Bacterial Diversity Analysis and Evaluation Proteins Hydrolysis during the Acid Whey and Fish Waste Fermentation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials
2.2. Selection of Proteolytic Strain
2.3. Fermentation
2.4. Proteolytic Activity
2.5. pH Measurement and Bacterial Counts
2.6. Protein Analyses
2.6.1. Protein Concentration Assay
2.6.2. Free Amino Acids and Peptide Analysis
2.7. Microbial Community Analyses
2.7.1. DNA Extraction and Quality Assessment
2.7.2. DNA Library Preparation and Sequencing
2.7.3. 16S-v4 Annotation
2.8. Statistical Analyses
2.9. Accession Number
3. Results
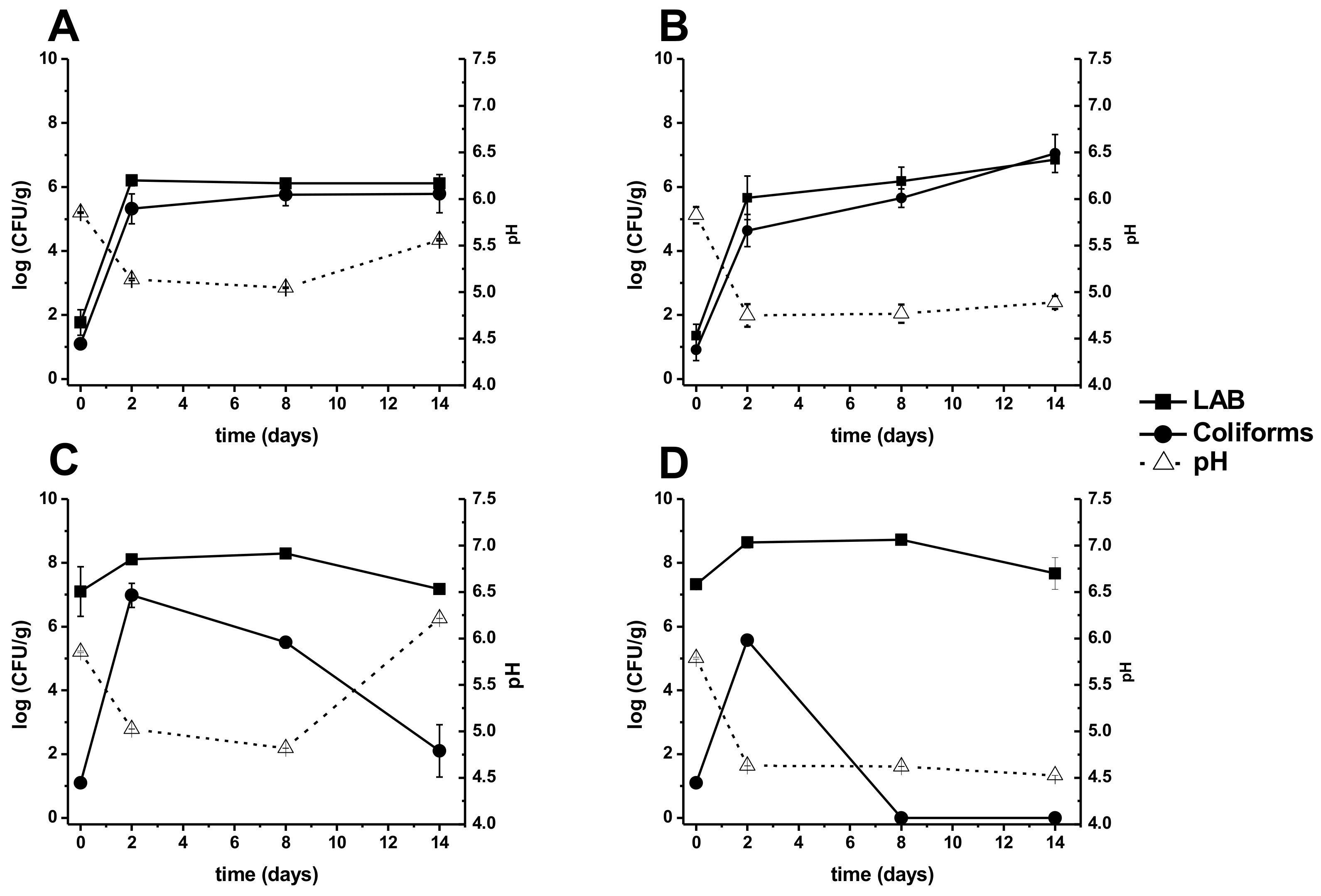
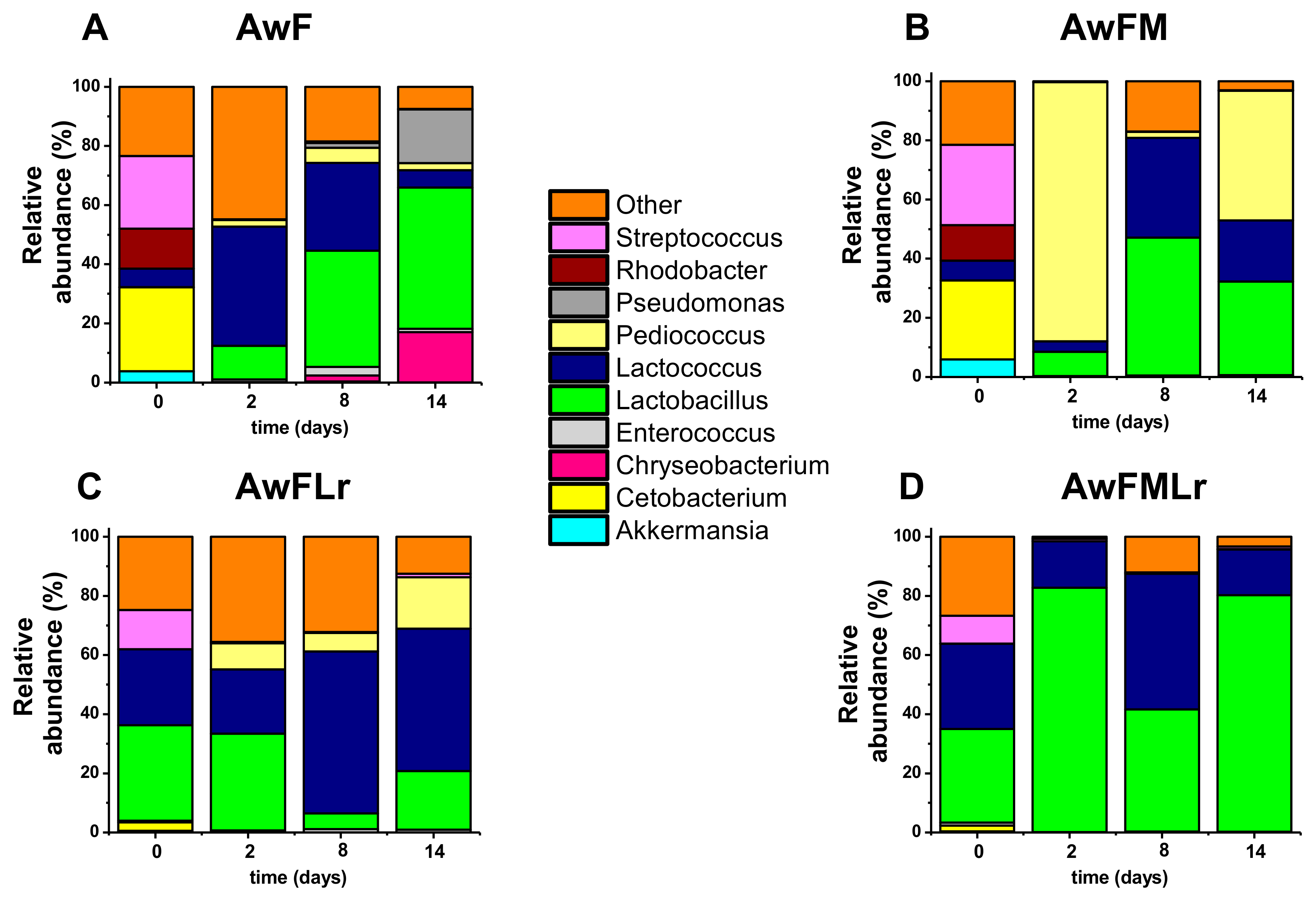
3.1. pH and Microbiology Community
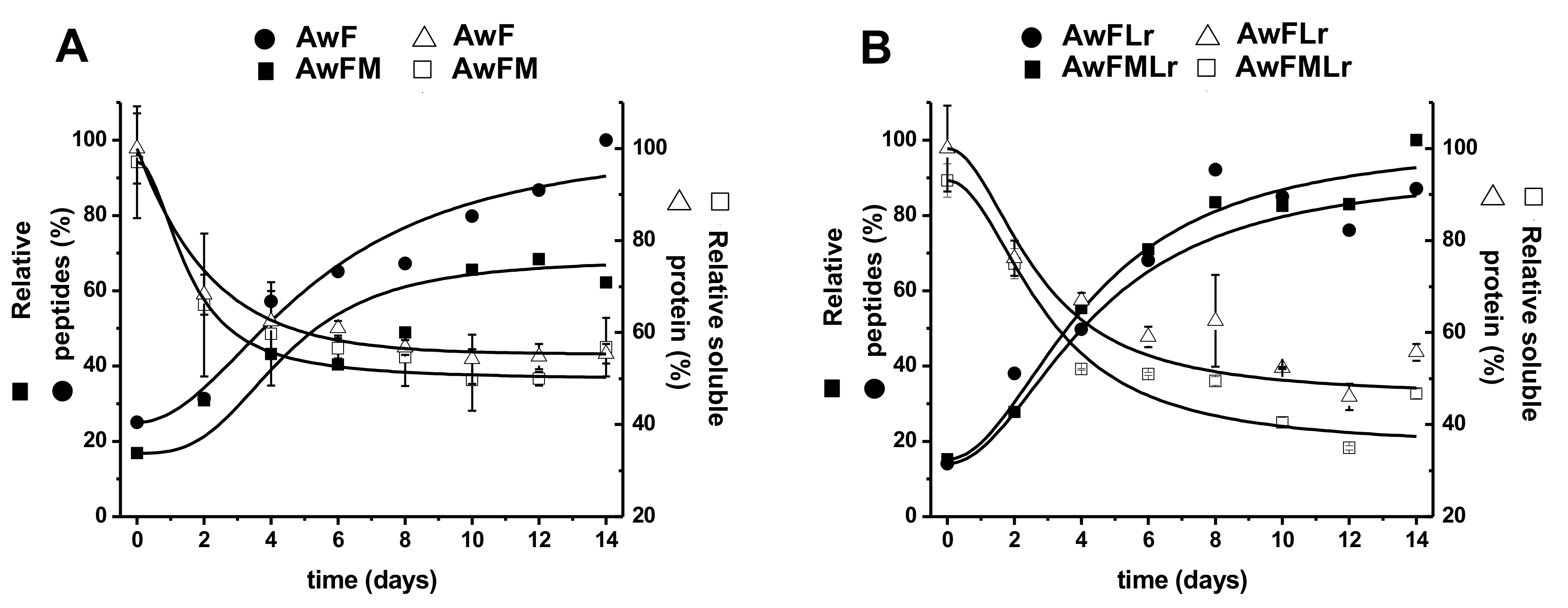
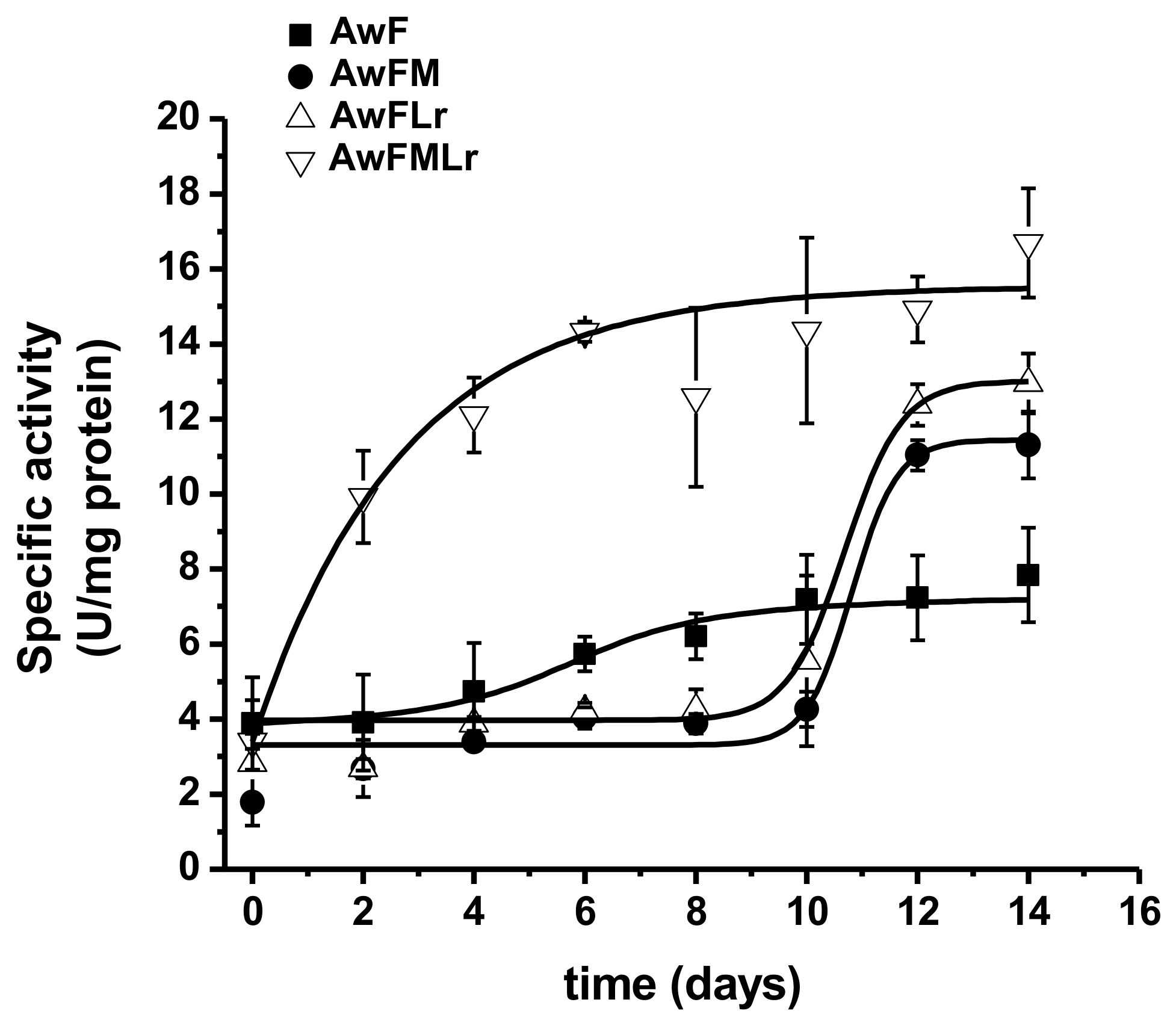
3.2. Protein Dynamics Through Fermentation
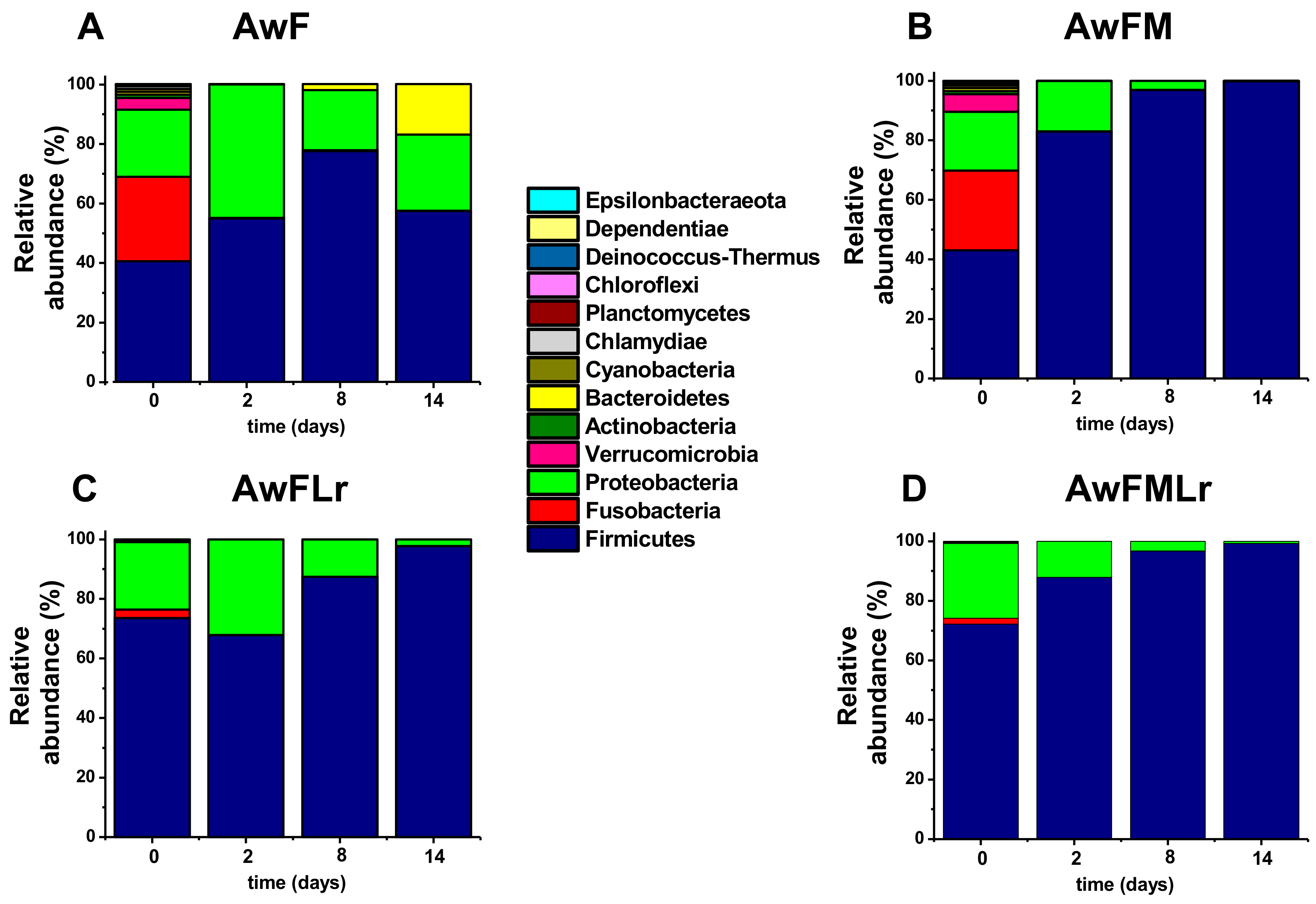
3.3. Metagenomic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chandrapala, J.; Duke, M.C.; Gray, S.R.; Weeks, M.; Palmer, M.; Vasiljevic, T. Nanofiltration and nanodiafiltration of acid whey as a function of pH and temperature. Sep. Purif. Technol. 2016, 160, 18–27. [Google Scholar] [CrossRef]
- Wherry, B.; Barbano, D.M.; Drake, M.A. Use of acid whey protein concentrate as an ingredient in nonfat cup set-style yogurt. J. Dairy Sci. 2019, 102, 8768–8784. [Google Scholar] [CrossRef] [PubMed]
- De Wit, J. Lecturer’s Handbook on Whey and Whey Products, 1st ed.; European Whey Products Association (EWPA): Brussels, Belgium, 2001; pp. 11–15. [Google Scholar]
- Prazeres, A.R.; Carvalho, F.; Rivas, J. Cheese whey management: A review. J. Environ Manag. 2012, 110, 48–68. [Google Scholar] [CrossRef] [PubMed]
- González-García, S.; Castanheira, É.G.; Dias, A.C.; Arroja, L. Environmental life cycle assessment of a dairy product: The yoghurt. Int. J. Life Cycle Assess 2012, 18, 796–811. [Google Scholar] [CrossRef]
- Siso, M.G. The biotechnological utilization of cheese whey: A review. Bioresour. Technol. 1996, 57, 1–11. [Google Scholar] [CrossRef]
- Cheison, S.C.; Kulozik, U. Impact of the environmental conditions and substrate pre-treatment on whey protein hydrolysis: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 418–453. [Google Scholar] [CrossRef]
- Liaset, B.; Lied, E.; Espe, M. Enzymatic hydrolysis of by-products from the fish-filleting industry; chemical characterisation and nutritional evaluation. J. Sci. Food Agric. 2000, 80, 581–589. [Google Scholar] [CrossRef]
- Rustad, T.; Storrø, I.; Slizyte, R. Possibilities for the utilisation of marine by-products. Int. J. Food Sci. Technol. 2011, 46, 2001–2014. [Google Scholar] [CrossRef]
- Rai, A.K.; Swapna, H.C.; Bhaskar, N.; Halami, P.M.; Sachindra, N.M. Effect of fermentation ensilaging on recovery of oil from freshwater fish viscera. Enzym. Microb. Technol. 2010, 46, 9–13. [Google Scholar] [CrossRef]
- Bhaskar, N.; Benila, T.; Radha, C.; Lalitha, R.G. Optimization of enzymatic hydrolysis of visceral waste proteins of Catla (Catla catla) for preparing protein hydrolysate using a commercial protease. Bioresour. Technol. 2008, 99, 335–343. [Google Scholar] [CrossRef]
- Arason, S.; Karlsdottir, M.; Valsdottir, T.; Slizyte, R.; Rustad, T.; Falch, E.; Eysturskarð, J.; Jakobsen, G. Maximum Resource Utilization—Value Added Fish by-Products; Nordic Council of Ministers: Copenhagen, Denmark, 2010; pp. 4–6. [Google Scholar]
- Tacon, A.G.J. Trends in Global Aquaculture and Aquafeed Production: 2000–2017. Rev Fish Sci. Aquac. 2019, 28, 43–56. [Google Scholar] [CrossRef]
- Villamil, O.; Vaquiro, H.; Solanilla, J.F. Fish viscera protein hydrolysates: Production, potential applications and functional and bioactive properties. Food Chem. 2017, 224, 160–171. [Google Scholar] [CrossRef]
- Yoshioka, T.; Konno, Y.; Konno, K. Below-zero storage of fish to suppress loss of freshness. Fish Sci. 2019, 85, 601–609. [Google Scholar] [CrossRef]
- Juarez-Castelan, C.; Garcia-Cano, I.; Escobar-Zepeda, A.; Azaola-Espinosa, A.; Alvarez-Cisneros, Y.; Ponce-Alquicira, E. Evaluation of the bacterial diversity of Spanish-type chorizo during the ripening process using high-throughput sequencing and physicochemical characterization. Meat Sci. 2019, 150, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cano, I.; Rocha-Mendoza, D.; Ortega-Anaya, J.; Wang, K.; Kosmerl, E.; Jimenez-Flores, R. Lactic acid bacteria isolated from dairy products as potential producers of lipolytic, proteolytic and antibacterial proteins. Appl. Microbiol. Biotechnol. 2019, 103, 5243–5257. [Google Scholar] [CrossRef] [Green Version]
- Gitton, C.; Meyrand, M.; Wang, J.; Caron, C.; Trubuil, A.; Guillot, A.; Mistou, M.Y. Proteomic signature of Lactococcus lactis NCDO763 cultivated in milk. Appl. Environ. Microbiol. 2005, 71, 7152–7163. [Google Scholar] [CrossRef] [Green Version]
- Mazzucotelli, C.A.; Ponce, A.G.; Kotlar, C.E.; Moreira, M. d. R. Isolation and characterization of bacterial strains with a hydrolytic profile with potential use in bioconversion of agroindustial by-products and waste. Food Sci. Technol. 2013, 33, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Xia, W.; Gao, P.; Xu, Y. Sarcoplasmic Protein Hydrolysis Activity of Lactobacillus plantarum 120 Isolated from Suanyu: A Traditional Chinese Low Salt Fermented Fish. J. Food Process. Pres. 2017, 41, e12821. [Google Scholar] [CrossRef]
- Jini, R.; Swapna, H.C.; Amit Kumar, R.; Halami, P.M.; Sachindra, N.M. Isolation and characterization of potential lactic acid bacteria (LAB) from freshwater fish processing wastes for application in fermentative utilisation of fish processing waste. Braz. J. Microbiol. 2011, 42, 1516–1525. [Google Scholar]
- Qureshi, A.S.; Khushk, I.; Ali, C.H.; Chisti, Y.; Ahmad, A.; Majeed, H. Coproduction of protease and amylase by thermophilic Bacillus sp. BBXS-2 using open solid-state fermentation of lignocellulosic biomass. Biocatal. Agric. Biotechnol. 2016, 8, 146–151. [Google Scholar] [CrossRef]
- Garcia-Cano, I.; Rocha-Mendoza, D.; Kosmerl, E.; Zhang, L.; Jimenez-Flores, R. Technically relevant enzymes and proteins produced by LAB suitable for industrial and biological activity. Appl. Microbiol. Biotechnol. 2020, 104, 1401–1422. [Google Scholar] [CrossRef] [PubMed]
- Anson, M.L. The estimation of pepsin, trypsin, papain, and cathepsin with hemoglobin. J. Gen. Physiol. 1938, 22, 79. [Google Scholar] [CrossRef] [PubMed]
- Doi, E.; Shibata, D.; Matoba, T. Modified colorimetric ninhydrin methods for peptidase assay. Anal. Biochem. 1981, 118, 173–184. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Xia, W.S.; Zhang, X.W.; Xu, Y.S.; Jiang, Q.X. A comparison of endogenous and microbial proteolytic activities during fast fermentation of silver carp inoculated with Lactobacillus plantarum. Food Chem. 2016, 207, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Chandrapala, J.; Duke, M.C.; Gray, S.R.; Weeks, M.; Palmer, M.; Vasiljevic, T. Strategies for maximizing removal of lactic acid from acid whey—Addressing the un-processability issue. Sep. Purif. Technol. 2017, 172, 489–497. [Google Scholar] [CrossRef]
- Chandrapala, J.; Chen, G.Q.; Kezia, K.; Bowman, E.G.; Vasiljevic, T.; Kentish, S.E. Removal of lactate from acid whey using nanofiltration. J. Food Eng. 2016, 177, 59–64. [Google Scholar] [CrossRef]
- Davies, S.J.; Guroy, D.; Hassaan, M.S.; El-Ajnaf, S.M.; El-Haroun, E. Evaluation of co-fermented apple-pomace, molasses and formic acid generated sardine based fish silages as fishmeal substitutes in diets for juvenile European sea bass (Dicentrachus labrax) production. Aquaculture 2020, 521, 735087. [Google Scholar] [CrossRef]
- Hasan, B. Fermentation of fish silage using Lactobacillus pentosus. J. Nat. Indones. 2003, 6, 11–15. [Google Scholar]
- Kröckel, L. Bacterial fermentation of meats. In Fermented Meats; Campbell-Platt, G., Cook, P.E., Eds.; Springer: Cambridge, MA, USA, 1995; pp. 69–109. [Google Scholar]
- Olsen, R.L.; Toppe, J. Fish silage hydrolysates: Not only a feed nutrient, but also a useful feed additive. Trends Food Sci. Technol. 2017, 66, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, J.A.; Nogueira, M.; Durán, A.; Prieto, M.A.; Rodríguez-Amado, I.; Rial, D.; González, M.P.; Murado, M.A. Preparation of marine silage of swordfish, ray and shark visceral waste by lactic acid bacteria. J. Food Eng. 2011, 103, 442–448. [Google Scholar] [CrossRef] [Green Version]
- Fadda, S.; Vignolo, G.; Holgado, A.P.; Oliver, G. Proteolytic activity of Lactobacillus strains isolated from dry fermented sausages on muscle sarcoplasmic proteins. Meat Sci. 1998, 49, 11–18. [Google Scholar] [CrossRef]
- Guan, L.; Cho, K.H.; Lee, J.H. Analysis of the cultivable bacterial community in jeotgal, a Korean salted and fermented seafood, and identification of its dominant bacteria. Food Microbiol. 2011, 28, 101–113. [Google Scholar] [CrossRef]
- Samaddar, A.; Kaviraj, A. Processing of fish offal waste through fermentation utilizing whey as inoculum. Int. J. Recycl. Org. Waste Agric. 2014, 3, 45. [Google Scholar] [CrossRef] [Green Version]
- Ozyurt, G.; Özkütük, A.; Boğa, M.; Durmuş, M.; Boğa, K.E. Biotransformation of seafood processing wastes fermented with natural lactic acid bacteria; the quality of fermented products and their use in animal feeding. Turk. J. Fish. Aquat. Sci. 2016, 17, 543–555. [Google Scholar]
- van’t Land, M.; Vanderperren, E.; Raes, K. The effect of raw material combination on the nutritional composition and stability of four types of autolyzed fish silage. Anim. Feed Sci. Technol. 2017, 234, 284–294. [Google Scholar] [CrossRef]
- Bhaskar, N.; Suresh, P.V.; Sakhare, P.Z.; Sachindra, N.M. Shrimp biowaste fermentation with Pediococcus acidilactici CFR2182: Optimization of fermentation conditions by response surface methodology and effect of optimized conditions on deproteination/demineralization and carotenoid recovery. Enzym. Microb. Technol. 2007, 40, 1427–1434. [Google Scholar] [CrossRef]
- Raa, J.; Gildberg, A. Fish silage: A review. Crit. Rev. Food Sci. Nutr. 1982, 16, 383–419. [Google Scholar] [CrossRef] [PubMed]
- Özyurt, G.; Ozogul, Y.; Kuley Boga, E.; Özkütük, A.S.; Durmuş, M.; Uçar, Y.; Ozogul, F. The effects of fermentation process with acid and lactic acid bacteria strains on the biogenic amine formation of wet and spray-dried fish silages of discards. J. Aquat. Food Prod. Technol. 2019, 28, 314–328. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Ramírez-Suárez, J.C.; Moreno-Hernández, J.M.; Pacheco-Aguilar, R. Seafood proteins. In Proteins in Food Processing, 2nd ed.; Rickey, Y.Y., Ed.; Woodhead Publishing: Sawston, UK, 2018; pp. 445–466. [Google Scholar]
- Javeed, A.; Mahendrakar, N.S. Effect of different levels of molasses and salt on acid production and volume of fermenting mass during ensiling of tropical freshwater fish viscera. J. Food Sci. Technol. 1995, 32, 115–118. [Google Scholar]
- Javeed, A.; Mahendrakar, N.S. Acceleration of fish viscera silage by prefermented starter culture. Ir. J. Agric. Food Res. 1996, 35, 171–177. [Google Scholar]
- Kameník, J. Hurdle technologies in fermented meat production. In Fermented Meat Products: Health Aspects; CRC Press: Boca Raton, FL, USA, 2017; Chapter 7. [Google Scholar]
- Bernardo, M.P.; Coelho, L.F.; Sass, D.C.; Contiero, J. L-(+)-Lactic acid production by Lactobacillus rhamnosus B103 from dairy industry waste. Braz. J. Microbiol. 2016, 47, 640–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mpofu, A.; Linnemann, A.R.; Nout, M.J.; Zwietering, M.H.; Smid, E.J.; den Besten, H.M. Inactivation of bacterial pathogens in yoba mutandabota, a dairy product fermented with the probiotic Lactobacillus rhamnosus yoba. Int. J. Food Microbiol. 2016, 217, 42–48. [Google Scholar] [CrossRef]
- Szajewska, H.; Kołodziej, M.; Gieruszczak-Białek, D.; Skórka, A.; Ruszczyński, M.; Shamir, R. Systematic review with meta-analysis: Lactobacillus rhamnosus GG for treating acute gastroenteritis in children—A 2019 update. Aliment. Pharmacol. Ther. 2019, 49, 1376–1384. [Google Scholar] [CrossRef]
- Lobb, B.; Hodgson, R.; Lynch, M.D.; Mansfield, M.J.; Cheng, J.; Charles, T.C.; Neufeld, J.D.; Craig, P.M.; Doxey, A.C. Time Series Resolution of the Fish Necrobiome Reveals a Decomposer Succession Involving Toxigenic Bacterial Pathogens. MSystems 2020, 5. [Google Scholar] [CrossRef]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in fish gastrointestinal microbiota research. Rev. Aquac. 2018, 10, 626–640. [Google Scholar] [CrossRef] [Green Version]
- Evariste, L.; Barret, M.; Mottier, A.; Mouchet, F.; Gauthier, L.; Pinelli, E. Gut microbiota of aquatic organisms: A key endpoint for ecotoxicological studies. Environ. Pollut. 2019, 248, 989–999. [Google Scholar] [CrossRef]
- Kashinskaya, E.; Simonov, E.; Kabilov, M.; Izvekova, G.; Andree, K.; Solovyev, M. Diet and other environmental factors shape the bacterial communities of fish gut in an eutrophic lake. J. Appl. Microbiol. 2018, 125, 1626–1641. [Google Scholar] [CrossRef] [PubMed]
- Schleifer, K.; Ludwig, W. Phylogenetic relationships of lactic acid bacteria. In The Genera of Lactic Acid Bacteria; Wood, B.J.B., Holzapfel, W.H., Eds.; Springer: Cambridge, MA, USA, 1995; Volume 2, pp. 7–18. [Google Scholar]
- Zabat, M.A.; Sano, W.H.; Wurster, J.I.; Cabral, D.J.; Belenky, P. Microbial community analysis of sauerkraut fermentation reveals a stable and rapidly established community. Foods 2018, 7, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.; Xia, W.; Jiang, Q.; Yang, F. Chemical and microbial properties of Chinese traditional low-salt fermented whole fish product Suan yu. Food Control 2013, 30, 590–595. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Yasokawa, D.; Nagashima, K.; Yamazaki, K.; Kurihara, H.; Ohta, T.; Kawai, Y. Microbiota during fermentation of chum salmon (Oncorhynchus keta) sauce mash inoculated with halotolerant microbial starters: Analyses using the plate count method and PCR-denaturing gradient gel electrophoresis (DGGE). Food Microbiol. 2010, 27, 509–514. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Ratio | ||||||
|---|---|---|---|---|---|---|
| Treatment | Content | Abbreviation | Acid Whey % | Fish % | Molasses % | LAB CFU/mL |
| 1 | Acid whey + Fish waste | AwF | 50 | 50 | - | - |
| 2 | Acid whey + Fish waste + Molasses | AwFM | 47.5 | 50 | 2.5 | - |
| 3 | Acid whey + Fish waste + L. rhamnosus | AwFLr | 50 | 50 | - | 7.2 × 1010 |
| 4 | Acid whey + Fish waste + Molasses + L. rhamnosus | AwFMLr | 47.5 | 50 | 2.5 | 7.2 × 1010 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayta-Apaza, A.C.; García-Cano, I.; Dabrowski, K.; Jiménez-Flores, R. Bacterial Diversity Analysis and Evaluation Proteins Hydrolysis during the Acid Whey and Fish Waste Fermentation. Microorganisms 2021, 9, 100. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010100
Mayta-Apaza AC, García-Cano I, Dabrowski K, Jiménez-Flores R. Bacterial Diversity Analysis and Evaluation Proteins Hydrolysis during the Acid Whey and Fish Waste Fermentation. Microorganisms. 2021; 9(1):100. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010100
Chicago/Turabian StyleMayta-Apaza, Alba C., Israel García-Cano, Konrad Dabrowski, and Rafael Jiménez-Flores. 2021. "Bacterial Diversity Analysis and Evaluation Proteins Hydrolysis during the Acid Whey and Fish Waste Fermentation" Microorganisms 9, no. 1: 100. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010100