
Microbial Diversity of Psychrotolerant Bacteria Isolated from Wild Flora of Andes Mountains and Patagonia of Chile towards the Selection of Plant Growth-Promoting Bacterial Consortia to Alleviate Cold Stress in Plants



 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Reagents, and Culture Media
2.2. Model and Phytopathogenic Bacterial Strains
2.3. Collection of Wild Flora Samples
2.4. Psychrotolerant Bacteria Isolation
2.5. Psychrotolerant Potential Test
2.6. Genomic DNA Extraction, Identification of Selected Bacteria, and Phylogenetic Analysis
2.7. Bacterial Consortia Selection
2.7.1. Auxin Production
2.7.2. Phosphate Solubilization and Detection of the nifH and the acdS Genes
2.7.3. Antimicrobial Activities against Phytopathogenic Bacteria
2.7.4. Compatibility Tests and Formulation of Bacterial Consortia
2.8. Plant Growth-Promoting Assays with Bacterial Consortia
2.8.1. Effect of Bacterial Consortia on Tomato Plants Growth
2.8.2. Effect of Bacterial Consortia on Tomato Seeds Germination
2.9. Bacterial Consortium Selection
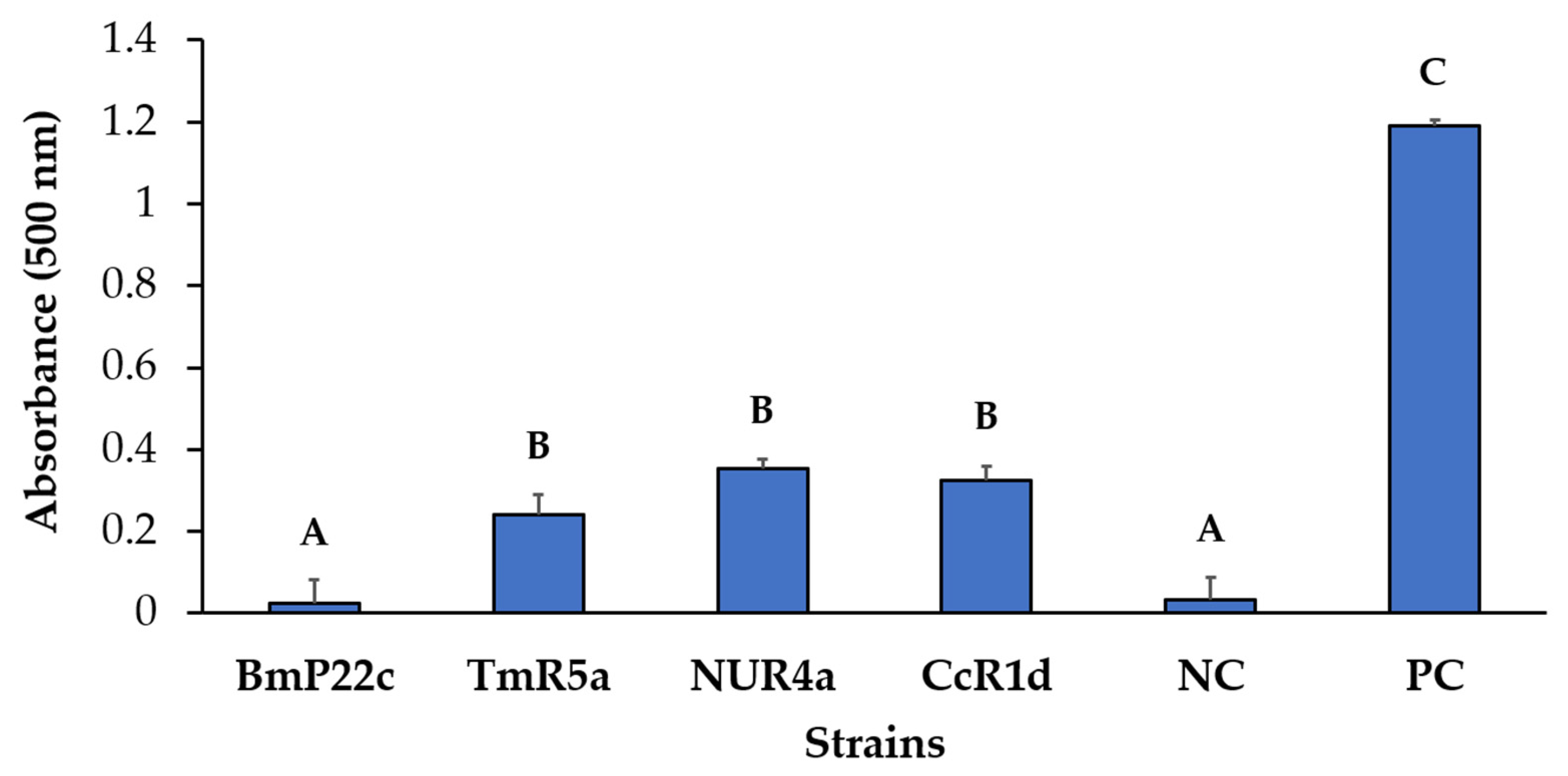
2.9.1. ACC Deaminase Activity
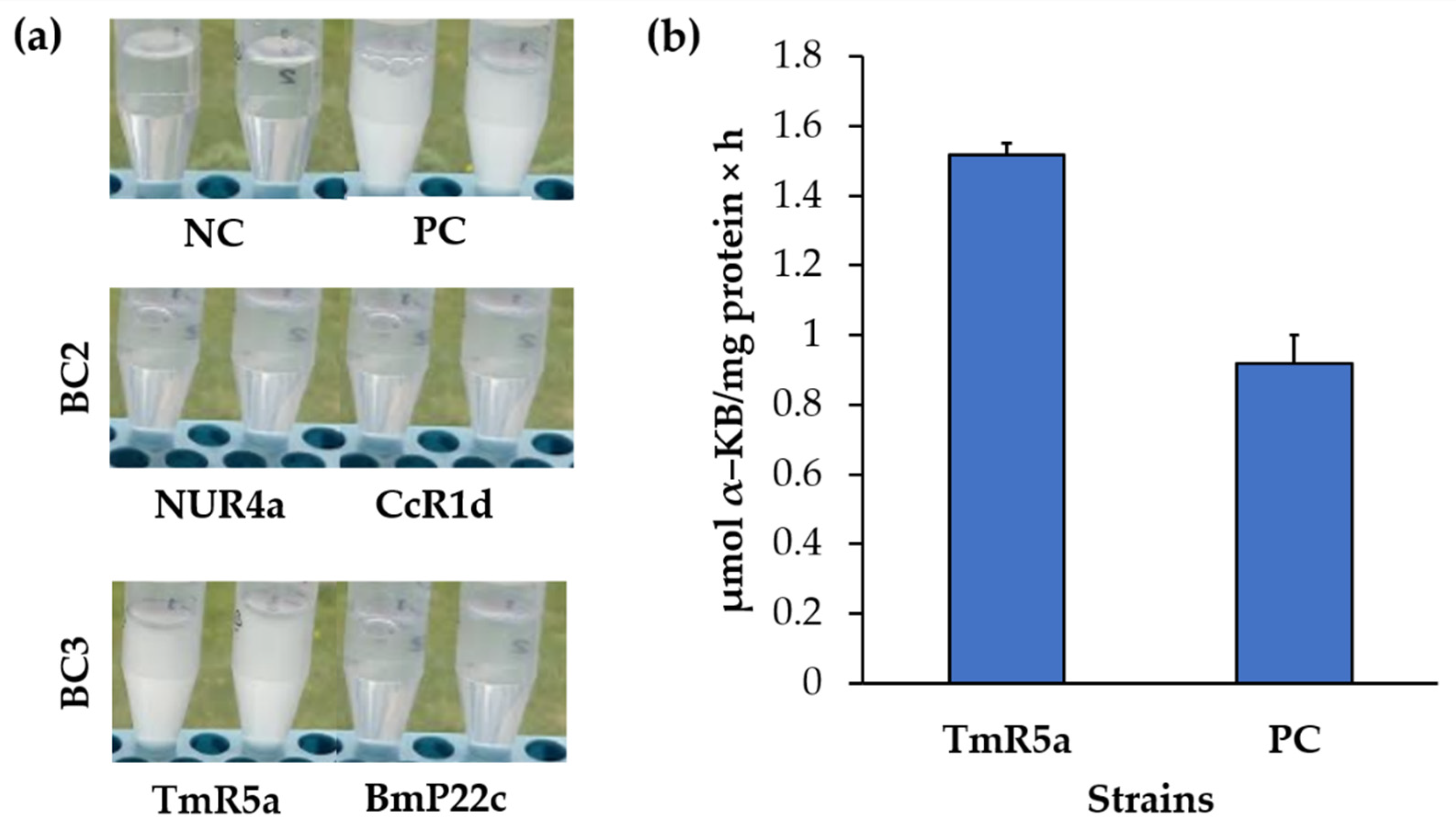
2.9.2. IRI Activity
2.10. Statistical Analysis
3. Results
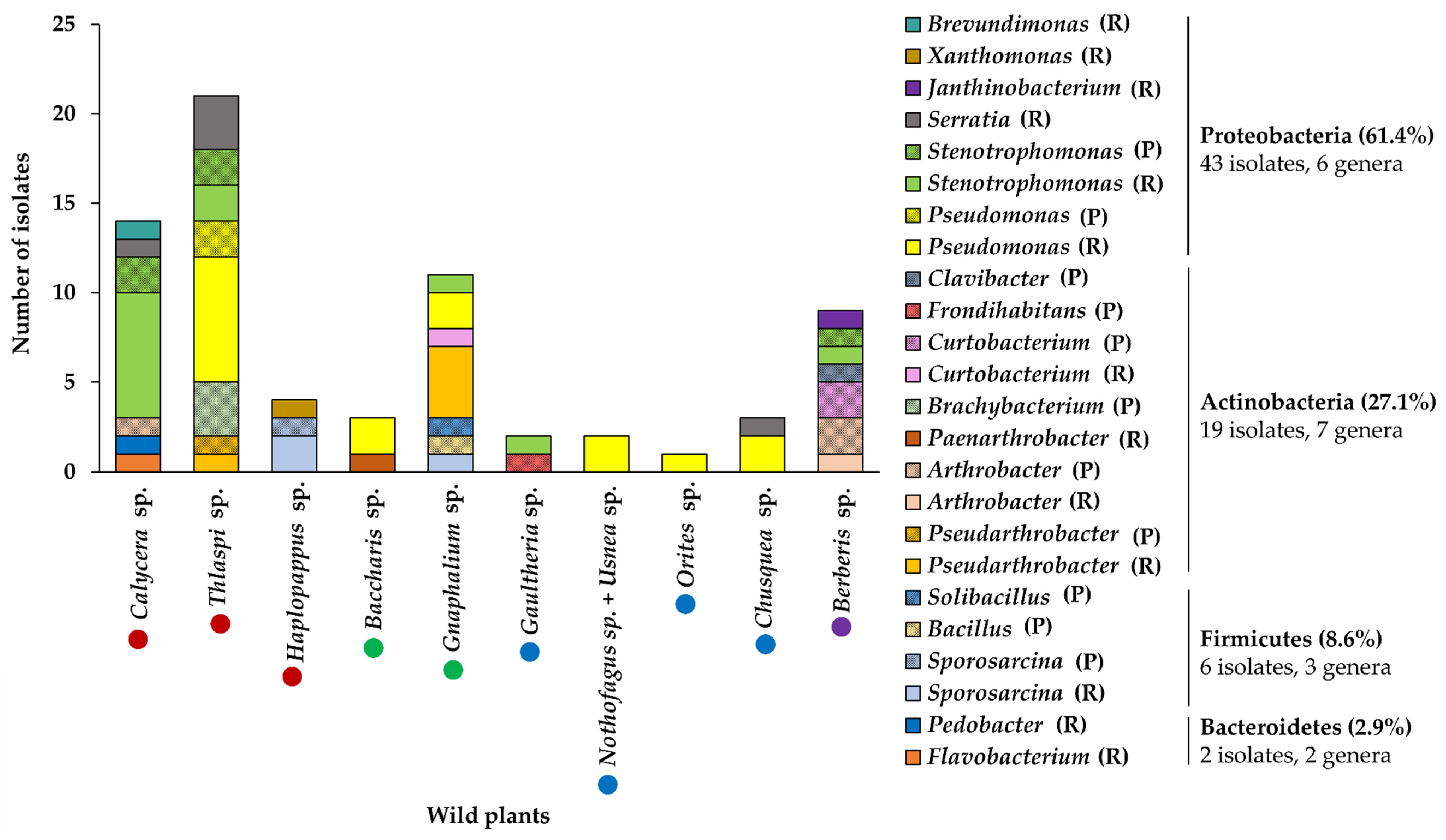
3.1. Collection of Wild Flora from Andean and Patagonia Areas of Chile and Isolation of Psychrotolerant Bacteria
3.2. Selection of Psychrotolerant Bacteria
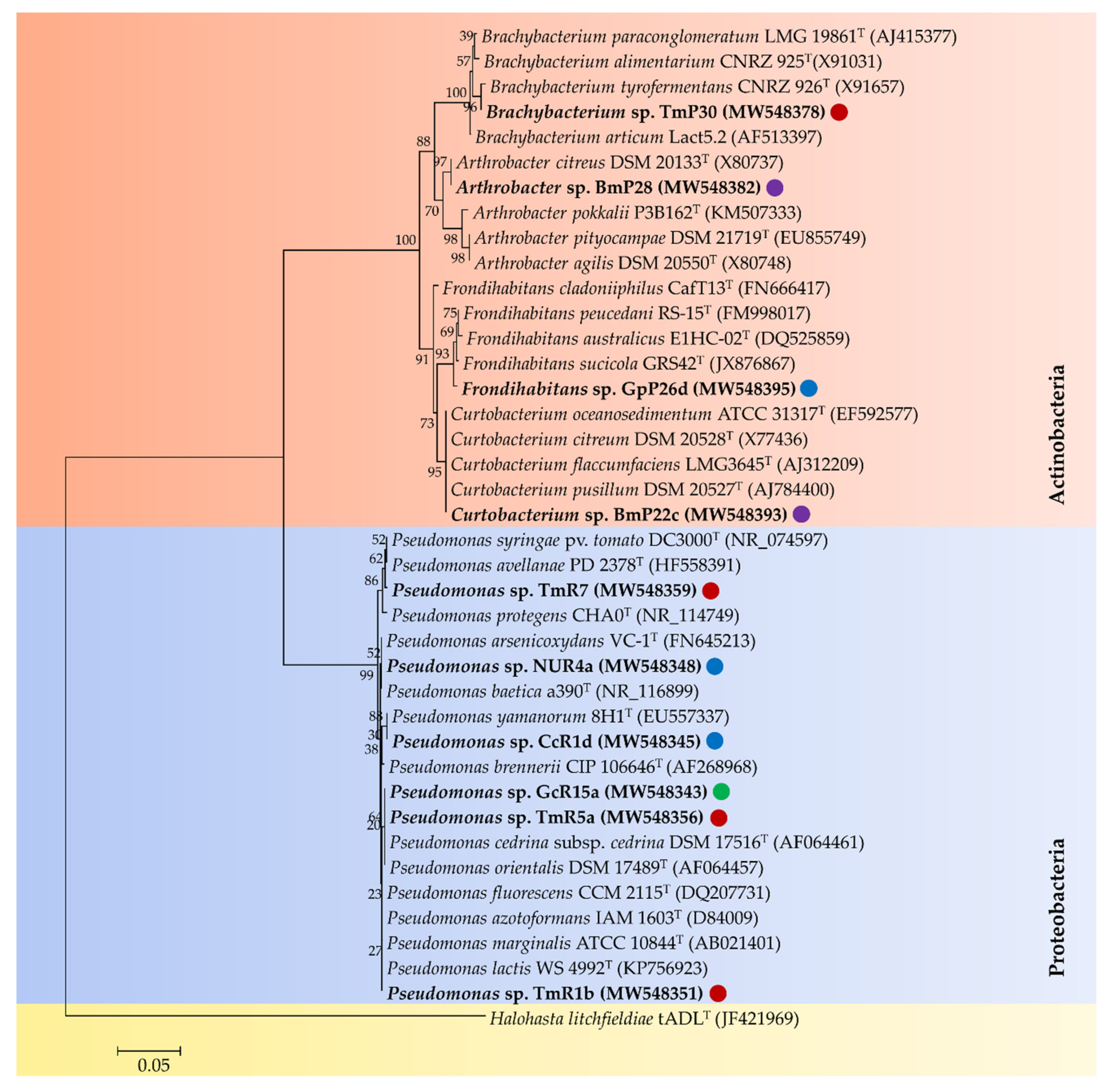
3.3. Identification of Selected Psychrotolerant Bacteria
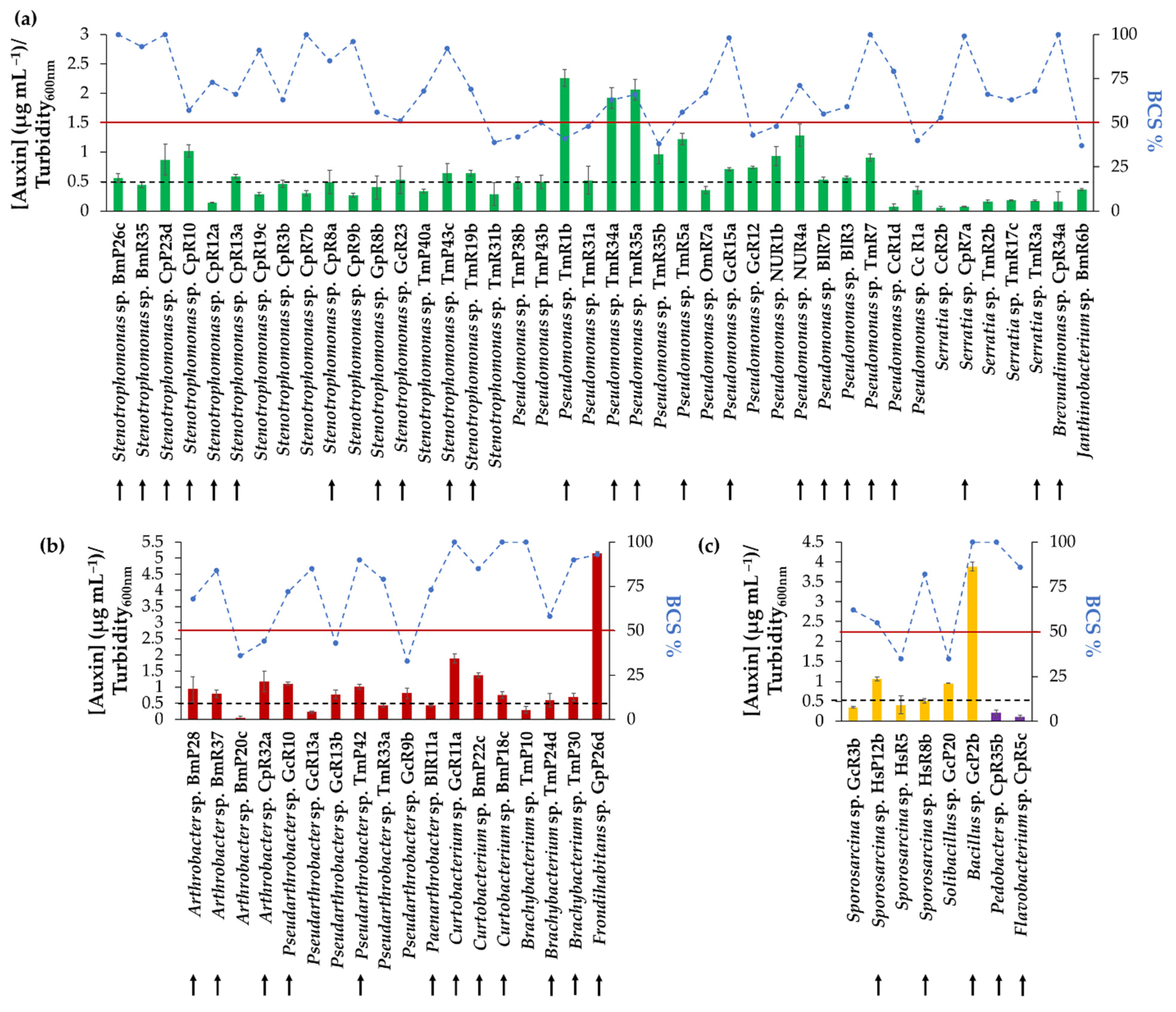
3.4. Auxin Biosynthesis of Selected Strains
3.5. Phosphate Solubilization and Presence of the nifH and the acdS Genes in Selected Strains
3.6. Antimicrobial Activities of Selected Strains
3.7. Compatibility Tests and Formulation of Bacterial Consortia
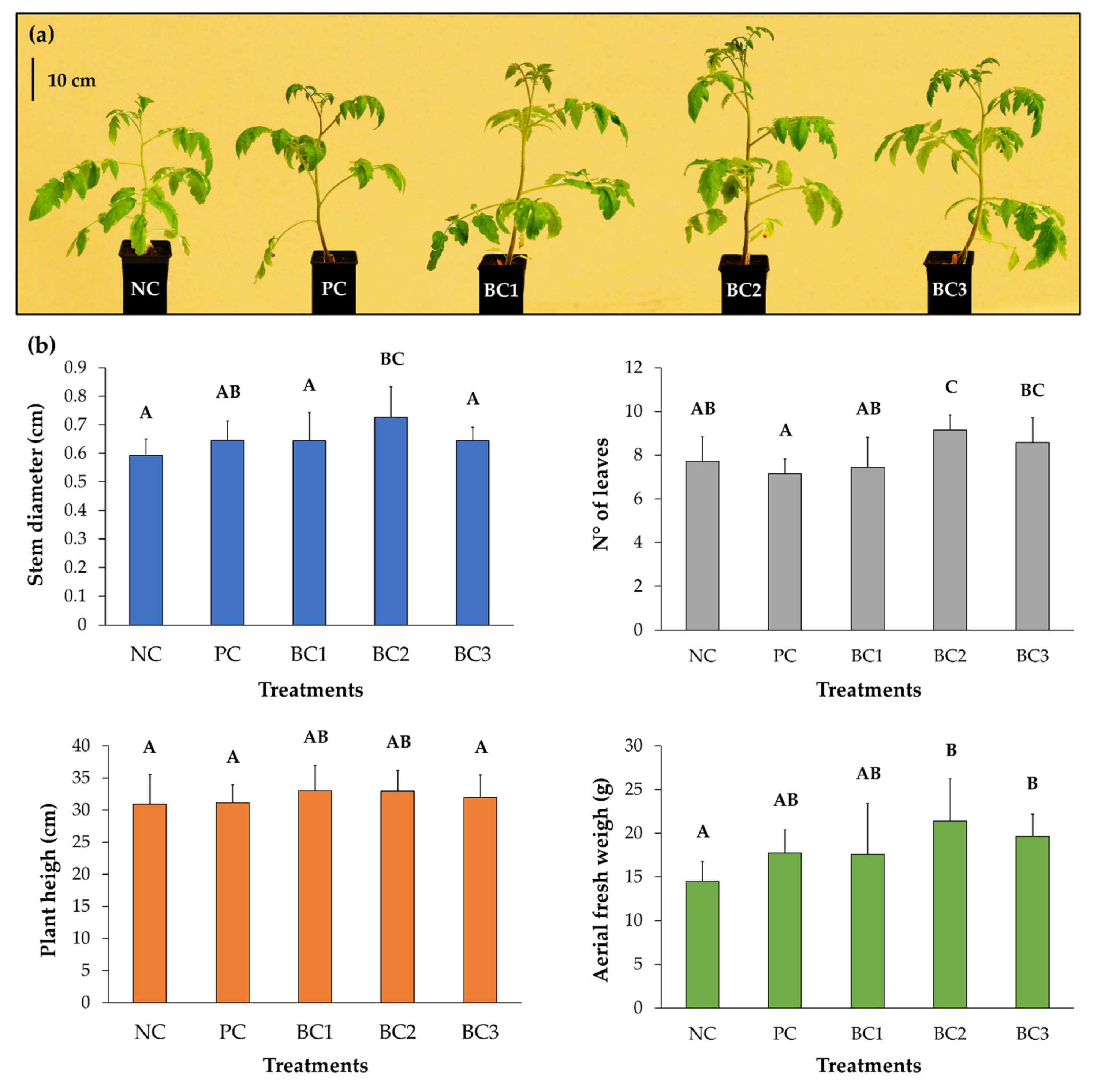
3.8. Plant Growth–Promotion on Tomato Plants
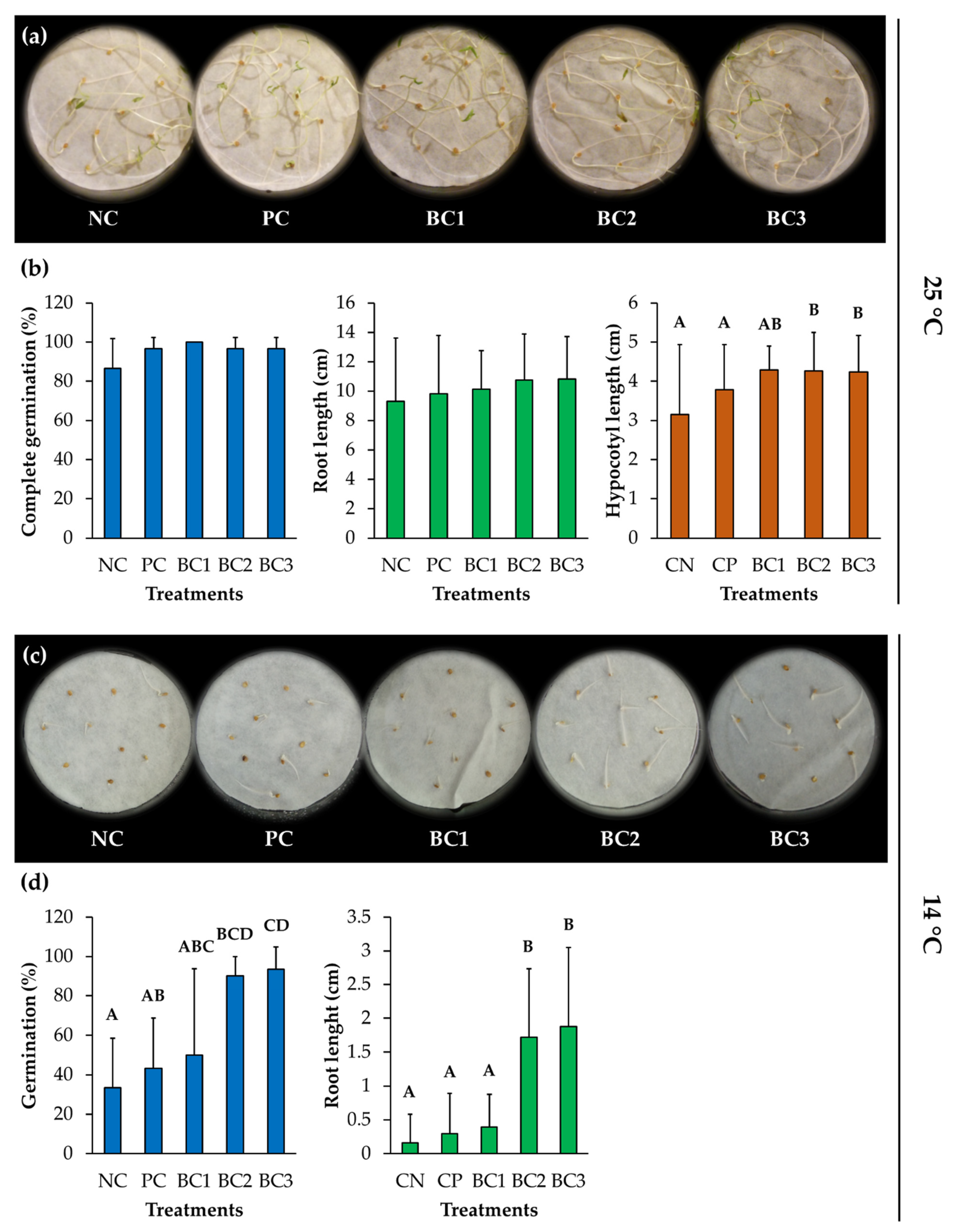
3.9. Plant Growth–Promotion on Tomato Seeds
3.10. Selection of One Bacterial Consortium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent advances in utilizing transcription factors to improve plant abiotic stress tolerance by transgenic technology. Front. Plant Sci. 2016, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- FAO. Frost Protection: Fundamentals, Practice and Economics; Food and Agriculture Organization of the United Nations: Rome, Italy, 2005. Available online: http://www.fao.org/3/y7223e/y7223e00.htm (accessed on 3 February 2021).
- Duman, J.G.; Wisniewski, M. The use of antifreeze proteins for frost protection in sensitive crop plants. Environ. Exp. Bot. 2014, 106, 60–69. [Google Scholar] [CrossRef]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Dang, P.; Liu, L.; He, C. Cold acclimation by the CBF–COR pathway in a changing climate: Lessons from Arabidopsis thaliana. Plant Cell Rep. 2019, 38, 511–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlow, K.M.; Christy, B.P.; O’Leary, G.L.; Riffkin, P.A.; Nuttall, J.G. Simulating the impact of extreme heat and frost events on wheat crop production: A review. Field Crops Res. 2015, 171, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Bagati, S.; Mahajan, R.; Nazir, M.; Dar, A.A.; Zargar, S.M. “Omics”: A gateway towards abiotic stress tolerance. In Abiotic Stress-mediated Sensing and Signaling in Plants: An Omics Perspective; Zargar, S., Zargar, M., Eds.; Springer: Singapore, 2018; pp. 1–45. [Google Scholar]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 1, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rilling, J.I.; Acuña, J.J.; Sadowsky, M.J.; Jorquera, M.A. Putative nitrogen-fixing bacteria associated with the rhizosphere and root endosphere of wheat plants grown in an andisol from Southern Chile. Front. Microbiol. 2018, 9, 2710. [Google Scholar] [CrossRef] [PubMed]
- Bakhshandeh, E.; Gholamhosseini, M.; Yaghoubian, Y.; Pirdashti, H. Plant growth promoting microorganisms can improve germination, seedling growth and potassium uptake of soybean under drought and salt stress. Plant Growth Regul. 2020, 90, 123–136. [Google Scholar] [CrossRef]
- Velásquez, A.; Vega-Celedón, P.; Fiaschi, G.; Agnolucci, M.; Avio, L.; Giovannetti, M.; D’Onofrio, C.; Seeger, M. Responses of Vitis vinifera cv. Cabernet Sauvignon roots to the arbuscular mycorrhizal fungus Funneliformis mosseae and the plant growth-promoting rhizobacterium Ensifer meliloti include changes in volatile organic compounds. Mycorrhiza 2020, 30, 161–170. [Google Scholar] [CrossRef]
- Mishra, P.K.; Bisht, S.C.; Ruwari, P.; Selvakumar, G.; Joshi, G.K.; Bisht, J.K.; Bhatt, J.C.; Gupta, H.S. Alleviation of cold stress in inoculated wheat (Triticum aestivum L.) seedlings with psychrotolerant Pseudomonas from NW Himalayas. Arch. Microbiol. 2011, 193, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Bisht, S.; Sood, A.; Aeron, A.; Sharma, G.D.; Maheshwari, D.K. Consortium of plant-growth-promoting bacteria: Future perspective in agriculture. In Bacteria in Agrobiology: Plant Probiotics; Maheshwari, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 185–200. [Google Scholar]
- Majeed, A.; Muhammad, Z.; Ahmad, H. Plant growth promoting bacteria: Role in soil improvement, abiotic and biotic stress management of crops. Plant Cell Rep. 2018, 37, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Bradáčová, K.; Florea, A.S.; Bar-Tal, A.; Minz, D.; Yermiyahu, U.; Shawahna, R.; Kraut-Cohen, J.; Zolti, A.; Erel, R.; Dietel, K.; et al. Microbial consortia versus single-strain inoculants: An advantage in PGPM-assisted tomato production? Agronomy 2019, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Menéndez, E.; Paço, A. Is the application of plant probiotic bacterial consortia always beneficial for plants? Exploring synergies between rhizobial and non-rhizobial bacteria and their effects on agro-economically valuable crops. Life 2020, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Griffith, M.; Pasternak, J.J.; Glick, B.R. Low temperature growth, freezing survival and production of antifreeze protein by the plant growth promoting rhizobacterium Pseudomonas putida GR12-2. Can. J. Microbiol. 1995, 41, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Park, E.; Glick, B.R. 1-Aminocyclopropane-1-carboxylate deaminase from Pseudomonas putida UW4 facilitates the growth of canola in the presence of salt. Can. J. Microbiol. 2007, 53, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Mishra, S.; Selvakumar, G.; Bisht, S.C.; Bisht, J.K.; Kundu, S.; Gupta, H.S. Characterization of a psychrotolerant plant growth promoting Pseudomonas sp. strain PGERs17 (MTCC 9000) isolated from North Western Indian Himalayas. Ann. Microbiol. 2008, 58, 561–568. [Google Scholar] [CrossRef]
- Selvakumar, G.; Kundu, S.; Joshi, P.; Gupta, A.D.; Nazim, S.; Mishra, P.K.; Gupta, H.S. Characterization of a cold-tolerant plant growth-promoting bacterium Pantoea dispersa 1A isolated from a sub-alpine soil in the North Indian Himalayas. World J. Microbiol. Biotechnol. 2008, 24, 955–960. [Google Scholar] [CrossRef]
- Selvakumar, G.; Mohan, M.; Kundu, S.; Gupta, A.D.; Joshi, P.; Nazim, S.; Gupta, H.S. Cold tolerance and plant growth promotion potential of Serratia marcescens strain SRM (MTCC 8708) isolated from flowers of summer squash (Cucurbita pepo). Lett. Appl. Microbiol. 2008, 46, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Mishra, S.; Bisht, S.C.; Selvakumar, G.; Kundu, S.; Bisht, J.K.; Gupta, H.S. Isolation, molecular characterization and growth-promotion activities of a cold tolerant bacterium Pseudomonas sp. NARs9 (MTCC9002) from the Indian Himalayas. Biol. Res. 2009, 42, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Theocharis, A.; Bordiec, S.; Fernandez, O.; Paquis, S.; Dhondt-Cordelier, S.; Baillieul, F.; Clément, C.; Barka, E.A. Burkholderia phytofirmans PsJN primes Vitis vinifera L. and confers a better tolerance to low nonfreezing temperatures. Mol. Plant-Microbe Interact. 2012, 25, 241–249. [Google Scholar] [CrossRef] [Green Version]
- Verma, P.; Yadav, A.N.; Khannam, K.S.; Panjiar, N.; Kumar, S.; Saxena, A.K.; Suman, A. Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum) from the northern hills zone of India. Ann. Microbiol. 2015, 65, 1885–1899. [Google Scholar] [CrossRef]
- Subramanian, P.; Kim, K.; Krishnamoorthy, R.; Mageswari, A.; Selvakumar, G.; Sa, T. Cold stress tolerance in psychrotolerant soil bacteria and their conferred chilling resistance in tomato (Solanum lycopersicum Mill.) under low temperatures. PLoS ONE 2016, 11, e0161592. [Google Scholar] [CrossRef] [PubMed]
- Yarzábal, L.A.; Monserrate, L.; Buela, L.; Chica, E. Antarctic Pseudomonas spp. promote wheat germination and growth at low temperatures. Polar Biol. 2018, 41, 2343–2354. [Google Scholar] [CrossRef] [Green Version]
- Zubair, M.; Hanif, A.; Farzand, A.; Sheikh, T.M.M.; Khan, A.R.; Suleman, M.; Ayaz, M.; Gao, X. Genetic screening and expression analysis of psychrophilic Bacillus spp. reveal their potential to alleviate cold stress and modulate phytohormones in wheat. Microorganisms 2019, 7, 337. [Google Scholar] [CrossRef] [Green Version]
- Kakar, K.U.; Ren, X.L.; Nawaz, Z.; Cui, Z.Q.; Li, B.; Xie, G.L.; Hassan, M.A.; Ali, E.; Sun, G.C. A consortium of rhizobacterial strains and biochemical growth elicitors improve cold and drought stress tolerance in rice (Oryza sativa L.). Plant Biol. 2016, 18, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, C.; Gao, Y.L.; Wang, Y.P.; Guo, J.H. A Consortium of three plant growth-promoting rhizobacterium strains acclimates Lycopersicon esculentum and confers a better tolerance to chilling stress. J. Plant Growth Regul. 2016, 35, 54–64. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Maruyama, F.; Ogram, A.V.; Navarrete, O.U.; Lagos, L.M.; Inostroza, N.G.; Acuña, J.J.; Rilling, J.I.; Mora, M.L. Rhizobacterial community structures associated with native plants grown in Chilean extreme environments. Microb. Ecol. 2016, 72, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Orellana, R.; Macaya, C.; Bravo, G.; Dorochesi, F.; Cumsille, A.; Valencia, R.; Rojas, C.; Seeger, M. Living at the frontiers of life: Extremophiles in Chile and their potential for bioremediation. Front. Microbiol. 2018, 9, 2309. [Google Scholar] [CrossRef] [PubMed]
- Jorquera, M.A.; Inostroza, N.G.; Lagos, L.M.; Barra, P.J.; Marileo, L.G.; Rilling, J.I.; Campos, D.C.; Crowley, D.E.; Richardson, A.E.; Mora, M.L. Bacterial community structure and detection of putative plant growth-promoting rhizobacteria associated with plants grown in Chilean agro-ecosystems and undisturbed ecosystems. Biol. Fertil. Soils 2014, 50, 1141–1153. [Google Scholar] [CrossRef]
- Araya, M.A.; Valenzuela, T.; Inostroza, N.G.; Maruyama, F.; Jorquera, M.A.; Acuña, J.J. Isolation and characterization of cold-tolerant hyper-ACC-degrading bacteria from the rhizosphere, endosphere, and phyllosphere of Antarctic vascular plants. Microorganisms 2020, 8, 1788. [Google Scholar] [CrossRef] [PubMed]
- Cavieres, L.A.; Papic, C.; Castor, C. Variación altitudinal en los síndromes de dispersión de semillas de la vegetación andina de la cuenca del Río Molina, Chile central (33 S). Gayana Bot. 1999, 56, 115–123. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Rösch, C.; Mergel, A.; Bothe, H. Biodiversity of denitrifying and dinitrogen-fixing bacteria in an acid forest soil. Appl. Environ. Microbiol. 2002, 68, 3818–3829. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kumar, A.; Pratush, A. Molecular diversity and functional variability of environmental isolates of Bacillus species. Springerplus 2014, 3, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega-Celedón, P.; Canchignia Martínez, H.; González, M.; Seeger, M. Biosíntesis de ácido indol-3-acético y promoción del crecimiento de plantas por bacterias. Cultiv. Trop. 2016, 37, 31–37. [Google Scholar]
- Oberhänsli, T.; Défago, G.; Haas, D. Indole-3-acetic acid (IAA) synthesis in the biocontrol strain CHA0 of Pseudomonas fluorescens: Role of tryptophan side chain oxidase. J. Gen. Microbiol. 1991, 137, 2273–2279. [Google Scholar] [CrossRef] [Green Version]
- Jousset, A.; Schuldes, J.; Keel, C.; Maurhofer, M.; Daniel, R.; Scheu, S.; Thuermer, A. Full-genome sequence of the plant growth-promoting bacterium Pseudomonas protegens CHA0. Genome Announc. 2014, 2, e00322-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cid, F.P.; Inostroza, N.G.; Graether, S.P.; Bravo, L.A.; Jorquera, M.A. Bacterial community structures and ice recrystallization inhibition activity of bacteria isolated from the phyllosphere of the Antarctic vascular plant Deschampsia antarctica. Polar Biol. 2017, 40, 1319–1331. [Google Scholar] [CrossRef]
- Barra, P.J.; Inostroza, N.G.; Acuña, J.J.; Mora, M.L.; Crowley, D.E.; Jorquera, M.A. Formulation of bacterial consortia from avocado (Persea americana Mill.) and their effect on growth, biomass and superoxide dismutase activity of wheat seedlings under salt stress. Appl. Soil Ecol. 2016, 102, 80–91. [Google Scholar] [CrossRef] [Green Version]
- Osorio, M.E.; Quiroz, K.A.; Carvajal, M.A.; Vergara, A.P.; Sánchez, E.Y.; González, C.E.; Catalán, K.S. Synthesis, anti-phytopathogenic and DPPH radical scavenging activities of C-prenylated acetophenones and benzaldehydes. J. Chil. Chem. Soc. 2016, 61, 3095–3101. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, M.; Besoain, X.; Durand, K.; Cesbron, S.; Fuentes, S.; Claverías, F.; Jacques, M.A.; Seeger, M. Clavibacter michiganensis subsp. michiganensis strains from Central Chile exhibit low genetic diversity and sequence types match strains in other parts of the world. Plant Pathol. 2018, 67, 1944–1954. [Google Scholar] [CrossRef]
- Kucheryava, N.; Fiss, M.; Auling, G.; Kroppenstedt, R.M. Isolation and characterization of epiphytic bacteria from the phyllosphere of apple, antagonistic in vitro to Venturia inaequalis, the causal agent of apple scab. Syst. Appl. Microbiol. 1999, 22, 472–478. [Google Scholar] [CrossRef]
- Barrientos-Díaz, L.; Gidekel, M.; Gutiérrez-Moraga, A. Characterization of rhizospheric bacteria isolated from Deschampsia antarctica Desv. World J. Microbiol. Biotechnol. 2008, 24, 2289–2296. [Google Scholar] [CrossRef]
- Strahsburger, E.; Retamales, P.; Estrada, J.; Seeger, M. Microdot method: Used with chromogenic agar is a useful procedure for sanitary monitoring in aquaculture. Lat. Am. J. Aquat. Res. 2016, 44, 742–749. [Google Scholar] [CrossRef]
- Méndez, V.; Fuentes, S.; Morgante, V.; Hernández, M.; González, M.; Moore, E.; Seeger, M. Novel hydrocarbonoclastic metal-tolerant Acinetobacter and Pseudomonas strains from Aconcagua river oil-polluted soil. J. Soil Sci. Plant Nutr. 2017, 17, 1074–1087. [Google Scholar] [CrossRef] [Green Version]
- Bravo, G.; Vega-Celedón, P.; Gentina, J.C.; Seeger, M. Bioremediation by Cupriavidus metallidurans strain MSR33 of mercury-polluted agricultural soil in a rotary drum bioreactor and its effects on nitrogen cycle microorganisms. Microorganisms 2020, 8, 1952. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Patten, C.; Glick, B. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 78, 3795–3801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Zhang, Q.; Guo, J.; Charkowski, A.; Glick, B.; Ibekwe, A.; Cooksey, D.; Yang, C.H. Global effect of indole-3-acetic acid biosynthesis on multiple virulence factors of Erwinia chrysanthemi 3937. Appl. Environ. Microbiol. 2007, 73, 1079–1088. [Google Scholar] [CrossRef] [Green Version]
- Vicente, C.S.L.; Nascimento, F.; Espada, M.; Barbosa, P.; Mota, M.; Glick, B.; Oliveira, S. Characterization of bacteria associated with pinewood nematode Bursaphelenchus xylophilus. PLoS ONE 2012, 7, e46661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Coman, M.M.; Verdenelli, M.C.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Boyko, N.; Cresci, A. In vitro evaluation of antimicrobial activity of Lactobacillus rhamnosus IMC 501®, Lactobacillus paracasei IMC 502® and SYNBIO® against pathogens. J. Appl. Microbiol. 2014, 117, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.A.; Ramos Solano, B.; Montes, F.; Ojeda, J.; Megias, M.; Gutierrez Mañero, F.J. Use of two PGPR strains in the integrated management of blast disease in rice (Oryza sativa) in Southern Spain. Field Crops Res. 2009, 114, 404–410. [Google Scholar] [CrossRef]
- Battini, F.; Bernardi, R.; Turrini, A.; Agnolucci, M.; Giovannetti, M. Rhizophagus intraradices or its associated bacteria affect gene expression of key enzymes involved in the rosmarinic acid biosynthetic pathway of basil. Mycorrhiza 2016, 26, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Penrose, D.M.; Glick, B. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; Hill, P.J.; Dodd, C.E.R.; Laybourn-Parry, J. Demonstration of antifreeze protein activity in Antarctic lake bacteria. Microbiology 2004, 150, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rademaker, J.L.W.; Norman, D.J.; Forster, R.L.; Louws, F.J.; Schultz, M.H.; De Bruijn, F.J. Classification and identification of Xanthomonas translucens isolates, including those pathogenic to ornamental asparagus. Phytopathology 2006, 96, 876–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, T.B.; Gurung, S.; Hansen, J.M.; Bonman, J.M. Pathogenic and genetic diversity of Xanthomonas translucens pv. undulosa in North Dakota. Phytopathology 2012, 102, 390–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, S.D.; Corton, C.; Brown, S.E.; Barron, A.; Clark, L.; Doggett, J.; Harris, B.; Ormond, D.; Quail, M.A.; May, G.; et al. Genome of the actinomycete plant pathogen Clavibacter michiganensis subsp. sepedonicus suggests recent niche adaptation. J. Bacteriol. 2008, 190, 2150–2160. [Google Scholar] [CrossRef] [Green Version]
- Méndez, V.; Valenzuela, M.; Salvà-Serra, F.; Jaén-Luchoro, D.; Besoain, X.; Moore, E.R.B.; Seeger, M. Comparative genomics of pathogenic Clavibacter michiganensis subsp. michiganensis strains from Chile reveals potential virulence features for tomato plants. Microorganisms 2020, 8, 1679. [Google Scholar] [CrossRef]
- Knief, C.; Delmotte, N.; Chaffron, S.; Stark, M.; Innerebner, G.; Wassmann, R.; Von Mering, C.; Vorholt, J.A. Metaproteogenomic analysis of microbial communities in the phyllosphere and rhizosphere of rice. ISME J. 2012, 6, 1378–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, X.; He, R.; Wang, S.; Jiao, C.; Huang, R.; He, X.; Zeng, J.; Zhao, D. The composition and assembly of bacterial communities across the rhizosphere and phyllosphere compartments of phragmites Australis. Diversity 2019, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Idris, R.; Trifonova, R.; Puschenreiter, M.; Wenzel, W.W.; Sessitsch, A. Bacterial communities associated with flowering plants of the Ni hyperaccumulator Thlaspi goesingense. Appl. Environ. Microbiol. 2004, 70, 2667–2677. [Google Scholar] [CrossRef] [Green Version]
- Castillo, U.F.; Browne, L.; Strobel, G.; Hess, W.M.; Ezra, S.; Pacheco, G.; Ezra, D. Biologically active endophytic streptomycetes from Nothofagus spp. and other plants in Patagonia. Microb. Ecol. 2007, 53, 12–19. [Google Scholar] [CrossRef]
- Navarro-Noya, Y.E.; Hernández-Mendoza, E.; Morales-Jiménez, J.; Jan-Roblero, J.; Martínez-Romero, E.; Hernández-Rodríguez, C. Isolation and characterization of nitrogen fixing heterotrophic bacteria from the rhizosphere of pioneer plants growing on mine tailings. Appl. Soil Ecol. 2012, 62, 52–60. [Google Scholar] [CrossRef]
- Tani, A.; Sahin, N.; Kimbara, K. Methylobacterium gnaphalii sp. nov., isolated from leaves of Gnaphalium spicatum. Int. J. Syst. Evol. Microbiol. 2012, 62, 2602–2607. [Google Scholar] [CrossRef]
- Ávila Martínez, E.G.; Lizarazo Forero, L.M.; Cortés Pérez, F. Promoción del crecimiento de Baccharis macrantha (Asteraceae) con bacterias solubilizadoras de fosfatos asociadas a su rizósfera. Acta Biol. Colomb. 2015, 20, 121–131. [Google Scholar] [CrossRef]
- Sánchez-López, A.S.; Ma Del Carmen, A.G.C.; Solís-Domínguez, F.A.; Carrillo-González, R.; Rosas-Saito, G.H. Leaf epiphytic bacteria of plants colonizing mine residues: Possible exploitation for remediation of air pollutants. Front. Microbiol. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Zhang, Q.; Acuña, J.J.; Inostroza, N.G.; Mora, M.L.; Radic, S.; Sadowsky, M.J.; Jorquera, M.A. Endophytic bacterial communities associated with roots and leaves of plants growing in Chilean extreme environments. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.J.; Preece, T.F. A note on Pseudomonas syringae pv. berberidis infections of Berberis: Aetiology of a leaf spot and leaf fall disease in England. J. Appl. Microbiol. 1984, 56, 507–513. [Google Scholar] [CrossRef]
- Russell, G.B.; Bowers, W.S.; Keesing, V.; Niemeyer, H.M.; Sevenet, T.; Vasanthaverni, S.; Wratten, S.D. Patterns of bioactivity and herbivory on Nothofagus species from Chile and New Zealand. J. Chem. Ecol. 2000, 26, 41–56. [Google Scholar] [CrossRef]
- Mølgaard, P.; Holler, J.G.; Asar, B.; Liberna, I.; Rosenbæk, L.B.; Jebjerg, C.P.; Jørgensen, L.; Lauritzen, J.; Guzman, A.; Adsersen, A.; et al. Antimicrobial evaluation of Huilliche plant medicine used to treat wounds. J. Ethnopharmacol. 2011, 138, 219–227. [Google Scholar] [CrossRef]
- González, B.; Vogel, H.; Razmilic, I.; San Martín, J.; Doll, U. Biomass, resin and essential oil content and their variability in natural populations of the Chilean crude drug “bailahuén” (Haplopappus spp.). Bol. Latinoam. Caribe Plantas Med. Aromát. 2012, 11, 66–73. [Google Scholar]
- Ranković, B.; Kosanić, M.; Stanojković, T.; Vasiljević, P.; Manojlović, N. Biological activities of Toninia candida and Usnea barbata together with their norstictic acid and usnic acid constituents. Int. J. Mol. Sci. 2012, 13, 14707–14722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urzúa, A.; Echeverría, J.; Espinoza, J. Lipophilicity and antibacterial activity of flavonols: Antibacterial activity of resinous exudates of Haplopappus litoralis, H. chrysantemifolius and H. scrobiculatus. Bol. Latinoam. Caribe Plantas Med. Aromát. 2012, 11, 369–376. [Google Scholar]
- Nikolić, M.; Marković, T.; Mojović, M.; Pejin, B.; Savić, A.; Perić, T.; Marković, D.; Stević, T.; Soković, M. Chemical composition and biological activity of Gaultheria procumbens L. essential oil. Ind. Crops Prod. 2013, 49, 561–567. [Google Scholar] [CrossRef]
- Concha, J.; Cavieres, L.A.; Sotes, G.J.; Hernández, V. Essential oil composition of Baccharis linearis (Ruiz & Pav.) Pers. and Baccharis paniculata DC. leaves from Chile. Am. J. Essent. Oils Nat. Prod. 2014, 1, 6–8. [Google Scholar]
- Pandey, B.P.; Thapa, R.; Upreti, A. Chemical composition, antioxidant and antibacterial activities of essential oil and methanol extract of Artemisia vulgaris and Gaultheria fragrantissima collected from Nepal. Asian Pac. J. Trop. Med. 2017, 10, 952–959. [Google Scholar] [CrossRef]
- Chaudhary, D.K.; Khulan, A.; Jaisoo, K. Development of a novel cultivation technique for uncultured soil bacteria. Sci. Rep. 2019, 9, 6666. [Google Scholar] [CrossRef]
- Acuña, J.J.; Marileo, L.G.; Araya, M.A.; Rilling, J.I.; Larama, G.A.; Mora, M.L.; Epstein, S.; Jorquera, M.A. In situ cultivation approach to increase the culturable bacterial diversity in the rhizobiome of plants. J. Soil Sci. Plant Nutr. 2020, 20. [Google Scholar] [CrossRef]
- Roh, S.W.; Quan, Z.X.; Do Nam, Y.; Chang, H.W.; Kim, K.H.; Kim, M.K.; Im, W.T.; Jin, L.; Kim, S.H.; Lee, S.T.; et al. Pedobacter agri sp. nov., from soil. Int. J. Syst. Evol. Microbiol. 2008, 58, 1640–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, R.P.; Monchy, S.; Cardinale, M.; Taghavi, S.; Crossman, L.; Avison, M.B.; Berg, G.; van der Lelie, D.; Dow, J.M. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat. Rev. Microbiol. 2009, 7, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Melnick, R.L.; Suárez, C.; Bailey, B.A.; Backman, P.A. Isolation of endophytic endospore-forming bacteria from Theobroma cacao as potential biological control agents of cacao diseases. Biol. Control 2011, 57, 236–245. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.C.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, L.M.; Tisa, L.S. Friend or foe? a review of the mechanisms that drive Serratia towards diverse lifestyles. Can. J. Microbiol. 2013, 59, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, H.C.; Zhou, Y.G.; Xin, Y.H. Microevolution and adaptive strategy of psychrophilic species Flavobacterium bomense sp. nov. isolated from glaciers. Front. Microbiol. 2019, 10, 1069. [Google Scholar] [CrossRef]
- Busse, H.J. Review of the taxonomy of the genus Arthrobacter, emendation of the genus Arthrobacter sensu lato, proposal to reclassify selected species of the genus Arthrobacter in the novel genera Glutamicibacter gen. nov., Paeniglutamicibacter gen. nov., Pseudoglutamicibacter gen. nov., Paenarthrobacter gen. nov., and Pseudarthrobacter gen. nov., and emended description of Arthrobacter roseus. Int. J. Syst. Evol. Microbiol. 2016, 66, 9–37. [Google Scholar] [CrossRef] [Green Version]
- Cid, F.P.; Maruyama, F.; Murase, K.; Graether, S.P.; Larama, G.; Bravo, L.A.; Jorquera, M.A. Draft genome sequences of bacteria isolated from the Deschampsia antarctica phyllosphere. Extremophiles 2018, 22, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Kay, M.; Cooper, K.K. Comparative genomics of cocci-shaped Sporosarcina strains with diverse spatial isolation. BMC Genom. 2018, 19, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Abraham, W.R.; Strömpl, C.; Meyer, H.; Lindholst, S.; Moore, E.R.; Christ, R.; Vancanneyt, M.; Tindall, B.J.; Bennasar, A.; Smit, J.; et al. Phylogeny and polyphasic taxonomy of Caulobacter species. Proposal of Maricaulis gen. nov. with Maricaulis maris (Poindexter) comb. nov. as the type species, and emended description of the genera Brevundimonas and Caulobacter. Int. J. Syst. Bacteriol. 1999, 49, 1053–1073. [Google Scholar] [CrossRef] [Green Version]
- Tak, E.J.; Kim, P.S.; Hyun, D.W.; Kim, H.S.; Lee, J.Y.; Kang, W.; Sung, H.; Shin, N.R.; Kim, M.S.; Whon, T.W.; et al. Phenotypic and genomic properties of Brachybacterium vulturis sp. nov. and Brachybacterium avium sp. nov. Front. Microbiol. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Greene, A.C.; Euzéby, J.P.; Tindall, B.J.; Patel, B.K.C. Proposal of Frondihabitans gen. nov. to replace the illegitimate genus name Frondicola Zhang et al. 2007. Int. J. Syst. Evol. Microbiol. 2009, 59, 447–448. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.D. Frondihabitans peucedani sp. nov., an actinobacterium isolated from rhizosphere soil, and emended description of the genus Frondihabitans Greene et al. 2009. Int. J. Syst. Evol. Microbiol. 2010, 60, 1740–1744. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, M.; Grube, M.; Berg, G. Frondihabitans cladoniiphilus sp. nov., an actinobacterium of the family Microbacteriaceae isolated from lichen, and emended description of the genus Frondihabitans. Int. J. Syst. Evol. Microbiol. 2011, 61, 3033–3038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Lim, J.M.; Ahn, J.H.; Weon, H.Y.; Hamada, M.; Suzuki, K.I.; Ahn, T.Y.; Kwon, S.W. Description of Galbitalea soli gen. nov., sp. nov., and Frondihabitans sucicola sp. nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Han, S.R.; Yu, S.C.; Kang, S.; Park, H.; Oh, T.J. Complete genome sequence of Frondihabitans sp. strain PAMC28766, a novel carotenoid-producing and radiation-resistant strain isolated from an Antarctic lichen. J. Biotechnol. 2016, 226, 20–21. [Google Scholar] [CrossRef]
- Eichenlaub, R.; Gartemann, K.-H. The Clavibacter michiganensis subspecies: Molecular investigation of Gram-positive bacterial plant pathogens. Annu. Rev. Phytopathol. 2011, 49, 445–464. [Google Scholar] [CrossRef]
- Timilsina, S.; Potnis, N.; Newberry, E.A.; Liyanapathiranage, P.; Iruegas-Bocardo, F.; White, F.F.; Goss, E.M.; Jones, J.B. Xanthomonas diversity, virulence and plant–pathogen interactions. Nat. Rev. Microbiol. 2020, 18, 415–427. [Google Scholar] [CrossRef]
- Kuddus, M.; Ramteke, P.W. A cold-active extracellular metalloprotease from Curtobacterium luteum (MTCC 7529): Enzyme production and characterization. J. Gen. Appl. Microbiol. 2008, 54, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Undabarrena, A.; Beltrametti, F.; Claverías, F.P.; González, M.; Moore, E.R.B.; Seeger, M.; Cámara, B. Exploring the diversity and antimicrobial potential of marine actinobacteria from the Comau Fjord in Northern Patagonia, Chile. Front. Microbiol. 2016, 7, 1135. [Google Scholar] [CrossRef]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Saxena, A.K. Bioprospecting of plant growth promoting psychrotrophic Bacilli from the cold desert of north western Indian Himalayas. Indian J. Exp. Biol. 2016, 54, 142–150. [Google Scholar] [PubMed]
- Yadav, A.N.; Yadav, N.; Sachan, S.G.; Saxena, A.K. Biodiversity of psychrotrophic microbes and their biotechnological applications. J. Appl. Biol. Biotechnol. 2019, 7, 99–108. [Google Scholar] [CrossRef]
- Ortíz-Ojeda, P.; Ogata-Gutiérrez, K.; Zúñiga-Dávila, D. Evaluation of plant growth promoting activity and heavy metal tolerance of psychrotrophic bacteria associated with maca (Lepidium meyenii Walp.) rhizosphere. AIMS Microbiol. 2017, 3, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J.; Mulet, M.; Gomila, M.; García-Valdés, E. Genomics in bacterial taxonomy: Impact on the genus Pseudomonas. Genes 2020, 11, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitaraman, R. Pseudomonas spp. as models for plant-microbe interactions. Front. Plant Sci. 2015, 6, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.; Villalobos, P.; Morgante, V.; González, M.; Reiff, C.; Moore, E.; Seeger, M. Isolation and characterization of a novel simazine degrading bacterium from agricultural soil of Central Chile, Pseudomonas sp. MHP41. FEMS Microbiol. Lett. 2008, 286, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canchignia, H.; Altimira, F.; Montes, C.; Sánchez, E.; Tapia, E.; Miccono, M.; Espinoza, D.; Aguirre, C.; Seeger, M.; Prieto, H. Candidate nematicidal proteins in a new Pseudomonas veronii isolate identified by its antagonistic properties against Xiphinema index. J. Gen. Appl. Microbiol. 2017, 63, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, M.; Morgante, V.; Ávila, M.; Villalobos, P.; Millares, P.; González, M.; Seeger, M. Novel s-triazine-degrading bacteria isolated from agricultural soils of Central Chile for herbicide bioremediation. Electron. J. Biotechnol. 2008, 11, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Hayward, A.C.; Fegan, N.; Fegan, M.; Stirling, G.R. Stenotrophomonas and Lysobacter: Ubiquitous plant-associated gamma-proteobacteria of developing significance in applied microbiology. J. Appl. Microbiol. 2010, 108, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Altimira, F.; Yáñez, C.; Bravo, G.; González, M.; Rojas, L.A.; Seeger, M. Characterization of copper-resistant bacteria and bacterial communities from copper-polluted agricultural soils of Central Chile. BMC Microbiol. 2012, 12, 193. [Google Scholar] [CrossRef] [Green Version]
- Wolf, A.; Fritze, A.; Hagemann, M.; Berg, G. Stenotrophomonas rhizophila sp. nov., a novel plant-associated bacterium with antifungal properties. Int. J. Syst. Evol. Microbiol. 2002, 52, 1937–1944. [Google Scholar] [CrossRef] [PubMed]
- Lindow, S.E. Methods of preventing frost injury through control of epiphytic ice nucleation active bacteria. Plant Dis. 1983, 67, 327–333. [Google Scholar] [CrossRef]
- Subramoni, S.; Nathoo, N.; Klimov, E.; Yuan, Z.C. Agrobacterium tumefaciens responses to plant-derived signaling molecules. Front. Plant Sci. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.J.; Yang, W.; Wang, C.; Gu, C.; Niu, D.D.; Liu, H.X.; Wang, Y.P.; Guo, J.H. Induction of drought tolerance in cucumber plants by a consortium of three plant growth-promoting rhizobacterium strains. PLoS ONE 2012, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Molina-Romero, D.; Baez, A.; Quintero-Hernández, V.; Castañeda-Lucio, M.; Fuentes-Ramírez, L.E.; Bustillos-Cristales, M.D.R.; Rodríguez-Andrade, O.; Morales-García, Y.E.; Munive, A.; Muñoz-Rojas, J. Compatible bacterial mixture, tolerant to desiccation, improves maize plant growth. PLoS ONE 2017, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Saikia, J.; Sarma, R.K.; Dhandia, R.; Yadav, A.; Bharali, R.; Gupta, V.K.; Saikia, R. Alleviation of drought stress in pulse crops with ACC deaminase producing rhizobacteria isolated from acidic soil of Northeast India. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Pandey, S. ACC deaminase producing bacteria with multifarious plant growth promoting traits alleviates salinity stress in French Bean (Phaseolus vulgaris) plants. Front. Microbiol. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Bacterial consortium for improved maize (Zea mays L.) production. Microorganisms 2019, 7, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahara, H.; Nakano, Y.; Omiya, K.; Muryoi, N.; Nishikawa, J.; Obata, H. Production of two types of ice crystal-controlling proteins in Antarctic bacterium. J. Biosci. Bioeng. 2004, 98, 220–223. [Google Scholar] [CrossRef]
- Yu, S.O.; Brown, A.; Middleton, A.J.; Tomczak, M.M.; Walker, V.K.; Davies, P.L. Ice restructuring inhibition activities in antifreeze proteins with distinct differences in thermal hysteresis. Cryobiology 2010, 61, 327–334. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Naveed, M.; Ayyub, M.; Khan, M.Y.; Ahmad, M.; Zahir, Z.A. Potential, limitations and future prospects of Pseudomonas spp. for sustainable agriculture and environment: A review. Soil Environ. 2016, 35, 106–145. [Google Scholar]
- Subramanian, P.; Mageswari, A.; Kim, K.; Lee, Y.; Sa, T. Psychrotolerant endophytic Pseudomonas sp. strains OB155 and OS261 induced chilling resistance in tomato plants (Solanum lycopersicum Mill.) by activation of their antioxidant capacity. Mol. Plant-Microbe Interact. 2015, 28, 1073–1081. [Google Scholar] [CrossRef] [Green Version]
- Chase, A.B.; Arevalo, P.; Polz, M.F.; Berlemont, R.; Martiny, J.B.H. Evidence for ecological flexibility in the cosmopolitan genus Curtobacterium. Front. Microbiol. 2016, 7, 1–11. [Google Scholar] [CrossRef]
- Díez-Méndez, A.; Rivas, R. Improvement of saffron production using Curtobacterium herbarum as a bioinoculant under greenhouse conditions. AIMS Microbiol. 2017, 3, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.; de Quadros, P.D.; Fulthorpe, R. Plantibacter flavus, Curtobacterium herbarum, Paenibacillus taichungensis, and Rhizobium selenitireducens endophytes provide host-specific growth promotion of Arabidopsis thaliana, basil, lettuce, and bok choy plants. Appl. Environ. Microbiol. 2019, 85, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barriuso, J.; Ramos Solano, B.; Gutiérrez Mañero, F.J. Protection against pathogen and salt stress by four plant growth-promoting rhizobacteria isolated from Pinus sp. on Arabidopsis thaliana. Phytopathology 2008, 98, 666–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardinale, M.; Ratering, S.; Suarez, C.; Zapata Montoya, A.M.; Geissler-Plaum, R.; Schnell, S. Paradox of plant growth promotion potential of rhizobacteria and their actual promotion effect on growth of barley (Hordeum vulgare L.) under salt stress. Microbiol. Res. 2015, 181, 22–32. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Ullah, I.; Ali, S.; Kang, S.M.; Lee, I.J. Alleviation of salt stress response in soybean plants with the endophytic bacterial isolate Curtobacterium sp. SAK1. Ann. Microbiol. 2019, 69, 797–808. [Google Scholar] [CrossRef]
- Vimal, S.R.; Patel, V.K.; Singh, J.S. Plant growth promoting Curtobacterium albidum strain SRV4: An agriculturally important microbe to alleviate salinity stress in paddy plants. Ecol. Indic. 2019, 105, 553–562. [Google Scholar] [CrossRef]
- Irizarry, I.; White, J.F. Application of bacteria from non-cultivated plants to promote growth, alter root architecture and alleviate salt stress of cotton. J. Appl. Microbiol. 2017, 122, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.; Koskella, B. Nutrient- and dose-dependent microbiome-mediated protection against a plant pathogen. Curr. Biol. 2018, 28, 2487–2492.e3. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Zhang, Q.; Chen, F.; Zhu, Q.; Wang, Y.; Liu, G. Remediation of resins-contaminated soil by the combination of electrokinetic and bioremediation processes. Environ. Pollut. 2020, 260, 114047. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Buyer, J.S. Culturable bacterial microflora associated with nectarine fruit and their potential for control of brown rot. Can. J. Microbiol. 2010, 56, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.; Lauga, B.; Miot-Sertier, C.; Mercier, A.; Lonvaud, A.; Soulas, M.L.; Soulas, G.; Masneuf-Pomarède, I. Characterization of epiphytic bacterial communities from grapes, leaves, bark and soil of grapevine plants grown, and their relations. PLoS ONE 2013, 8, e73013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandel, S.L.; Firrincieli, A.; Joubert, P.M.; Okubara, P.A.; Leston, N.D.; McGeorge, K.M.; Mugnozza, G.S.; Harfouche, A.; Kim, S.H.; Doty, S.L. An in vitro study of bio-control and plant growth promotion potential of Salicaceae endophytes. Front. Microbiol. 2017, 8, 386. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Wang, J.T.; Zhang, Z.F.; Li, W.; Chen, W.; Cai, L. Microbiota in the rhizosphere and seed of rice from China, with reference to their transmission and biogeography. Front. Microbiol. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5’-3’) | Gene Target | Size (pb) | Reference |
|---|---|---|---|---|
| 27F | AGAGTTTGATCMTGGCTCAG | 16S ARNr | 1465 | [35] |
| 1492R | TACGGYTACCTTGTTACGACTT | |||
| nifH-F-Rösch | AAAGGYGGWATCGGYAARTCCACCA | nifH | 458 | [36] |
| nifH-R-Rösch | TTGTTSGCSGCRTACATSGCCATCAT | |||
| DegACC-F | GGBGGVAAYAARMYVMGSAAGCTYGA | acdS | 750 | [37] |
| DegACC-R | TTDCCHKYRTANACBGGRTC |
| Number of Isolates | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rhizosphere | Phyllosphere | |||||||||||||
| Wild plant (Code for Isolates) | Total | Id | L | M | H | VH | E | Total | Id | L | M | H | VH | E |
| Calycera sp. (Cp) | 16 | 11 | 0 | 5 | 0 | 4 | 7 | 3 | 3 | 0 | 0 | 1 | 0 | 2 |
| Thlaspi sp. (Tm) | 17 | 13 | 3 | 1 | 4 | 7 | 2 | 10 | 8 | 0 | 2 | 2 | 2 | 4 |
| Haplopappus sp. (Hs) | 9 | 3 | 0 | 6 | 1 | 0 | 2 | 2 | 1 | 1 | 0 | 0 | 1 | 0 |
| Baccharis sp. (Bl) | 10 | 3 | 1 | 6 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Gnaphalium sp. (Gc) | 18 | 9 | 3 | 6 | 4 | 2 | 3 | 2 | 2 | 0 | 0 | 1 | 0 | 1 |
| Gaultheria sp. (Gp) | 8 | 1 | 1 | 6 | 0 | 1 | 0 | 2 | 1 | 1 | 0 | 0 | 0 | 1 |
| Nothofagus sp. + Usnea sp. (NU) | 7 | 2 | 0 | 5 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| Orites sp. (Om) | 3 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chusquea sp. (Cc) | 10 | 3 | 1 | 6 | 2 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Berberis sp. (Bm) | 5 | 3 | 0 | 2 | 1 | 0 | 2 | 6 | 6 | 0 | 0 | 1 | 2 | 3 |
| Nothofagus sp. (Nd) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total isolates | 103 | 49 | 10 | 44 | 13 | 19 | 17 | 27 | 21 | 3 | 3 | 5 | 5 | 11 |
| Strains | PS at RT/PS at 4 °C | nifH/ acdS | Strains | PS at RT/ 4 °C | nifH/ acdS |
|---|---|---|---|---|---|
| Pseudomonas sp. TmR1b * | +/+ | +/− | Stenotrophomonas sp. TmP43c * | −/− | +/+ |
| Pseudomonas sp. TmR34a | +/+ | +/− | Stenotrophomonas sp. TmR19b | −/− | +/− |
| Pseudomonas sp. TmR35a | +/+ | +/− | Brevundimonas sp. CpR34a | −/− | +/− |
| Pseudomonas sp. TmR5a * | +/+ | +/+ | Arthrobacter sp. BmP28 * | +/− | +/+ |
| Pseudomonas sp. GcR15a * | −/− | +/+ | Arthrobacter sp. BmR37 | +/− | +/− |
| Pseudomonas sp. NUR4a * | +/+ | −/+ | Arthrobacter sp. CpR32a | −/− | +/− |
| Pseudomonas sp. BlR7b * | +/+ | +/+ | Paenarthrobacter sp. BlR11a | −/− | +/− |
| Pseudomonas sp. BlR3 * | +/+ | +/+ | Pseudarthrobacter sp. TmP42 | −/− | +/− |
| Pseudomonas sp. TmR7 * | +/+ | +/+ | Pseudarthrobacter sp. GcR10 | −/− | +/− |
| Pseudomonas sp. CcR1d * | +/+ | +/− | Curtobacterium sp. GcR11a | −/− | +/− |
| Serratia sp. CpR7a | +/+ | −/− | Curtobacterium sp. BmP22c * | −/− | −/+ |
| Serratia sp. TmR3a * | +/+ | +/− | Curtobacterium sp. BmP18c * | −/− | −/− |
| Stenotrophomonas sp. BmP26c | −/− | +/− | Brachybacterium sp. TmP24d | −/− | +/+ |
| Stenotrophomonas sp. BmR35 * | −/− | +/+ | Brachybacterium sp. TmP30 | −/− | +/+ |
| Stenotrophomonas sp. CpP23d * | −/− | +/+ | Frondihabitans sp. GpP26d * | −/− | +/+ |
| Stenotrophomonas sp. CpR10 * | −/− | +/+ | Sporosarcina sp. HsP12b * | −/− | +/+ |
| Stenotrophomonas sp. CpR13a | −/− | +/+ | Sporosarcina sp. HsR8 * | −/− | +/+ |
| Stenotrophomonas sp. CpR8a | −/− | +/+ | Bacillus sp. GcP2b | +/− | −/− |
| Stenotrophomonas sp. CpR12a | −/− | +/− | Pedobacter sp. CpR35b | −/− | −/− |
| Stenotrophomonas sp. GpR8b | −/− | +/+ | Flavobacterium sp. CpR5c * | +/+ | +/− |
| Stenotrophomonas sp. GcR23 * | −/− | +/+ |
| Strains | Pss | Pc | Cmm | At | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| YM | MH | KB | YM | MH | KB | YM | MH | YM | MH | |
| Pseudomonas sp. TmR1b * | + | − | ++ | + | − | +/− | + | ++ | − | − |
| Pseudomonas sp. TmR5a * | − | − | + | − | − | − | + | + | − | − |
| Pseudomonas sp. GcR15a * | +/− | − | + | + | − | +/− | ++ | + | − | − |
| Pseudomonas sp. NUR4a * | ++ | − | + | ++ | − | − | ++ | ++ | + | − |
| Pseudomonas sp. BlR7b | − | − | − | − | − | − | − | − | − | − |
| Pseudomonas sp. BlR3 | − | − | − | − | − | − | − | − | − | − |
| Pseudomonas sp. TmR7 * | − | − | − | +/− | − | − | +/− | +/− | − | − |
| Pseudomonas sp. CcR1d * | − | − | + | − | − | +/− | ++ | ++ | + | − |
| Serratia sp. TmR3a | − | − | − | − | − | − | + | − | − | − |
| Stenotrophomonas sp. BmR35 | − | − | − | − | − | − | − | − | − | − |
| Stenotrophomonas sp. CpP23d | − | − | − | − | − | − | − | + | − | − |
| Stenotrophomonas sp. CpR10 | − | − | − | − | − | − | − | − | − | − |
| Stenotrophomonas sp. GcR23 | − | − | − | − | − | − | − | − | − | − |
| Stenotrophomonas sp. TmP43c | − | − | − | − | − | − | − | − | − | − |
| Arthrobacter sp. BmP28 * | + | − | + | + | − | + | ++ | − | ++ | − |
| Curtobacterium sp. BmP22c * | + | − | − | ++ | − | − | ++ | +/− | ++ | − |
| Brachybacterium sp. TmP30 * | − | + | − | − | − | − | − | +/− | − | − |
| Frondihabitans sp. GpP26d * | + | − | − | + | − | − | ++ | + | + | + |
| Sporosarcina sp. HsP12b | − | − | − | − | − | − | − | − | − | − |
| Sporosarcina sp. HsR8 | − | − | − | − | − | − | +/− | +/− | − | − |
| Flavobacterium sp. CpR5c | − | − | − | − | − | − | − | +/− | − | − |
| Pseudomonas protegens CHA0 | + | − | + | ++ | − | + | ++ | + | + | − |
| Strains | TmR1b | TmR5a | GcR15a | NUR4a | TmR7 | CcR1d | BmP28 | BmP22c | TmP30 | GpP26d |
|---|---|---|---|---|---|---|---|---|---|---|
| TmR1b | + | + | − | − | − | − | + | − | − | |
| TmR5a | + | − | − | − | − | − | + | − | − | |
| GcR15a | + | − | − | − | − | − | − | − | − | |
| NUR4a | − | − | − | − | + | − | − | − | − | |
| TmR7 | − | − | − | − | − | + | + | − | + | |
| CcR1d | − | − | − | + | − | − | − | − | − | |
| BmP28 | − | − | − | − | + | − | − | + | + | |
| BmP22c | + | + | − | − | + | − | − | + | − | |
| TmP30 | − | − | − | − | − | − | + | + | − | |
| GpP26d | − | − | − | − | + | − | + | − | − |
| N° | Bacterial Consortia | PT Potential | Plant Growth-Promoting Activities/Determinations | Antimicrobial Activities | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BCS% | [Auxin] */ OD600nm | PS at RT/ PS at 4 °C | nifH | acdS | Pss | Pc | Cmm | At | ||
| BC1 | Pseudomonas sp. TmR7 | 100 | 0.91 | +/+ | + | + | − | + | + | − |
| Frondihabitans sp. GpP26d | 93.26 | 5.146 | −/− | + | + | + | + | + | + | |
| BC2 | Pseudomonas sp. CcR1d | 78.88 | 0.074 | +/+ | + | − | + | + | + | + |
| Pseudomonas sp. NUR4a | 70.55 | 1.342 | +/+ | − | + | + | + | + | + | |
| BC3 | Pseudomonas sp. TmR5a | 56.23 | 1.223 | +/+ | + | + | + | − | + | − |
| Curtobacterium sp. BmP22c | 84.5 | 1.37 | −/− | − | + | + | + | + | + | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vega-Celedón, P.; Bravo, G.; Velásquez, A.; Cid, F.P.; Valenzuela, M.; Ramírez, I.; Vasconez, I.-N.; Álvarez, I.; Jorquera, M.A.; Seeger, M. Microbial Diversity of Psychrotolerant Bacteria Isolated from Wild Flora of Andes Mountains and Patagonia of Chile towards the Selection of Plant Growth-Promoting Bacterial Consortia to Alleviate Cold Stress in Plants. Microorganisms 2021, 9, 538. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030538
Vega-Celedón P, Bravo G, Velásquez A, Cid FP, Valenzuela M, Ramírez I, Vasconez I-N, Álvarez I, Jorquera MA, Seeger M. Microbial Diversity of Psychrotolerant Bacteria Isolated from Wild Flora of Andes Mountains and Patagonia of Chile towards the Selection of Plant Growth-Promoting Bacterial Consortia to Alleviate Cold Stress in Plants. Microorganisms. 2021; 9(3):538. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030538
Chicago/Turabian StyleVega-Celedón, Paulina, Guillermo Bravo, Alexis Velásquez, Fernanda P. Cid, Miryam Valenzuela, Ingrid Ramírez, Ingrid-Nicole Vasconez, Inaudis Álvarez, Milko A. Jorquera, and Michael Seeger. 2021. "Microbial Diversity of Psychrotolerant Bacteria Isolated from Wild Flora of Andes Mountains and Patagonia of Chile towards the Selection of Plant Growth-Promoting Bacterial Consortia to Alleviate Cold Stress in Plants" Microorganisms 9, no. 3: 538. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030538