Biofilm Formation among Stenotrophomonas maltophilia Isolates Has Clinical Relevance: The ANSELM Prospective Multicenter Study

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Multilocus Sequence Typing (MLST)
2.3. Microbiological and Clinical Data
2.4. Standardized Inoculum Preparation
2.5. Microtiter Plate (MTP) Assay for Biofilm Quantification
2.6. Antibiotic Susceptibility of Planktonic Cells
2.7. Antibiotic Activity Against Biofilm Formation and Mature Biofilm
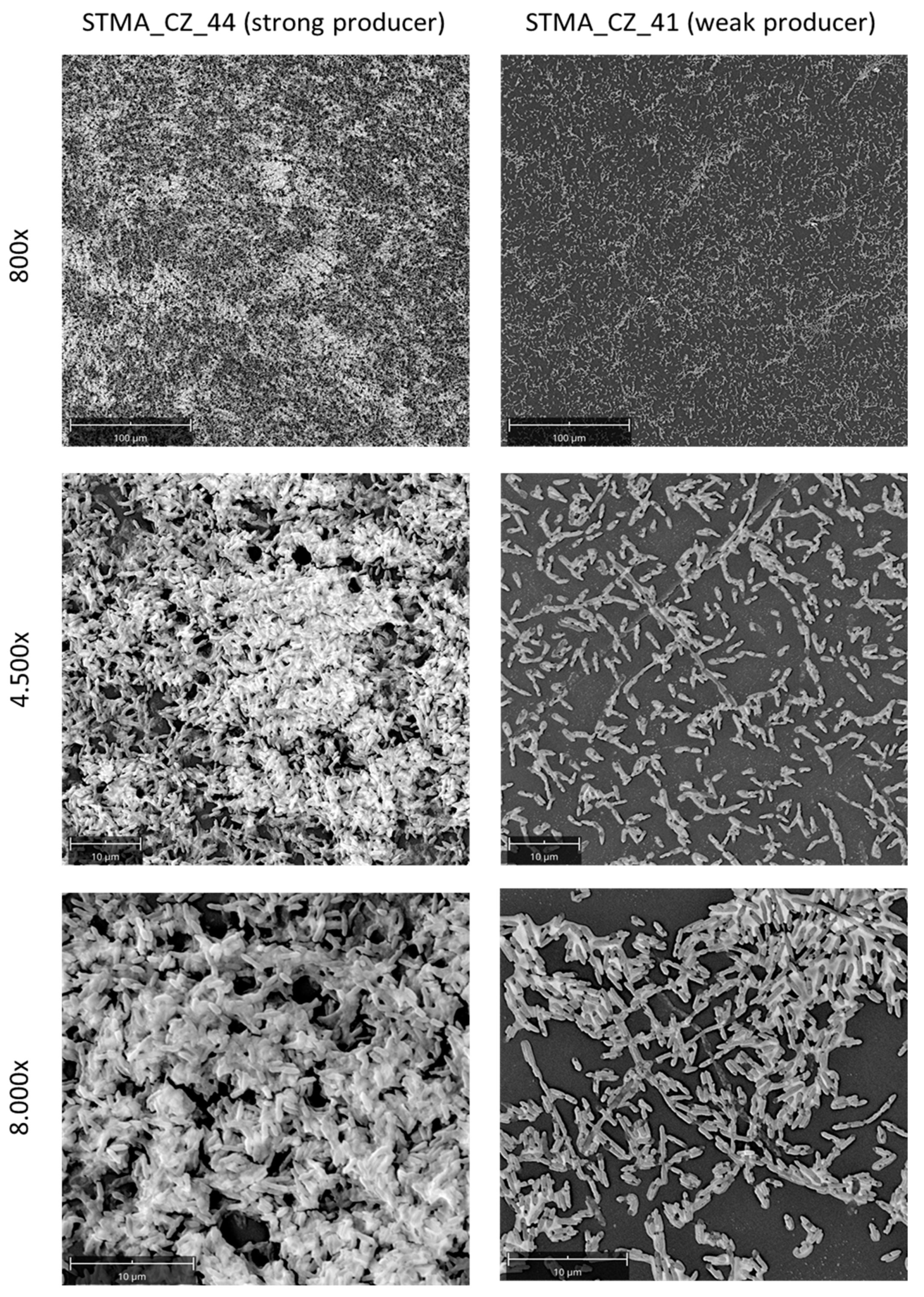
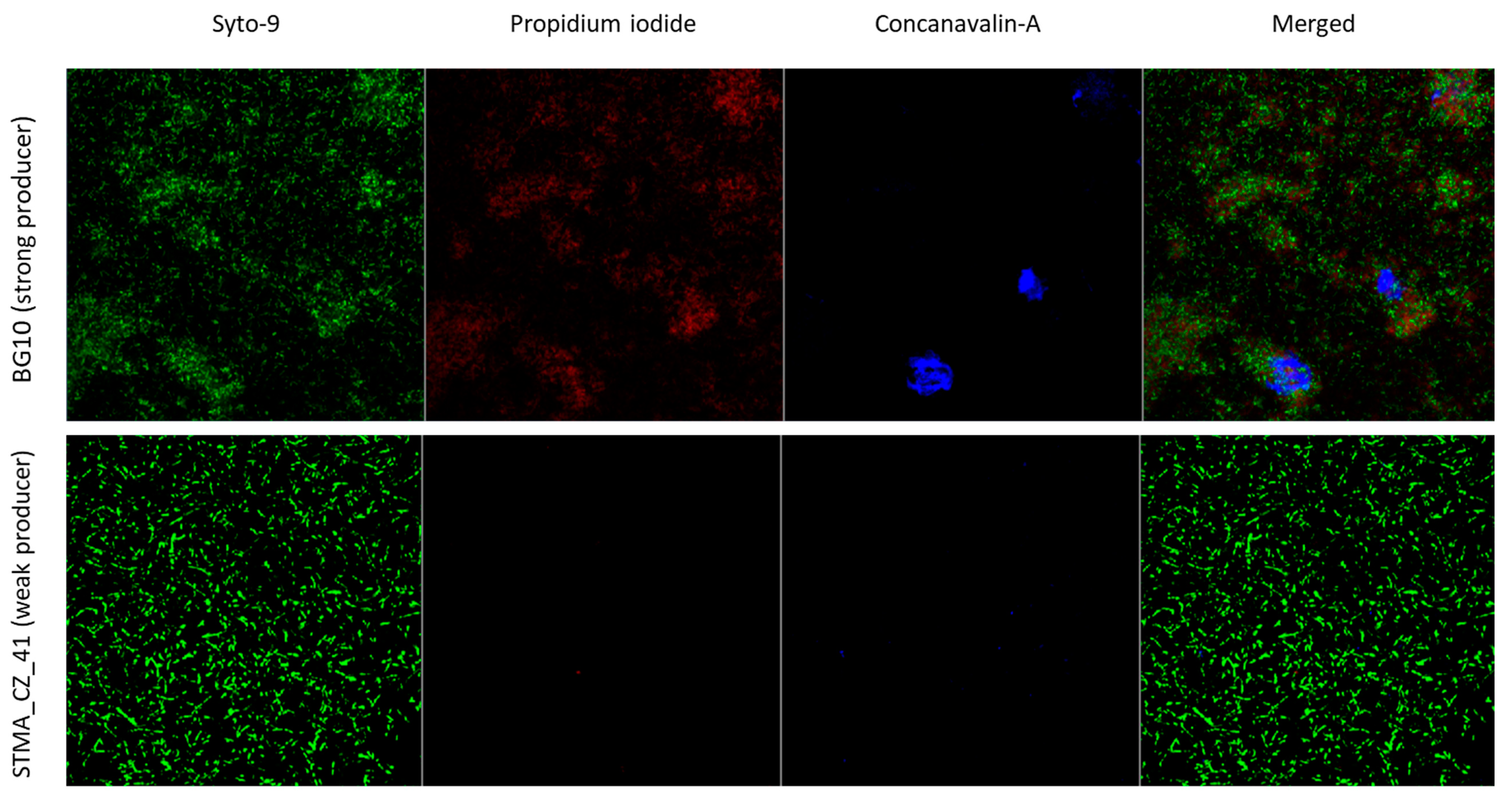
2.8. Evaluation of Biofilms by Microscopic Analysis
2.9. Statistical Analysis
3. Results
3.1. Microbiological and Clinical Features
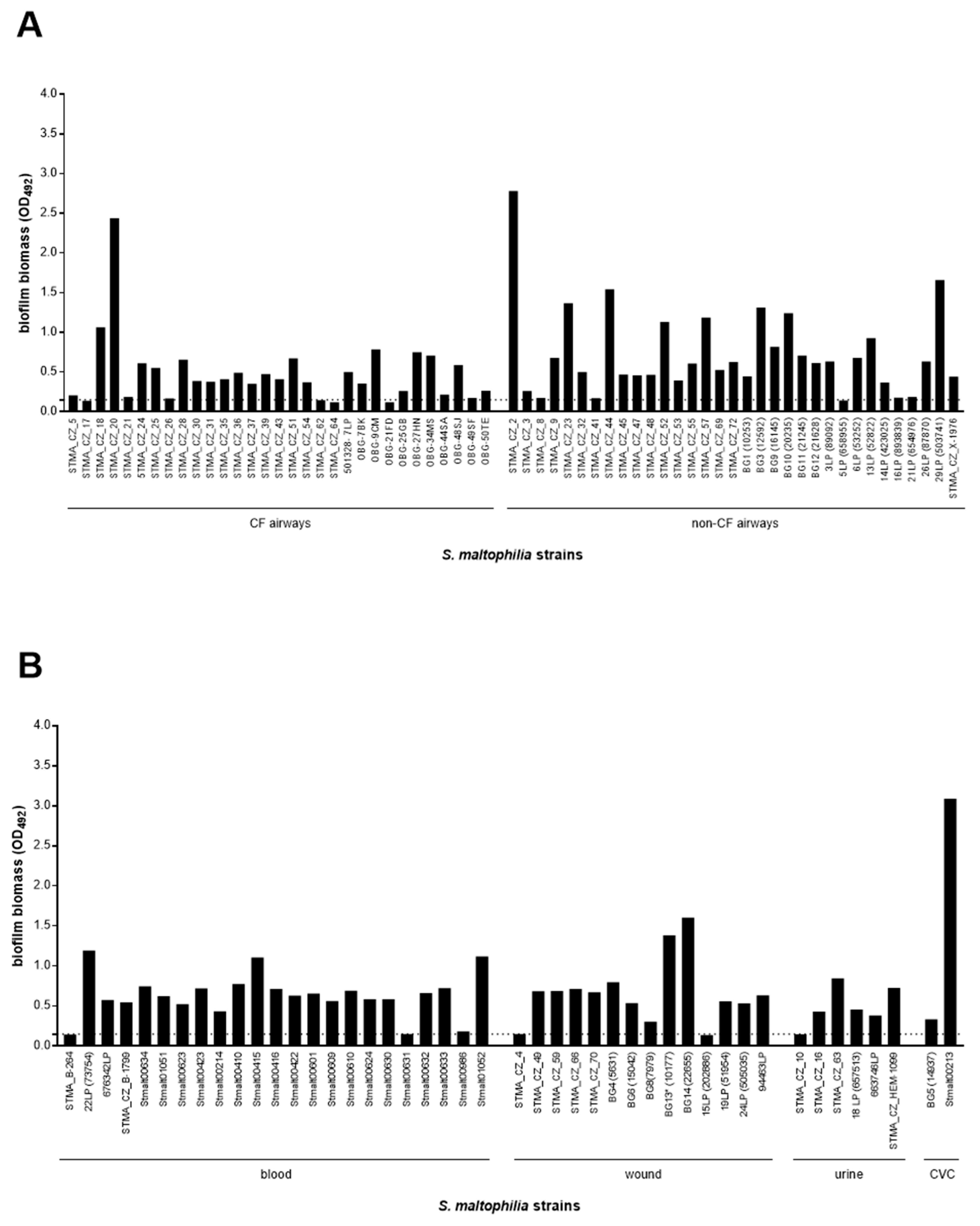
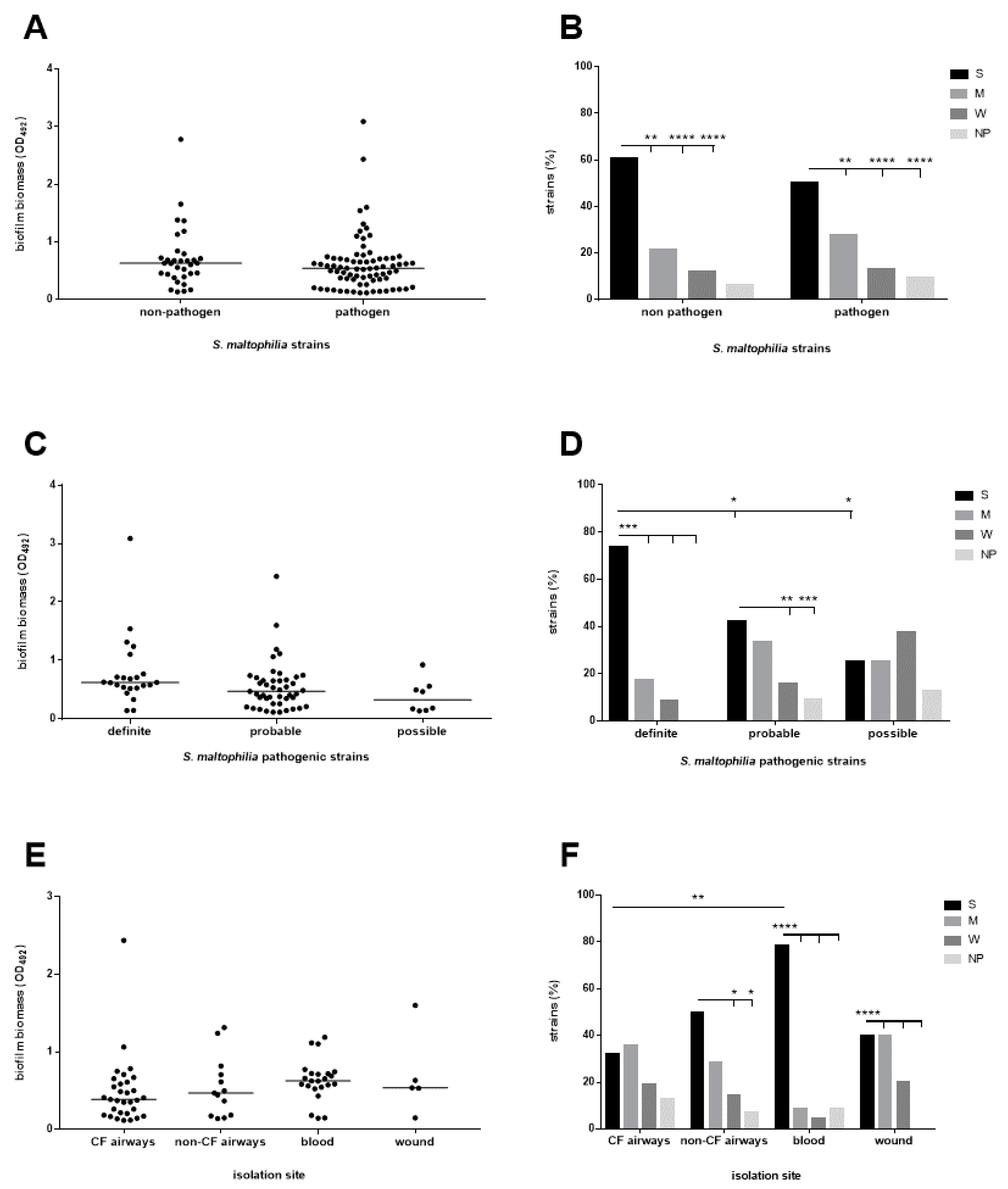
3.2. The Biofilm Forming Ability Is Highly Preserved in S. maltophilia, Although Strains with a Definite Etiological Role, Particularly Those From Blood, Show a Higher Efficiency
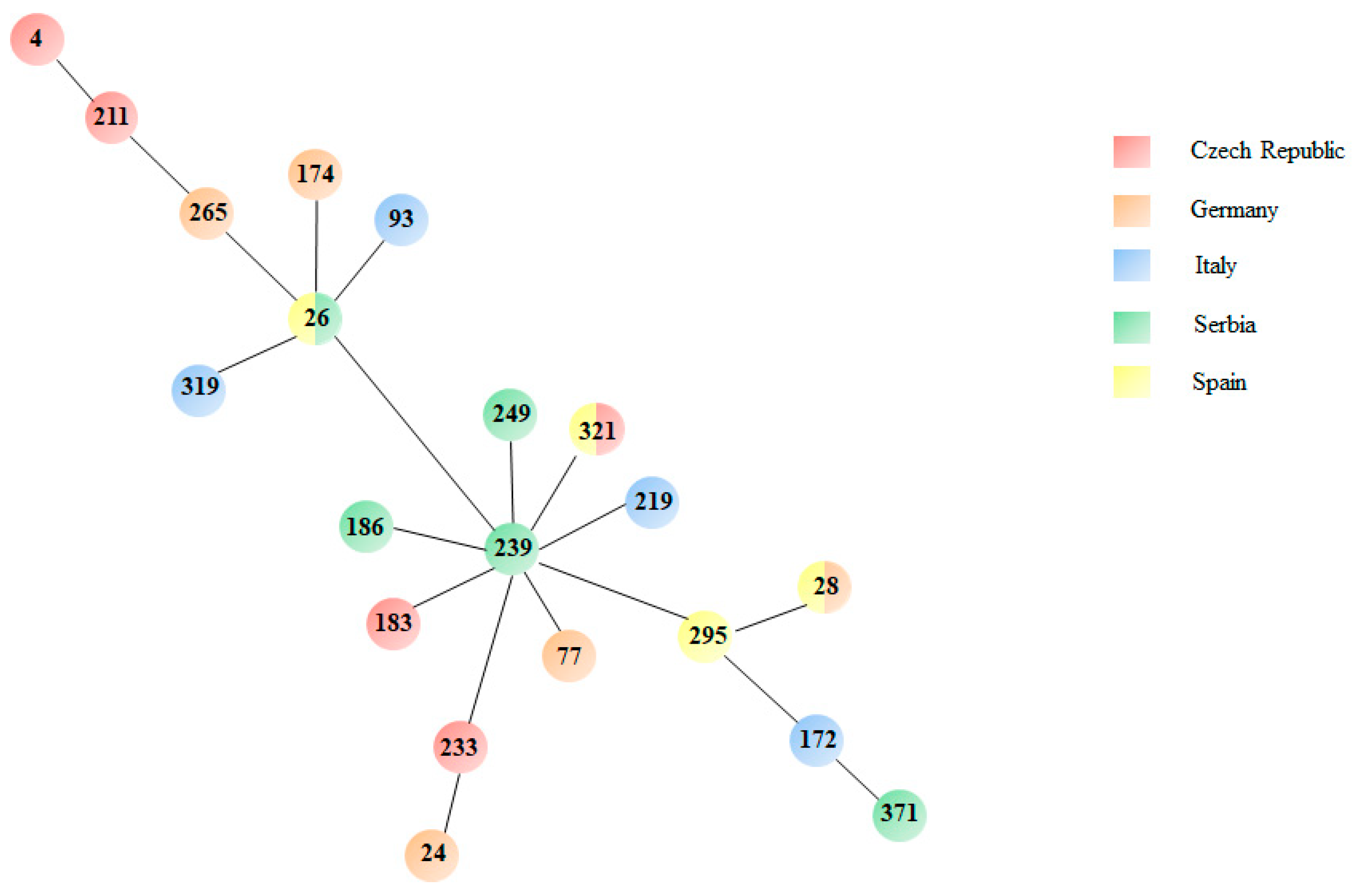
3.3. MLST Analysis Reveals High Genetic Diversity
3.4. Crystal Violet Assay Is Highly Predictive in the Quantitative Analysis of Biofilm Formation
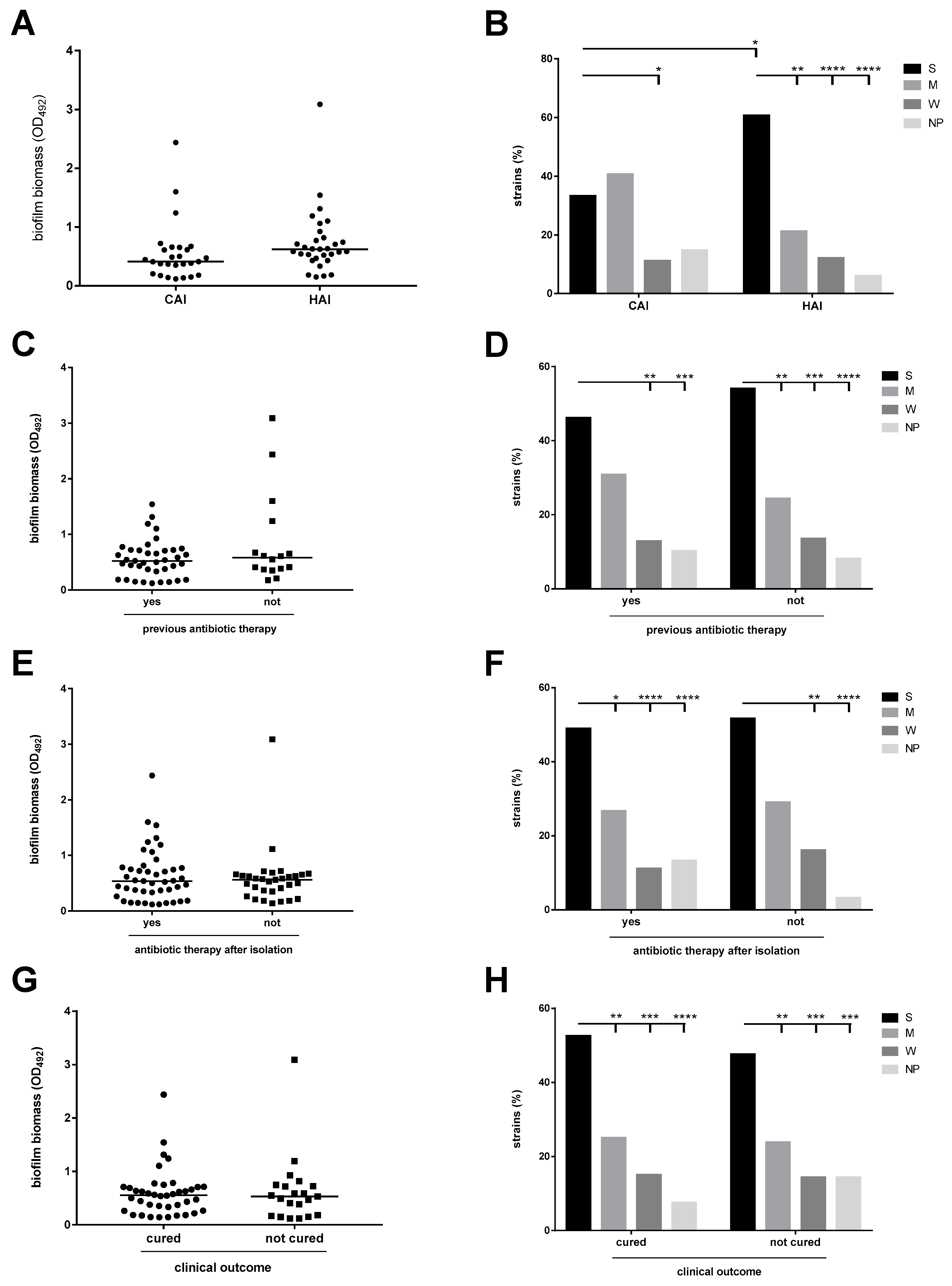
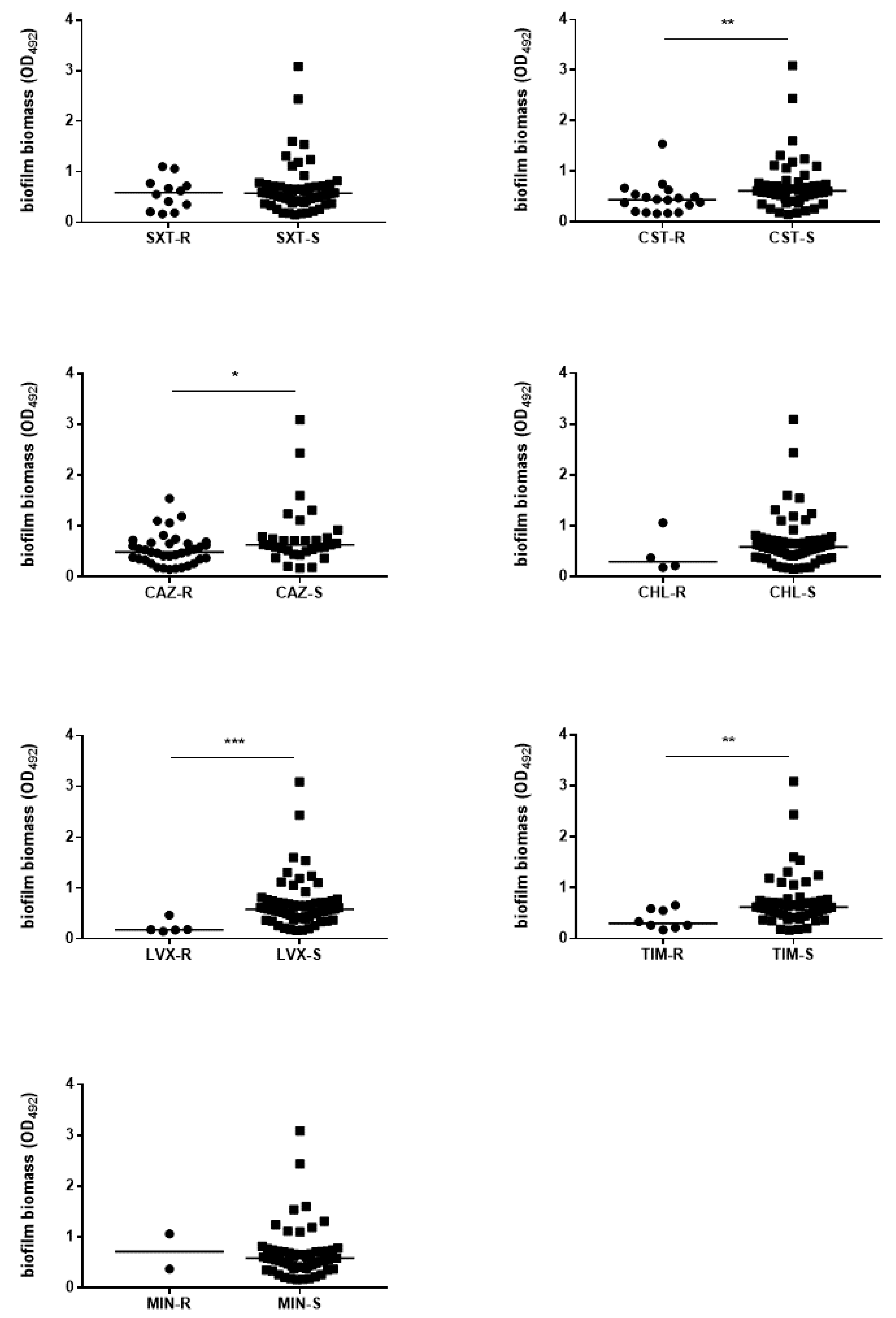
3.5. The Antibiotic Resistance Level Is Higher Among Pathogenic Strains
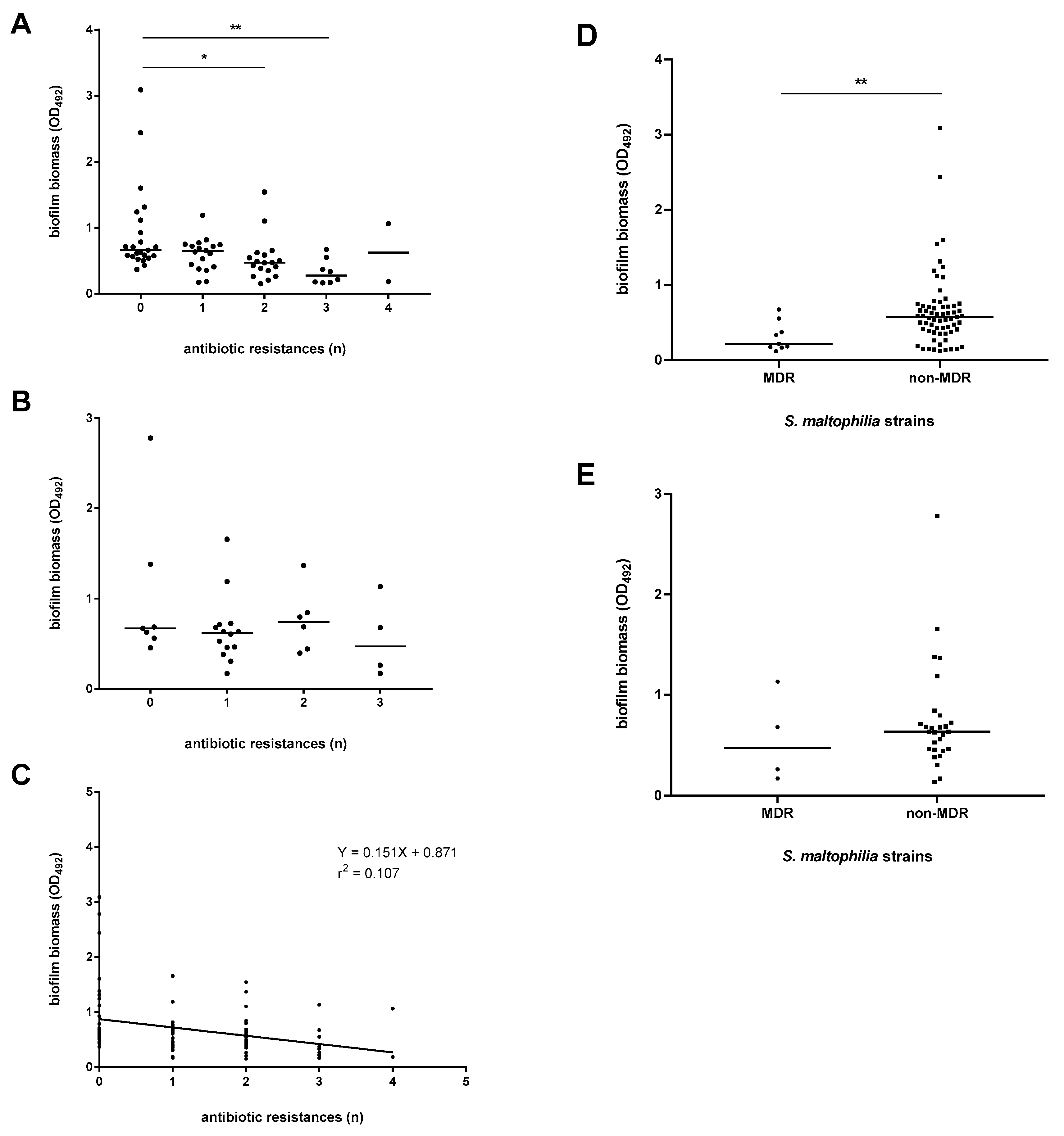
3.6. The Efficiency of Biofilm Formation Is Affected by Antibiotic Resistance in Pathogenic Strains Only
3.7. The Preformed S. maltophilia Biofilm Is Highly Resistant to Both Cotrimoxazole and Levofloxacin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pompilio, A.; Crocetta, V.; Ghosh, D.; Chakrabarti, M.; Gherardi, G.; Vitali, L.A.; Fiscarelli, E.; Di Bonaventura, G. Stenotrophomonas maltophilia phenotypic and genotypic diversity during a 10-year colonization in the lungs of a cystic fibrosis patient. Front. Microbiol. 2016, 7, 1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pompilio, A.; Crocetta, V.; De Nicola, S.; Verginelli, F.; Fiscarelli, E.V.; Di Bonaventura, G. Cooperative pathogenicity in cystic fibrosis: Stenotrophomonas maltophilia modulates Pseudomonas aeruginosa virulence in mixed biofilm. Front. Microbiol. 2015, 6, 951. [Google Scholar] [CrossRef] [PubMed]
- Pompilio, A.; Crocetta, V.; Scocchi, M.; Pomponio, S.; Di Vincenzo, V.; Mardirossian, M.; Gherardi, G.; Fiscarelli, E.V.; Dicuonzo, G.; Gennaro, R.; et al. Potential novel therapeutic strategies in cystic fibrosis: Antimicrobial and anti-biofilm activity of natural and designed α-helical peptides against Staphylococcus aureus, Pseudomonas aeruginosa, and Stenotrophomonas maltophilia. BMC Microbiol. 2012, 12, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pompilio, A.; Pomponio, S.; Crocetta, V.; Gherardi, G.; Verginelli, F.; Fiscarelli, E.V.; Dicuonzo, G.; Savini, V.; D’Antonio, D.; Di Bonaventura, G. Phenotypic and genotypic characterization of Stenotrophomonas maltophilia isolates from patients with cystic fibrosis: Genome diversity, biofilm formation, and virulence. BMC Microbiol. 2011, 11, 159. [Google Scholar] [CrossRef] [Green Version]
- Savini, V.; Catavitello, C.; D’Aloisio, M.; Balbinot, A.; Astolfi, D.; Masciarelli, G.; Pompilio, A.; Di Bonaventura, G.; D’Antonio, D. Chloramphenicol and rifampin may be the only options against Stenotrophomonas maltophilia. A tale of a colonized bladder device in a patient with myelofibrosis. Infez. Med. 2010, 18, 193–197. [Google Scholar]
- Pompilio, A.; Crocetta, V.; Confalone, P.; Nicoletti, M.; Petrucca, A.; Guarnieri, S.; Fiscarelli, E.V.; Savini, V.; Piccolomini, R.; Di Bonaventura, G. Adhesion to and biofilm formation on IB3-1 bronchial cells by Stenotrophomonas maltophilia isolates from cystic fibrosis patients. BMC Microbiol. 2010, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- Pompilio, A.; Catavitello, C.; Picciani, C.; Confalone, P.; Piccolomini, R.; Savini, V.; Fiscarelli, E.; D’Antonio, D.; Di Bonaventura, G. Subinhibitory concentrations of moxifloxacin decrease adhesion and biofilm formation of Stenotrophomonas maltophilia from cystic fibrosis. J. Med. Microbiol. 2010, 59, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Pompilio, A.; Piccolomini, R.; Picciani, C.; D’Antonio, D.; Savini, V.; Di Bonaventura, G. Factors associated with adherence to and biofilm formation on polystyrene by Stenotrophomonas maltophilia: The role of cell surface hydrophobicity and motility. FEMS Microbiol. Lett. 2008, 287, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Di Bonaventura, G.; Prosseda, G.; Del Chierico, F.; Cannavacciuolo, S.; Cipriani, P.; Petrucca, A.; Superti, F.; Ammendolia, M.; Concato, C.; Fiscarelli, E.; et al. Molecular characterization of virulence determinants of Stenotrophomonas maltophilia strains isolated from patients affected by cystic fibrosis. Int. J. Immunopathol. Pharmacol. 2007, 20, 529–537. [Google Scholar] [CrossRef] [Green Version]
- Di Bonaventura, G.; Stepanović, S.; Picciani, C.; Pompilio, A.; Piccolomini, R. Effect of environmental factors on biofilm formation by clinical Stenotrophomonas maltophilia isolates. Folia Microbiol. 2007, 52, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Di Bonaventura, G.; Spedicato, I.; D’Antonio, D.; Robuffo, I.; Piccolomini, R. Biofilm formation by Stenotrophomonas maltophilia: Modulation by quinolones, trimethoprim-sulfamethoxazole, and ceftazidime. Antimicrob. Agents Chemother. 2004, 48, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcaraz, E.; Garcia, C.; Papalia, M.; Vay, C.; Friedman, L.; De Rossi, B.P. Stenotrophomonas maltophilia isolated from patients exposed to invasive devices in a university hospital in Argentina: Molecular typing, susceptibility and detection of potential virulence factors. J. Med. Microbiol. 2018, 67, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Flores-Treviño, S.; Gutiérrez-Ferman, J.L.; Morfín-Otero, R.; Rodríguez-Noriega, E.; Estrada-Rivadeneyra, D.; Rivas-Morales, C.; Llaca-Díaz, J.M.; Camacho-Ortíz, A.; Mendoza-Olazarán, S.; Garza-González, E. Stenotrophomonas maltophilia in Mexico: Antimicrobial resistance, biofilm formation and clonal diversity. J. Med. Microbiol. 2014, 63, 1524–1530. [Google Scholar] [CrossRef] [PubMed]
- Madi, H.; Lukić, J.; Vasiljević, Z.; Biočanin, M.; Kojić, M.; Jovčić, B.; Lozo, J. Genotypic and phenotypic characterization of Stenotrophomonas maltophilia strains from a pediatric tertiary care hospital in Serbia. PLoS ONE 2016, 11, e0165660. [Google Scholar] [CrossRef]
- Sun, E.; Liang, G.; Wang, L.; Wei, W.; Lei, M.; Song, S.; Han, R.; Wang, Y.; Qi, W. Antimicrobial susceptibility of hospital acquired Stenotrophomonas maltophilia isolate biofilms. Braz. J. Infect. Dis. 2016, 20, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Córdova, A.; Mancilla-Rojano, J.; Luna-Pineda, V.M.; Escalona-Venegas, G.; Cázares-Domínguez, V.; Ormsby, C.; Franco-Hernández, I.; Zavala-Vega, S.; Hernández, M.A.; Medina-Pelcastre, M.; et al. Molecular epidemiology, antibiotic resistance, and virulence traits of Stenotrophomonas maltophilia strains associated with an outbreak in a Mexican tertiary care hospital. Front. Cell. Infect. Microbiol. 2020, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Pompilio, A.; Savini, V.; Fiscarelli, E.V.; Gherardi, G.; Di Bonaventura, G. Clonal diversity, biofilm formation, and antimicrobial resistance among Stenotrophomonas maltophilia strains from cystic fibrosis and non-cystic fibrosis patients. Antibiotics 2020, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Nys, C.; Cherabuddi, K.; Venugopalan, V.; Klinker, K.P. Clinical and microbiologic outcomes in patients with monomicrobial Stenotrophomonas maltophilia infections. Antimicrob. Agents Chemother. 2019, 63, 00788–00819. [Google Scholar] [CrossRef]
- Duan, Z.; Qin, J.; Li, C.; Ying, C. Clinical and molecular epidemiology of Stenotrophomonas maltophilia in pediatric patients from a Chinese teaching hospital. Front. Cell. Infect. Microbiol. 2020, 10, 411. [Google Scholar] [CrossRef]
- Bostanghadiri, N.; Ghalavand, Z.; Fallah, F.; Yadegar, A.; Ardebili, A.; Tarashi, S.; Pournajaf, A.; Mardaneh, J.; Shams, S.; Hashemi, A. Characterization of phenotypic and genotypic diversity of Stenotrophomonas maltophilia strains isolated from selected hospitals in Iran. Front. Microbiol. 2019, 10, 1191. [Google Scholar] [CrossRef] [PubMed]
- Garner, J.S.; Jarvis, W.R.; Emori, T.; Horan, T.C.; Hughes, J.M. CDC definitions for nosocomial infections, 1988. Am. J. Infect. Control 1988, 16, 128–140. [Google Scholar] [CrossRef]
- Sonnenwirth, A.C. Stains and staining procedures. In Gradwohl’s Clinical Laboratory Methods and Diagnosis, 8th ed.; Sonnenwirth, A.C., Jarett, L., Eds.; C.V. Mosby Co.: St. Louis, MO, USA, 1980; p. 380. [Google Scholar]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 26th ed.; CLSI Supplement M100S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016; ISBN 1-56238-923-8/1-56238-924-6. [Google Scholar]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.J.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Soto, S.M.; Smithson, A.; Martinez, J.; Horcajada, J.; Mensa, J.; Vila, J. Biofilm formation in uropathogenic Escherichia coli strains: Relationship with prostatitis, urovirulence factors and antimicrobial resistance. J. Urol. 2007, 177, 365–368. [Google Scholar] [CrossRef]
- Salo, J.; Sevander, J.-J.; Tapiainen, T.; Ikäheimo, I.; Pokka, T.; Koskela, M.; Uhari, M. Biofilm formation by Escherichia coli isolated from patients with urinary tract infections. Clin. Nephrol. 2009, 71, 501–507. [Google Scholar] [CrossRef]
- Pompilio, A.; Crocetta, V.; Savini, V.; Petrelli, D.; Di Nicola, M.; Bucco, S.; Amoroso, L.; Bonomini, M.; Di Bonaventura, G. Phylogenetic relationships, biofilm formation, motility, antibiotic resistance and extended virulence genotypes among Escherichia coli strains from women with community-onset primitive acute pyelonephritis. PLoS ONE 2018, 13, e0196260. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Solano, L.; Macia, M.D.; Fajardo, A.; Oliver, A.; Martinez, J.L. Chronic Pseudomonas aeruginosa infection in chronic obstructive pulmonary disease. Clin. Infect. Dis. 2008, 47, 1526–1533. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.-K.; Liaw, S.-J.; Yu, C.-J.; Teng, L.-J.; Hsueh, P.-R. Extensively drug-resistant Stenotrophomonas maltophilia in a tertiary care hospital in Taiwan: Microbiologic characteristics, clinical features, and outcomes. Diagn. Microbiol. Infect. Dis. 2008, 60, 205–210. [Google Scholar] [CrossRef]
- Steinmann, J.; Mamat, U.; Abda, E.M.; Kirchhoff, L.; Streit, W.R.; Schaible, U.E.; Niemann, S.; Kohl, T.A. Analysis of phylogenetic variation of Stenotrophomonas maltophilia reveals human-specific branches. Front. Microbiol. 2018, 9, 806. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Cha, M.K.; Kang, C.-I.; Ko, J.-H.; Huh, K.; Cho, S.Y.; Chung, O.R.; Peck, K.R. Pathogenic significance of hemorrhagic pneumonia in hematologic malignancy patients with Stenotrophomonas maltophilia bacteremia: Clinical and microbiological analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 38, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Willsey, G.G.; Eckstrom, K.; LaBauve, A.E.; Hinkel, L.A.; Schutz, K.; Meagher, R.J.; Lipuma, J.J.; Wargo, M.J. Stenotrophomonas maltophilia differential gene expression in synthetic cystic fibrosis sputum reveals shared and cystic fibrosis strain-specific responses to the sputum environment. J. Bacteriol. 2019, 201, 00074–00119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gherardi, G.; Creti, R.; Pompilio, A.; Di Bonaventura, G. An overview of various typing methods for clinical epidemiology of the emerging pathogen Stenotrophomonas maltophilia. Diagn. Microbiol. Infect. Dis. 2015, 81, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Welcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Yang, W.; Meng, J.; Lv, Y.; Wang, J.; Huang, B. Co-infection of Pseudomonas aeruginosa and Stenotrophomonas maltophilia in hospitalized pneumonia patients has a synergic and significant impact on clinical outcomes. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2231–2235. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-M.; Yu, J.; Liu, G.-X.; Hu, L.-Y.; Li, L.; Li, F.; Yang, H. Electron microscopic analysis of biofilm on tracheal tubes removed from intubated neonates and the relationship between biofilm and lower respiratory infection. Chin. J. Pediatr. 2007, 45, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Barros, C.E.D.; Almeida, J.A.D.; Ayres, G.H.D.S.; Oliveira, C.G.D.; Braga, C.A.D.S.B.; Avelino, M.A.G. Pediatric tracheostomy: Epidemiology and characterization of tracheal secretion—A literature review. Rev. Assoc. Méd. Bras. 2019, 65, 1502–1507. [Google Scholar] [CrossRef]
- Donlan, R.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Cepas, V.; López, Y.; Muñoz, E.; Rolo, D.; Ardanuy, C.; Martí, S.; Xercavins, M.; Horcajada, J.P.; Bosch, J.; Soto, S.M. Relationship between biofilm formation and antimicrobial resistance in gram-negative bacteria. Microb. Drug Resist. 2019, 25, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Xia, J.; Xu, Y.; Gong, M.; Zhou, Y.; Xie, L.; Fang, X. Biological features of biofilm-forming ability of Acinetobacter baumannii strains derived from 121 elderly patients with hospital-acquired pneumonia. Clin. Exp. Med. 2014, 16, 73–80. [Google Scholar] [CrossRef]
- Fàbrega, A.; Soto, S.M.; Ballesté-Delpierre, C.; Fernández-Orth, D.; Jiménez de Anta, M.T.; Vila, J. Impact of quinolone-resistance acquisition on biofilm production and fitness in Salmonella enterica. J. Antimicrob. Chemother. 2014, 69, 1815–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.L.; Scipione, M.R.; Dubrovskaya, Y.; Papadopoulos, J. Monotherapy with fluoroquinolone or trimethoprim-sulfamethoxazole for treatment of Stenotrophomonas maltophilia infections. Antimicrob. Agents Chemother. 2014, 58, 176–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Baño, J.; Martí, S.; Soto, S.; Fernández-Cuenca, F.; Cisneros, J.; Pachón, J.; Pascual, A.; Martínez-Martínez, L.; McQueary, C.; Actis, L.; et al. Biofilm formation in Acinetobacter baumannii: Associated features and clinical implications. Clin. Microbiol. Infect. 2008, 14, 276–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmberg, A.; Lood, R.; Mörgelin, M.; Söderquist, B.; Holst, E.; Collin, M.; Christensson, B.; Rasmussen, M. Biofilm formation by Propionibacterium acnes is a characteristic of invasive isolates. Clin. Microbiol. Infect. 2009, 15, 787–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuck, A.-M.; Johnson, J.R.; Dunny, G.M. A Widely used in vitro biofilm assay has questionable clinical significance for enterococcal endocarditis. PLoS ONE 2014, 9, e107282. [Google Scholar] [CrossRef]
- McDaniel, M.S.; Schoeb, T.; Swords, W.E. Cooperativity between Stenotrophomonas maltophilia and Pseudomonas aeruginosa during polymicrobial airway infections. Infect. Immun. 2020, 88, 00855-19. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. (%) of S. maltophilia with the Following Etiological Role: a | |||||
|---|---|---|---|---|---|
| Overall (n = 109) | Definite (n = 23) | Probable (n = 45) | Possible (n = 8) | Non-Pathogen (n = 33) | |
| Isolation Site | |||||
| Airways (CF b and non-CF) | 64 (58.7) | (21.7) | 34 (75.5) | 6 (75.0) | 19 (57.6) |
| Blood | 23 (21.1) | 14 (60.8) | 8 (17.7) | 1 (12.5) | |
| Wound | 14 (12.8) | 2 (8.7) | 2 (4.4) | 1 (12.5) | 9 (27.3) |
| Urine | 6 (5.5) | 1 (2.2) | 5 (15.1) | ||
| CVC c | 2 (1.8) | 2 (8.7) | |||
| Source of Infection d | |||||
| Community-acquired | 41 (44.1) | 3 (7.3) | 22 (53.6) | 2 (4.9) | 14 (34.1) |
| Hospital-acquired | 52 (55.9) | 18 (34.6) | 11 (21.1) | 4 (7.7) | 19 (36.5) |
| Patient | |||||
| Age (mean ± SD) | 43.8 ± 28.3 | 58.8 ± 25.5 | 27.5 ± 22.4 | 65.7 ± 14.1 | 49.8 ± 28.8 |
| Gender (Male) | 63 (57.8) | 15 (65.2) | 16 (35.5) | 6 (75.0) | 24 (72.7) |
| Clinical Diagnosis | |||||
| CF | 32 (29.3) | 32 (71.1) | |||
| Non-CF airways infection | 15 (13.7) | 5 (21.7) | 3 (6.6) | 5 (62.5) | 2 (6.0) |
| Neoplasia e | 8 (7.3) | 3 (13.0) | 2 (4.4) | 1 (12.5) | 2 (6.0) |
| Wound infection | 6 (5.5) | 1 (4.3) | 1 (2.2) | 4 (12.1) | |
| Sepsis | 6 (5.5) | 2 (8.7) | 3 (6.6) | 1 (3.0) | |
| CVC infection | 4 (3.6) | 3 (13.0) | 1 (2.2) | ||
| Intracranial injury/bleeding | 4 (3.6) | 1 (4.3) | 3 (9.1) | ||
| Other f | 34 (31.2) | 8 (34.8) | 4 (4.4) | 2 (25.0) | 20 (60.6) |
| Risk Factors | |||||
| Previous antibiotic therapy | 55 (50.4) | 15 (65.2) | 18 (40.0) | 6 (75.0) | 16 (48.5) |
| Prolonged hospital/ICU stay | 17 (15.6) | 3 (13.0) | 3 (6.6) | 1 (12.5) | 10 (30.3) |
| Chemotherapy | 12 (11.0) | 4 (17.4) | 3 (6.6) | 1 (12.5) | 4 (12.1) |
| Mechanical ventilation | 11 (10.1) | 3 (13.0) | 1 (2.2) | 7 (21.2) | |
| Outcome after Antibiotic Therapy g | |||||
| Cleared | 40 (65.6) g | 16 (69.6) | 20 (44.4) | 4 (50.0) | NA h |
| Uncleared | 21 (34.4) g | 6 (26.0) | 12 (26.7) | 3 (37.5) | NA |
| Strain | Sequence Type | Etiological Role | Biofilm Class | Isolation Country |
|---|---|---|---|---|
| STMA_CZ_44 | 4 | Definite | Strong | Czech Republic |
| STMA_CZ_B1799 | 233 | Definite | Strong | Czech Republic |
| STMA_CZ_25 | 183 | Probable | Strong | Czech Republic |
| STMA_CZ_30 | 211 | Probable | Moderate | Czech Republic |
| STMA_CZ_4 | 321 | Possible | Weak | Czech Republic |
| STMA_CZ_70 | 4 | Non-pathogen | Strong | Czech Republic |
| BG1 | 239 | Definite | Moderate | Serbia |
| BG10 | 26 | Definite | Strong | Serbia |
| BG11 | 249 | Definite | Strong | Serbia |
| BG9 | 371 | Probable | Strong | Serbia |
| BG14 | 26 | Probable | Strong | Serbia |
| BG8 | 186 | Non-pathogen | Weak | Serbia |
| 94463 | 295 | Definite | Strong | Spain |
| 676342 | 321 | Definite | Strong | Spain |
| 24LP | 28 | Definite | Moderate | Spain |
| 7LP | 26 | Probable | Moderate | Spain |
| 21LP | 26 | Possible | Weak | Spain |
| 29LP | 28 | Non-pathogen | Strong | Spain |
| 00623 | 77 | Definite | Moderate | Germany |
| 00410 | 28 | Definite | Strong | Germany |
| 00416 | 265 | Definite | Strong | Germany |
| 00610 | 174 | Definite | Strong | Germany |
| 00630 | 28 | Definite | Strong | Germany |
| 01052 | 24 | Probable | Strong | Germany |
| 9CM | 319 | Probable | Strong | Italy |
| 7BK | 93 | Probable | Moderate | Italy |
| 48SJ | 219 | Probable | Strong | Italy |
| 50TE | 172 | Probable | Moderate | Italy |
| Strain | Source | MIC | MBIC | MBIC/MIC | MBC | MBEC | MBEC/MBC |
|---|---|---|---|---|---|---|---|
| Definite Pathogen | |||||||
| STMA_CZ44 | non-CF airways | 1/19 | 32/608 | 32 | 16/304 | >32/608 | >2 |
| BG3 | non-CF airways | 0.25/4.75 | >32/608 | >128 | 1/19 | >32/608 | >32 |
| BG10 | non-CF airways | 0.25/4.75 | >32/608 | >128 | 0.5/9.5 | >32/608 | >64 |
| BG11 | non-CF airways | 0.25/4.75 | >32/608 | >128 | 4/76 | >32/60 | >8 |
| 00422 | blood | 1/19 | 32/608 | 128 | 4/76 | >32/608 | >8 |
| 00610 | blood | 2/38 | >32/608 | >16 | 8/152 | >32/608 | >4 |
| 00624 | blood | 2/38 | >32/608 | >16 | 8/152 | >32/608 | >4 |
| 00630 | blood | 2/38 | >32/608 | >16 | 8/152 | >32/608 | >4 |
| 01052 | blood | 1/19 | >32/608 | >32 | 8/152 | >32/608 | >4 |
| Probable Pathogen | |||||||
| BG9 | non-CF airways | 1/19 | 32/608 | 32 | 8/152 | >32/608 | >4 |
| BG12 | non-CF airways | 0.25/4.75 | 64/1216 | 256 | 4/76 | 64/1216 | 16 |
| BG14 | wound | 0.125/2.375 | 32/608 | 256 | 0.5/9.5 | >32/608 | >64 |
| STMA_CZ20 | CF airways | 1/19 | 32/608 | 32 | 4/76 | >32/608 | >8 |
| Possible pathogen | |||||||
| 00213 | blood | 0.5/9.5 | >32/608 | >64 | 4/76 | >32/608 | >8 |
| 00634 | blood | 1/19 | >32/608 | >32 | 8/152 | >32/608 | >4 |
| Non-pathogen | |||||||
| STMA_CZ2 | non-CF airways | 1/19 | >32/608 | >32 | 32/608 | >32/608 | >1 |
| Strain | Source | MIC | MBIC | MBIC/MIC | MBC | MBEC | MBEC/MBC |
|---|---|---|---|---|---|---|---|
| Definite Pathogen | |||||||
| STMA_CZ44 | non-CF airways | 1 | 1 | 1 | 1 | 32 | 32 |
| BG3 | non-CF airways | 0.25 | >256 | >1024 | 0.5 | 4 | 8 |
| BG10 | non-CF airways | 0.5 | 4 | 8 | 1 | >128 | >128 |
| BG11 | non-CF airways | 0.5 | 2 | 4 | 2 | 128 | 64 |
| 00422 | blood | 1 | 8 | 8 | 2 | >256 | >128 |
| 00610 | blood | 1 | 4 | 4 | 4 | 512 | 128 |
| 00624 | blood | 0.5 | 2 | 4 | 1 | >128 | >128 |
| 00630 | blood | 1 | 8 | 8 | 1 | 128 | 128 |
| 01052 | blood | 1 | 256 | 256 | 2 | >256 | >128 |
| Probable Pathogen | |||||||
| BG9 | non-CF airways | 2 | 4 | 2 | 2 | 256 | 128 |
| BG12 | non-CF airways | 1 | 1 | 1 | 1 | 128 | 128 |
| BG14 | wound | 1 | 16 | 16 | 1 | >128 | >128 |
| STMA_CZ20 | CF airways | 1 | 1 | 1 | 1 | 8 | 8 |
| Possible pathogen | |||||||
| 00213 | blood | 2 | >256 | >128 | 2 | >256 | >128 |
| 00634 | blood | 1 | 2 | 2 | 2 | >256 | >128 |
| Non-Pathogen | |||||||
| STMA_CZ2 | non-CF airways | 1 | 4 | 4 | 1 | >128 | >128 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pompilio, A.; Ranalli, M.; Piccirilli, A.; Perilli, M.; Vukovic, D.; Savic, B.; Krutova, M.; Drevinek, P.; Jonas, D.; Fiscarelli, E.V.; et al. Biofilm Formation among Stenotrophomonas maltophilia Isolates Has Clinical Relevance: The ANSELM Prospective Multicenter Study. Microorganisms 2021, 9, 49. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010049
Pompilio A, Ranalli M, Piccirilli A, Perilli M, Vukovic D, Savic B, Krutova M, Drevinek P, Jonas D, Fiscarelli EV, et al. Biofilm Formation among Stenotrophomonas maltophilia Isolates Has Clinical Relevance: The ANSELM Prospective Multicenter Study. Microorganisms. 2021; 9(1):49. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010049
Chicago/Turabian StylePompilio, Arianna, Marco Ranalli, Alessandra Piccirilli, Mariagrazia Perilli, Dragana Vukovic, Branislava Savic, Marcela Krutova, Pavel Drevinek, Daniel Jonas, Ersilia V. Fiscarelli, and et al. 2021. "Biofilm Formation among Stenotrophomonas maltophilia Isolates Has Clinical Relevance: The ANSELM Prospective Multicenter Study" Microorganisms 9, no. 1: 49. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010049