Bacterial Succession through the Artisanal Process and Seasonal Effects Defining Bacterial Communities of Raw-Milk Adobera Cheese Revealed by High Throughput DNA Sequencing

 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. DNA Extraction
2.3. Construction of the 16S rRNA Libraries and High Throughput DNA Sequencing
2.4. Bioinformatics
3. Results
3.1. Bioinformatic Analysis
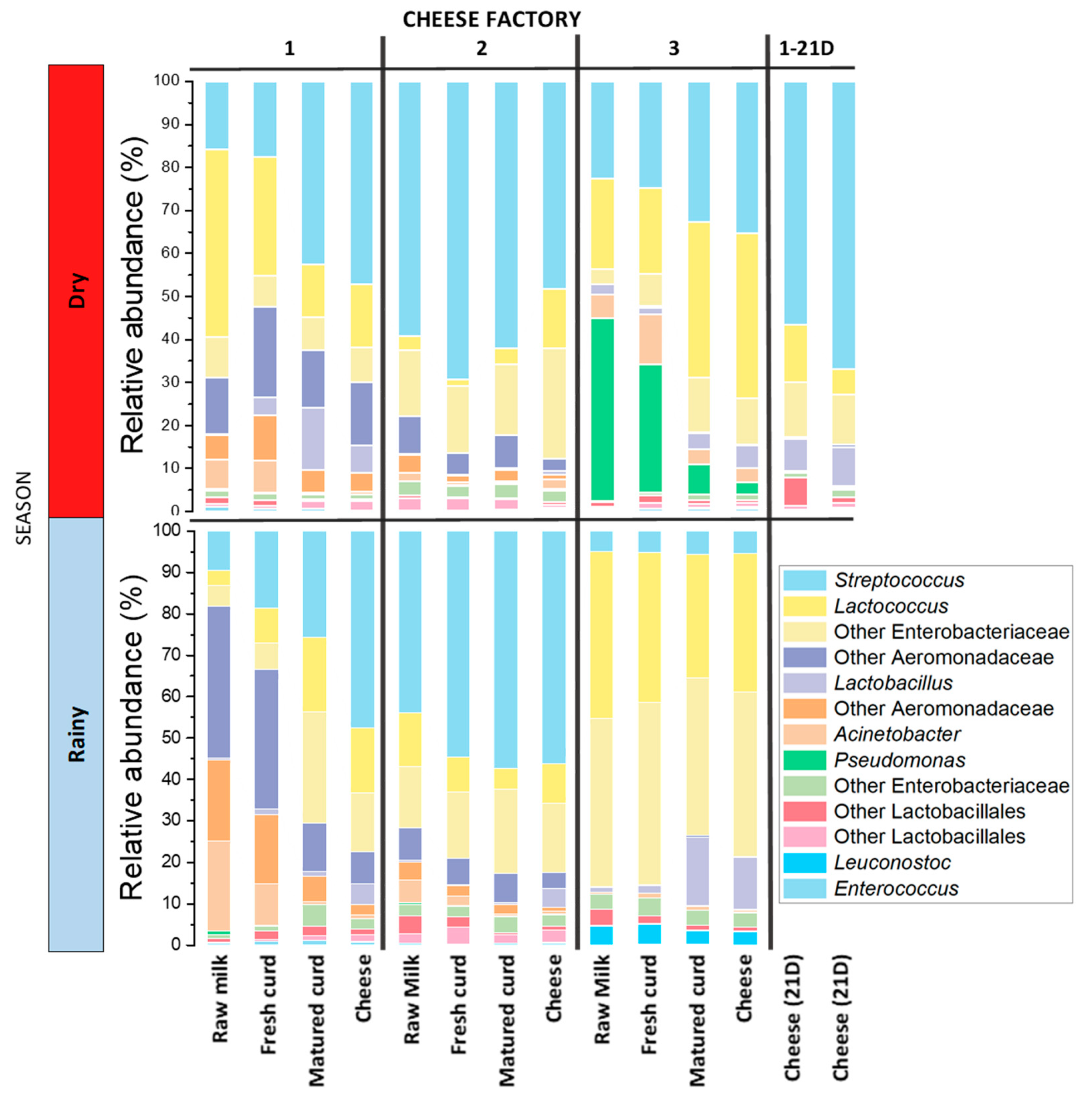
3.2. Differences in Microbiota Composition: From Milk to Cheese
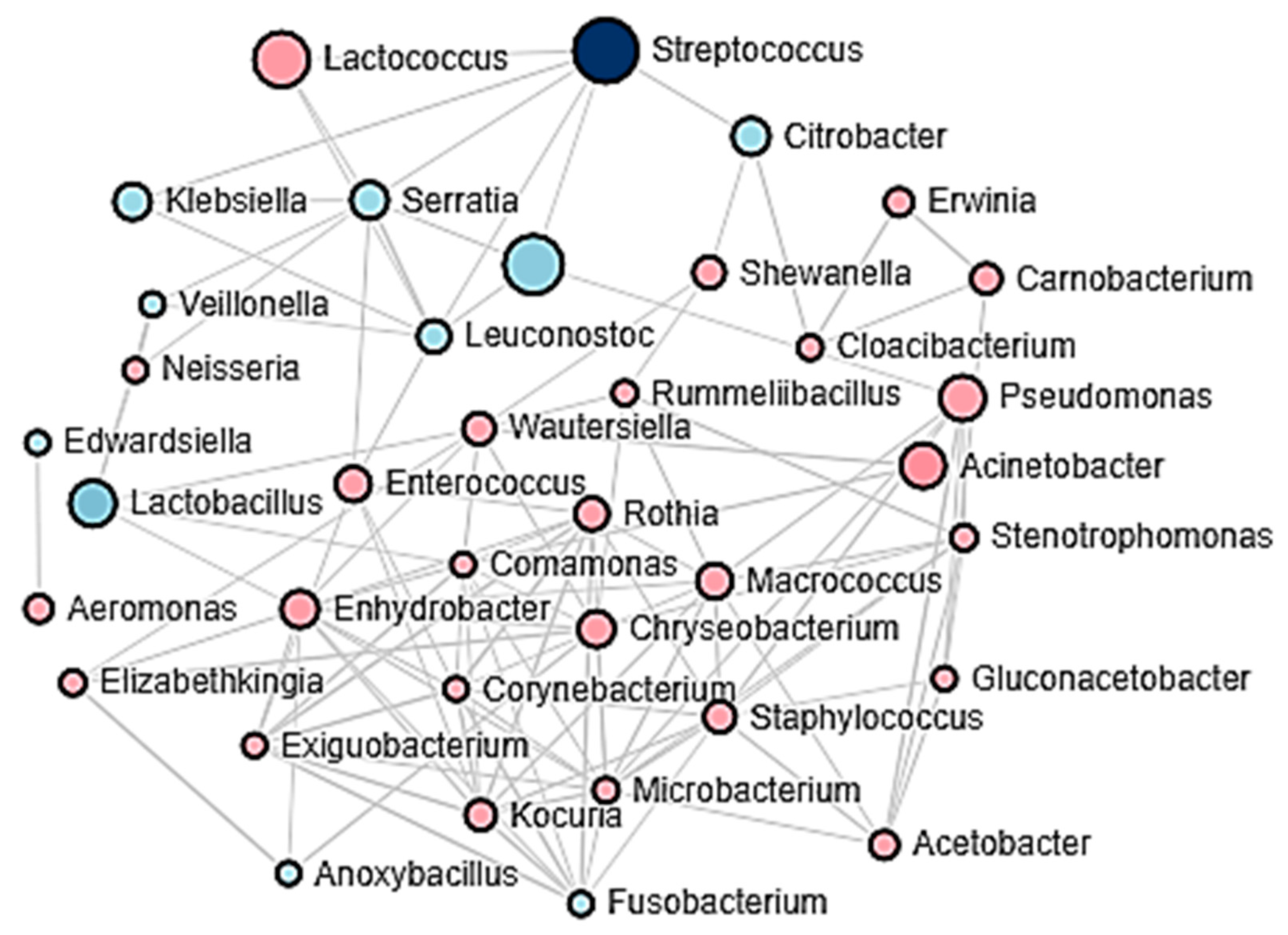
3.3. Bacterial Communities of Adobera Cheese during Process
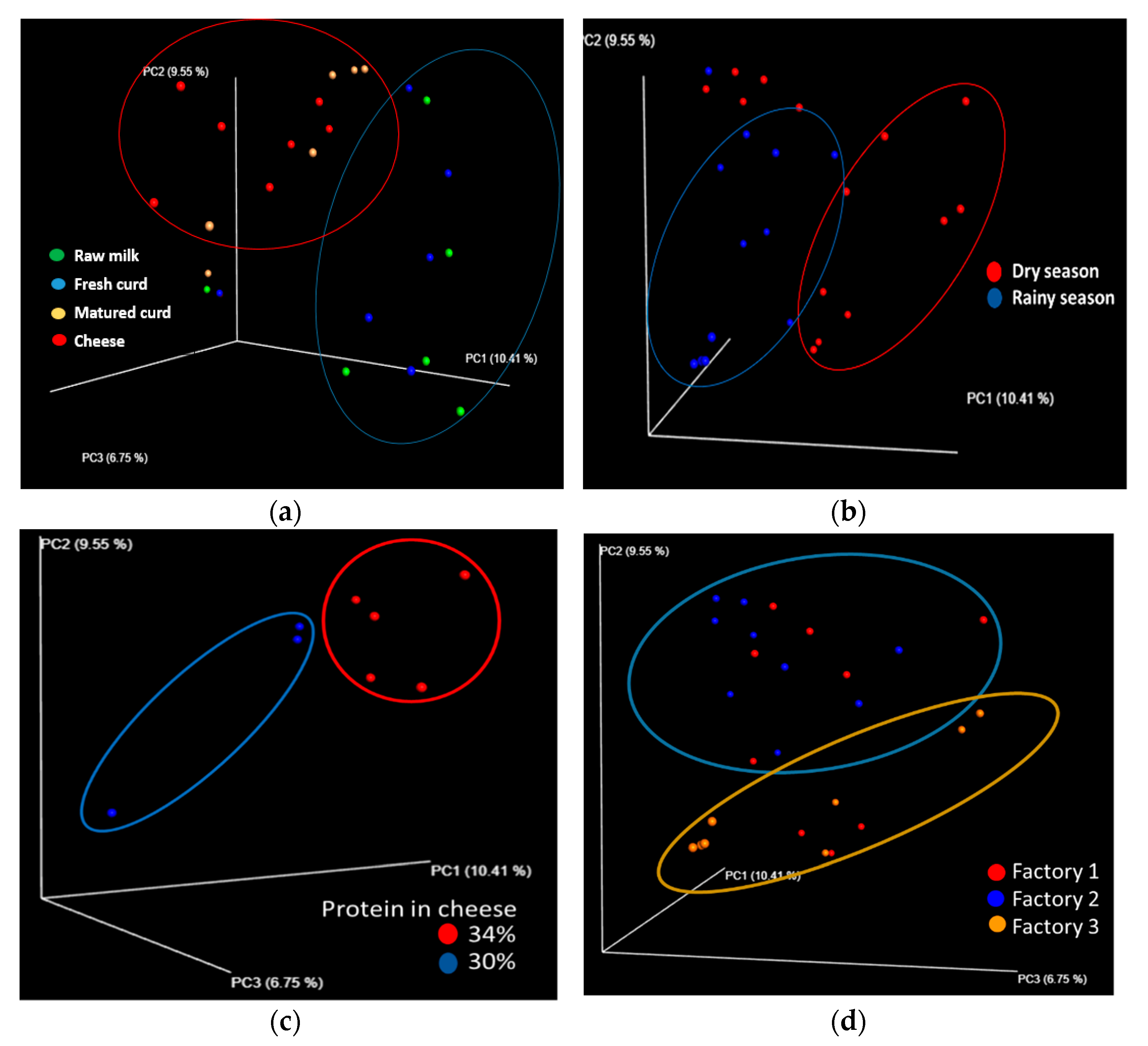
3.4. Factors Driving Changes in the Bacterial Community of Artisanal Adobera Cheese
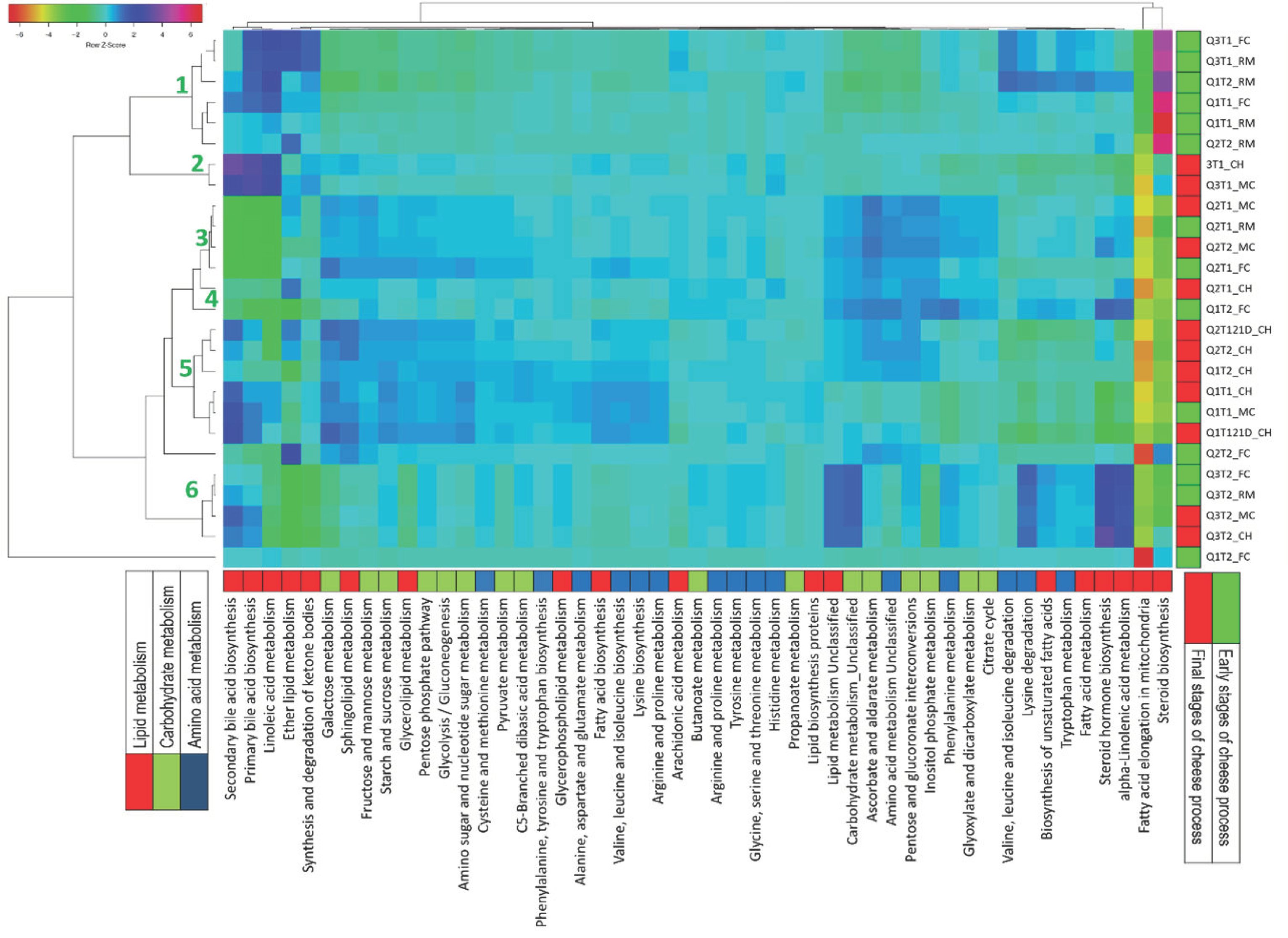
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef]
- Macori, G.; Cotter, P.D. Novel insights into the microbiology of fermented dairy foods. Curr. Opin. Biotechnol. 2018, 49, 172–178. [Google Scholar] [CrossRef]
- Licitra, G.; Caccamo, M.; Lortal, S. Artisanal Products Made with Raw Milk. In Raw Milk; Nero, L.A., Fernandes de Carvalho, A., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 175–221. [Google Scholar]
- Frank, J.F. Milk and dairy products. In Food Microbiology—Fundamental and Frontiers; Doyle, P., Beuchat, R., Montville, J., Eds.; ASM Press: Herndon, VA, USA, 1997. [Google Scholar]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. Molecular approaches to analysing the microbial composition of raw milk and raw milk cheese. Int. J. Food Microbiol. 2011, 150, 81–94. [Google Scholar] [CrossRef]
- Coppola, S.; Blaiotta, G.; Ercolini, D. Dairy products. In Molecular Techniques in the Microbial Ecology of Fermented Foods; Cocolin, L., Ercolini, D., Eds.; Springer: New York, NY, USA, 2008; pp. 31–90. [Google Scholar]
- Lavezzo, E.; Barzon, L.; Toppo, S.; Palu, G. Third generation sequencing technologies applied to diagnostic microbiology: Benefits and challenges in applications and data analysis. Expert Rev. Mol. Diagn. 2016, 16, 1011–1023. [Google Scholar] [CrossRef]
- Olsen, G.J.; Woese, C.R. Ribosomal RNA: A key to phylogeny. FASEB J. 1993, 7, 113–123. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley & Sons: New York, NY, USA, 1991; pp. 115–176. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Alegría, Á.; Szczesny, P.; Mayo, B.; Bardowski, J.; Kowalczyk, M. Biodiversity in Oscypek, a Traditional Polish Cheese, Determined by Culture-Dependent and -Independent Approaches. Appl. Environ. Microbiol. 2012, 78, 1890–1898. [Google Scholar] [CrossRef] [Green Version]
- Aldrete-Tapia, A.; Escobar-Ramírez, M.C.; Tamplin, M.L.; Hernández-Iturriaga, M. High-throughput sequencing of microbial communities in Poro cheese, an artisanal Mexican cheese. Food Microbiol. 2014, 44, 136–141. [Google Scholar] [CrossRef]
- De Filippis, F.; La Storia, A.; Stellato, G.; Gatti, M.; Ercolini, D. A selected core microbiome-e drives the early stages of three popular Italian cheese manufactures. PLoS ONE 2014, 9, e89680. [Google Scholar] [CrossRef] [PubMed]
- De Pasquale, I.; Di Cagno, R.; Buchin, S.; De Angelis, M.; Gobbetti, M. Microbial Ecology Dynamics Reveal a Succession in the Core Microbiota Involved in the Ripening of Pasta Filata Caciocavallo Pugliese Cheese. Appl. Environ. Microbiol. 2014, 80, 6243–6255. [Google Scholar] [CrossRef] [Green Version]
- Galimberti, A.; Bruno, A.; Mezzasalma, V.; De Mattia, F.; Bruni, I.; Labra, M. Emerging DNA-based technologies to characterize food ecosystems. Food Res. Int. 2015, 69, 424–433. [Google Scholar] [CrossRef]
- Salazar, J.K.; Carstens, C.K.; Ramachandran, P.; Shazer, A.G.; Narula, S.S.; Reed, E.; Ottesen, A.; Schill, K.M. Metagenomics of pasteurized and unpasteurized gouda cheese using targeted 16S rRNA sequencing. BMC Microbiol. 2018, 18, 189. [Google Scholar] [CrossRef] [PubMed]
- Dalmasso, A.; Rio, M.D.L.D.S.D.; Civera, T.; Pattono, D.; Cardazzo, B.; Bottero, M.T. Characterization of microbiota in Plaisentif cheese by high-throughput sequencing. LWT 2016, 69, 490–496. [Google Scholar] [CrossRef]
- Zheng, X.; Liu, F.; Shi, X.; Wang, B.; Li, K.; Li, B.; Zhuge, B. Dynamic correlations between microbiota succession and flavor development involved in the ripening of Kazak artisanal cheese. Food Res. Int. 2018, 105, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Grass, R.J.F.; Cesín, V.A. Situación actual y retrospectiva de los quesos genuinos de Chiautla de Tapia, México. Agric. Soc. Desarro. 2014, 11, 201–221. [Google Scholar] [CrossRef] [Green Version]
- Agudelo-López, M. Abraham Villegas de Gante, Fernando Cervantes Escoto, Alfredo Cesín Vargas, Angélica Espinoza Ortega, Arturo Hernández Montes, Armando Santos Moreno y Ángel Roberto Martínez Campos. 2014. Atlas de los quesos mexicanos genuinos. Agric. Soc. Desarro. 2015, 12, 257–260. [Google Scholar] [CrossRef]
- Cervantes-Escoto, F.; Villegas de Gante, A.; Cesín, V.J.A.; Espinoza, O.A. Los Quesos Mexicanos Genuinos/Genuine Mexican Cheeses: Patrimonio Cultural que Debe Rescatarse, 1st ed.; Mundi Prensa México: México City, México, 2008. [Google Scholar]
- González-Córdova, A.F.; Yescas, C.; Ortiz-Estrada, Á.M.; Alcaraz, M.D.L.; Ángeles, D.L.R.; Hernández-Mendoza, A.; Vallejo-Cordoba, B. Invited review: Artisanal Mexican cheeses. J. Dairy Sci. 2016, 99, 3250–3262. [Google Scholar] [CrossRef] [Green Version]
- Aldrete-Tapia, A.; Escobar-Ramírez, C.M.; Tamplin, M.L.; Hernández-Iturriaga, M. Characterization of Bacterial Communities in Mexican Artisanal Raw Milk “Bola de Ocosingo” Cheese by High-Throughput Sequencing. Front. Microbiol. 2018, 9, 2598. [Google Scholar] [CrossRef]
- Ruvalcaba-Gómez, J.M.; Ruiz-Espinosa, H.; Arteaga-Garibay, R.I.; Rojas-López, M.; Amador-Espejo, G.G.; Anaya-Esparza, L.M.; Delgado-Macuil, R.J. Texture, physicochemical and sensory properties of artisanal Adobera cheese from Los Altos de Jalisco, a genuine Mexican cheese. Int. J. Dairy. Technol. 2020, 73, 411–420. [Google Scholar] [CrossRef]
- Murugesan, S.; Reyes-Mata, M.P.; Nirmalkar, K.; Chavez-Carbajal, A.; Juárez-Hernández, J.I.; Torres-Gómez, R.E.; Piña-Escobedo, A.; Maya, O.; Hoyo-Vadillo, C.; Ramos-Ramírez, E.G.; et al. Profiling of bacterial and fungal communities of Mexican cheeses by high throughput DNA sequencing. Food Res. Int. 2018, 113, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Vega, H.; Arteaga-Garibay, R.I.; Maya-Lucas, O.; Gómez-Rodríguez, V.M.; Chávez-Díaz, I.F.; Ruvalcaba-Gómez, J.M.; Heredia-Nava, D.; Loperena-Martínez, R.; Zelaya-Molina, L. The Bacterial Community Associated with the Amarillo Zamorano Maize (Zea mays) Landrace Silage Process. Microorganisms 2020, 8, 1503. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. GigaScience 2013, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Moreno, C.E. Métodos Para Medir la Biodiversidad, 1st ed.; Manuales y Tesis SEA: Zaragoza, España, 2001; pp. 20–60. [Google Scholar]
- Parente, E.; Cocolin, L.; De Filippis, F.; Zotta, T.; Ferrocino, I.; O’Sullivan, O.; Neviani, E.; De Angelis, M.; Cotter, P.D.; Ercolini, D. FoodMicrobionet: A database for the visualisation and exploration of food bacterial communities based on network analysis. Int. J. Food Microbiol. 2016, 219, 28–37. [Google Scholar] [CrossRef]
- Jagadeesan, B.; Gerner-Smidt, P.; Allard, M.W.; Leuillet, S.; Winkler, A.; Xiao, Y.; Chaffron, S.; Van Der Vossen, J.; Tang, S.; Katase, M.; et al. The use of next generation sequencing for improving food safety: Translation into practice. Food Microbiol. 2019, 79, 96–115. [Google Scholar] [CrossRef]
- Loman, N.J.; Constantinidou, C.; Christner, M.; Rohde, H.; Chan, J.Z.M.; Quick, J.; Weir, J.C.; Quince, C.; Smith, G.P.; Betley, J.R.; et al. A Culture-Independent Sequence-Based Metagenomics Approach to the Investigation of an Outbreak of Shiga-Toxigenic Escherichia coli O104:H4. JAMA 2013, 309, 1502–1510. [Google Scholar] [CrossRef]
- Huang, A.D.; Luo, C.; Pena-Gonzalez, A.; Weigand, M.R.; Tarr, C.L.; Konstantinidis, K.T. Metagenomics of Two Severe Foodborne Outbreaks Provides Diagnostic Signatures and Signs of Coinfection Not Attainable by Traditional Methods. Appl. Environ. Microbiol. 2016, 83, e02577-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugli, G.A.; Duranti, S.; Albert, K.; Mancabelli, L.; Napoli, S.; Viappiani, A.; Anzalone, R.; Longhi, G.; Milani, C.; Turroni, F.; et al. Unveiling Genomic Diversity among Members of the Species Bifidobacterium pseudolongum, a Widely Distributed Gut Commensal of the Animal Kingdom. Appl. Environ. Microbiol. 2019, 85, e03065-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, V.D.S.; Carlot, M.; Pakroo, S.; Tarrah, A.; Lombardi, A.; Santiago, H.; Corich, V.; Giacomini, A. Comparative evaluation of cheese whey microbial composition from four Italian cheese factories by viable counts and 16S rRNA gene amplicon sequencing. Int. Dairy J. 2020, 104, 104656. [Google Scholar] [CrossRef]
- Escobar-Zepeda, A.; Sanchez-Flores, A.; Baruch, M.Q. Metagenomic analysis of a Mexican ripened cheese reveals a unique complex microbiota. Food Microbiol. 2016, 57, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, B.A.; Cabral, L.; Noronha, M.F.; Baptista, R.C.; Nascimento, H.M.; Sant’Ana, A.S. Amplicon sequencing reveals the bacterial diversity in milk, dairy premises and Serra da Canastra artisanal cheeses produced by three different farms. Food Microbiol. 2020, 89, 103453. [Google Scholar] [CrossRef] [PubMed]
- Ceniti, C.; Britti, M.; Santoro, A.M.L.; Musarella, R.; Ciambrone, L.; Casalinuovo, F.; Costanzo, N. Phenotypic Antimicrobial Resistance Profile of Isolates Causing Clinical Mastitis in Dairy Animals. Ital. J. Food Saf. 2017, 6, 6612. [Google Scholar] [CrossRef] [Green Version]
- Tilocca, B.; Costanzo, N.; Morittu, V.M.; Spina, A.A.; Soggiu, A.; Britti, D.; Roncada, P.; Piras, C. Milk microbiota: Characterization methods and role in cheese production. J. Proteom. 2020, 210, 103534. [Google Scholar] [CrossRef]
- Masoud, W.; Vogensen, F.K.; Lillevang, S.; Abu Al-Soud, W.; Sørensen, S.J.; Jakobsen, M. The fate of indigenous microbiota, starter cultures, Escherichia coli, Listeria innocua and Staphylococcus aureus in Danish raw milk and cheeses determined by pyrosequencing and quantitative real time (qRT)-PCR. Int. J. Food Microbiol. 2012, 153, 192–202. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Mancini, L.; Fox, P.F. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening. Trends Food Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Blaya, J.; Barzideh, Z.; Lapointe, G. Symposium review: Interaction of starter cultures and nonstarter lactic acid bacteria in the cheese environment. J. Dairy Sci. 2018, 101, 3611–3629. [Google Scholar] [CrossRef]
- Settanni, L.; Di Grigoli, A.; Tornambé, G.; Bellina, V.; Francesca, N.; Moschetti, G.; Bonanno, A. Persistence of wild Streptococcus thermophilus strains on wooden vat and during the manufacture of a traditional Caciocavallo type cheese. Int. J. Food Microbiol. 2012, 155, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M.; Calasso, M. Streptococcus: Introduction. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: London, UK, 2014; pp. 535–553. [Google Scholar]
- Batt, C.A. Lactococcus: Introduction. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: London, UK, 2014; pp. 439–441. [Google Scholar]
- Endo, A.; Maeno, S.; Liu, S.Q. Lactic Acid Bacteria: Leuconostoc spp. In Reference Module in Food Science; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 1–6. [Google Scholar]
- Giraffa, G.; De Fernando, G.G. Lactic Acid Bacteria: Enterococcus in Milk and Dairy Products. In Reference Module in Food Science; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 1–7. [Google Scholar]
- Encyclopedia of Food Microbiology. In Encyclopedia of Food Microbiology; Elsevier BV: Amsterdam, The Netherlands, 2014; pp. 11–17.
- Rafei, R.; Hamze, M.; Pailhoriès, H.; Eveillard, M.; Marsollier, L.; Joly-Guillou, M.-L.; Dabboussi, F.; Kempf, M. Extrahuman Epidemiology of Acinetobacter baumannii in Lebanon. Appl. Environ. Microbiol. 2015, 81, 2359–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atrouni, A.E.; Ejoly-Guillou, M.-L.; Ehamze, M.; Kempf, M. Reservoirs of Non-baumannii Acinetobacter Species. Front. Microbiol. 2016, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yunita, D.; Dodd, C.E. Microbial community dynamics of a blue-veined raw milk cheese from the United Kingdom. J. Dairy Sci. 2018, 101, 4923–4935. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, H.A. Spoilage potential of Pseudomonas spp. isolated from domiati cheese. Assiut Vet. Med. J. 2015, 61, 18–23. [Google Scholar]
- Nucera, D.M.; Lomonaco, S.; Morra, P.; Ortoffi, M.; Giaccone, D.; Grassi, M.A. Dissemination and Persistence of Pseudomonas spp. in Small-Scale Dairy Farms. Ital. J. Food Saf. 2016, 5, 5652. [Google Scholar] [CrossRef] [Green Version]
- Samaržija, D.; Zamberlin, Š. Psychrotrophic Bacteria: Pseudomonas spp. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Singh, N.; Anand, S. Enterobacteriaceae. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Torres-Vitela, M.R.; Mendoza-Bernardo, M.; Castro-Rosas, J.; Gómez-Aldapa, C.A.; Garay-Martinez, L.E.; Navarro-Hidalgo, V.; Villarruel-López, A. Incidence of Salmonella, Listeria monocytogenes, Escherichia coli O157:H7, and Staphylococcal Enterotoxin in Two Types of Mexican Fresh Cheeses. J. Food Prot. 2012, 75, 79–84. [Google Scholar] [CrossRef]
- O’Brien, B.; Guinee, T. Seasonal Effects on Processing Properties of Cows’ Milk. In Reference Module in Food Science; Elsevier BV: Amsterdam, The Netherlands, 2016; pp. 598–606. [Google Scholar]
- Carminati, D.; Meucci, A.; Tidona, F.; Zago, M.; Giraffa, G. Multifunctional Lactic Acid Bacteria Cultures to Improve Quality and Nutritional Benefits in Dairy Products. In Advances in Food Biotechnology; Wiley: Hoboken, NJ, USA, 2015; pp. 263–276. [Google Scholar]
- Tidona, F.; Francolino, S.; Ghiglietti, R.; Locci, F.; Carminati, D.; LaForce, P.; Giraffa, G. Characterization and pre-industrial validation of Streptococcus thermophilus strains to be used as starter cultures for Crescenza, an Italian soft cheese. Food Microbiol. 2020, 92, 103599. [Google Scholar] [CrossRef]
- Caro, I.; Quinto, E.J.; Fuentes, L.; Alessandria, V.; Cocolin, L.; Redondo-Del-Río, M.; Mayo, B.; Flórez, A.B.; Mateo, J. Characterization of Lactococcus strains isolated from artisanal Oaxaca cheese. LWT 2020, 122, 109041. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; Reyes-Gavilán, C.G.D.L.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, A.D.M.D.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Factories 2017, 16, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Moyano, S.; Dos Santos, M.T.P.G.; Galván, A.I.; Merchán, A.V.; González, E.; Córdoba, M.D.G.; Benito, M.J. Screening of autochthonous lactic acid bacteria strains from artisanal soft cheese: Probiotic characteristics and prebiotic metabolism. LWT 2019, 114, 108388. [Google Scholar] [CrossRef]
- Brooks, J.; Martinez, B.; Stratton, J.; Bianchini, A.; Krokstrom, R.; Hutkins, R. Survey of raw milk cheeses for microbiological quality and prevalence of foodborne pathogens. Food Microbiol. 2012, 31, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.; Lee, S.; Choi, K.-H. Microbial benefits and risks of raw milk cheese. Food Control. 2016, 63, 201–215. [Google Scholar] [CrossRef]
- Silvetti, T.; Capra, E.; Morandi, S.; Cremonesi, P.; Decimo, M.; Gavazzi, F.; Giannico, R.; Cattaneo, S.; Brasca, M. Microbial population profile during ripening of Protected Designation of Origin (PDO) Silter cheese, produced with and without autochthonous starter culture. LWT 2017, 84, 821–831. [Google Scholar] [CrossRef]
- De Pasquale, I.; Di Cagno, R.; Buchin, S.; De Angelis, M.; Gobbetti, M. Use of autochthonous mesophilic lactic acid bacteria as starter cultures for making Pecorino Crotonese cheese: Effect on compositional, microbiological and biochemical attributes. Food Res. Int. 2019, 116, 1344–1356. [Google Scholar] [CrossRef]
- Ferrari, I.D.S.; De Souza, J.V.; Ramos, C.L.; Da Costa, M.M.; Schwan, R.F.; Dias, F.S. Selection of autochthonous lactic acid bacteria from goat dairies and their addition to evaluate the inhibition of Salmonella typhi in artisanal cheese. Food Microbiol. 2016, 60, 29–38. [Google Scholar] [CrossRef]
- Scatassa, M.L.; Gaglio, R.; Cardamone, C.; Macaluso, G.; Arcuri, L.; Todaro, M.; Mancuso, I. Anti-Listeria Activity of Lactic Acid Bacteria in Two Traditional Sicilian Cheeses. Ital. J. Food Saf. 2017, 6, 6191. [Google Scholar] [CrossRef] [Green Version]
- Castro, R.; De Oliveira, A.P.D.; Souza, E.; Correia, T.M.A.; De Souza, J.V.; Dias, F.S. Lactic Acid Bacteria as Biological Control of Staphylococcus aureus in Coalho Goat Cheese. Food Technol. Biotechnol. 2018, 56, 431–440. [Google Scholar] [CrossRef]
- Ruvalcaba-Gómez, J.M.; Arteaga-Garibay, R.I.; Domínguez-Araujo, G.; Galindo-Barboza, A.J.; Salazar-Gutiérrez, G.; Martínez-Peña, M.D.; Delgado-Macuil, R.J. Uso de bacterias ácido lácticas para descontaminación de estiércol porcino mediante ensilaje experimental. Rev. Int. Cont. Amb. 2019, 35, 247–257. [Google Scholar] [CrossRef]
- Settanni, L.; Moschetti, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiol. 2010, 27, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Torres-Llanez, M.; González-Córdova, A.; Hernández-Mendoza, A.; Garcia, H.; Vallejo-Cordoba, B. Angiotensin-converting enzyme inhibitory activity in Mexican Fresco cheese. J. Dairy Sci. 2011, 94, 3794–3800. [Google Scholar] [CrossRef] [PubMed]
- Ahmadova, A.; Todorov, S.D.; Choiset, Y.; Rabesona, H.; Zadi, T.M.; Kuliyev, A.; Franco, B.D.G.D.M.; Chobert, J.-M.; Haertlé, T. Evaluation of antimicrobial activity, probiotic properties and safety of wild strain Enterococcus faecium AQ71 isolated from Azerbaijani Motal cheese. Food Control 2013, 30, 631–641. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Ferrocino, I.; Botta, C.; Ercolini, D.; Cocolin, L.; Rantsiou, K. Probiotic potential of a Lactobacillus rhamnosus cheese isolate and its effect on the fecal microbiota of healthy volunteers. Food Res. Int. 2019, 119, 305–314. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
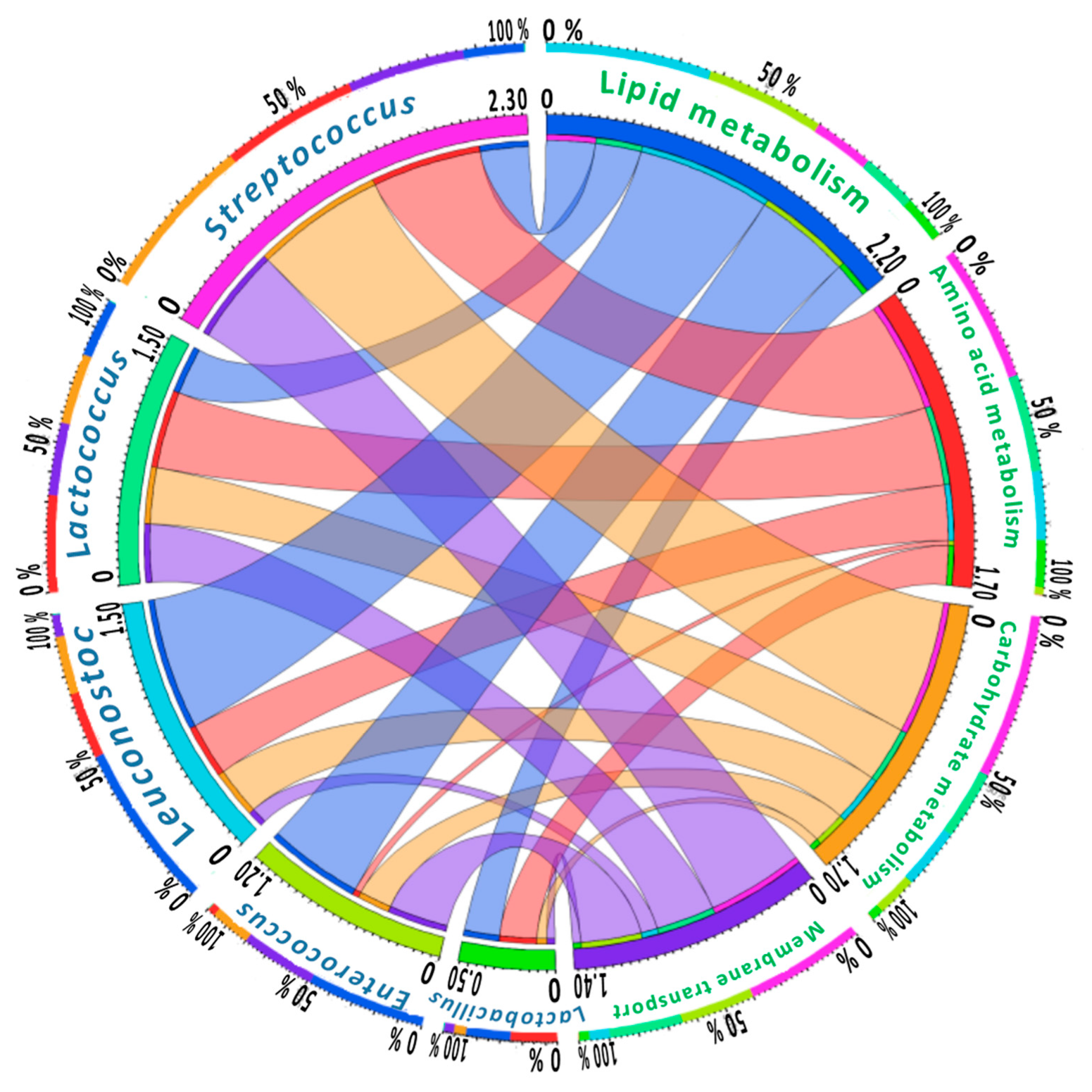{kind=link}
| Cheese | Season | Sample | Valid Reads | Chao1 | Shannon | Simpson | Simpson Reciprocal |
|---|---|---|---|---|---|---|---|
| 1 | Dry | Raw Milk | 112,441 | 3954.71 | 6.539 | 0.948 | 19.23 |
| Fresh curd | 41,138 | 4143.00 | 6.821 | 0.968 | 31.09 | ||
| Matured curd | 299,377 | 4304.96 | 6.128 | 0.967 | 29.95 | ||
| Cheese | 480,795 | 4525.90 | 6.105 | 0.965 | 28.78 | ||
| Rainy | Raw Milk | 69,215 | 3049.56 | 6.199 | 0.962 | 34.76 | |
| Fresh curd | 336,678 | 4719.12 | 6.250 | 0.955 | 22.20 | ||
| Matured curd | 90,918 | 5156.78 | 6.957 | 0.979 | 47.70 | ||
| Cheese | 8674 | 24.20 | 3.102 | 0.878 | 8.26 | ||
| 2 | Dry | Raw Milk | 516,917 | 5170.39 | 6.687 | 0.978 | 45.62 |
| Fresh curd | 293,445 | 4477.34 | 6.202 | 0.965 | 28.50 | ||
| Matured curd | 183,288 | 4293.72 | 6.417 | 0.947 | 38.15 | ||
| Cheese | 106,058 | 5357.36 | 6.906 | 0.974 | 38.26 | ||
| Rainy | Raw Milk | 50,256 | 3774.49 | 7.071 | 0.984 | 62.72 | |
| Fresh curd | 38,019 | 3350.41 | 6.833 | 0.981 | 51.78 | ||
| Matured curd | 99,925 | 4379.79 | 6.545 | 0.975 | 39.50 | ||
| Cheese | 75,107 | 4554.86 | 6.744 | 0.977 | 43.23 | ||
| 3 | Dry | Raw Milk | 119,219 | 4075.30 | 6.590 | 0.971 | 33.94 |
| Fresh curd | 67,669 | 4417.30 | 7.011 | 0.981 | 51.32 | ||
| Matured curd | 98,183 | 6429.53 | 7.041 | 0.972 | 35.32 | ||
| Cheese | 61,937 | 5861.76 | 6.970 | 0.971 | 34.46 | ||
| Rainy | Raw Milk | 41,368 | 1741.43 | 5.351 | 0.949 | 19.57 | |
| Fresh curd | 58,128 | 2746.97 | 5.639 | 0.950 | 19.92 | ||
| Matured curd | 17,047 | 1252.38 | 5.873 | 0.956 | 22.55 | ||
| Cheese | 10,540 | 1196.24 | 5.919 | 0.958 | 23.54 | ||
| 21 d post-elaboration | |||||||
| 1 | Dry | Cheese | 92,545 | 4453.98 | 6.179 | 0.968 | 31.25 |
| 2 | Dry | Cheese | 67,213 | 4559.55 | 6.636 | 0.974 | 38.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruvalcaba-Gómez, J.M.; Delgado-Macuil, R.J.; Zelaya-Molina, L.X.; Maya-Lucas, O.; Ruesga-Gutiérrez, E.; Anaya-Esparza, L.M.; Villagrán-de la Mora, Z.; López-de la Mora, D.A.; Arteaga-Garibay, R.I. Bacterial Succession through the Artisanal Process and Seasonal Effects Defining Bacterial Communities of Raw-Milk Adobera Cheese Revealed by High Throughput DNA Sequencing. Microorganisms 2021, 9, 24. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010024
Ruvalcaba-Gómez JM, Delgado-Macuil RJ, Zelaya-Molina LX, Maya-Lucas O, Ruesga-Gutiérrez E, Anaya-Esparza LM, Villagrán-de la Mora Z, López-de la Mora DA, Arteaga-Garibay RI. Bacterial Succession through the Artisanal Process and Seasonal Effects Defining Bacterial Communities of Raw-Milk Adobera Cheese Revealed by High Throughput DNA Sequencing. Microorganisms. 2021; 9(1):24. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010024
Chicago/Turabian StyleRuvalcaba-Gómez, José M., Raúl J. Delgado-Macuil, Lily X. Zelaya-Molina, Otoniel Maya-Lucas, Edmundo Ruesga-Gutiérrez, Luis M. Anaya-Esparza, Zuamí Villagrán-de la Mora, David A. López-de la Mora, and Ramón I. Arteaga-Garibay. 2021. "Bacterial Succession through the Artisanal Process and Seasonal Effects Defining Bacterial Communities of Raw-Milk Adobera Cheese Revealed by High Throughput DNA Sequencing" Microorganisms 9, no. 1: 24. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010024