Effects of Ectomycorrhizal Fungi and Heavy Metals (Pb, Zn, and Cd) on Growth and Mineral Nutrition of Pinus halepensis Seedlings in North Africa

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Growth Conditions
2.2. Sampling and Physico-Chemical Analyses of Contaminated and Control Soils
2.3. Sampling, Dry Mass Measurements and Plant Tissue Analysis of Ectomycorrhizal Pinus halepensis Seedlings
2.4. Statistical Analyses
3. Results
3.1. Soil Physicochemical Properties
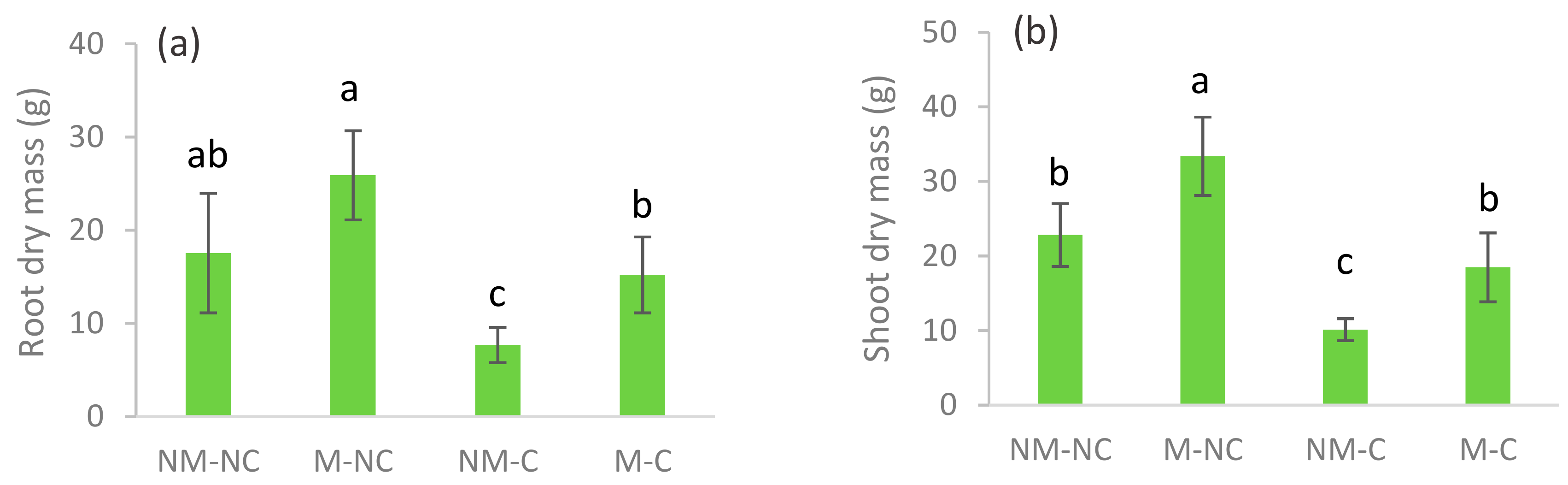
3.2. Growth and Seedling Morphology
3.3. Mineral Nutrient Contents
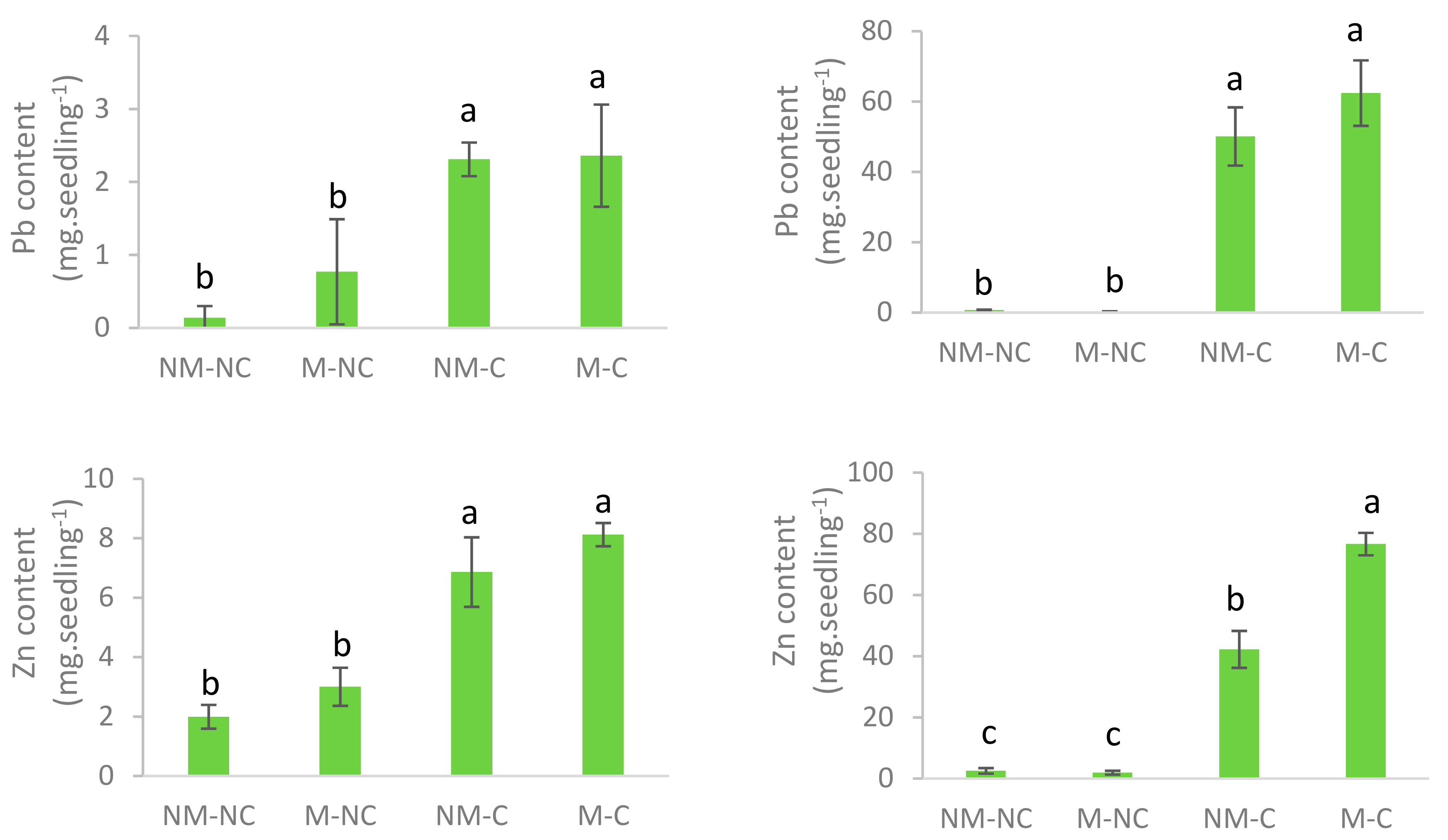
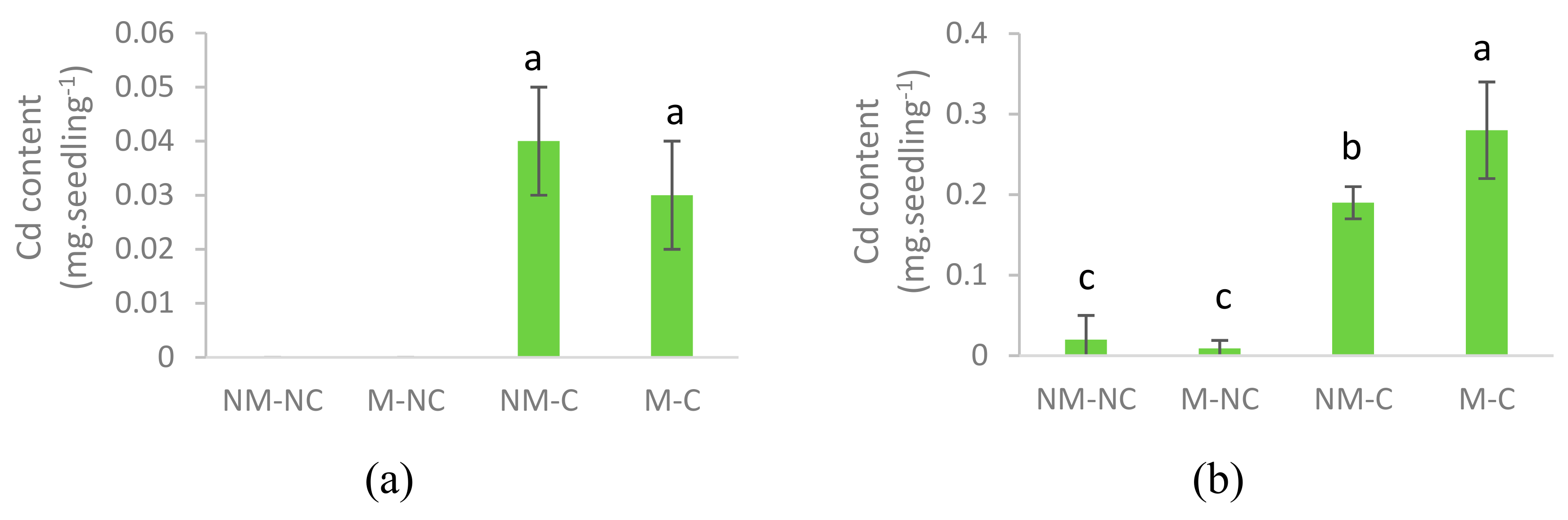
3.4. Heavy Metal Content
4. Discussion
5. Conclusion and Research Needs
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yabe, J.; Ishizuka, M.; Umemura, T. Current levels of heavy metal pollution in Africa. J. Vet. Med. Sci. 2010, 72, 1257–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO; ITPS. Status of the World’s Soil Resources (SWSR)—Main Report; Food and Agriculture Organization of the United Nations and Intergovernmental Technical Panel on Soils: Rome, Italy, 2015; p. 650. [Google Scholar]
- Strzebonska, M.; Jarosz-Krzemińska, E.; Adamiec, E. Assessing historical mining and smelting effects on heavy metal pollution of river systems over span of two decades. Water Air Soil Poll. 2017, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaca, O.; Cameselle, C.; Reddy, K.R. Mine tailing disposal sites: Contamination problems, remedial options and phytocaps for sustainable remediation. Rev. Environ. Sci. Biotechnol. 2018, 17, 205–228. [Google Scholar] [CrossRef]
- Ogundele, L.T.; Owoade, O.K.; Hopke, P.K.; Olise, F.S. Heavy metals in industrially emitted particulate matter in Ile-Ife, Nigeria. Environ. Res. 2017, 156, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Doumas, P.; Munoz, M.; Banni, M.; Becerra, S.; Bruneel, O.; Casiot, C.; Cleyet-Marel, J.C.; Gardon, J.; Noack, Y.; Sappin-Didier, V. Polymetallic pollution from abandoned mines in Mediterranean regions: A multidisciplinary approach to environmental risks. Reg. Environ. Chang. 2016, 18, 677–692. [Google Scholar] [CrossRef]
- Ghorbel, M.; Munoz, M.; Courjault-Rade, P.; Destrigneville, C.; Souissi, R.; Souissi, F.; Ben Mammou, A.; Abdeljaouad, S. Health risk assessment for human exposure by direct ingestion of Pb, Cd, Zn bearing dust in the former miner’s village of Jebel Ressas (NE Tunisia). Eur. J. Mineral. 2010, 22, 639–649. [Google Scholar] [CrossRef]
- El Khalil, H.; El Hamiani, O.; Bitton, G.; Ouazzani, N.; Boularbah, A. Heavy metal contamination from mining sites in South Morocco: Monitoring metal content and toxicity of soil runoff and groundwater. Environ. Monit. Assess. 2008, 136, 147–160. [Google Scholar] [CrossRef]
- Elouear, Z.; Bouhamed, F.; Boujelben, N.; Bouzid, J. Assessment of toxic metals dispersed from improperly disposed tailing, Jebel Ressas mine, NE Tunisia. Environ. Earth Sci. 2016, 75, 1–7. [Google Scholar] [CrossRef]
- Hudson-Edwards, K.A.; Jamieson, H.E.; Lottermoser, B.G. Minewastes: Past, present, future. Elements 2011, 7, 375–380. [Google Scholar] [CrossRef]
- Othmani, M.A.; Souissi, F.; Benzaazoua, M.; Bouzahzah, H.; Bussiere, B.; Mansouri, A. The geochemical behavior of mine tailings from the Touiref Pb–Zn district in Tunisia in weathering cells leaching tests. Mine Water Environ. 2013, 32, 28–41. [Google Scholar] [CrossRef]
- FAO. Soil Pollution a Hidden Reality; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; p. 142. [Google Scholar]
- Ghorbel, M. Contamination Métallique Issue des Déchets de L’ancien site Minier de Jebel Ressas: Modélisation des Mécanismes de Transfert et Conception de Cartes D’aléa Post-Mine Dans un Contexte Carbonaté et Sous un Climat Semi-Aride. Evaluation du Risque Pour la Santé Humaine. Ph.D. Thesis, Paul Sabatier-Toulouse III University, Toulouse, France, 2012. [Google Scholar]
- Gupta, G.; Khan, J.; Singh, N.K. Phytoremediation of Metal-Contaminated Sites. In Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives II; Hasanuzzaman, M., Ed.; Springer Nature: Singapore, 2020; pp. 725–745. [Google Scholar] [CrossRef]
- Otero-Blanca, A.; Folch-Mallol, J.L.; Lira-Ruan, V.; del Rayo Sánchez Carbente, M.; Batista-García, R.A. Phytoremediation and fungi: An underexplored binomial. In Approaches in Bioremediation: The New Era of Environmental Microbiology and Nanobiotechnology; Prasad, R., Aranda, E., Eds.; Springer: Cham, Switzerland, 2018; pp. 79–95. [Google Scholar]
- Marx, D.H. Mycorrhizae and establishment of trees on strip-mined land. Ohio J. Sci. 1975, 75, 288–297. [Google Scholar]
- Grossnickle, S.C. Ectomycorrhizae: A viable alternative for successful mined land reclamation. In Proceedings of the America Society of Mining and Reclamation, Princeton, WV, USA, 15 August 1985; pp. 306–313. [Google Scholar] [CrossRef]
- Urban, A.; Puschenreiter, M.; Strauss, J.; Gorfer, M. Diversity and structure of ectomycorrhizal and co-associated fungal communities in a serpentine soil. Mycorrhiza 2008, 18, 339–354. [Google Scholar] [CrossRef]
- Gadd, G.M. Interactions of fungi with toxic metals. New Phytol. 1993, 124, 25–60. [Google Scholar] [CrossRef]
- Jentschke, G.; Godbold, D.L. Metal toxicity and ectomycorrhizas. Physiol. Plant 2000, 109, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Colpaert, J.V.; Wevers, J.H.L.; Krznaric, E.; Adriaensen, K. How metal-tolerant ecotypes of ectomycorrhizal fungi protect plants from heavy metal pollution. Ann. For. Sci. 2011, 68, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Blaudez, D.; Jacob, C.; Turnau, K.; Colpaert, J.V.; Ahonen-Jonnarth, U.; Finlay, R.; Botton, B.; Chalot, M. Differential responses of ectomycorrhizal fungi to heavy metals in vitro. Mycol. Res. 2000, 104, 1366–1371. [Google Scholar] [CrossRef]
- Bojarczuk, K.; Karliński, L.; Hazubska-Przybył, T.; Kieliszewska-Rokicka, B. Influence of mycorrhizal inoculation on growth of micropropagated Populus × canescens lines in metal-contaminated soils. New For. 2015, 46, 195–215. [Google Scholar] [CrossRef] [Green Version]
- Siddiquee, S.; Rovina, K.; Al, A.S.; Naher, L.; Suryani, S.; Chaikaew, P. Microbial & biochemical technology heavy metal contaminants removal from wastewater using the potential filamentous fungi biomass: A review. Microbiol. Biochem. Technol. 2015, 7, 384–393. [Google Scholar]
- Lamhamedi, M.S.; Fortin, J.A.; Bernier, P.Y. La génétique de Pisolithus sp.: Une nouvelle approche de biotechnologie forestière pour assurer une meilleure survie des plants en conditions de sécheresse. Sécheresse 1991, 2, 251–258. [Google Scholar]
- Lamhamedi, M.S.; Bernier, P.Y.; Fortin, J.A. Hydraulic conductance and soil water potential at the soil-root interface of Pinus pinaster seedlings inoculated with different dikaryons of Pisolithus sp. Tree Physiol. 1992, 10, 231–244. [Google Scholar] [CrossRef]
- Luo, Z.B.; Li, K.; Jiang, X.; Polle, A. Ectomycorrhizal fungus (Paxillus involutus) and hydrogels affect performance of Populus euphratica exposed to drought stress. Ann. For. Sci. 2009, 66, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Beniwal, R.S.; Langenfeld-Heyser, R.; Polle, A. Ectomycorrhiza and hydrogel protect hybrid poplar from water deficit and unravel plastic responses of xylem anatomy. Environ. Exp. Bot. 2010, 69, 189–197. [Google Scholar] [CrossRef]
- Rajtor, M.; Piotrowska-Seget, Z. Prospects for arbuscular mycorrhizal fungi (AMF) to assist in phytoremediation of soil hydrocarbon contaminants. Chemosphere 2016, 162, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Garg, N. Recent perspectives on cross talk between cadmium, zinc, and arbuscular mycorrhizal fungi in plants. J. Plant Growth Regul. 2018, 37, 680–693. [Google Scholar] [CrossRef]
- Lamhamedi, M.S.; Fortin, J.A.; Kope, H.H.; Kropp, B.R. Genetic variation in ectomycorrhiza formation by Pisolithus arhizus on Pinus pinaster and Pinus banksiana. New Phytol. 1990, 115, 689–697. [Google Scholar] [CrossRef]
- Lamhamedi, M.S.; Ammari, Y.; Fecteau, B.; Fortin, J.A.; Margolis, H. Problèmatique des pépinères forestières en Afrique du Nord et strategies de développement. Cahiers Agric. 2000, 9, 369–380. [Google Scholar]
- Lamhamedi, M.S.; Abourouh, M.; Fortin, J.A. Technological transfer: The use of ectomycorrhizal fungi in conventional and modern forest tree nurseries in northern Africa. In Advances in Mycorrhizal Science and Technology; Khasa, D., Piché, Y., Coughlan, A.P., Eds.; NRC Research Press: Ottawa, ON, Canada, 2009; pp. 139–152. [Google Scholar]
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Ne’eman, G.; Trabaud, L. Ecology, Biogeography and Management of Pinus Halepensis and P. Brutia Forest Ecosystems in the Mediterranean Basin; Backhuys Publishers: Leiden, The Netherlands, 2000; p. 407. ISBN 90-5782-055-2. [Google Scholar]
- Fuentes, D.; Disante, K.B.; Valdecantos, A.; Cortina, J.; Ramón-Vallejo, V. Response of Pinus halepensis Mill. seedlings to biosolids enriched with Cu, Ni and Zn in three Mediterranean forest soils. Environ. Pollut. 2007, 145, 316–323. [Google Scholar] [CrossRef]
- Querejeta, J.I.; Barberá, G.G.; Granados, A.; Castillo, V.M. Afforestation method affects the isotopic composition of planted Pinus halepensis in a semiarid region of Spain. For. Ecol. Manag. 2008, 254, 56–64. [Google Scholar] [CrossRef]
- Scarascia-Mugnozza, G.T. Recherches sur l’écophysiologie de Pinus halepensis Mill. In Le pin d’Alep et le Pin Brutia Dans la Sylviculture Méditerranéenne; Options Méditerranéennes; Paris-CIHEAM: Série Etudes, France, 1986; pp. 89–97. [Google Scholar]
- Barbéro, M.; Loisel, R.; Quézel, P.; Richardson, D.M.; Romane, F. Pines of the Mediterranean basin. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 153–170. [Google Scholar]
- Parraga-Aguado, I.; Álvarez-Rogel, J.; González-Alcaraz, M.N.; Jiménez-Cárceles, F.J.; Conesa, H.M. Assessment of metal(loid)s availability and their uptake by Pinus halepensis in a Mediterranean forest impacted by abandoned tailings. Ecol. Eng. 2013, 58, 84–90. [Google Scholar] [CrossRef]
- Parraga-Aguado, I.; Querejeta, J.I.; González-Alcaraz, M.N.; Conesa, H.M. Metal(loid) allocation and nutrient retranslocation in Pinus halepensis trees growing on semiarid mine tailings. Sci. Total Environ. 2014, 485, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J.; Rodá, F. Changes in nutrient use efficiency, status and retranslocation in young post-fire regeneration Pinus halepensis in response to sudden N and P input, irrigation and removal of competing vegetation. Trees 2005, 19, 233–250. [Google Scholar] [CrossRef]
- Jagtap, U.B.; Bapat, V.A. Genetic engineering of plants for heavy metal removal from soil. In Heavy Metal Contamination of Soils; Sherameti, I., Varma, A., Eds.; Springer: Cham, Switzerland, 2015; pp. 433–470. [Google Scholar] [CrossRef]
- FAO; Bleu, P.; Mediterranea, S. FFEM: État des Forêts Méditerranéennes; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; p. 207. [Google Scholar]
- McDowell, N.G.; Allen, C.D. Darcy’s law predicts widespread forest mortality under climate warming. Nat. Clim. Chang. 2015, 5, 669–672. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.; Llusià, J.; Ogaya, R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O.; et al. Impacts of global change on Mediterranean forests and their services. Forests 2017, 8, 463. [Google Scholar] [CrossRef] [Green Version]
- Lima, A.T.; Mitchell, K.; O’Connell, D.W.; Verhoeven, J.; Van Cappellen, P. The legacy of surface mining: Remediation, restoration, reclamation and rehabilitation. Environ. Sci. Policy 2016, 66, 227–233. [Google Scholar] [CrossRef]
- Rayment, G.E.; Lyons, D.J. Soil Chemical Methods: Australasia, 3rd ed.; CSIRO Publishing: Melbourne, Australia, 2011. [Google Scholar]
- Cameselle, C.; Gouveia, S. Phytoremediation of mixed contaminated soil enhanced with electric current. J. Hazard. Mater. 2019, 361, 95–102. [Google Scholar] [CrossRef]
- USEPA. US Environmental Protection Agency (USEPA) Method 3050B—Acid Digestion of Sediments, Sludges, and Soils. 1996. Available online: https://www.epa.gov/sites/production/files/2015-06/documents/epa-3050b.pdf (accessed on 5 May 2019).
- Lamhamedi, M.S.; Labbe, L.; Margolis, H.A.; Stowe, D.C.; Blais, L.; Renaud, M. Spatial variability of substrate water content and growth of white spruce seedlings. Soil Sci. Soc. Am. J. 2006, 70, 108–120. [Google Scholar] [CrossRef] [Green Version]
- Lamhamedi, M.S.; Renaud, M.; Auger, I.; Fortin, J.A. Granular calcite stimulates natural mycorrhization and growth of white spruce seedlings in peat-based substrates in forest nursery. Microorganisms 2020, 8, 1088. [Google Scholar] [CrossRef]
- Agerer, R. Colour Atlas of Ectomycorrhizae, 1st ed.; Einhorn—Verlag Eduard: Munich, Germany, 1998; ISBN 3-921703-77-8. [Google Scholar]
- Timmer, V.R.; Miller, B.D. Effects of contrasting fertilization and moisture regimes on biomass, nutrients, and water relations of container grown red pine seedlings. New Forest. 1991, 5, 335–348. [Google Scholar] [CrossRef]
- Lamhamedi, M.; Renaud, M.; Desjardins, P.; Veilleux, L. Root growth, plug cohesion, mineral nutrition, and carbohydrate content of 1+0 Picea mariana seedlings in response to a short-day treatment. Tree Planters Note 2013, 56, 35–46. [Google Scholar]
- Mackay, D.; Fraser, A. Bioaccumulation of persistent organic chemicals: Mechanisms and models. Environ. Pollut. 2000, 110, 375–391. [Google Scholar] [CrossRef] [PubMed]
- Steel, G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics: A Biometrical Approach, 3rd ed.; The McGraw-Hill Companies Inc.: New York, NY, USA, 1997. [Google Scholar]
- Marx, D.H. Ectomycorrhizal fungus inoculations: A tool for improving forestation practices. In Tropical Mycorrhiza Research; Mikola, P., Ed.; Oxford University Press: London, UK, 1980; pp. 13–71. [Google Scholar]
- Walker, R.F.; West, D.C.; McLaughlin, S.B.; Amundsen, C.C. Growth, xylem pressure potential and nutrient absorption of loblolly pine on a reclaimed surface mine as affected by an induced Pisolithus tinctorius infection. For. Sci. 1989, 35, 569–581. [Google Scholar]
- Sousa, N.R.; Ramos, M.A.; Marques, A.P.G.C.; Castro, P.M.L. The effect of ectomycorrhizal fungi forming symbiosis with Pinus pinaster seedlings exposed to cadmium. Sci. Total Environ. 2012, 414, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Onwuchekwa, N.E.; Zwiazek, J.J.; Quoreshi, A.; Khasa, D.P. Growth of mycorrhizal jack pine (Pinus banksiana) and white spruce (Picea glauca) seedlings planted in oil sands reclaimed areas. Mycorrhiza 2014, 24, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Adeleke, R.; Cloete, T.E.; Bertrand, A.; Khasa, D.P. Relationship between plant growth and organic acid exudates from ectomycorrhizal and non-ectomycorrhizal Pinus patula. S. Afr. J. Plant Soil 2015, 32, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Sebastiana, M.; Pereira, V.T.; Alcântara, A.; Pais, M.S.; Silva, A.B. Ectomycorrhizal inoculation with Pisolithus tinctorius increases the performance of Quercus suber L. (cork oak) nursery and field seedlings. New Forest. 2013, 44, 937–949. [Google Scholar] [CrossRef]
- Plassard, C.; Bonafox, B.; Touraine, B. Differential effects of mineral and organic N sources, and of ectomycorrhizal infection by Hebeloma cylindrosporum, on growth and N utilization in Pinus pinaster. Plant Cell. Environ. 2000, 23, 1195–1205. [Google Scholar] [CrossRef]
- Corrêa, A.; Gurevitch, J.; Martins-Loução, M.A.; Cruz, C. C allocation to the fungus is not a cost to the plant in ectomycorrhizae. Oikos 2012, 121, 449–463. [Google Scholar] [CrossRef]
- Lehto, T.; Zwiazek, J.J. Ectomycorrhizas and water relations of trees: A review. Mycorrhiza 2011, 2, 71–90. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Kou, Y. Ectomycorrhizal fungi: Participation in nutrient turnover and community assembly pattern in forest ecosystems. Forest 2020, 11, 453. [Google Scholar] [CrossRef]
- Krupa, Z.; Siedlecka, A.; Skórzynska-Polit, E.; Maksymiec, W. Heavy metal interactions with plant nutrients. In Physiology and Biochemistry of Metal Toxicity and Tolerance in Plants; Prasad, M.N., Strzalka, K., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 287–301. [Google Scholar]
- Futai, K.; Taniguchi, T.; Kataoka, R. Ectomycorrhizae and their importance in forest ecosystems. In Mycorrhizae: Sustainable Agriculture and Forestry; Siddiqui, Z.A., Akhtar, M.S., Futai, K., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 241–285. [Google Scholar] [CrossRef]
- Jentschke, G.; Brandes, B.; Kuhn, A.J.; Schroder, W.H.; Godbold, D.L. Interdependence of phosphorus, nitrogen, potassium and magnesium translocation by the ectomycorrhizal fungus Paxillus involutus. New Phytol. 2001, 149, 327–337. [Google Scholar] [CrossRef]
- Nehls, U.; Plassard, C. Nitrogen and phosphate metabolism in ectomycorrhizas. New Phytol. 2018, 220, 1047–1058. [Google Scholar] [CrossRef] [Green Version]
- Plassard, C.; Dell, B. Phosphorus nutrition of mycorrhizal trees. Tree Physiol. 2010, 30, 1129–1139. [Google Scholar] [CrossRef] [Green Version]
- Plassard, C.; Louche, J.; Ali, M.A.; Duchemin, M.; Legname, E.; Cloutier-Hurteau, B. Diversity in phosphorus mobilisation and uptake in ectomycorrhizal fungi. Ann. For. Sci. 2011, 68, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Zhao, Y.; Tian, K.; He, X.; Jia, Y.; He, Z.; Tian, X. Insight into nitrogen and phosphorus enrichment on cadmium phytoextraction of hydroponically grown Salix matsudana Koidz. cuttings. Environ. Sci. Pollut. Res. 2020, 27, 8406–8417. [Google Scholar] [CrossRef]
- Lamhamedi, M.S.; Renaud, M.; Veilleux, L. Les effets de l’augmentation du pH des substrats sur la croissance des plants forestiers produits dans les pépinières forestières. In Production de Plants Forestiers au Québec: La Culture de L’innovation. In Proceedings of the Colloque de transfert de connaissances et de savoir-faire, Carrefour Forêt Innovations, Québec, QC, Canada, 4–6 October 2011; pp. 33–45. [Google Scholar]
- Kasowska, D.; Gediga, K.; Spiak, Z. Heavy metal and nutrient uptake in plants colonizing post-flotation copper tailings. Environ. Sci. Pollut. Res. 2018, 25, 824–835. [Google Scholar] [CrossRef] [Green Version]
- Godbold, D.L.; Kettner, C. Lead influences root growth and mineral nutrition of Picea abies seedlings. J. Plant Physiol. 1991, 139, 95–99. [Google Scholar] [CrossRef]
- Małkowski, E.; Kurtyka, R.; Kita, A.; Karcz, W. Accumulation of Pb and Cd and its effect on Ca distribution in maize seedlings (Zea mays L.). Pol. J. Environ. Stud. 2005, 14, 203–207. [Google Scholar]
- Gu, H.H.; Zhou, Z.; Gao, Y.Q.; Yuan, X.T.; Ai, Y.J.; Zhang, J.Y.; Li, F.P. The influences of arbuscular mycorrhizal fungus on phytostabilization of lead/zinc tailings using four plant species. Int. J. Phytoremediation 2017, 19, 739–745. [Google Scholar] [CrossRef]
- Schützendübel, A.; Polle, A. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization. J. Exp. Bot. 2002, 53, 1351–1365. [Google Scholar] [CrossRef] [PubMed]
- Ahonen-Jonnarth, U.; van Hees, P.A.W.; Lundström, U.S.; Finlay, R.D. Production of organic acids by mycorrhizal and non-mycorrhizal Pinus sylvestris L. seedlings exposed to elevated concentrations of aluminium and heavy metals. New Phytol. 2000, 146, 557–567. [Google Scholar] [CrossRef]
- Bellion, M.; Courbot, M.; Jacob, C.; Blaudez, D.; Chalot, M. Extracellular and cellular mechanisms sustaining metal tolerance in ectomycorrhizal fungi. FEMS Microbiol. Lett. 2006, 254, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Khullar, S.; Reddy, M.S. Ectomycorrhizal fungi and its role in metal homeostasis through metallothionein and glutathione mechanisms. Curr. Biotechnol. 2018, 7, 231–241. [Google Scholar] [CrossRef]
- Ma, Y.L.; He, J.L.; Ma, C.F.; Luo, J.; Li, H.; Liu, T.X.; Polle, A.; Peng, C.H.; Luo, Z.B. Ectomycorrhizas with Paxillus involutus enhance cadmium uptake and tolerance in Populus x canescens. Plant Cell Environ. 2014, 37, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Adriano, D.C. Trace Elements in Terrestrial Environments: Biogeochemistry, Bioavailability, and Risks of Metals; Springer: Berlin, Germany, 2001; p. 867. [Google Scholar]
- Pourrut, B.; Perchet, G.; Silvestre, J.; Cecchi, M.; Guiresse, M.; Pinelli, E. Potential role of NADPH-oxidase in early steps of lead-induced oxidative burst in Vicia faba roots. J. Plant Physiol. 2008, 165, 571–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Contaminated Soil | Control Soil | |
|---|---|---|
| pHwater | 8.87 ± 0.02 a | 7.59 ± 0.08 b |
| pHCaCl2 | 7.73 ± 0.11 a | 6.42 ± 0.14 b |
| EC (µS·cm−1) | 255.3 ± 3.51 a | 152.8 ± 9.25 b |
| Carbon (mg·g−1) | 44.300 ± 4.050 a | 10.433 ± 1.484 b |
| Pb (mg·g−1) | 15.587 ± 0.796 a | 0.009 ± 0.0004 b |
| Zn (mg·g−1) | 37.766 ± 3.210 a | 0.021 ± 0.002 b |
| Cd (mg·g−1) | 0.181 ± 0.033 | 0 |
| N (mg·g−1) | <1 | <1 |
| P (mg·g−1) | 0.622 ± 0.042 a | 0.084 ± 0.001 b |
| K (mg·g−1) | 2.236 ± 0.299 a | 0.899 ± 0.073 b |
| Ca (mg·g−1) | 252.483 ± 8.765 a | 2.076 ± 0.169 b |
| Mg (mg·g−1) | 6.034 ± 0.232 a | 0.461 ± 0.015 b |
| Fe (mg·g−1) | 7.824 ± 0.504 a | 4.205 ± 0.559 b |
| Mn (mg·g−1) | 0.307 ± 0.004 a | 0.070 ± 0.004 b |
| Cu (mg·g−1) | 0.036 ± 0.002 a | 0.017 ± 0.014 b |
| B (mg·g−1) | 0.028 ± 0.002 a | 0.016 ± 0.0003 b |
| Na (mg·g−1) | 0.694 ± 0.068 a | 0.350 ± 0.042 b |
| Shoots (mg.Seedling−1) | Roots (mg.Seedling−1) | Soil (mg.g−1) | ||||
|---|---|---|---|---|---|---|
| Elements | Mycorrhizal | Non-mycorrhizal | Mycorrhizal | Non-mycorrhizal | Mycorrhizal | Non-mycorrhizal |
| Pb | 0.023 ± 0.009 a | 0.024 ± 0.002 a | 0.007 ± 0.0005 b | 0.009 ± 0.0006 a | 0.011 ± 0.0008 b | 0.016 ± 0.001 a |
| Zn | 0.101 ± 0.0005 b | 0.128 ± 0.002 a | 0.036 ± 0.001 a | 0.034 ± 0.001 a | 0.038 ± 0.002 b | 0.050 ± 0.002 a |
| Cd | 0 | 0 | 0 | 0 | 0 | 0 |
| N | 31.641 ± 0.199 a | 27.183 ± 0.562 b | 6.272 ± 0.148 a | 4.725 ± 0.129 b | 1.633 ± 0.057 a | 1.733 ± 0.057 a |
| P | 4.864 ± 0.097 a | 4.675 ± 0.137 a | 1.546 ± 0.001 a | 1.919 ± 1.061 a | 0.265 ± 0.024 a | 0.273 ± 0.017 a |
| K | 14.585 ± 0.367 a | 13.811 ± 0.459 a | 3.238 ± 0.009 a | 2.599 ± 0.057 b | 1.087 ± 0.066 a | 1.167 ± 0.046 a |
| Ca | 12.480 ± 0.132 b | 13.644 ± 0.337 a | 7.328 ± 0.353 a | 5.862 ± 0.116 b | 4.700 ± 0.284 b | 5.247 ± 0.074 a |
| Mg | 3.848 ± 0.044 b | 4.033 ± 0.029 a | 1.646 ± 0.045 a | 1.469 ± 0.026 b | 1.207 ± 0.077 a | 1.234 ± 0.106 a |
| Fe | 0.856 ± 0.022 b | 1.323 ± 0.081 a | 2.496 ± 0.389 a | 2.578 ± 0.146 a | 8.441 ± 0.676 a | 9.319 ± 2.306 a |
| Mineral Nutrient Contents (mg.Seedling−1) | |||||||
|---|---|---|---|---|---|---|---|
| Treatment | N | P | K | Ca | Mg | Fe | |
| Shoots | NM-NC | 178.18 ± 17.87 c | 25.37 ± 10.48 b | 148.60 ± 26.36 b | 187.66 ± 33.11 b | 51.18 ± 11.75 b | 7.22 ± 0.57 a |
| M-NC | 477.84 ± 12.39 a | 49.69 ± 9.11 a | 220.79 ± 16.32 a | 258.20 ± 21.59 a | 72.43 ± 5.63 a | 7.57 ± 1.49 a | |
| NM-C | 114.07 ± 7.65 d | 11.20 ± 1.69 c | 87.36 ± 3.84 c | 185.38 ± 14.58 b | 43.21 ± 4.79 b | 5.22 ± 0.31 b | |
| M-C | 266.11 ± 5.05 b | 32.01 ± 4.39 b | 115.44 ± 18.81 b | 264.79 ± 15.25 a | 54.56 ± 7.75 b | 5.47 ± 0.10 b | |
| Roots | NM-NC | 143.71 ± 9.69 b | 16.20 ± 2.12 b | 109.32 ± 13.23 ab | 335.64 ± 50.63 b | 48.84 ± 9.76 a | 50.95 ± 4.48 b |
| M-NC | 205.14 ± 19.08 a | 23.06 ± 4.55 a | 128.73 ± 26.61 a | 382.70 ± 44.75 b | 54.06 ± 9.73 a | 75.83 ± 15.49 a | |
| NM-C | 54.78 ± 1.72 d | 6.40 ± 0.29 c | 45.10 ± 3.73 c | 348.78 ± 36.38 b | 26.59 ± 2.45 b | 29.14 ± 16.42 c | |
| M-C | 117.16 ± 4.36 c | 19.24 ± 0.93 ab | 90.54 ± 7.74 b | 711.18 ± 135.02 a | 44.43 ± 6.55 a | 32.71 ± 3.24 c | |
| Metallic Element | |||
|---|---|---|---|
| Pb | Zn | Cd | |
| TF | |||
| NM-C | 0.030 ± 0.006 a | 0.124 ± 0.009 a | 0.191 ± 0.007 a |
| M-C | 0.031 ± 0.008 a | 0.086 ± 0.005 b | 0.106 ± 0.004 b |
| BAF | |||
| NM-C | 0.161 ± 0.026 a | 0.062 ± 0.009 a | 0.062 ± 0.009 a |
| M-C | 0.100 ± 0.010 b | 0.054 ± 0.002 a | 0.044 ± 0.006 b |
| Final concentration in soil | |||
| NM-C (mg.g−1) | 9.678 ± 0.320 a | 27.218 ± 8.839 a | 0.116 ± 0.030 a |
| M-C (mg.g−1) | 10.128 ± 1.554 a | 23.672 ± 4.353 a | 0.104 ± 0.007 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hachani, C.; Lamhamedi, M.S.; Cameselle, C.; Gouveia, S.; Zine El Abidine, A.; Khasa, D.P.; Béjaoui, Z. Effects of Ectomycorrhizal Fungi and Heavy Metals (Pb, Zn, and Cd) on Growth and Mineral Nutrition of Pinus halepensis Seedlings in North Africa. Microorganisms 2020, 8, 2033. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122033
Hachani C, Lamhamedi MS, Cameselle C, Gouveia S, Zine El Abidine A, Khasa DP, Béjaoui Z. Effects of Ectomycorrhizal Fungi and Heavy Metals (Pb, Zn, and Cd) on Growth and Mineral Nutrition of Pinus halepensis Seedlings in North Africa. Microorganisms. 2020; 8(12):2033. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122033
Chicago/Turabian StyleHachani, Chadlia, Mohammed S. Lamhamedi, Claudio Cameselle, Susana Gouveia, Abdenbi Zine El Abidine, Damase P. Khasa, and Zoubeir Béjaoui. 2020. "Effects of Ectomycorrhizal Fungi and Heavy Metals (Pb, Zn, and Cd) on Growth and Mineral Nutrition of Pinus halepensis Seedlings in North Africa" Microorganisms 8, no. 12: 2033. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122033