
Genomic Characterization of a New Biofilm-Forming and Adhesive ST398 Human-Adapted MSSA Lineage Causing Septic Knee Arthritis Following Surgical Reconstruction

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
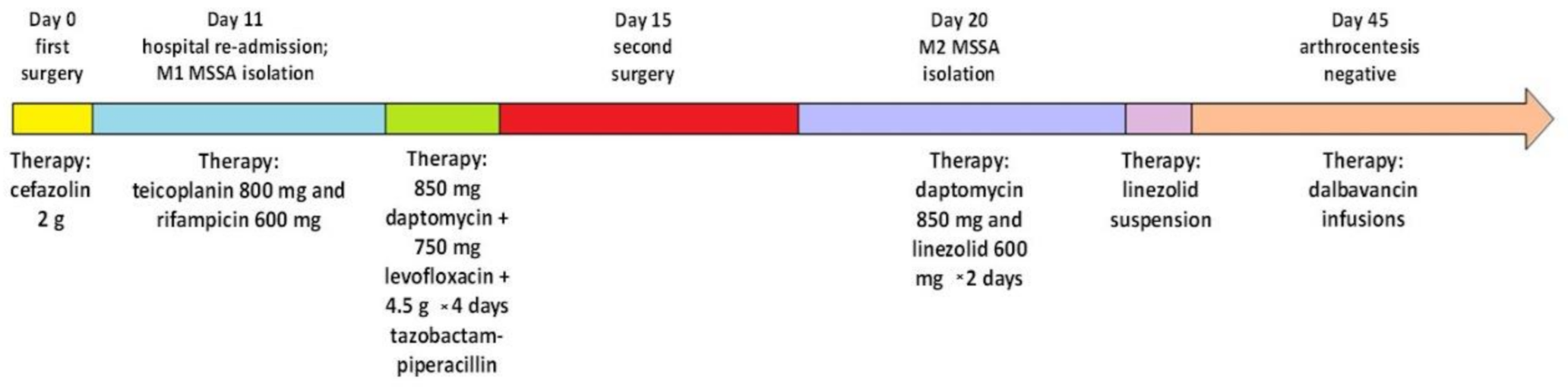
2.1. Clinical Case
2.2. Antibiotype
2.3. Biofilm and Slime Production
2.4. Haemolytic Ability
2.5. Whole Genome Sequencing (WGS)
2.6. De Novo Genome Assembly
2.7. Gene Annotation
2.8. Genomic Data Accession Number
2.9. Whole Genome Single Nucleotide Polymorphisms (wgSNPs)
2.10. Whole Genome Single Nucleotide Polymorphisms Effect Prediction
2.11. Genomic Epidemiology
2.12. Core Genome Single Nucleotide Polymorphisms
3. Results
3.1. Phenotypic Description of the Strains (Antibiotype Biofilm Haemolytic Properties)
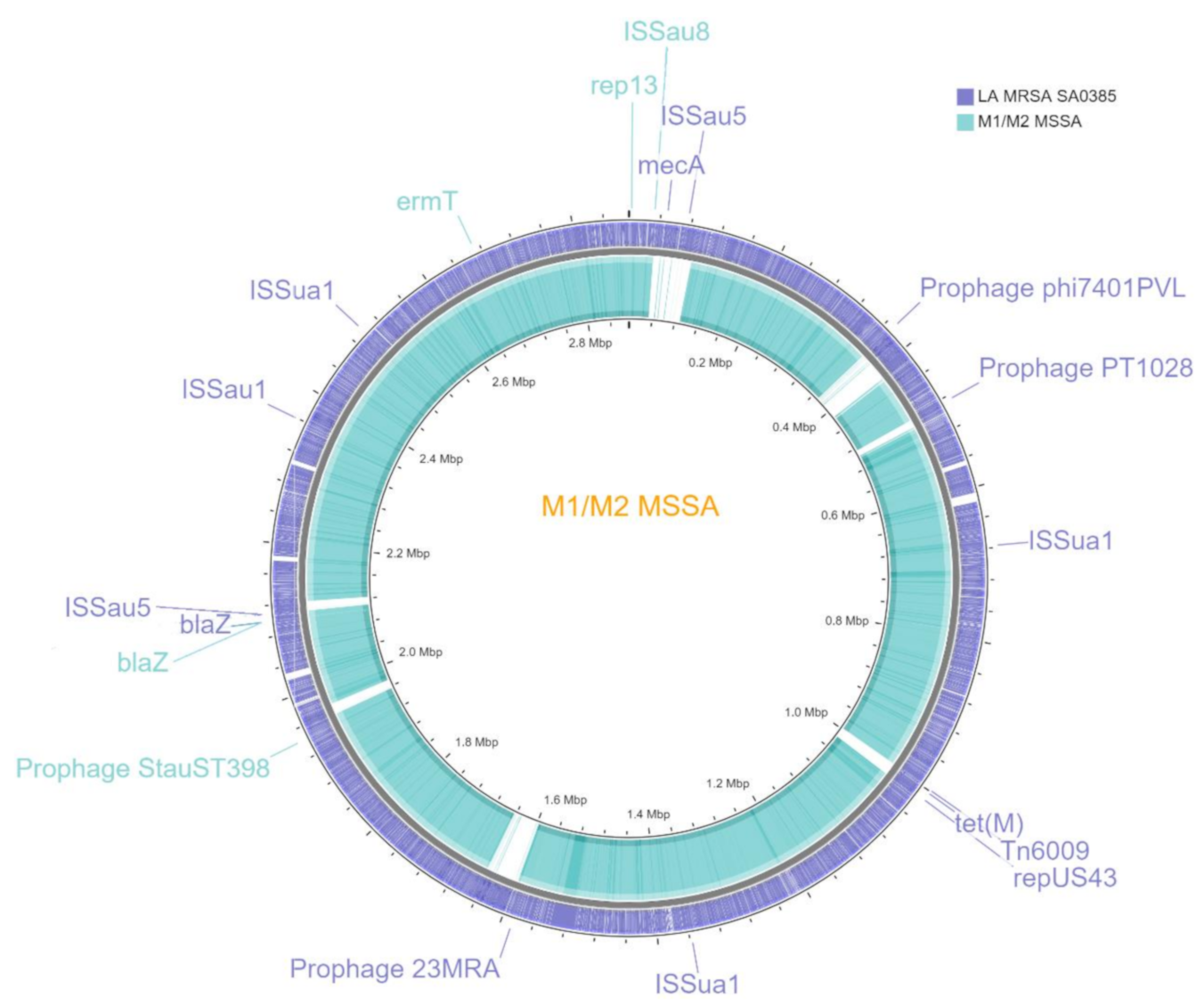
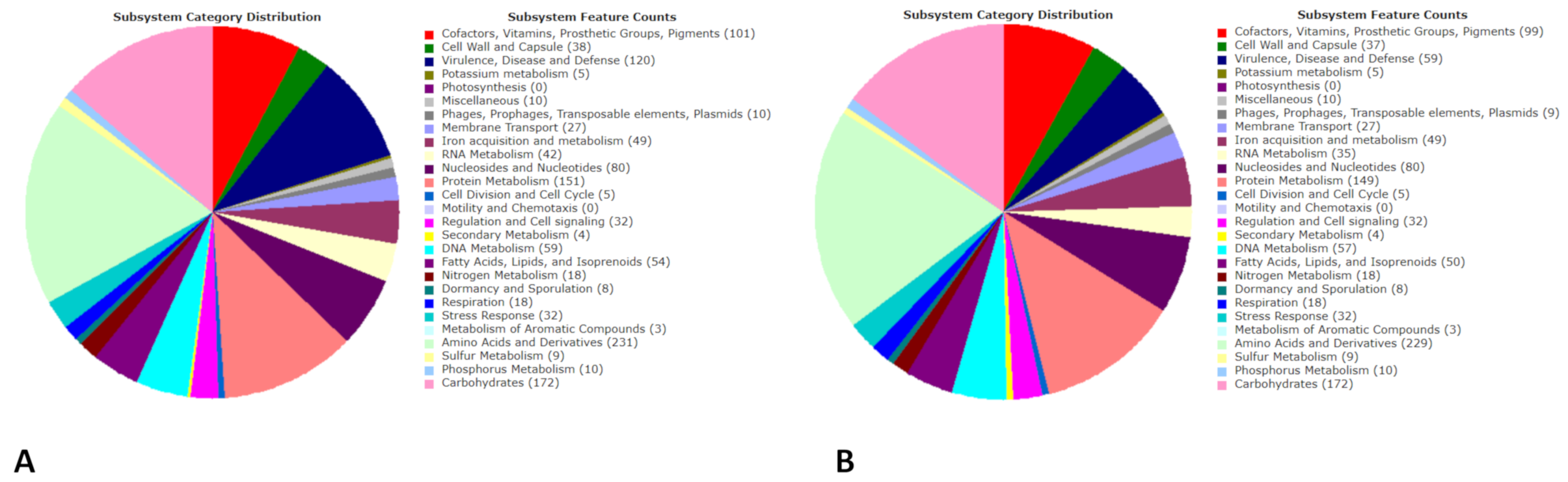
3.2. Genomics
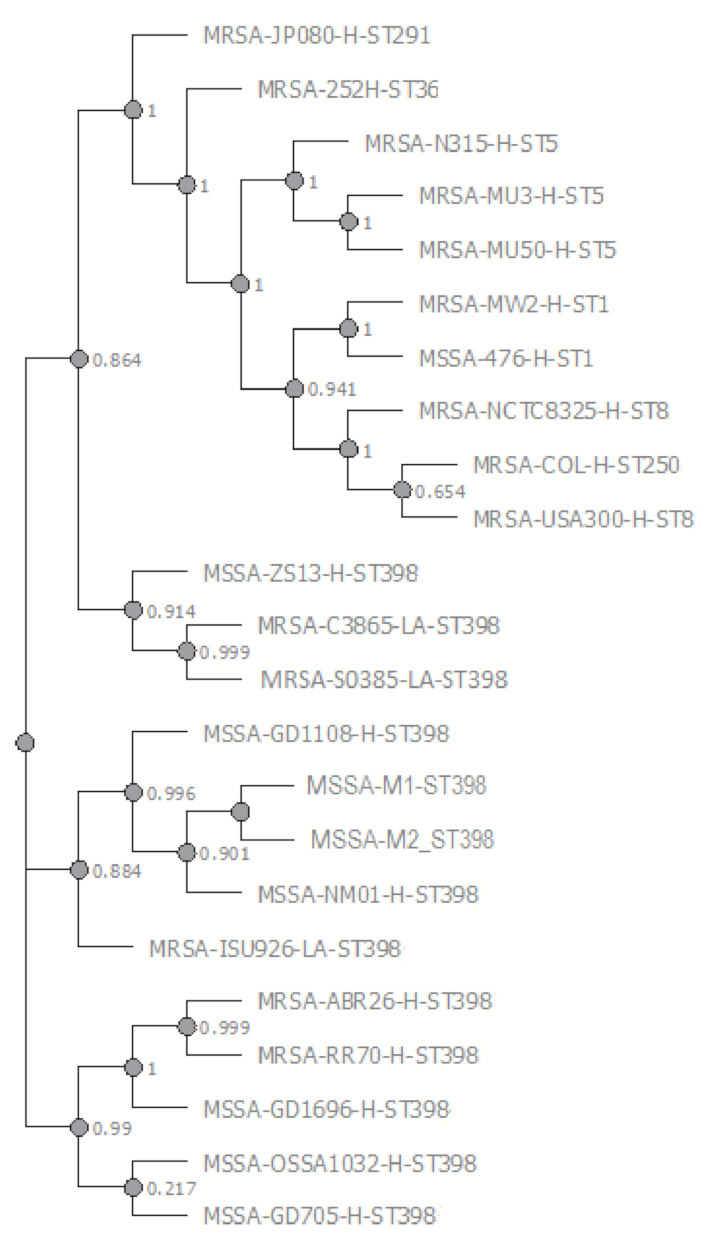
3.3. Phylogenomics—M1/M2 ST398 MSSA Comparative Genomics—wgSNPomes
3.4. Resistomics
3.5. Pathogenomics
3.6. M1 and M2 Shared HI and MI Effect wgSNPs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Combs, K.; Cox, K. Clinical outcomes involving patients that develop septic arthritis with methicillin sensitive staphylococcus aureus versus methicillin resistant staphylococcus aureus. J. Orthop. 2018, 15, 9–12. [Google Scholar] [CrossRef]
- Lindsay, J.A.; Holden, M.T.G. Understanding the rise of the superbug: Investigation of the evolution and genomic variation of Staphylococcus aureus. Funct. Integr. Genom. 2006, 6, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, M.; Wozniak, D.J.; Stoodley, P.; Hall-Stoodley, L. Prevention and treatment of Staphylococcus aureus biofilms. Expert Rev. Anti-Infect. Ther. 2015, 13, 1499–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harro, J.M.; Peters, B.M.; O’May, G.A.; Archer, N.; Kerns, P.; Prabhakara, R.; Shirtliff, M.E. Vaccine development in Staphylococcus aureus: Taking the biofilm phenotype into consideration. FEMS Immunol. Med. Microbiol. 2010, 59, 306–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, N.K.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Genet. 2014, 12, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Armand-Lefevre, L.; Ruimy, R.; Andremont, A. Clonal Comparison of Staphylococcus aureus Isolates from Healthy Pig Farmers, Human Controls, and Pigs. Emerg. Infect. Dis. 2005, 11, 711–714. [Google Scholar] [CrossRef]
- Price, L.B.; Stegger, M.; Hasman, H.; Aziz, M.; Larsen, J.; Andersen, P.S.; Pearson, T.; Waters, A.E.; Foster, J.T.; Schupp, J.; et al. Staphylococcus aureus CC398: Host Adaptation and Emergence of Methicillin Resistance in Livestock. mBio 2012, 3, e00305-11. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, A.J.; Van Wamel, W.; VandenDriessche, S.; Larsen, J.; Denis, O.; Garcia-Graells, C.; Uhlemann, A.-C.; Lowy, F.D.; Skov, R.; Lindsay, J.A. Staphylococcus aureus CC398 Clade Associated with Human-to-Human Transmission. Appl. Environ. Microbiol. 2012, 78, 8845–8848. [Google Scholar] [CrossRef] [Green Version]
- Cuny, C.; Abdelbary, M.; Layer, F.; Werner, G.; Witte, W. Prevalence of the immune evasion gene cluster in Staphylococcus aureus CC398. Vet. Microbiol. 2015, 177, 219–223. [Google Scholar] [CrossRef]
- Schijffelen, M.J.; Boel, C.H.E.; Van Strijp, J.A.G.; Fluit, A.C. Whole genome analysis of a livestock-associated methicillin-resistant Staphylococcus aureus ST398 isolate from a case of human endocarditis. BMC Genom. 2010, 11, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallin, M.; De Mendonça, R.; Denis, O.; Lefort, A.; El Garch, F.; Butaye, P.; Hermans, K.; Struelens, M.J. Diversity of accessory genome of human and livestock-associated ST398 methicillin resistant Staphylococcus aureus strains. Infect. Genet. Evol. 2011, 11, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Huijsdens, X.W.; Van Dijke, B.J.; Spalburg, E.; Van Santen-Verheuvel, M.G.; Heck, M.; Pluister, G.N.; Voss, A.; Wannet, W.J.B.; De Neeling, A.J. Community-acquired MRSA and pig-farming. Ann. Clin. Microbiol. Antimicrob. 2006, 5, 26. [Google Scholar] [CrossRef] [Green Version]
- Witte, W.; Strommenger, B.; Stanek, C.; Cuny, C. Methicillin-resistant Staphylococcus aureusST398 in Humans and Animals, Central Europe. Emerg. Infect. Dis. 2007, 13, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.; Dumortier, C.; Taylor, B.S.; Miller, M.; Vasquez, G.; Yunen, J.; Brudney, K.; Rodriguez-Taveras, C.; Rojas, R.; Leon, P.; et al. Staphylococcus aureusST398, New York City and Dominican Republic. Emerg. Infect. Dis. 2009, 15, 285–287. [Google Scholar] [CrossRef]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method for detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Cafiso, V.; Bertuccio, T.; Santagati, M.; Demelio, V.; Spina, D.; Nicoletti, G.; Stefani, S. agr-Genotyping and transcriptional analysis of biofilm-producing Staphylococcus aureus. FEMS Immunol. Med. Microbiol. 2007, 51, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zheng, Y.; Gao, H.; Xu, P.; Wang, M.; Li, A.; Miao, M.; Xie, X.; Deng, Y.; Zhou, H.; et al. Identification and Characterization of Staphylococcus aureus Strains with an Incomplete Hemolytic Phenotype. Front. Cell. Infect. Microbiol. 2016, 6, 146. [Google Scholar] [CrossRef] [Green Version]
- Cafiso, V.; Stracquadanio, S.; Verde, F.L.; Gabriele, G.; Mezzatesta, M.L.; Caio, C.; Pigola, G.; Ferro, A.; Stefani, S. Colistin Resistant A. baumannii: Genomic and Transcriptomic Traits Acquired Under Colistin Therapy. Front. Microbiol. 2019, 9, 3195. [Google Scholar] [CrossRef]
- Cafiso, V.; Stracquadanio, S.; Verde, F.L.; De Guidi, I.; Zega, A.; Pigola, G.; Stefani, S. Genomic and Long-Term Transcriptomic Imprints Related to the Daptomycin Mechanism of Action Occurring in Daptomycin- and Methicillin-Resistant Staphylococcus aureus Under Daptomycin Exposure. Front. Microbiol. 2020, 11, 1893. [Google Scholar] [CrossRef]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartels, M.; Petersen, A.; Worning, P.; Nielsen, J.B.; Larner-Svensson, H.; Johansen, H.K.; Andersen, L.P.; Jarløv, J.O.; Boye, K.; Larsen, A.R.; et al. Comparing Whole-Genome Sequencing with Sanger Sequencing for spa Typing of Methicillin-Resistant Staphylococcus aureus. J. Clin. Microbiol. 2014, 52, 4305–4308. [Google Scholar] [CrossRef] [Green Version]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-Time Whole-Genome Sequencing for Routine Typing, Surveillance, and Outbreak Detection of Verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Hasman, H. PlasmidFinder and In Silico pMLST: Identification and Typing of Plasmid Replicons in Whole-Genome Sequencing (WGS). Toxic. Assess. 2020, 2075, 285–294. [Google Scholar] [CrossRef]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: Mobile Element Finder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A Fast Phage Search Tool. Nucleic Acids Res. 2011, 39, W347–W352. [Google Scholar] [CrossRef]
- Van Der Mee-Marquet, N.; Corvaglia, A.; Valentin, A.-S.; Hernandez, D.; Bertrand, X.; Girard, M.; Kluytmans, J.; Donnio, P.-Y.; Quentin, R.; François, P. Analysis of prophages harbored by the human-adapted subpopulation of Staphylococcus aureus CC398. Infect. Genet. Evol. 2013, 18, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Sauget, M.; Bouiller, K.; Richard, M.; Chagrot, J.; Cholley, P.; Hocquet, D.; Bertrand, X. Increasing incidence of bloodstream infections due to Staphylococcus aureus clonal complex 398 in a French hospital between 2010 and 2017. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 2127–2132. [Google Scholar] [CrossRef]
- Li, X.; Huang, T.; Xu, K.; Li, C.; Li, Y. Molecular characteristics and virulence gene profiles of Staphylococcus aureus isolates in Hainan, China. BMC Infect. Dis. 2019, 19, 873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubin, G.G.; Lepelletier, D.; Reynaud, A.; Lavigne, J.-P.; Corvec, S. Methicillin-susceptible Staphylococcus aureus CC398: First description in prosthetic joint infection and genetic background comparison with nasal carriage isolates. J. Infect. Chemother. 2014, 20, 394–396. [Google Scholar] [CrossRef] [PubMed]
- Valour, F.; Tasse, J.; Trouillet-Assant, S.; Rasigade, J.-P.; Lamy, B.; Chanard, E.; Verhoeven, P.; Decousser, J.-W.; Marchandin, H.; Bés, M.; et al. Methicillin-susceptible Staphylococcus aureus clonal complex 398: High prevalence and geographical heterogeneity in bone and joint infection and nasal carriage. Clin. Microbiol. Infect. 2014, 20, O772–O775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senneville, E.; Brière, M.; Neut, C.; Messad, N.; Lina, G.; Richard, J.-L.; Sotto, A.; Lavigne, J.-P. First report of the predominance of clonal complex 398 Staphylococcus aureus strains in osteomyelitis complicating diabetic foot ulcers: A national French study. Clin. Microbiol. Infect. 2014, 20, O274–O277. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Ballhausen, B.; Kahl, B.C.; Köck, R. The clinical impact of livestock-associated methicillin-resistant Staphylococcus aureus of the clonal complex 398 for humans. Vet. Microbiol. 2017, 200, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.C.; Wardyn, S.E. Human Infections with Staphylococcus aureus CC398. Curr. Environ. Health Rep. 2015, 2, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, R.A.; Kriegeskorte, A.; Kahl, B.C.; Ebecker, K.; Löffler, B.; Epeters, G. Staphylococcus aureus Small Colony Variants (SCVs): A road map for the metabolic pathways involved in persistent infections. Front. Cell. Infect. Microbiol. 2014, 4, 99. [Google Scholar] [CrossRef]
- Herrmann, M.; Vaudaux, P.E.; Pittet, D.; Auckenthaler, R.; Lew, P.D.; Perdreau, F.S.; Peters, G.; Waldvogel, F.A. Fibronectin, Fibrinogen, and Laminin Act as Mediators of Adherence of Clinical Staphylococcal Isolates to Foreign Material. J. Infect. Dis. 1988, 158, 693–701. [Google Scholar] [CrossRef]
- Rydén, C.; Tung, H.-S.; Nikolaev, V.; Engström, Å.; Oldberg, Å. Staphylococcus aureus causing osteomyelitis binds to a nonapeptide sequence in bone sialoprotein. Biochem. J. 1997, 327, 825–829. [Google Scholar] [CrossRef] [Green Version]
- Yacoub, A.; Lindahl, P.; Rubin, K.; Wendel, M.; Heinegard, D.; Ryden, C. Purification of a bone sialoprotein-binding protein from Staphylococcus aureus. JBIC J. Biol. Inorg. Chem. 1994, 222, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Köck, R.; Schaumburg, F.; Mellmann, A.; Köksal, M.; Jurke, A.; Becker, K.; Friedrich, A.W. Livestock-Associated Methicillin-Resistant Staphylococcus aureus (MRSA) as Causes of Human Infection and Colonization in Germany. PLoS ONE 2013, 8, e55040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlemann, A.-C.; Porcella, S.F.; Trivedi, S.; Sullivan, S.B.; Hafer, C.; Kennedy, A.D.; Barbian, K.D.; McCarthy, A.J.; Street, C.; Hirschberg, D.L.; et al. Identification of a Highly Transmissible Animal-Independent Staphylococcus aureus ST398 Clone with Distinct Genomic and Cell Adhesion Properties. mBio 2012, 3, 00027-12. [Google Scholar] [CrossRef] [Green Version]
- Bongiorno, D.; Musso, N.; Lazzaro, L.M.; Mongelli, G.; Stefani, S.; Campanile, F. Detection of methicillin-resistant Staphylococcus aureus persistence in osteoblasts using imaging flow cytometry. Microbiology 2020, 9, e1017. [Google Scholar] [CrossRef] [Green Version]
- Gemmell, C.G.; Goutcher, S.C.; Reid, R.; Sturrock, R.D. Role of certain virulence factors in a murine model of Staphylococcus aureus arthritis. J. Med. Microbiol. 1997, 46, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Albus, A.; Arbeit, R.D.; Lee, J.C. Virulence of Staphylococcus aureus mutants altered in type 5 capsule production. Infect. Immun. 1991, 59, 1008–1014. [Google Scholar] [CrossRef] [Green Version]
- Essawi, T.; Na’Was, T.; Hawwari, A.; Wadi, S.; Doudin, A.; Fattom, A.I. Molecular, antibiogram and serological typing of Staphylococcus aureus isolates recovered from Al-Makased Hospital in East Jerusalem. Trop. Med. Int. Heal. 1998, 3, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Na’Was, T.; Hawwari, A.; Hendrix, E.; Hebden, J.; Edelman, R.; Martin, M.; Campbell, W.; Naso, R.; Schwalbe, R.; Fattom, A.I. Phenotypic and Genotypic Characterization of Nosocomial Staphylococcus aureus Isolates from Trauma Patients. J. Clin. Microbiol. 1998, 36, 414–420. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, I.M.; Lee, J.C.; Bremell, T.; Rydén, C.; Tarkowski, A. The role of staphylococcal polysaccharide microcapsule expression in septicemia and septic arthritis. Infect. Immun. 1997, 65, 4216–4221. [Google Scholar] [CrossRef] [Green Version]
- Lowe, A.M.; Beattie, D.T.; Deresiewicz, R.L. Identification of novel staphylococcal virulence genes by in vivo expression technology. Mol. Microbiol. 1998, 27, 967–976. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MICs (mg/L) | Biofilm | Slime Production | Virulome | Resistome | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DAP | VAN | TEC | GEN | ERY | CLI | FOX | TET | MUP | CIP | SXT | RIF | LZD | DAL | |||||||
| M1 | 0.5 | 0.75 | 1 | 0.25 | >32 | ind | 4 | 0.5 | 0.25 | 0.75 | 0.016 | 4 | 4 | 0.125 | Producer | Positive | ebp | blaZ (β-lactam-R) ermT (Macrolide/Lincosamide and Streptogramin B-R) | ||
| isdA-G | ||||||||||||||||||||
| clfA/B | ||||||||||||||||||||
| fnbA/B | ||||||||||||||||||||
| cna | ||||||||||||||||||||
| map | ||||||||||||||||||||
| sdrCDE | ||||||||||||||||||||
| hlgABC | ||||||||||||||||||||
| hlb | ||||||||||||||||||||
| hld | ||||||||||||||||||||
| hly/hla | ||||||||||||||||||||
| aur | ||||||||||||||||||||
| sspABC | ||||||||||||||||||||
| lip | ||||||||||||||||||||
| strB | ||||||||||||||||||||
| spa | ||||||||||||||||||||
| sbi | ||||||||||||||||||||
| scn | ||||||||||||||||||||
| chp | ||||||||||||||||||||
| adsA | ||||||||||||||||||||
| geh | ||||||||||||||||||||
| essABC | ||||||||||||||||||||
| esaAB | ||||||||||||||||||||
| esxA | ||||||||||||||||||||
| M2 | 0.5 | 0.75 | 1 | 0.25 | >32 | ind | 4 | 0.5 | 0.25 | 0.75 | 0.016 | 4 | 4 | 0.125 | Producer | Positive | ebp | blaZ (β-lactam-R) ermT (Macrolide/Lincosamide and Streptogramin B-R) | ||
| isdA-G | ||||||||||||||||||||
| clfA/B | ||||||||||||||||||||
| fnbA/B | ||||||||||||||||||||
| cna | ||||||||||||||||||||
| map | ||||||||||||||||||||
| sdrCDE | ||||||||||||||||||||
| hlgABC | ||||||||||||||||||||
| hlb | ||||||||||||||||||||
| hld | ||||||||||||||||||||
| hly/hla | ||||||||||||||||||||
| aur | ||||||||||||||||||||
| sspABC | ||||||||||||||||||||
| lip | ||||||||||||||||||||
| strB | ||||||||||||||||||||
| spa | ||||||||||||||||||||
| sbi | ||||||||||||||||||||
| scn | ||||||||||||||||||||
| chp | ||||||||||||||||||||
| adsA | ||||||||||||||||||||
| geh | ||||||||||||||||||||
| essABC | ||||||||||||||||||||
| esaAB | ||||||||||||||||||||
| esxA | ||||||||||||||||||||
| Strain | mec Gene | MLST | spa-Type | Capsular Type | agr-Group | Plasmid rep Gene | MGEs | Prophage | Resistome | Adhesin Genes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| clfA | clfB | fnbA | fnbB | CoA | sdrE | sdrC | sdrD | cna | ||||||||||
| LA-MRSA SA0385 | mecA | ST398 | t011 | Cap5 | I | repUS43 CDS12738 (DOp1) | ISSau5 (2 copies) ISSau1 (4 copies) Tn6009 | φ7401PVL PT1028 23MRA | mecA blaZ tetM | truncated | truncated | WT | truncated | Δ81bp | absent | Δ174bp | 3SNPs | WT |
| Human-animal independent MSSA NM01 | - | ST398 | t571 | Cap5 | I | rep13 (pSSP1) | CT Cn_30859-ISSau1 (4 copies) | φNM3 R8A2B | blaZ ermT | WT | WT | WT | WT | WT | WT | WT | Δ54bp | ΔBdomain |
| M1/M2 MSSA | - | ST398 | t1451 | Cap5 | I | rep13 (pC194) | ISSau8 (1 copy) | StauST398-4 | blaZ ermT | WT | WT | WT | WT | WT | WT | WT | 3SNPs | WT |
| SNPs Content on LA ST398 MRSA RefGen Mapping (CP003045.1) | SNPs Content on Human-Animal Independent ST398 MSSA RefGen Mapping (AM990992.1) | ||||
|---|---|---|---|---|---|
| M1 | M2 | M1 | M2 | ||
| cgSNPomes | cgSNPs vs RefGen | 852 | 851 | 265 | 264 |
| cgSNPs M1 vs M2 | 1 | 1 | 1 | 1 | |
| wgSNPomes | Variant Rate | 1318 | 1401 | 531 | 1890 |
| Variant Number | 2178 | 2091 | 5106 | 1436 | |
| MISSENSE SNPs (%) | 45.846 | 44.88 | 58.3106 | 57.164 | |
| NONSENSE SNPs (%) | 2.116 | 1.365 | 2.914 | 4.874 | |
| SILENT SNPs (%) | 52.08 | 53.74 | 38.98 | 37.962 | |
| HI Eff SNPs (%) | 4.02 | 3.44 | 1.35 | 6.163 | |
| MI Eff SNPs (%) | 33.501 | 33.01 | 6.53 | 27.978 | |
| LI Eff SNPs (%) | 34.87 | 35.323 | 4.32 | 18.09 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cafiso, V.; Lo Verde, F.; Zega, A.; Pigola, G.; Rostagno, R.; Borrè, S.; Stefani, S. Genomic Characterization of a New Biofilm-Forming and Adhesive ST398 Human-Adapted MSSA Lineage Causing Septic Knee Arthritis Following Surgical Reconstruction. Microorganisms 2021, 9, 305. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020305
Cafiso V, Lo Verde F, Zega A, Pigola G, Rostagno R, Borrè S, Stefani S. Genomic Characterization of a New Biofilm-Forming and Adhesive ST398 Human-Adapted MSSA Lineage Causing Septic Knee Arthritis Following Surgical Reconstruction. Microorganisms. 2021; 9(2):305. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020305
Chicago/Turabian StyleCafiso, Viviana, Flavia Lo Verde, Alessandra Zega, Giuseppe Pigola, Roberto Rostagno, Silvio Borrè, and Stefania Stefani. 2021. "Genomic Characterization of a New Biofilm-Forming and Adhesive ST398 Human-Adapted MSSA Lineage Causing Septic Knee Arthritis Following Surgical Reconstruction" Microorganisms 9, no. 2: 305. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9020305