Osmotic Adaptation and Compatible Solute Biosynthesis of Phototrophic Bacteria as Revealed from Genome Analyses


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation and DNA Extraction
2.2. Sequencing and Assembly
2.3. Genome Annotation and Submission to GenBank
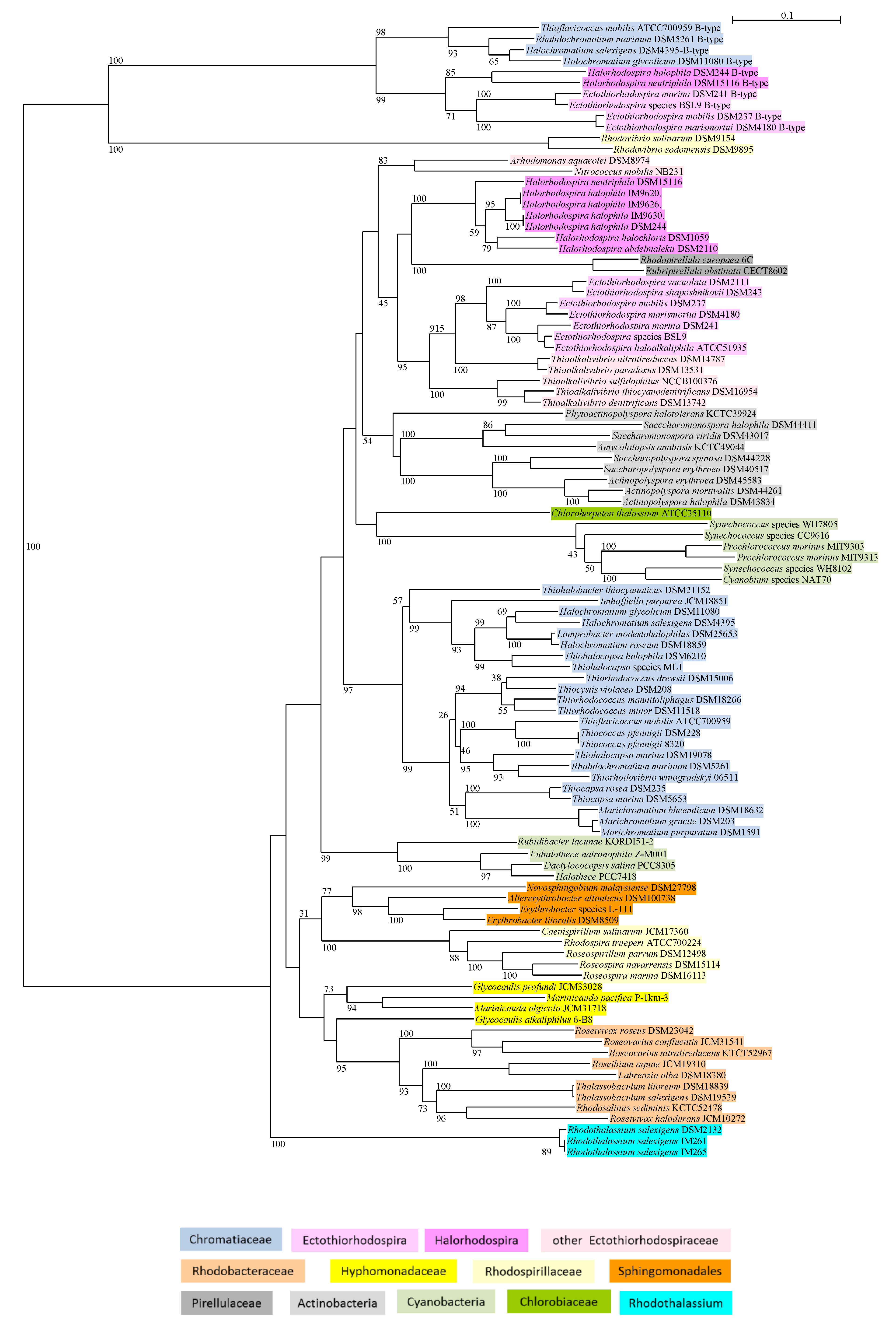
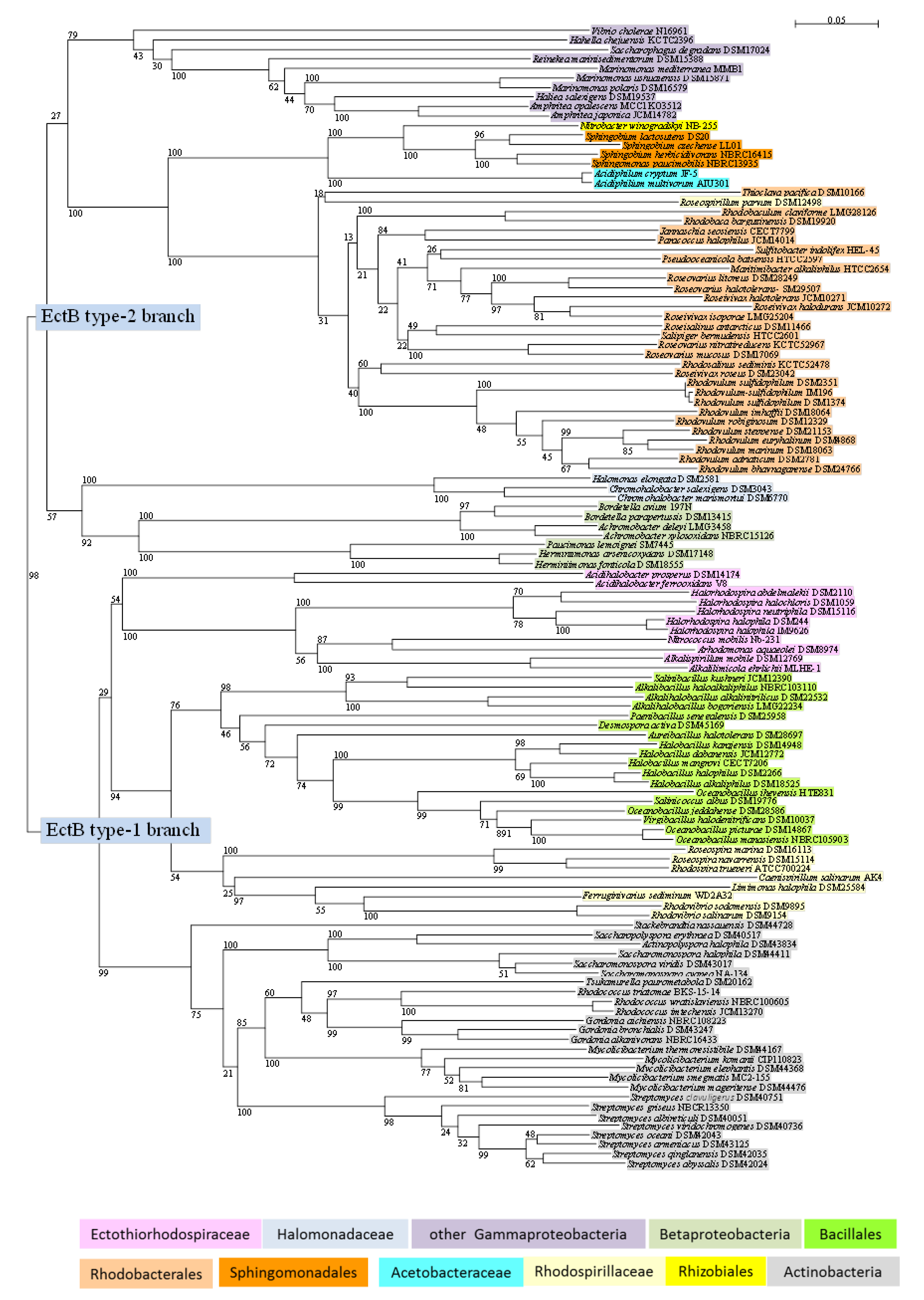
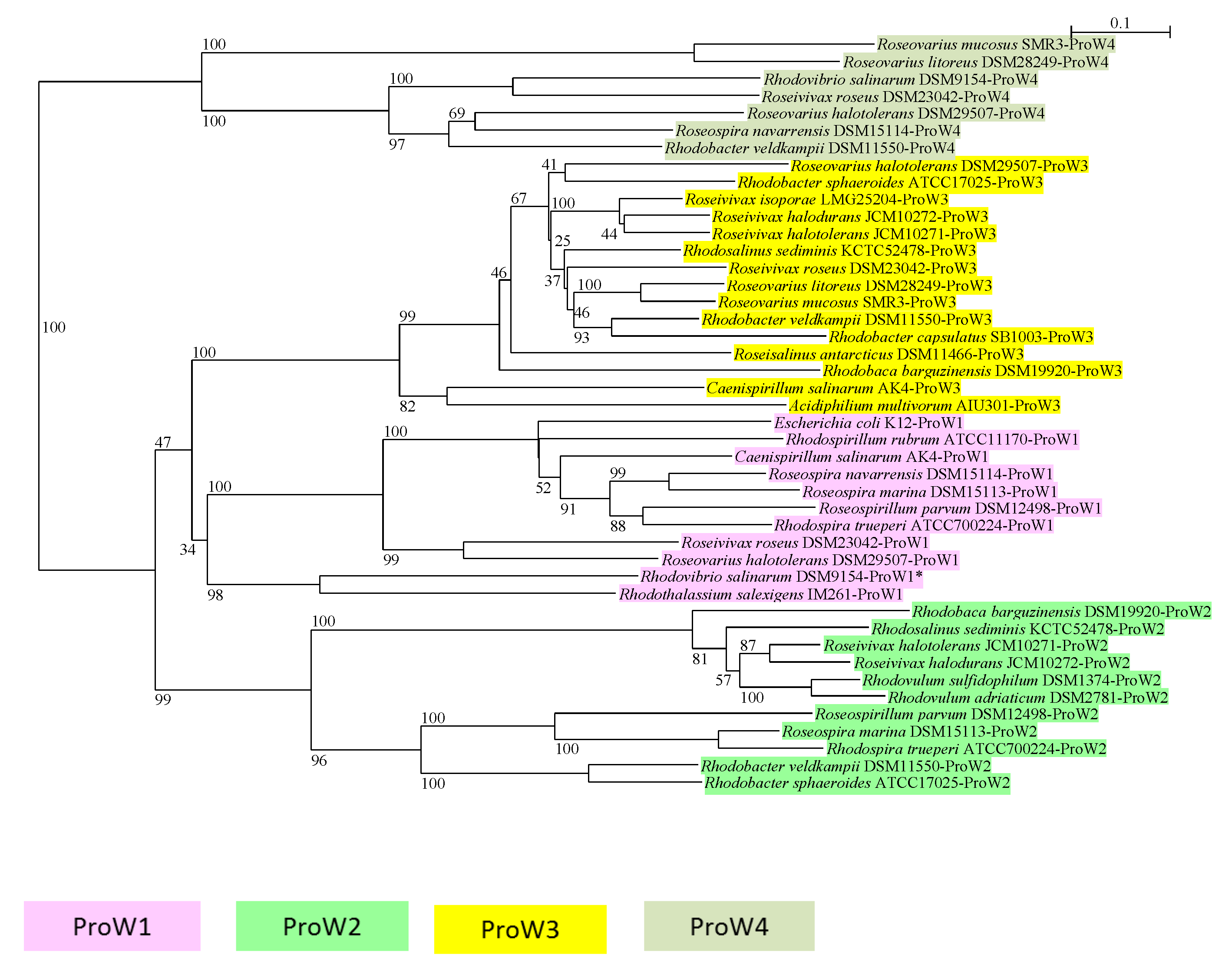
2.4. Phylogenetic Sequence Analyses
3. Results and Discussion
3.1. Osmotic Adaptation in Phototrophic Bacteria
3.1.1. Cyanobacteria
3.1.2. Chlorobiaceae
3.1.3. Chloroflexi
3.1.4. Marine Rhizobiales
3.1.5. Marine and Halotolerant Rhodobacteraceae
3.1.6. Acetobacteraceae
3.1.7. Marine and Halophilic Rhodospirillaceae
3.1.8. Halophilic Rhodothalassium salexigens
3.1.9. Chromatiaceae
3.1.10. Ectothiorhodospiraceae
3.2. Phylogeny of Glycine-Methyltransferase GMT
3.2.1. Chromatiaceae
3.2.2. Ectothiorhodospiraceae
3.2.3. Cyanobacteria
3.2.4. Alphaproteobacteria
- Rhodospirillaceae, including Caenispirillum salinarum, Rhodospira trueperi, Roseospira navarrensis, Roseospira marina, and Roseospirillum parvum (but not the Rhodovibrio species) have a fused GMT/DMT gene of the methyltransferases included in a small cluster with S-adenosylmethionine synthase (MAT).
- Rhodothalassium salexigens, which represents the most distant line to all other Alphaproteobacteria, has a small gene cluster with just the two methyltransferases.
- Sphingomonadales include Novosphingobium malayensis, Erythrobacter litoralis, Altererythrobacter atlanticus; in Erythrobacter litoralis just the two methyltransferases form a small gene cluster.
- Hyphomonadaceae include the Glycocaulis and Marinicauda species with a small gene cluster of the two methyltransferases only.
- Rhodobacteraceae included in the study, as indicated in Figure 1, form a subbranch together with Thalassobaculum litoreum and Thalassobaculum salexigens (according to 16S rRNA phylogeny forming a branch with the Oceanobaculum species at an almost equal distance to the Rhodobacteraceae and Rhodospirillaceae species; data not shown). A fused GMT/DMT gene is associated with the MAT gene in Roseivivax halodurans, with the MS-MAT genes in Rhodosalinus sediminis, and with the MTHFR-MS-MAT-SAHase genes in Roseivivax roseus and Roseivarius nitratireducens (Table 2).
3.2.5. Actinobacteria
3.2.6. Chlorobiaceae
3.2.7. Pirellulaceae
3.3. Phylogeny of EctB
- The Ectothiorhodospiraceae form a branch with representatives of the genera Halorhodospira, Alkalilimnicola, Alkalispirillum, Nitrococcus, Arhodomonas, and Acidihalobacter, but lack species of Ectothiorhodospira, Thiorhodospira, and Ectothiorhodosinus, in which ectoine biosynthesis is absent. A cluster of the ectABC genes that lacks the regulatory gene is found in the extremely halophilic Halorhodospira species.
- The marine and halophilic Rhodospirillaceae also have an ectABC gene cluster and form deeply branching separate lineages with Rhodospira trueperi and the Roseospira species in one, Rhodovibrio sodomensis and Rhodovibrio salinarum and the non-phototrophic Ferruginivarius sediminum and Limimonas halophila in another, and Caenispirillum salinarum in a third lineage (Figure 2).
- Different lineages of the Bacillales branch are represented by species of the genera Halobacillus, of Oceanobacillus (including Salinicoccus albus (Staphylococcaceae) and Virgibacillus halodenitrificans), of Salinibacillus, Alkalibacillus and Alkalihalobacillus. Distinct separate lines of Aureibacillus halotolerans (Bacillaceae), Paenibacillus senegaliensis (Paenibacillaceae) and Desmospora activa (Thermoactinomycetaceae) are found (Figure 2).
- The branch of Actinobacteria shows distinct subbranches of Streptomyces, Mycolicibacterium, Gordonia, and Rhodococcus (associated with the Tsukamurella paurometabola) species, and of Saccharomonospora, Actinopolyspora, and Saccharopolyspora species (Figure 2). Distinct from these and as an outsider of the group is Stackebrandtia nassauensis. Common among many of the Actinobacteria is the ability to form hydroxyectoine, as demonstrated here by genome analysis of Mycolicibacterium thermoresistibile, and BLAST search with the EctD sequences from these bacteria (data not shown).
3.4. Betaine Synthesis from Choline—Distribution of Bet Genes in Phototrophic Bacteria
3.5. Transport System for Uptake of Glycine Betaine and Choline
3.6. Evolutionary Considerations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Imhoff, J.F. Minireview—True marine and halophilic anoxygenic phototrophic bacteria. Arch. Microbiol. 2001, 176, 243–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhoff, J.F. Diversity of anaerobic anoxygenic phototrophic purple bacteria. In Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Hallenbeck, P.C., Ed.; Springer: Cham, Switzerland, 2017; pp. 47–85. [Google Scholar]
- Brown, A.D. Microbial water stress. Bacteriol. Rev. 1976, 40, 803–846. [Google Scholar] [CrossRef] [PubMed]
- Galinski, E.A.; Trüper, H.G. Betaine, a compatible solute in the extremely halophilic phototrophic bacterium Ectothiorhodospira halochloris. FEMS Microbiol. Lett. 1982, 13, 357–360. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Rodriguez-Valera, F. Betaine is the main compatible solute of halophilic eubacteria. J. Bacteriol. 1984, 160, 478–479. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, J.F. Osmoregulation and compatible solutes in eubacteria. FEMS Microbiol. Rev. 1986, 39, 57–66. [Google Scholar] [CrossRef]
- Imhoff, J.F. Osmotic adaptation in halophilic and halotolerant microorganisms. In The Biology of Halophilic Bacteria; Vreeland, R.W., Hochstein, L.J., Eds.; The CRC Press: Boca Raton, FL, USA, 1993; pp. 211–253. [Google Scholar]
- Trüper, H.G.; Galinski, E.A. Biosynthesis and fate of compatible solutes in extremely halophilic phototrophic eubacteria. FEMS Microbiol. Rev. 1990, 75, 247–254. [Google Scholar] [CrossRef]
- Galinski, E.A.; Pfeiffer, H.-P.; Trüper, H.G. 1,4,5,6-Tetra-hydro-2-methyl-4-pyrimidinecarboxylic acid: A novel cyclic amino acid from halophilic phototrophic bacteria of the genus Ectothiorhodospira. Eur. J. Biochem. 1985, 149, 135–139. [Google Scholar] [CrossRef]
- Czech, L.; Hermann, L.; Stöveken, N.; Richter, A.A.; Höeppner, A.; Smits, S.H.J.; Heider, J.; Bremer, E. Role of the Extremolytes Ectoine and Hydroxyectoine as Stress Protectants and Nutrients: Genetics, Phylogenomics, Biochemistry, and Structural Analysis. Genes 2018, 9, 177. [Google Scholar] [CrossRef] [Green Version]
- Mackay, M.A.; Norton, R.S.; Borowitzka, L.J. Organic Osmoregulatory Solutes in Cyanobacteria. J. Gen. Microbiol. 1984, 130, 2177–2191. [Google Scholar] [CrossRef] [Green Version]
- Severin, J.; Wohlfarth, A.; Galinski, E.A. The predominant role of recently discovered tetrahydropyrimidines for the osmoadaptation of halophilic eubacteria. J. Gen. Microbiol. 1992, 138, 1629–1638. [Google Scholar] [CrossRef] [Green Version]
- Welsh, D.T.; Herbert, R.A. Identification of organic solutes accumulated by purple and green sulfur bacteria during osmotic stress using natural abundance 13C nuclear magnetic resonance spectroscopy. FEMS Micobiol. Ecol. 1993, 13, 145–150. [Google Scholar] [CrossRef]
- Nyyssölä, A.; Kerovuo, J.; Kaukinen, P.; Von Weymarn, N.; Reinikainen, T. Extreme Halophiles Synthesize Betaine from Glycine by Methylation. J. Biol. Chem. 2000, 275, 22196–22201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyyssölä, A.; Reinikainen, T.; Leisola, M. Characterization of Glycine Sarcosine N-Methyltransferase and Sarcosine Dimethylglycine N-Methyltransferase. Appl. Environ. Microbiol. 2001, 67, 2044–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waditee, R.; Tanaka, Y.; Aoki, K.; Hibino, T.; Jikuya, H.; Takano, J.; Takabe, T.; Takabe, T. Isolation and functional characterization of N-methyltransferases that catalyze betaine synthesis from glycine in a halotolerant photosynthetic organism Aphanothece halophytica. J. Biol. Chem. 2003, 278, 4932–4942. [Google Scholar] [CrossRef] [Green Version]
- Andresen, P.A.; Kaasen, I.; Styrvold, O.B.; Boulnois, G.; Strøm, A.R. Molecular cloning, physical mapping and expression of the bet genes governing the osmoregulatory choline-glycine betaine pathway of Escherichia coli. J. Gen. Microbiol. 1988, 134, 1737–1746. [Google Scholar] [CrossRef] [Green Version]
- Laloknam, S.; Tanaka, K.; Buaboocha, T.; Waditee, R.; Incharoensakdi, A.; Hibino, Y.; Tanaka, Y.; Takabe, T. Halotolerant cyanobacterium Aphanothece halophytica contains a betaine transporter active at alkaline pH and high salinity. Appl. Environ. Microbiol. 2006, 72, 6018–6026. [Google Scholar] [CrossRef] [Green Version]
- Peters, P.; Galinski, E.A.; Trüper, H.G. The biosynthesis of ectoine. FEMS Microbiol. Lett. 1990, 71, 157–162. [Google Scholar] [CrossRef]
- Ono, H.; Sawada, K.; Khunajakr, N.; Tao, T.; Yamamoto, M.; Hiramoto, M.; Shinmyo, A.; Takano, M.; Murooka, Y. Characterization of biosynthetic enzymes for ectoine as a compatible solute in a moderately halophilic eubacterium Halomonas elongata. J. Bacteriol. 1999, 181, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Galinski, E.A. Characterization of genes for the biosynthesis of the compatible solute ectoine from Marinococcus halophilus and osmoregulated expression in Escherichia coli. Microbiology 1997, 143, 1141–1149. [Google Scholar] [CrossRef] [Green Version]
- Inbar, L.; Labidot, A. The structure and biosynthesis of new tetrahydropyrimidine derivatives in actinomycin D producer Streptomyces parvulus. Use of 13C- and 15N-labeled L-glutamate and 13C and 15N NMR spectroscopy. J. Biol. Chem. 1988, 263, 16014–16022. [Google Scholar]
- Widderich, N.; Höppner, A.; Pittelkow, M.; Heider, J.; Smits, S.H.; Bremer, E. Biochemical properties of ectoine hydroxylase from extremophiles and their wide taxonomic distribution among microorganisms. PLoS ONE 2014, 9, e93809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stöveken, N.; Pittelkow, M.; Sinner, T.; Jensen, R.A.; Heider, J.; Bremer, E. A specialized aspartokinase enhances the biosynthesis of the osmoprotectants ectoine and hydroxyectoine in Pseudomonas stutzeri A1501. J. Bacteriol. 2011, 193, 4456–4468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reshetnikov, A.S.; Khmelenina, V.N.; Trotsenko, Y.A. Characterization of the ectoine biosynthesis genes of haloalkalotolerant obligate methanotroph “Methylomicrobium alcaliphilum 20Z”. Arch. Microbiol. 2006, 184, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Sleator, R.D.; Hill, C. Bacterial osmoadaptation: The role of osmolytes in bacterial stress and virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef] [Green Version]
- Kempf, B.; Bremer, E. Uptake and synthesis of compatible solutes as microbial stress response to high-osmolality environments. Arch. Microbiol. 1998, 170, 319–330. [Google Scholar] [CrossRef]
- Pfennig, N.; Trüper, H.G. Isolation of members of the families Chromatiaceae and Chlorobiaceae. In The Prokaryotes, a Handbook on Habitats, Isolation and Identification of Bacteria; Starr, M.P., Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: New York, NY, USA, 1981; pp. 279–289. [Google Scholar]
- Imhoff, J.F. The Chromatiaceae. In The Prokaryotes. A Handbook on the Biology of Bacteria, 3rd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 6, pp. 846–873. [Google Scholar]
- Imhoff, J.F. Anoxygenic phototrophic bacteria. In Methods in Aquatic Bacteriology; Austin, B., Ed.; John Wiley & Sons: Chichester, UK, 1988; pp. 207–240. [Google Scholar]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Neulinger, S.C. New insights into the metabolic potential of the phototrophic purple bacterium Rhodopila globiformis DSM 161T from its draft genome sequence and evidence for a vanadium-dependent nitrogenase. Arch. Microbiol. 2018, 200, 847–857. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Morgulis, A.; Gertz, E.M.; Schäffer, A.A.; Agarwala, R. A Fast and Symmetric DUST Implementation to Mask Low-Complexity DNA Sequences. J. Comput. Biol. 2006, 13, 1028–1040. [Google Scholar] [CrossRef]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A. Assembling genomes and mini-metagenomes from highly chimeric reads. In Research in Computational Molecular Biology; Deng, M., Jiang, R., Sun, F., Zhang, X., Eds.; Springer: Heidelberg, Germany, 2013; pp. 158–170. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Boetzer, M.; Henkel, C.V.; Jansen, H.J.; Butler, D.; Pirovano, W. Scaffolding pre-assembled contigs using SSPACE. Bioinformatics 2011, 27, 578–579. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Perrière, G.; Gouy, M. WWW-query: An on-line retrieval system for biological sequence banks. Biochimie 1996, 78, 364–369. [Google Scholar] [CrossRef]
- Reed, R.; Stewart, W.D.P. Osmotic adjustment and organic solute accumulation in unicellular cyanobacteria from freshwater and marine habitats. Mar. Biol. 1985, 88, 1–9. [Google Scholar] [CrossRef]
- Reed, R.; Borowitzka, L.J.; Mackay, M.A.; Chudek, J.A.; Foster, R.; Warr, S.C.; Moort, D.J.; Stewart, W.D.P. Organic solute accumulation in osmotically stressed cyanobacteria. FEMS Microbiol. Rev. 1986, 39, 51–56. [Google Scholar] [CrossRef]
- Klähn, S.; Hagemann, M. Compatible solute biosynthesis in cyanobacteria. Environ. Microbiol. 2011, 13, 551–562. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Neulinger, S.C. Phylogeny of anoxygenic photosynthesis based on protein sequences of the photosynthetic reaction center and of bacteriochlorophyll biosynthesis. Microorganisms 2019, 7, 576. [Google Scholar] [CrossRef] [Green Version]
- Imhoff, J.F.; Petri, R.; Süling, J. Reclassification of species of the spiral-shaped phototrophic purple nonsulfur bacteria of the alpha-proteobacteria: Description of the new genera Phaeospirillum gen. nov., Rhodovibrio gen. nov., Rhodothalassium gen. nov. and Roseospira gen. nov. as well as transfer of Rhodospirillum fulvum to Phaeospirillum fulvum comb. nov., of Rhodospirillum molischianum to Phaeospirillum molischianum comb. nov., of Rhodospirillum salinarum to Rhodovibrio salinarum comb. nov., of Rhodospirillum sodomense to Rhodovibrio sodomensis comb. nov., of Rhodospirillum salexigens to Rhodothalassium salexigens comb. nov., and of Rhodospirillum mediosalinum to Roseospira mediosalina comb. nov. Int. J. Syst. Bacteriol. 1998, 48, 793–798. [Google Scholar] [PubMed] [Green Version]
- Ramana, V.V.; Chakravarthy, S.K.; Ramaprasad, E.V.V.; Thiel, V.; Imhoff, J.F.; Sasikala, C.; Ramana, C.V. Emended description of the genus Rhodothalassium Imhoff et al., 1998 and proposal of Rhodothalassiaceae fam. nov. and Rhodothalassiales ord. nov. Syst. Appl. Microbiol. 2013, 36, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, D.; Vargas, C.; Csonka, L.N.; Ventosa, A.; Nieto, J.J. Synthesis of Glycine Betaine from Exogenous Choline in the Moderately Halophilic Bacterium Halomonas elongata. Appl. Environ. Microbiol. 1998, 64, 4095–4097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, S.; Cressatti, M.; Mendoza, K.E.; Coumoundouros, C.N.; Plater, S.M.; Culham, D.E.; Kimber, M.S.; Wood, J.M. YehZYXW of Escherichia coli is a low-affinity, non-osmoregulatory betaine-specific ABC transporter. Biochemistry 2015, 54, 5735–5747. [Google Scholar] [CrossRef] [PubMed]
- Christian, J.H.G.; Whalto, J.A. Solute concentrations within cells of halophilic and non-halophilic bacteria. Biochim. Biophys. Acta 1962, 65, 506–508. [Google Scholar] [CrossRef]
- Kushner, D.J. (Ed.) Life in high salt and solute concentrations: Halophilic bacteria. In Microbial Life in Extreme Environments; Academic Press: New York, NY, USA; London, UK, 1978; pp. 317–368. [Google Scholar]
- Imhoff, J.F.; Hashwa, F.; Trüper, H.G. Isolation of extremely halophilic phototrophic bacteria from the alkaline Wadi Natrun, Egypt. Arch. Hydrobiol. 1978, 84, 381–388. [Google Scholar]
- Imhoff, J.F.; Sahl, H.G.; Soliman, G.S.H.; Trüper, H.G. The Wadi Natrun: Chemical composition and microbial mass developments in alkaline brines of Eutrophic Desert Lakes. Geomicrobiol. J. 1979, 1, 219–234. [Google Scholar] [CrossRef]
- Imhoff, J.F. Anoxygenic phototrophic bacteria from extreme environments. In Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Hallenbeck, P.C., Ed.; Springer: Cham, Switzerland, 2017; pp. 427–480. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Gene Repertoire of Various Orders of Phototrophic Bacteria for Osmotic Adaptation | ||||||||
|---|---|---|---|---|---|---|---|---|
| Family | Species | Strain | Salt | Betaine | biosynthesis | Osmolyte transport | ||
| response | from glycine | from choline | ||||||
| GMT-DMT | betAB | betT | opuA /opuC | proW1 | ||||
| opuD | proW2 | |||||||
| Acidobacteria/Acidobacteriales | ||||||||
| Acidobacteriaceae | Chloracidobacterium thermophilum | B-G2 | F | o | o | o | o | o |
| Chlorobi/Chlorobiales | ||||||||
| Chlorobiaceae | Chlorobaculum thiosulfatophilum | DSM 249 | F | o | o | o | o | o |
| Chlorobiaceae | Chlorobium limicola | DSM 245 | F | o | o | o | o | o |
| Chlorobiaceae | Chlorobium phaeovibrioides | DSM 265 | F | o | o | o | o | o |
| Chlorobiaceae | Chloroherpeton thalassium | ATCC 35110 | M | GMT-DMT-betT | o | o | o | o |
| Chlorobiaceae | Prosthecochloris aestuarii | DSM 271 | M | o | o | o | o | o |
| Chlorobiaceae | Prosthecochloris vibrioformis | DSM 260 | M | o | o | o | proVopuAB,AC | o |
| Chloroflexi/Chloroflexales | ||||||||
| Chloroflexaceae | Chloroflexus aggregans | DSM 9485 | F | o | o | o | ||
| Chloroflexaceae | Chloroflexus aurantiacus | J-10-fl | F | o | o | o | o | |
| Roseiflexaceae | Roseiflexus castenholzii | DSM 13941 | F | o | o | opuCB-proXV | o | |
| Roseiflexaceae | Roseiflexus sp. | RS-1 | F | o | o | opuCB-proXV | o | |
| Cyanobacteria/Synechococcales | ||||||||
| Prochloraceae | Prochlorococcus marinus | MIT 9313 | M | GMT-DMTproVWX | o | o | o | o |
| Synechococcaceae | Synechococcus species | WH8102 | M | GMT-DMTproVWX | o | o | o | o |
| Synechococcaceae | Dactylococcopsis salina | PCC 8305 | H | GMT-DMT | o | o | o | W1 |
| Cyanobacteria/Chroococcales | ||||||||
| Aphanothecaceae | Halothece sp./Aphanothece halophytica | PCC 7418 | H | GMT-DMT | o | /betT/ | o | W1 |
| Aphanothecaceae | Euhalothece natronophila | Z-M001 | H | GMT-DMT | o | /betT/ | o | W1 |
| Firmicutes/Clostridiales | ||||||||
| Heliobacteriaceae | Heliobacterium modesticaldum | Ice1 | F | o | o | o | o | o |
| Gemmatimonadetes/Gemmatimonadales | ||||||||
| Gemmatimonadaceae | Gemmatimonas phototrophica | AP64 | F | o | o | o | o | o |
| Betaproteobacteria/Burkholderiales | ||||||||
| Burkholderiaceae | Polynucleobacter duraquae | MWH-MoK4 | F | o | o | o | o | o |
| Comamonadaceae | Rhodoferax antarcticus | DSM 24876 | F | o | o | o | o | W2 |
| Comamonadaceae | Rhodoferax fermentans | DSM 10138 | F | o | o | o | opuD | o |
| uncl. Burkholderiales | Rubrivivax gelatinosus | IL144 | F | o | o | o | opuD | o |
| uncl. Burkholderiales | Rubrivivax gelatinosus | DSM 1709 | F | o | o | o | opuD | o |
| uncl. Burkholderiales | Rubrivivax gelatinosus | IM 151 | F | o | o | o | opuD | o |
| uncl. Burkholderiales | Rubrivivax gelatinosus 155 | DSM 149 | F | o | o | o | opuD | o |
| Betaproteobacteria/Rhodocyclales | ||||||||
| Rhodocyclaceae | Rhodocyclus purpureus | TEM | F | o | o | o | o | o |
| Rhodocyclaceae | Rhodocyclus tenuis | IM 230 | F | o | o | o | o | o |
| Gene Repertoire of Phototrophic Alphaproteobacteria for Osmotic Adaptation | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Family | Species | Strain | Salt | Ectoine | Betaine | biosynthesis | Osmolyte transport | |||||
| response | biosynthesis | from glycine | from choline | |||||||||
| ectABC | GMT-DMT | betAB | betT | opuA | proW1 | proW2 | proW3 | proW4 | ||||
| Rhizobiales | opuD | |||||||||||
| Beijerinckiaceae | Methylocella silvestris | BL2 | F | o | o | o | o | o | o | o | o | o |
| Beijerinckiaceae | Rhodoblastus acidophilus | DSM 137 | F | o | o | o | o | o | o | o | o | o |
| Beijerinckiaceae | Rhodoblastus sphagnicola | DSM 16996 | F | o | o | o | o | o | o | o | o | o |
| Bradyrhizobiaceae | Bradyrhizobium oligotrophicum | S58 | F | o | o | o | o | o | o | o | o | o |
| Bradyrhizobiaceae | Rhodopseudomonas palustris | DSM 126 | F | o | o | /betA/ 3x /betB/ | o | o | o | o | o | o |
| Bradyrhizobiaceae | Rhodopseudomonas pseudopalustris | DSM 123T | F | o | o | o | o | o | o | o | o | o |
| Hyphomicrobiaceae | Blastochloris tepida | GI | F | o | o | o | o | o | o | o | o | o |
| Hyphomicrobiaceae | Blastochloris viridis | DSM 133 | F | o | o | o | o | o | o | o | o | o |
| Hyphomicrobiaceae | Rhodomicrobium vannielii | ATCC 17100 | F | o | o | o | o | o | o | o | o | o |
| Hyphomicrobiaceae | Rhodoplanes elegans | DSM 11907 | F | o | o | o | o | o | o | o | o | o |
| Aurantimonadaceae | Fulvimarina pelagi | HTCC2506 | M | ectB/ | o | betABCIproXWV | /betT/ 3x | o | W1 | W2 | W3 | o |
| Phyllobacteriaceae | Hoeflea phototrophica | DFL-43 | M | o | o | betABCI | /betT/ | o | o | W2 | o | W4 |
| Rhodobiaceae | Afifella marina 125/2 | IM 162 | M | o | o | proXbetIA /betB/ | /betT/ | o | W1 | o | o | o |
| Rhodobiaceae | Afifella marina 125/4) | IM 163 | M | o | o | proXbetIA /betB/ | /betT/ | o | W1 | o | o | o |
| Rhodobiaceae | Afifella marina 985, 126 (166) | DSM 2698 | M | o | o | proXbetIA /betB/proWV | /betT/ | o | W1 | o | W3 | o |
| Rhodobiaceae | Afifella pfennigii | DSM 17143 | M | o | o | proXbetIA /betB/ | /betT/ | o | W1 | o | o | o |
| Rhodobiaceae | Rhodobium orientis | DSM 11290 | M | o | o | betABIbetCproXWV | /betT/ | o | o | W2 | W3 | o |
| Rhodobacterales | ||||||||||||
| Rhodobacteraceae | Rhodobacter capsulatus | SB 1003 | F | o | o | betABIproXWV | o | o | o | W3 | o | |
| Rhodobacteraceae | Rhodobacter sphaeroides | ATCC 17025 | F | o | o | betABIproXWV | o | o | W2 | W3 | o | |
| Rhodobacteraceae | Rhodobacter veldkampii | DSM 11550 | F | o | o | betABIproXWV | o | o | W2 | W3 | W4 | |
| Rhodobacteraceae | Rhodobaca barguzinensis | DSM19920 | M | ectRABCask-ect | o | betAB>proXWV | /betT/ 2x | o | o | W2 | W3 | o |
| Rhodobacteraceae | Rhodobaculum claviforme | GOR B7-4 | M | ectRABCask-ect | o | o | /betT/ 2x | o | o | o | o | o |
| Rhodobacteraceae | Rhodovulum adriaticum | DSM2781 | M | ectRABCask-ect | o | o | /betT/ | o | o | W2 | o | o |
| Rhodobacteraceae | Rhodovulum imhoffii | DSM 18064 | M | ectRABCask-ect | o | o | /betT/ 2x | o | o | o | o | o |
| Rhodobacteraceae | Rhodovulum sulfidophilum | IM 196 | M | ectRABCask-ect | o | o | /betT/ | o | o | W2 | o | o |
| Rhodobacteraceae | Rhodovulum sulfidophilum | DSM 2351 | M | ectRABCask-ect | o | o | /betT/ | o | o | W2 | o | o |
| Rhodobacteraceae | Rhodovulum sulfidophilum | DSM 1374 | M | ectRABCask-ect | o | o | /betT/ | o | o | W2 | o | o |
| Rhodobacteraceae | Roseisalinus antarcticus | DSM 11466 | M | ectR-X-ectABCask-ect | o | betABNCIproXWV/betAT | /betT/ 2x | o | o | o | W3 | o |
| Rhodobacteraceae | Roseivivax isoporae | LMG 25204 | M/H | ectRABCask-ect | o | betABC /<bet>proWX | /betT/ 4x | o | o | o | o | |
| Rhodobacteraceae | Roseivivax halotolerans | JCM 10271 | M/H | ectRABCask-ect | o | betABC /betI>proWV | /betT/ 3x | o | o | W2 | o | |
| Rhodobacteraceae | Roseivivax halodurans | JCM 10272 | M/H | ectRABCask-ect | fusedMT-MAT | betABTC /betAT/bet>proWX | /betT/ 3x | o | o | W2 | o | |
| Rhodobacteraceae | Roseivivax roseus | DSM 23042 | H | ectRABCask-ect | fusedMT-MTHFR-MS-MAT-SAHase | betABCIproX/proWV | /betT/ 5x | o | W1 | o | W4 | |
| Rhodobacteraceae | Rhodosalinus sediminis | KCTC52478 | H | ectRABCask-ect | fusedMT- MS-MAT | betABIproXWV | betT/ | o | o | W2 | W3 | o |
| Rhodobacteraceae | Roseovarius nitratireducens | KTCT52967 | M/H | ectRABCask-ect | fusedMT-MTHFR-MS-MAT-SAHase | betACINproXWV | betT/ | o | o | W2 | W3 | W4 |
| Rhodobacteraceae | Roseovarius halotolerans | DSM 29507 | M/H | ectRABCask-ect | o | betABCINproXWV | /betT/ 2x | o | W1 | o | W3 | W4 |
| Rhodobacteraceae | Roseovarius mucosus | SMR3 | M | ectRABCask-ect | o | betACIproXWV | /betT/ 2x | o | o | o | W3 | W4 |
| Rhodobacteraceae | Roseovarius litoreus | DSM 28249 | M | ectRABCask-ect | o | betABCINproXWV | /betT/ 2x | o | o | W3 | W4 | |
| Rhodospirillales | ||||||||||||
| Acetobacteraceae | Acidiphilium cryptum | JF-5 | F | ectRABCDask_ect | o | proXWVbetBA | o | o | o | o | W3 | o |
| Acetobacteraceae | Acidiphilium multivorum | AIU301 | F | ectRABCDask_ect | o | proXWVbetBA | o | o | o | o | W3 | o |
| Acetobacteraceae | Paracraurococcus ruber | DSM 15382 | F | o | o | o | o | o | o | W2 | o | o |
| Acetobacteraceae | Rhodopila globiformis | DSM 161 | F | o | o | o | o | o | o | o | o | o |
| Rhodospirillaceae | Pararhodospirillum photometricum | DSM 122 | F | o | o | o | o | o | o | o | o | o |
| Rhodospirillaceae | Phaeospirillum fulvum | MGU-K5 | F | o | o | o | o | o | o | o | o | o |
| Rhodospirillaceae | Phaeospirillum molischianum | DSM 120 | F | o | o | o | o | o | o | o | o | o |
| Rhodospirillaceae | Rhodospirillum rubrum | DSM 1068 | F | o | o | proXbetAB/betI/ | o | o | o | o | o | o |
| Rhodospirillaceae | Rhodospirillum rubrum 220 | DSM 107 | F | o | o | proXbetAB/betI/ | o | o | o | o | o | o |
| Rhodospirillaceae | Rhodospirillum rubrum | ATCC 11170 | F | o | o | o | o | o | W1 | o | o | o |
| Rhodospirillaceae | Rhodospirillum rubrum | FR1Mutante-IV | F | o | o | o | o | o | W1 | o | o | o |
| Rhodospirillaceae | Rhodovibrio salinarum | DSM 9154 | H | ectBC/2 x ectA | GMT-B-DMT-MAT-SAHase | betABIproX | betTproX | proVWopuAC | o | o | W4 | |
| Rhodospirillaceae | Rhodovibrio sodomensis | DSM 9895 | H | ectBC/2 x ectA | GMT-B-DMT-MAT-SAHase | betABIproX | /betT/ 6x | o | W1 | o | o | W4 |
| Rhodospirillaceae | Caenispirillum salinarum | AK4 | M | ectABCD/A/D | fusedMT-MAT | betABIproXWV | /betT/ | opuD | W1 | o | W3 | o |
| Rhodospirillaceae | Rhodospira trueperi | ATCC 700224 | M | ectABC | fusedMT-MAT | o | /betT/ 2x | o | W1 | W2 | o | o |
| Rhodospirillaceae | Roseospira marina | DSM 15113 | M | ectABC | MAT-fusedMT | betIBAproX | /betT/ 2x | o | W1 | W2 | o | o |
| Rhodospirillaceae | Roseospira navarrensis | DSM 15114 | M | ectABC | MAT-fusedMT | betIBAproX | /betT/ | o | W1 | o | o | W4 |
| Rhodospirillaceae | Roseospirillum parvum | DSM 12498 | M | ectRABC | MAT-fusedMT | o | /betT/ 2x | o | W1 | W2 | o | o |
| Rhodothalassiales | ||||||||||||
| Rhodothalassiaceae | Rhodothalassium salexigens | IM 261 | H | o | GMT-DMT | o | /betT/ 3x | o | W1 | o | o | o |
| Rhodothalassiaceae | Rhodothalassium salexigens | IM 265 | H | o | GMT-DMT | o | /betT/ 3x | o | W1 | o | o | o |
| Rhodothalassiaceae | Rhodothalassium salexigens | DSM 2132 | H | o | GMT-DMT | o | /betT/ 3x | o | o | o | o | o |
| Sphingomonadales | ||||||||||||
| Erythrobacteraceae | Erythrobacter litoralis | DSM 8509 | M | o | GMT-DMT | o | o | o | o | o | o | |
| Gene repertoire of Phototrophic Gammaproteobacteria for Osmotic Adaptation | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Family | Species | Strain | Salt | Ectoine | Betaine biosynthesis | Osmolyte transport | |||
| response | biosynthesis | from glycine | from choline | ||||||
| ectABC | GMT-DMT | betAB | betT | opuA | proW1 | ||||
| opuD | |||||||||
| Cellvibrionales | |||||||||
| Halieaceae | Congregibacter litoralis | KT71 | M | o | o | no genes of osmotic stress synthesis and transport | |||
| Chromatiales | |||||||||
| Chromatiaceae | Allochromatium vinosum | MT86 | F | o | o | o | o | o | o |
| Chromatiaceae | Allochromatium vinosum | DSM 180 | F | o | o | o | o | o | o |
| Chromatiaceae | Allochromatium warmingii | DSM 173 | F | o | o | o | o | o | o |
| Chromatiaceae | Chromatium okenii 6010 | DSM 169 | F | o | o | o | o | o | o |
| Chromatiaceae | Chromatium weissei IM 5910 | DSM 5161 | F | o | o | o | o | o | o |
| Chromatiaceae | Lamprocystis purpurea | DSM 4197 | F | o | o | o | o | o | o |
| Chromatiaceae | Thiocystis minor | DSM 178 | F | o | o | o | o | o | o |
| Chromatiaceae | Thiocystis violacea | DSM 207 | F | o | o | o | o | o | o |
| Chromatiaceae | Thiocystis violascens | DSM 198 | F | o | GMTtruncated,DMT | o | o | o | o |
| Chromatiaceae | Thiocapsa imhoffii | DSM 21303 | F | o | o | o | betT/ | o | o |
| Chromatiaceae | Thiocapsa roseopersicina | DSM 217 | F | o | o | o | betT/ | o | o |
| Chromatiaceae | Thiocapsa marina 5811 | DSM 5653 | M | o | GMT-DMT-betT | o | opuAA,AB,AC-N-GMT,DMT,betT | o | |
| Chromatiaceae | Marichromatium gracile 5210 | DSM 203 | M | o | GMT-DMT | betBAT | betT/ 2x | opuAA,AB,AC-N-opuAC-NN-betT | o |
| Chromatiaceae | Marichromatium purpuratum 984 | DSM 1591 | M | o | GMT-DMT | o | betT/ | opuAA,AB,AC,N,AC,AC,NN,betT | o |
| Chromatiaceae | Thiocystis violacea | DSM 208 | M | o | GMT-DMT | o | betTopuAC | opuAA,AB,AC | o |
| Chromatiaceae | Imhoffiella purpurea AK35 | AK35 | M | o | GMT-DMT | o | betTopuAC | opuAA,AB,AC | o |
| Chromatiaceae | Thiorhodococcus drewsii | AZ1 | M | o | GMT-DMT | betBAT/betAB | betT-opuAC/betT | opuAA,AB,AC | o |
| Chromatiaceae | Thiorhodococcus mannitoliphagus | DSM 18266 | M | o | GMT-DMT | o | betT-opuAC/betT | opuAA,AB,AC | o |
| Chromatiaceae | Thiorhodococcus minor | DSM 11518 | M | o | GMT-DMT | o | betT/ 4x | o | W1 |
| Chromatiaceae | Thioflavicoccus mobilis 8321 | ATCC700959 | M | o | GMT-DMT /GMT-B | o | betT/ 2x | o | W1-N-betT |
| Chromatiaceae | Thiococcus pfennigii 4252 | DSM 228 | M | o | GMT-DMT | o | betT/ 2x | o | W1 |
| Chromatiaceae | Thiococcus pfennigii 4254 | Pfennig 8320 | M | o | GMT-DMT | o | betT/ 3x | o | W1 |
| Chromatiaceae | Thiorhodovibrio winogradskyi | 06511 | M | o | GMT-DMT | o | betT/ 3x | proVW1-opuAC | |
| Chromatiaceae | Rhabdochromatium marinum | DSM 5261 | M | o | GMT-DMT /GMT-B | o | betT/ 3x | proVW1-opuAC | |
| Chromatiaceae | Lamprobacter modestohalophilus | DSM 25653 | M | o | GMT-DMT | o | betT/ 3x | o | W1 |
| Chromatiaceae | Halochromatium roseum | DSM 18859 | M | o | GMT-DMT | o | betT/ 2x | o | W1 |
| Chromatiaceae | Halochromatium glycolicum | DSM 11080 | H | o | GMT-DMT /GMT-B | o | betT/ 4x | W1-NN-betTopuAC | |
| Chromatiaceae | Halochromatium salexigens IM6310 | DSM 4395 | H | o | GMT-DMT /GMT-B | o | betT/ 2x | W1-N-betTopuAC-NN-betT | |
| Chromatiaceae | Thiohalocapsa halophila IM4270 | DSM 6210 | H | o | GMT-DMT | o | betT/ 5x | o | W1 |
| Ectothiorhodospiraceae | Ectothiorhodospira mobilis | DSM 237 | M | o | GMT-DMT-MAT /GMT-B | o | betT/ | betT-opuAA,AB,AC | o |
| Ectothiorhodospiraceae | Ectothiorhodospira marismortui | DSM 4180T | M/H | o | GMT-DMT-MAT /GMT-B | o | betT/ | betT-opuAA,AB,AC | o |
| Ectothiorhodospiraceae | Ectothiorhodospira marina | DSM 241T | M/H | o | GMT-DMT-MAT /GMT-B | betABIproX | betT | betT-opuAA,AB,AC | o |
| Ectothiorhodospiraceae | Ectothiorhodospira haloalkaliphila | ATCC 51935 | M/H | o | GMT-DMT-MAT /GMT-B | betABIproX | betT/ 2x | betT-opuAA,AB,AC | OpuD |
| Ectothiorhodospiraceae | Ectothiorhodospira sp. | BSL-9 | M | o | GMT-DMT-MAT /GMT-B | betABIproX | betT/ | betT-opuAA,AB,AC | o |
| Ectothiorhodospiraceae | Ectothiorhodospira shaposhnikovii | DSM 243 | M | ectB/ | GMT-DMT-MAT | betABIproX | betT-opuAA,AB,AC | o | |
| Ectothiorhodospiraceae | Ectothiorhodospira magna B7-7 | DSM 22250 | M | o | o | o | betT-opuAA,AB,AC | o | |
| Ectothiorhodospiraceae | Ectothiorhodosinus mongolicus M9 | DSM 15479 | M | o | o | o | betT/ | o | W1 |
| Ectothiorhodospiraceae | Thiorhodospira sibirica | ATCC 700588 | M | o | o | o | o | o | o |
| Ectothiorhodospiraceae | Halorhodospira abdelmalekii | DSM 2110 | H | ectAB/C | GMT-DMT-MAT-SAHase-MTHFR | o | see proU | opuD/ 2x | W1-betT |
| Ectothiorhodospiraceae | Halorhodospira halochloris | DSM 1059 | H | ectABC | GMT-DMT-MAT-SAHase-MTHFR | o | betT/ 2x | opuD | o |
| Ectothiorhodospiraceae | Halorhodospira halophila | IM 9626 | H | ectABC | GMT-DMT-MAT-SAHase-MTHFR/GMT-B | o | betT/ 5x | opuD | W1 |
| Ectothiorhodospiraceae | Halorhodospira halophila SL1 | DSM 244 | H | ectABC | GMT-DMT-MAT-SAHase-MTHFR/GMT-B | o | betT/ 4x | opuD | W1-N-betT |
| Ectothiorhodospiraceae | Halorhodospira halophila D | IM 9620 | H | ectABC | GMT-DMT-MAT-SAHase-MTHFR/GMT-B | o | betT/ 5x | opuD | W1-N-betT |
| Ectothiorhodospiraceae | Halorhodospira halophila 51/3 | IM 9630 | H | ectABC | GMT-DMT-MAT-SAHase-MTHFR/GMT-B | o | betT/ 5x | opuD | W1 |
| Ectothiorhodospiraceae | Halorhodospira neutriphila | DSM 15116 | H | ectABC | GMT-DMT-MAT-SAHase-MTHFR/GMT-B | o | betT/ 3x | o | W1 |
| Family | Species | Strain | Ectoine | Betaine | biosynthesis |
| biosynthesis | from glycine | from choline | |||
| ectABC | GMT-DMT | betAB | |||
| Actinobacteria | |||||
| Actinopolysporaceae | Actinopolyspora halophila | DSM 43834 | ectABC | fusedMT-MAT-o-MS-SAHase-MTHFR | o |
| Gordoniaceae | Gordonia alkanivorans | NBRC16433 | ectABC | o | betABT |
| Mycobacteriaceae | Mycolicibacterium thermoresistibile | DSM 44167 | ectABCD | o | betA/betB |
| Streptomycetaceae | Streptomyces clavuligerus | ATCC 27064 | ectABCD/AB | o | betAB/betI/betT |
| Bacilli/Bacillales | |||||
| Bacillaceae | Halobacillus halophilus | DSM 2266 | ectABC/D | o | betABbetIopuAC |
| Proteobacteria/Alphaproteobacteria | |||||
| Rhodobacteraceae | Jannaschia seosinensis | CECT7799 | ectRectABCask-ect | o | proVWXbetICBA |
| Rhodobacteraceae | Maritimibacter alkaliphilus | HTCC2654 | ectRectABCask-ect | o | proVWooXbetICBA |
| Rhodobacteraceae | Paracoccus halophilus | JCM 14014 | ectRectABCask-ect | o | proVWXbetIBA |
| Rhodobacteraceae | Salipiger bermudensis | HTCC2601 | ectRectABCask-ect | o | proVWXbetICBA |
| Rhodobacteraceae | Thioclava pacifica | DSM 10166 | ectRectABCask-ect | o | proVWoXbetIBA |
| Rhodospirillaceae | Ferruginivarius sediminum | WD2A32 | ectRectABC/B/C/D | o | proXbetIBA |
| Proteobacteria/Betaproteobacteria | |||||
| Alcaligenaceae | Achromobacter xylosoxidans | SOLR10 | ectRABCD | o | betA/betB |
| Alcaligenaceae | Bordetella avium | 197N | ectRABCD | o | betA/ |
| Burkholderiaceae | Paucimonas limoigeni | DSM 7445 | ectRABCD | o | betA/ |
| Oxalobacteraceae | Herminiimonas arsenicooxidans | DSM 17148 | ectRABCD | o | betA/ |
| Proteobacteria/Gammaproteobacteria | |||||
| Halomonadaceae | Halomonas elongata | DSM 2581 | ectABC/D | o | proXbetIBA |
| Halomonadaceae | Chromohalobacter salexigens | DSM 3043 | ectABC/D | o | proXbetIBA |
| Halomonadaceae | Chromohalobacter marismortui | DSM 6770 | ectABC/D | o | proXbetIBA |
| Haliaceae | Haliea salexigens | DSM 19537 | ectRectABCask-ect | o | betA/ |
| Chromatiaceae | Nitrosococcus oceani | ATCC 19707 | ectRCask-ect/ectABD | o | betA/ |
| Chromatiaceae | Nitrosococcus halophilus | Nc4 | ectRCask-ect/ectABD | o | o |
| Hahellaceae | Hahella chejuensis | KCTC2396 | ectABCD | o | betIBAproXWV |
| Oceanospirillaceae | Marinomonas mediterranea | MMB1 | ectABC/R/ask-ect | o | betAB / betIproXWV |
| Ectothiorhodospiraceae | Nitrococcus mobilis | NB-231 | ectABC | GMT-DMT | betA/ |
| Chromatiaceae | Acidihalobacter prosperus | DSM 14174 | ectABC | o | o |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imhoff, J.F.; Rahn, T.; Künzel, S.; Keller, A.; Neulinger, S.C. Osmotic Adaptation and Compatible Solute Biosynthesis of Phototrophic Bacteria as Revealed from Genome Analyses. Microorganisms 2021, 9, 46. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010046
Imhoff JF, Rahn T, Künzel S, Keller A, Neulinger SC. Osmotic Adaptation and Compatible Solute Biosynthesis of Phototrophic Bacteria as Revealed from Genome Analyses. Microorganisms. 2021; 9(1):46. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010046
Chicago/Turabian StyleImhoff, Johannes F., Tanja Rahn, Sven Künzel, Alexander Keller, and Sven C. Neulinger. 2021. "Osmotic Adaptation and Compatible Solute Biosynthesis of Phototrophic Bacteria as Revealed from Genome Analyses" Microorganisms 9, no. 1: 46. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010046