21.04 ± 3.9 | 31.11 ± 3.5 | 22.06 ± 2.1 |
| 90 | 21.01 ± 4.2 | 40.38 ± 3.7 | 22.61 ± 2.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salar-Garcia, M.J.; Obata, O.; Kurt, H.; Chandran, K.; Greenman, J.; Ieropoulos, I.A. Impact of Inoculum Type on the Microbial Community and Power Performance of Urine-Fed Microbial Fuel Cells. Microorganisms 2020, 8, 1921. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121921
Salar-Garcia MJ, Obata O, Kurt H, Chandran K, Greenman J, Ieropoulos IA. Impact of Inoculum Type on the Microbial Community and Power Performance of Urine-Fed Microbial Fuel Cells. Microorganisms. 2020; 8(12):1921. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121921
Chicago/Turabian StyleSalar-Garcia, Maria Jose, Oluwatosin Obata, Halil Kurt, Kartik Chandran, John Greenman, and Ioannis A. Ieropoulos. 2020. "Impact of Inoculum Type on the Microbial Community and Power Performance of Urine-Fed Microbial Fuel Cells" Microorganisms 8, no. 12: 1921. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121921