SARS-Cov-2 Interactome with Human Ghost Proteome: A Neglected World Encompassing a Wealth of Biological Data



Abstract
:1. Introduction
2. Material and Methods
Ghost Proteins Databases
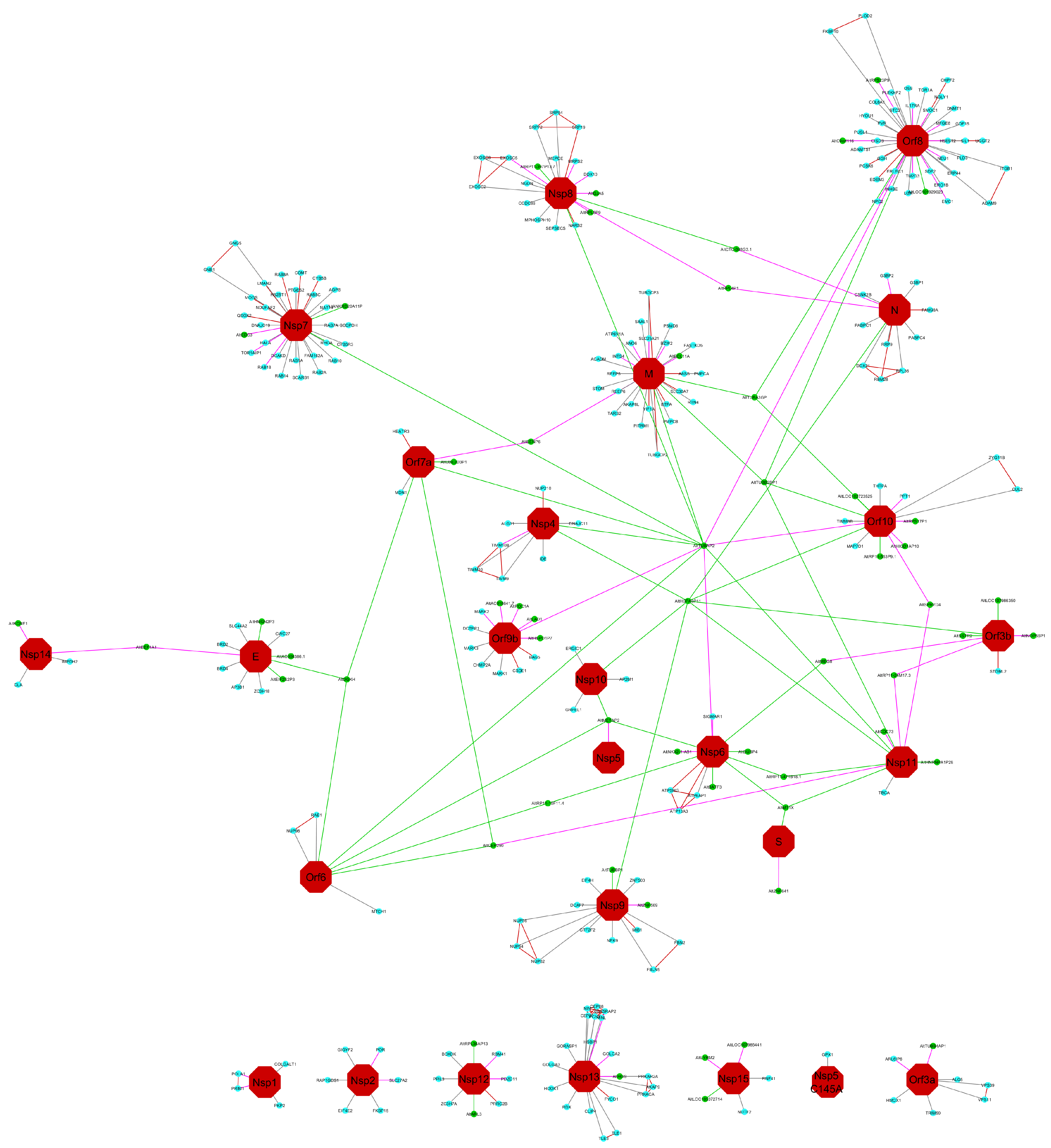
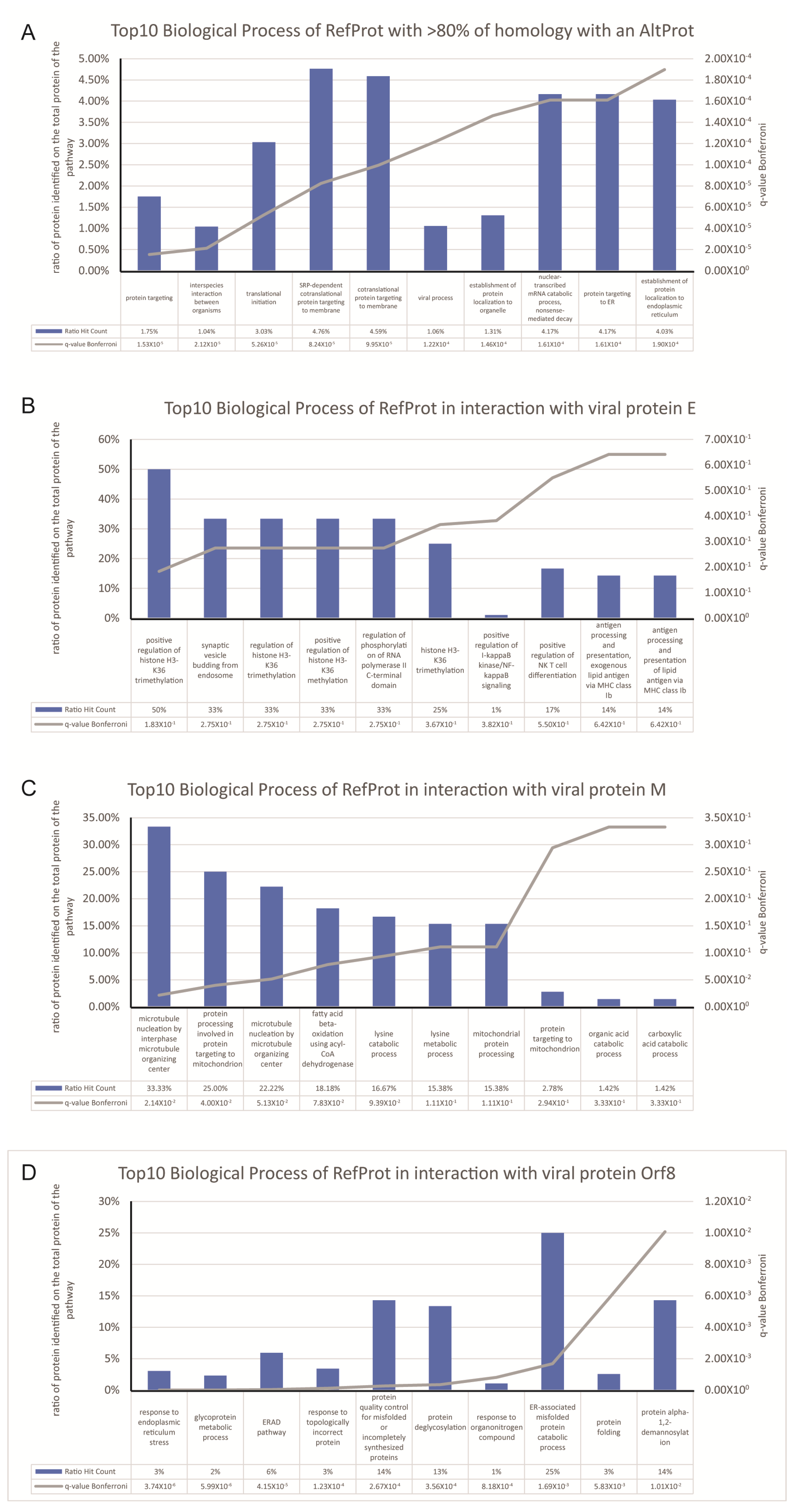
3. Results and Discussion
Authors Contribution
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Zubarev, R.A. The challenge of the proteome dynamic range and its implications for in-depth proteomics. Proteomics 2013, 13, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Meier, F.; Geyer, P.E.; Virreira Winter, S.; Cox, J.; Mann, M. BoxCar acquisition method enables single-shot proteomics at a depth of 10,000 proteins in 100 minutes. Nat. Methods 2018, 15, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Boeckmann, B.; Bairoch, A.; Apweiler, R.; Blatter, M.-C.; Estreicher, A.; Gasteiger, E.; Martin, M.J.; Michoud, K.; O’donovan, C.; Phan, I.; et al. The SWISS-PROT protein knowledgebase and its supplement TrEMBL in 2003. Nucleic Acids Res. 2003, 31, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M. Regulation of translation in eukaryotic systems. Annu. Rev. Cell Biol. 1992, 8, 197–225. [Google Scholar] [CrossRef]
- Ingolia, N.T.; Ghaemmaghami, S.; Newman, J.R.S.; Weissman, J.S. Genome-wide analysis in vivo of translation with nucleotide resolution using ribosome profiling. Science 2009, 324, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Ingolia, N.T.; Lareau, L.F.; Weissman, J.S. Ribosome profiling of mouse embryonic stem cells reveals the complexity and dynamics of mammalian proteomes. Cell 2011, 147, 789–802. [Google Scholar] [CrossRef] [Green Version]
- Bazzini, A.A.; Johnstone, T.G.; Christiano, R.; MacKowiak, S.D.; Obermayer, B.; Fleming, E.S.; Vejnar, C.E.; Lee, M.T.; Rajewsky, N.; Walther, T.C.; et al. Identification of small ORFs in vertebrates using ribosome footprinting and evolutionary conservation. EMBO J. 2014, 33, 981–993. [Google Scholar] [CrossRef] [Green Version]
- Vanderperre, B.; Lucier, J.-F.; Bissonnette, C.; Motard, J.; Tremblay, G.; Vanderperre, S.; Wisztorski, M.; Salzet, M.; Boisvert, F.-M.; Roucou, X.; et al. Direct Detection of Alternative Open Reading Frames Translation Products in Human Significantly Expands the Proteome. PLoS ONE 2013, 8, e70698. [Google Scholar] [CrossRef] [Green Version]
- Mouilleron, H.; Delcourt, V.; Roucou, X. Death of a dogma: Eukaryotic mRNAs can code for more than one protein. Nucleic Acids Res. 2016, 44, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Cardon, T.; Franck, J.; Coyaud, E.; Laurent, E.M.N.; Damato, M.; Maffia, M.; Vergara, D.; Fournier, I.; Salzet, M. Alternative proteins are functional regulators in cell reprogramming by PKA activation. Nucleic Acids Res. 2020. [Google Scholar] [CrossRef]
- Murgoci, A.N.; Cardon, T.; Aboulouard, S.; Duhamel, M.; Fournier, I.; Cizkova, D.; Salzet, M. Reference and Ghost Proteins Identification in Rat C6 Glioma Extracellular Vesicles. iScience 2020, 23, 101045. [Google Scholar] [CrossRef] [PubMed]
- Samandi, S.; Roy, A.V.; Delcourt, V.; Lucier, J.F.; Gagnon, J.; Beaudoin, M.C.; Vanderperre, B.; Breton, M.A.; Motard, J.; Jacques, J.F.; et al. Deep transcriptome annotation enables the discovery and functional characterization of cryptic small proteins. Elife 2017, 6, e27860. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, V.; Franck, J.; Leblanc, E.; Narducci, F.; Robin, Y.M.; Gimeno, J.P.; Quanico, J.; Wisztorski, M.; Kobeissy, F.; Jacques, J.F.; et al. Combined Mass Spectrometry Imaging and Top-down Microproteomics Reveals Evidence of a Hidden Proteome in Ovarian Cancer. EBioMedicine 2017, 21, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delcourt, V.; Franck, J.; Quanico, J.; Gimeno, J.P.; Wisztorski, M.; Raffo-Romero, A.; Kobeissy, F.; Roucou, X.; Salzet, M.; Fournier, I. Spatially-Resolved Top-down Proteomics Bridged to MALDI MS Imaging Reveals the Molecular Physiome of Brain Regions. Mol. Cell. Proteom. 2018, 17, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Khitun, A.; Na, Z.; Phoodokmai, T.; Sappakhaw, K.; Olatunji, E.; Uttamapinant, C.; Slavoff, S.A. Alt-RPL36 downregulates the PI3K-AKT-mTOR signaling pathway by interacting with TMEM24. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.L.; Meller, A.; Samandi, S.; Brunelle, M.; Frion, J.; Brunet, M.A.; Toupin, A.; Beaudoin, M.C.; Jacques, J.F.; Lévesque, D.; et al. UBB pseudogene 4 encodes functional ubiquitin variants. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Chen, J.; Brunner, A.D.; Cogan, J.Z.; Nuñez, J.K.; Fields, A.P.; Adamson, B.; Itzhak, D.N.; Li, J.Y.; Mann, M.; Leonetti, M.D.; et al. Pervasive functional translation of noncanonical human open reading frames. Science 2020, 367, 140–146. [Google Scholar] [CrossRef]
- Brunet, M.A.; Brunelle, M.; Lucier, J.F.; Delcourt, V.; Levesque, M.; Grenier, F.; Samandi, S.; Leblanc, S.; Aguilar, J.D.; Dufour, P.; et al. OpenProt: A more comprehensive guide to explore eukaryotic coding potential and proteomes. Nucleic Acids Res. 2019, 47, D403–D410. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Rijkers, D.T.S.S.; Post, H.; Heck, A.J.R.R. Proteome-wide profiling of protein assemblies by cross-linking mass spectrometry. Nat. Methods 2015, 12, 1179–1184. [Google Scholar] [CrossRef]
- Cardon, T.; Salzet, M.; Franck, J.; Fournier, I. Nuclei of HeLa cells interactomes unravel a network of ghost proteins involved in proteins translation. Biochim. Biophys. Acta-Gen. Subj. 2019. [Google Scholar] [CrossRef]
- Gordon, D.; Jang, G.; Bouhaddou, M.; Xu, J.; Obernier, K.; O’Meara, M.; Guo, J.; Swaney, D.; Tummino, T.; Hüttenhain, R.; et al. A SARS-CoV-2-Human Protein-Protein Interaction Map Reveals Drug Targets and Potential Drug-Repurposing. bioRxiv Prepr. Serv. Biol. 2020, 19, 4. [Google Scholar] [CrossRef] [Green Version]
- Delcourt, V.; Brunelle, M.; Roy, A.V.; Jacques, J.-F.; Salzet, M.; Fournier, I.; Roucou, X. The Protein Coded by a Short Open Reading Frame, Not by the Annotated Coding Sequence, Is the Main Gene Product of the Dual-Coding Gene MIEF1. Mol. Cell. Proteom. 2018, 17, 2402–2411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Goenawan, I.H.; Bryan, K.; Lynn, D.J. DyNet: Visualization and analysis of dynamic molecular interaction networks. Bioinformatics 2016, 32, 2713–2715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, J.Y.; Yang, J.S.; Kim, J.W.; Kim, V.N.; Chang, H. The Architecture of SARS-CoV-2 Transcriptome. Cell 2020, 181, 914–921.e10. [Google Scholar] [CrossRef]
- Jungreis, I.; Nelson, C.W.; Ardern, Z.; Finkel, Y.; Krogan, N.J.; Sato, K.; Ziebuhr, J.; Stern-Ginossar, N.; Pavesi, A.; Firth, A.E.; et al. Conflicting and ambiguous names of overlapping ORFs in SARS-CoV-2: A homology-based resolution. Biochemistry 2020. [Google Scholar] [CrossRef]
- Brunet, M.A.; Roucou, X. Mass spectrometry-based proteomics analyses using the openprot database to unveil novel proteins translated from non-canonical open reading frames. J. Vis. Exp. 2019, 2019, 59589. [Google Scholar] [CrossRef] [Green Version]
- Bouhaddou, M.; Memon, D.; Meyer, B.; White, K.M.; Rezelj, V.V.; Marrero, M.C.; Polacco, B.J.; Melnyk, J.E.; Ulferts, S.; Kaake, R.M.; et al. The Global Phosphorylation Landscape of SARS-CoV-2 Infection. Cell 2020. [Google Scholar] [CrossRef]
- Hillen, H.S.; Kokic, G.; Farnung, L.; Dienemann, C.; Tegunov, D.; Cramer, P. Structure of replicating SARS-CoV-2 polymerase. Nature 2020, 584, 154–156. [Google Scholar] [CrossRef]
- Kirchdoerfer, R.N.; Ward, A.B. Structure of the SARS-CoV nsp12 polymerase bound to nsp7 and nsp8 co-factors. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subissi, L.; Posthuma, C.C.; Collet, A.; Zevenhoven-Dobbe, J.C.; Gorbalenya, A.E.; Decroly, E.; Snijder, E.J.; Canard, B.; Imbert, I. One severe acute respiratory syndrome coronavirus protein complex integrates processive RNA polymerase and exonuclease activities. Proc. Natl. Acad. Sci. USA 2014, 111, E3900–E3909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jodele, S.; Köhl, J. Tackling COVID-19 infection through complement-targeted immunotherapy. Br. J. Pharmacol. 2020. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Accession | GN | TA | AltProt | Type | Location | Viral Protein in Interaction |
|---|---|---|---|---|---|---|
| IP_555327 | AC006386.1 | ENST00000624491 | AltAC006386.1 | mRNA | 3’UTR | E |
| IP_581922 | AC018641.7 | ENST00000435950 | AltAC018641.7 | ncRNA | - | orf9b |
| IP_659614 | ANKRD20A11P | ENST00000442192 | AltANKRD20A11P | ncRNA | - | nsp7 |
| IP_187691 | ARL3 | NM_004311.3 | AltARL3 | mRNA | 3’UTR | nsp12 |
| IP_077449 | BATF3 | NM_018664.2 | AltBATF3 | mRNA | CDS | nsp6 |
| IP_2387661 | BCL11A | XM_017004337.1 | AltBCL11A | mRNA | 5’UTR | M |
| IP_565887 | C9orf116 | ENST00000371789 | AltC9orf116 | mRNA | 5’UTR | orf8 |
| IP_075271 | CDC73 | XM_006711537.3 | AltCDC73 | mRNA | CDS | nsp11 |
| IP_766056 | CEP290 | ENST00000547691 | AltCEP290 | mRNA | 5’UTR | nsp11 orf6 orf7a |
| IP_691726 | CTC-398G3.1 | ENST00000483614 | AltCTC-398G3.1 | ncRNA | - | N nsp8 |
| IP_219869 | DGKH | NM_152910.5 | AltDGKH | mRNA | CDS | E orf6 orf7a |
| IP_2336782 | DUSP4 | XM_011544428.2 | AltDUSP4 | mRNA | 5’UTR | nsp6 |
| IP_235699 | EDC3 | ENST00000565602 | AltEDC3 | mRNA | 3’UTR | nsp7 |
| IP_594707 | EEF1A1 | ENST00000309268 | AltEEF1A1 | mRNA | 5’UTR | E nsp14 |
| IP_788706 | EIF2S2P3 | ENST00000428356 | AltEIF2S2P3 | ncRNA | - | E |
| IP_2396759 | GJA5 | XM_017001044.1 | AltGJA5 | mRNA | 5’UTR | nsp8 |
| IP_711582 | HGS | ENST00000577012 | AltHGS | mRNA | 5’UTR | nsp13 |
| IP_775502 | HIGD1AP10 | ENST00000527837 | AltHIGD1AP10 | ncRNA | - | orf10 |
| IP_724315 | HMGN2P3 | ENST00000433603 | AltHMGN2P3 | ncRNA | - | E |
| IP_557348 | HNRNPA1P28 | ENST00000424481 | AltHNRNPA1P28 | ncRNA | - | nsp11 |
| IP_572435 | HSPA8P11 | ENST00000508840 | AltHSPA8P11 | ncRNA | - | N nsp10 nsp11 nsp4 nsp9 orf10 orf3b |
| IP_658154 | HSPD1P7 | ENST00000447985 | AltHSPD1P7 | ncRNA | - | orf9b |
| IP_289249 | KCNE1 | XM_017028342.1 | AltKCNE1 | mRNA | 3’UTR | nsp14 |
| IP_075761 | LAD1 | ENST00000631576 | AltLAD1 | mRNA | CDS | orf9b |
| IP_671071 | LOC101929023 | ENST00000434879 | AltLOC101929023 | ncRNA | - | orf8 |
| IP_2361135 | LOC102723525 | XR_925379.2 | AltLOC102723525 | ncRNA | - | orf10 |
| IP_2268667 | LOC105372714 | XM_017028195.1 | AltLOC105372714 | mRNA | 5’UTR | nsp15 |
| IP_2266298 | LOC107985441 | XR_001754616.1 | AltLOC107985441 | ncRNA | - | nsp15 |
| IP_2354489 | LOC107986350 | XR_001742414.1 | AltLOC107986350 | ncRNA | - | orf3b |
| IP_143572 | LYRM2 | NM_020466.4 | AltLYRM2 | mRNA | 3’UTR | nsp15 |
| IP_745252 | MEG8 | ENST00000553465 | AltMEG8 | ncRNA | - | nsp6 orf3b |
| IP_213668 | METAP2 | XM_005268583.3 | AltMETAP2 | mRNA | CDS | nsp10 nsp6 orf6 |
| IP_729791 | MT1X | ENST00000568370 | AltMT1X | mRNA | 3’UTR | nsp11 nsp6 |
| IP_230046 | NKX2-1-AS1 | ENST00000521292 | AltNKX2-1-AS1 | ncRNA | - | nsp6 |
| IP_597201 | NOP56P1 | ENST00000440030 | AltNOP56P1 | ncRNA | - | orf3b |
| IP_105102 | POC1A | XM_011533561.1 | AltPOC1A | mRNA | CDS | orf9b |
| IP_581419 | RP11-10F11.4 | ENST00000634439 | AltRP11-10F11.4 | ncRNA | - | nsp6 orf6 |
| IP_734708 | RP11-24M17.3 | ENST00000567565 | AltRP11-24M17.3 | ncRNA | - | nsp11 orf3b |
| IP_667059 | RP11-397P13.7 | ENST00000427282 | AltRP11-397P13.7 | ncRNA | - | nsp8 |
| IP_591742 | RP11-471B18.1 | ENST00000407538 | AltRP11-471B18.1 | ncRNA | - | nsp11 nsp6 |
| IP_612631 | RP11-553P9.1 | ENST00000509116 | AltRP11-553P9.1 | ncRNA | - | orf10 |
| IP_639311 | RPL36AP13 | ENST00000457490 | AltRPL36AP13 | ncRNA | - | nsp12 |
| IP_750273 | RPL4P1 | ENST00000496596 | AltRPL4P1 | ncRNA | - | N nsp8 |
| IP_637436 | RPL5P9 | ENST00000448118 | AltRPL5P9 | ncRNA | - | nsp8 |
| IP_597129 | RPS17P1 | ENST00000396783 | AltRPS17P1 | ncRNA | - | orf10 |
| IP_668819 | RPS23P9 | ENST00000448848 | AltRPS23P9 | ncRNA | - | orf8 |
| IP_594653 | SENP6 | ENST00000474906 | AltSENP6 | ncRNA | - | M orf7a |
| IP_769089 | SPRYD4 | ENST00000338146 | AltSPRYD4 | mRNA | 3’UTR | nsp11 orf10 |
| IP_713094 | SSTR2 | ENST00000357585 | AltSSTR2 | mRNA | 3’UTR | orf3b |
| IP_656465 | TUBA3GP | ENST00000410028 | AltTUBA3GP | ncRNA | - | M orf10 orf8 |
| IP_774695 | TUBAP2 | ENST00000530835 | AltTUBAP2 | ncRNA | - | M nsp11 nsp4 nsp6 nsp7 nsp8 orf10 orf6 orf7a orf8 orf9b |
| IP_557241 | TUBB4AP1 | ENST00000450755.1 | Alt TUBB4AP1 | ncRNA | - | orf3a |
| IP_593099 | TUBB2BP1 | ENST00000404155 | AltTUBB2BP1 | ncRNA | - | M nsp11 orf10 orf8 |
| IP_572422 | TUBBP1 | ENST00000518096 | AltTUBBP1 | ncRNA | - | nsp9 |
| IP_665452 | UBE2D3P1 | ENST00000436669 | AltUBE2D3P1 | ncRNA | - | orf7a |
| IP_274314 | ZNF569 | XM_006723046.2 | AltZNF569 | mRNA | 3’UTR | nsp9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardon, T.; Fournier, I.; Salzet, M. SARS-Cov-2 Interactome with Human Ghost Proteome: A Neglected World Encompassing a Wealth of Biological Data. Microorganisms 2020, 8, 2036. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122036
Cardon T, Fournier I, Salzet M. SARS-Cov-2 Interactome with Human Ghost Proteome: A Neglected World Encompassing a Wealth of Biological Data. Microorganisms. 2020; 8(12):2036. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122036
Chicago/Turabian StyleCardon, Tristan, Isabelle Fournier, and Michel Salzet. 2020. "SARS-Cov-2 Interactome with Human Ghost Proteome: A Neglected World Encompassing a Wealth of Biological Data" Microorganisms 8, no. 12: 2036. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122036