
Actinotignum schaalii: Relation to Concomitants and Connection to Patients’ Conditions in Polymicrobial Biofilms of Urinary Tract Catheters and Urines

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Data Collection
2.2. Preanalytical Procedure
2.3. PCR-DGGE-S and Culture
2.3.1. PCR-DGGE-S
2.3.2. Culture
2.3.3. Culture and PCR-DGGE-S Data Analysis and Interpretation
2.4. 16S rRNA Amplicon Sequencing
16S rRNA Amplicon Sequencing Data Analysis and Interpretation
3. Results
3.1. Culture and PCR-DGGE-S
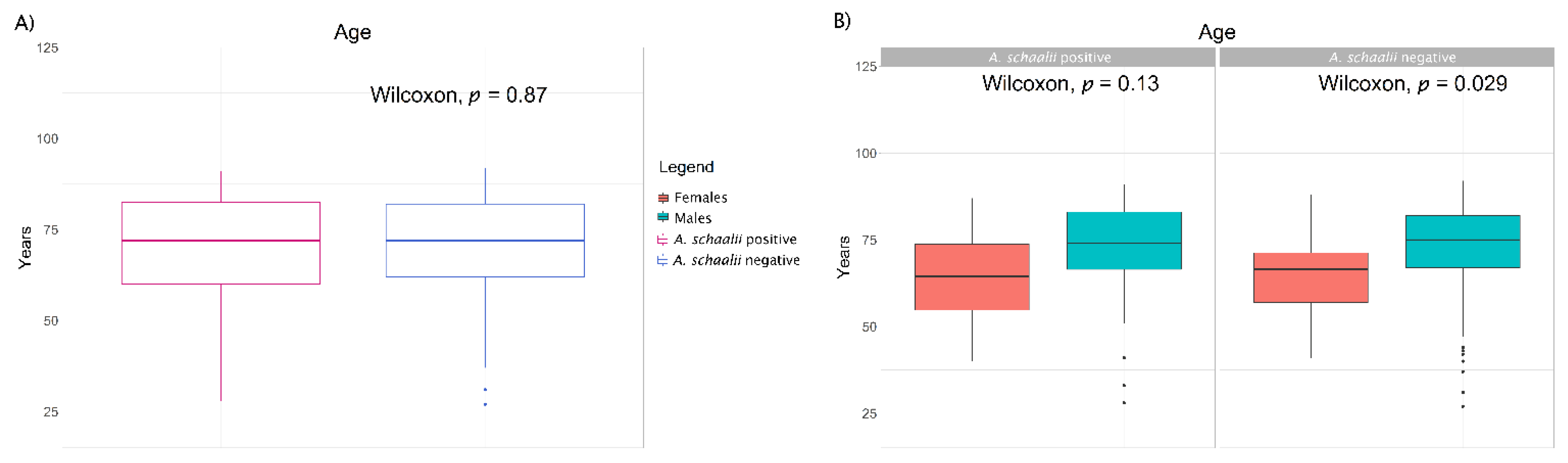
3.1.1. A. schaalii and Explanatory Variables
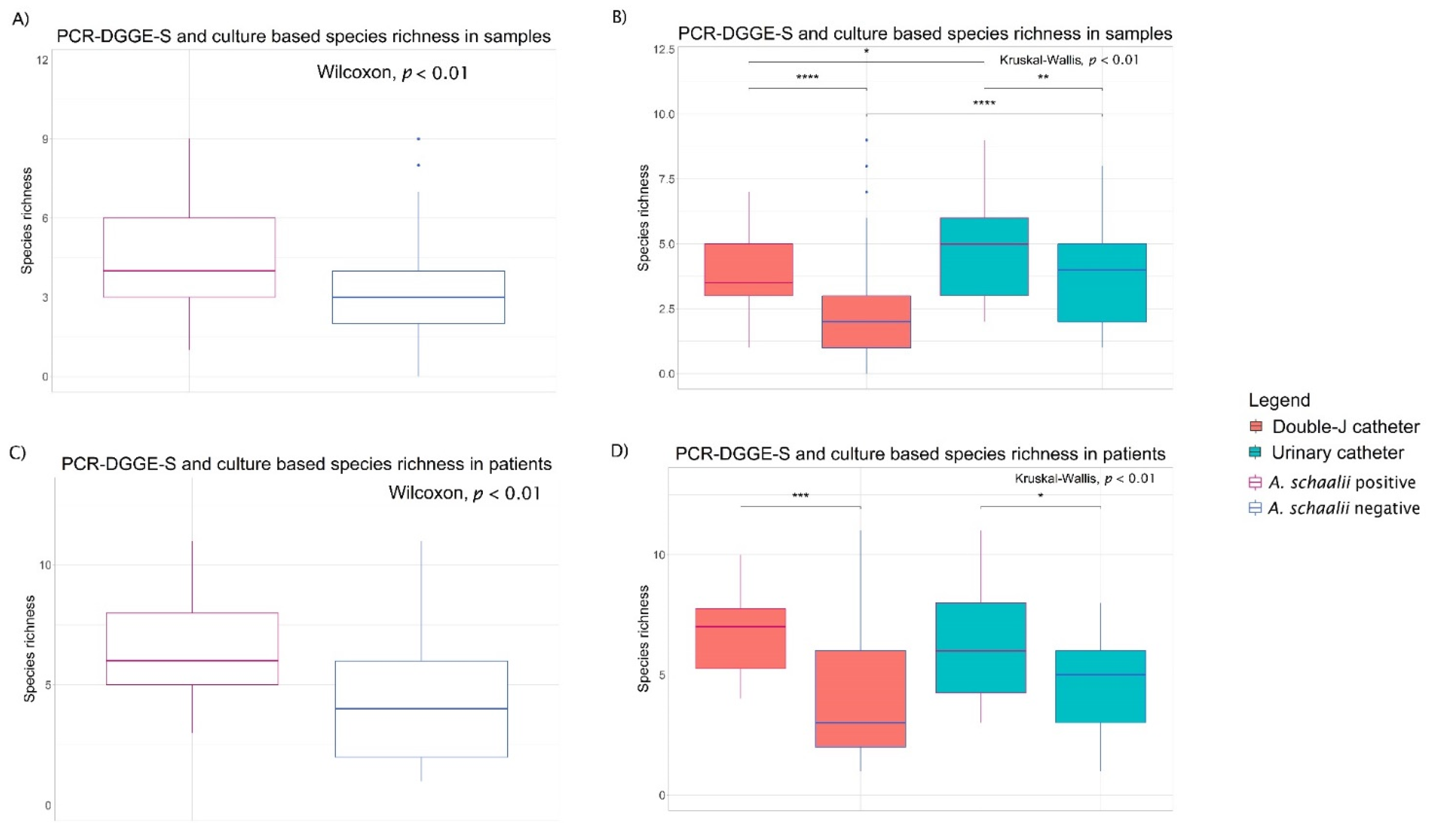
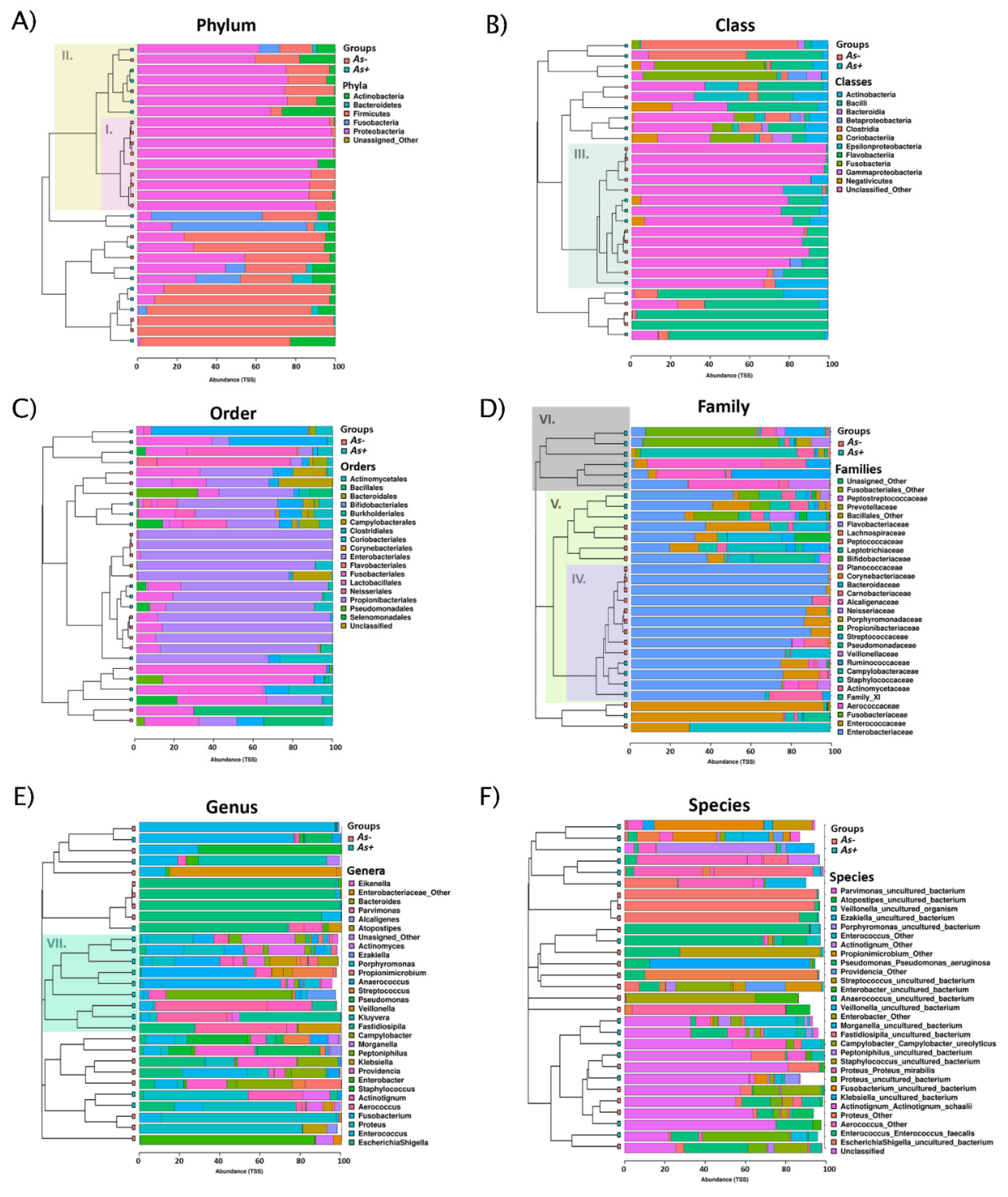
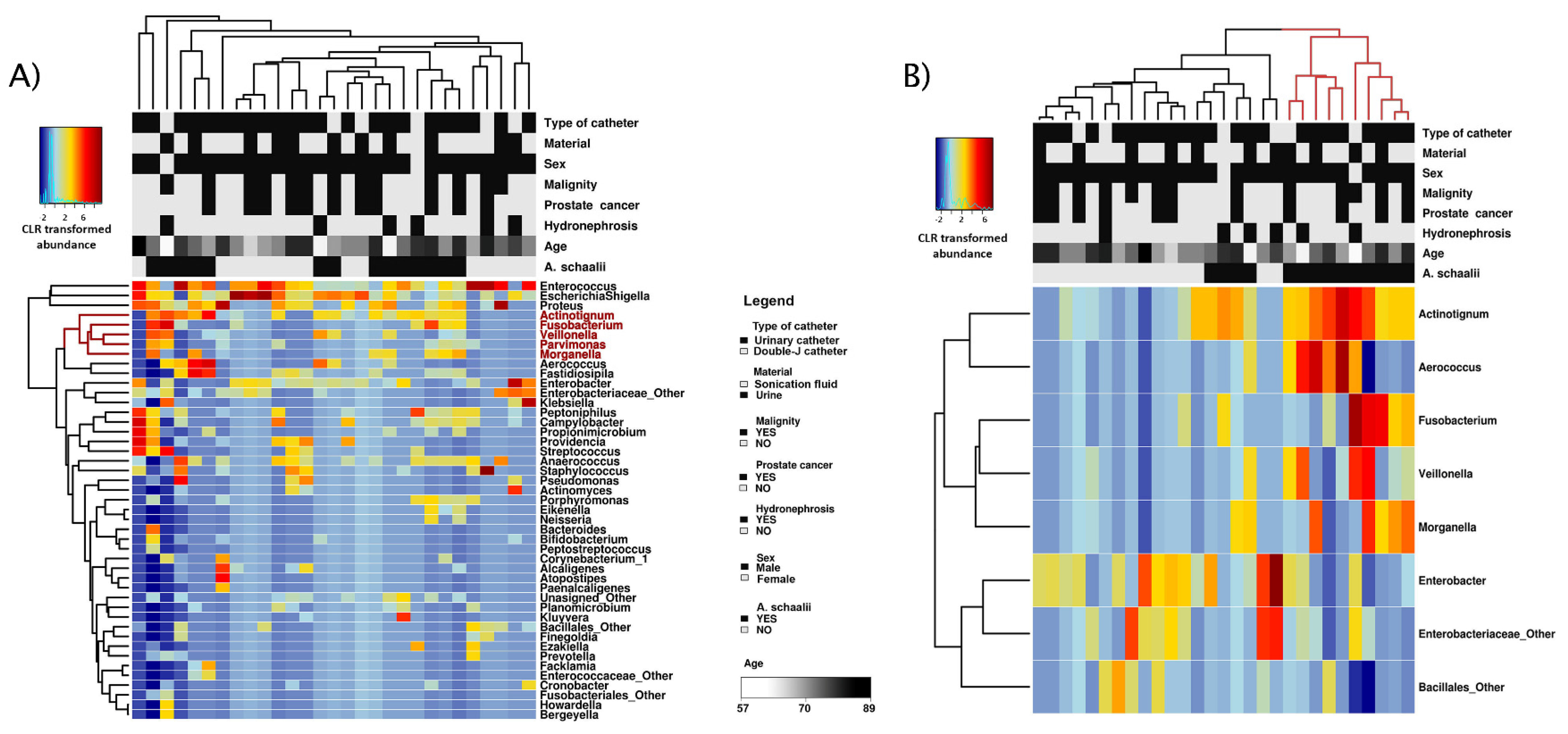
3.1.2. Diversity of Bacterial Communities
3.1.3. Concomitant Species of A. schaalii-Positive Patients
3.2. 16S rRNA Amplicon Sequencing
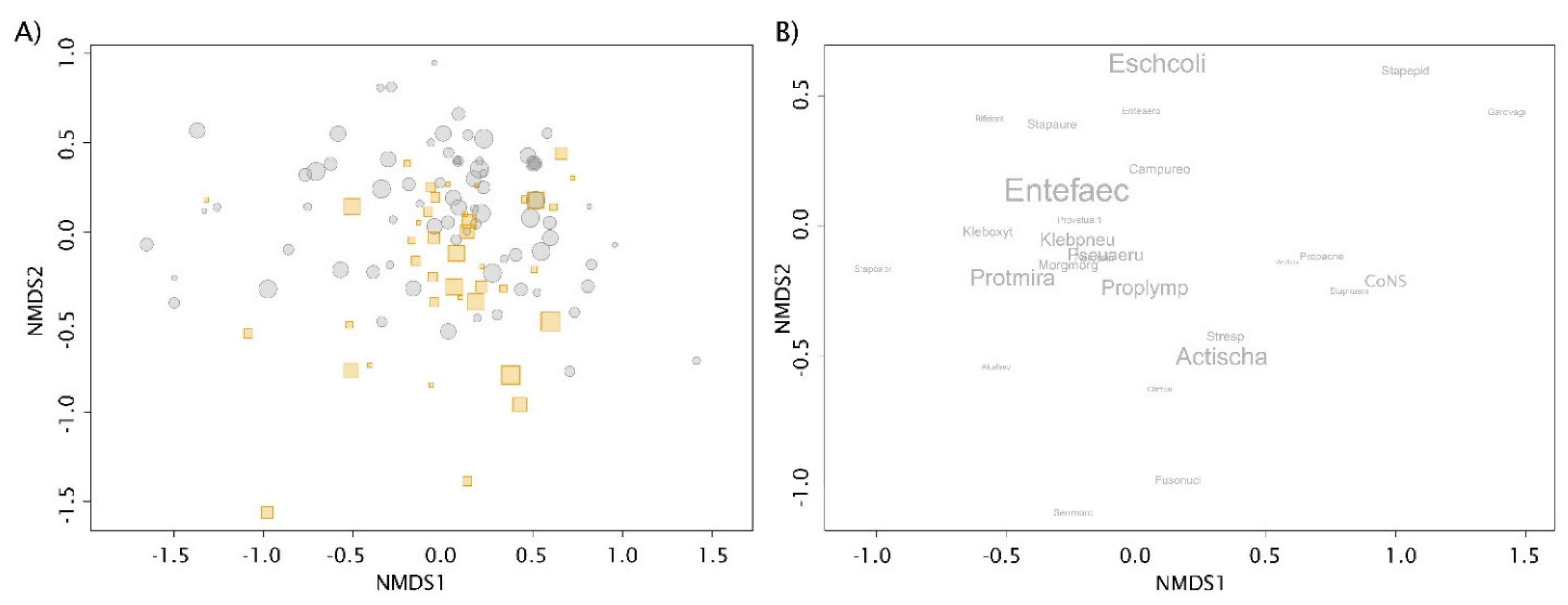
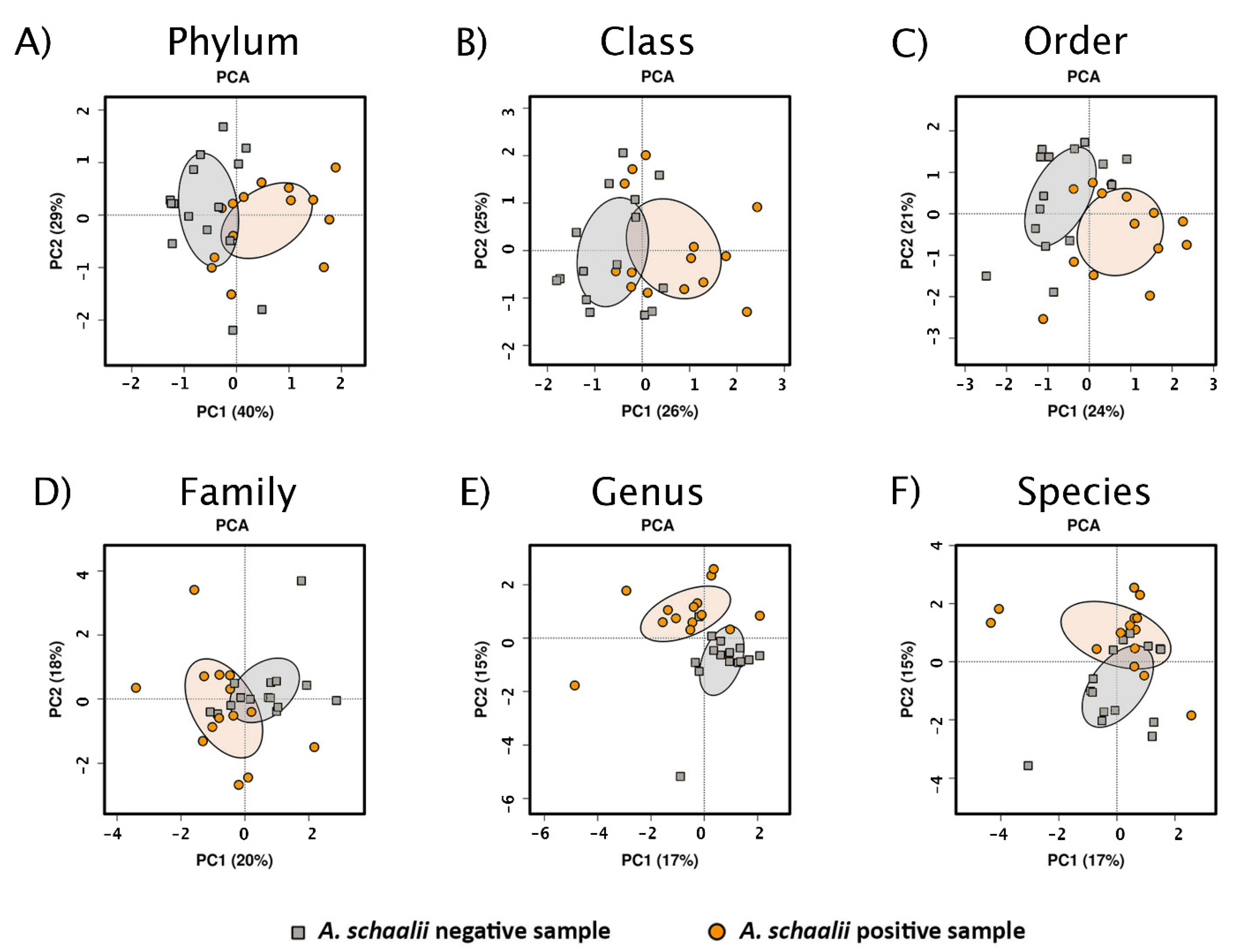
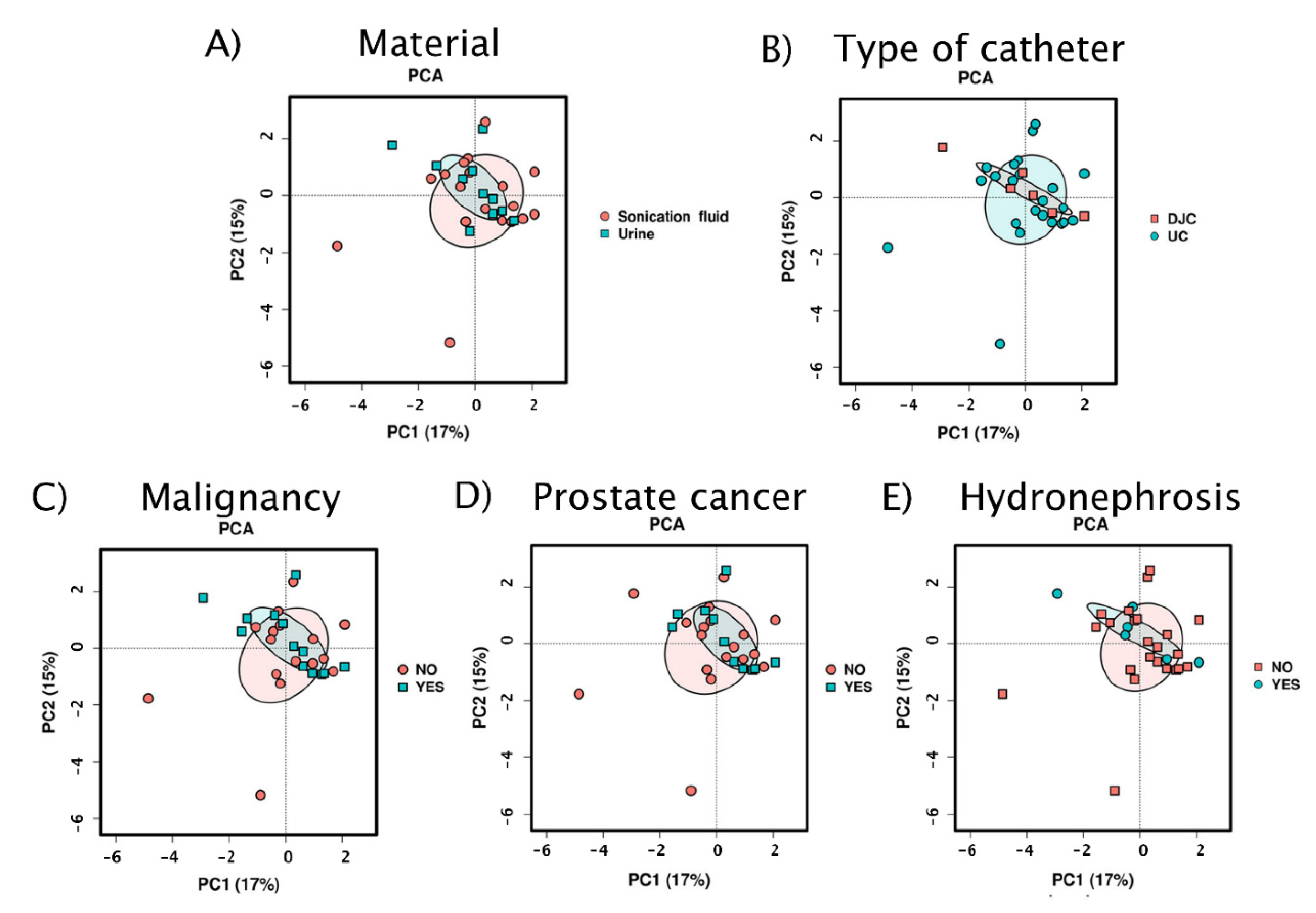
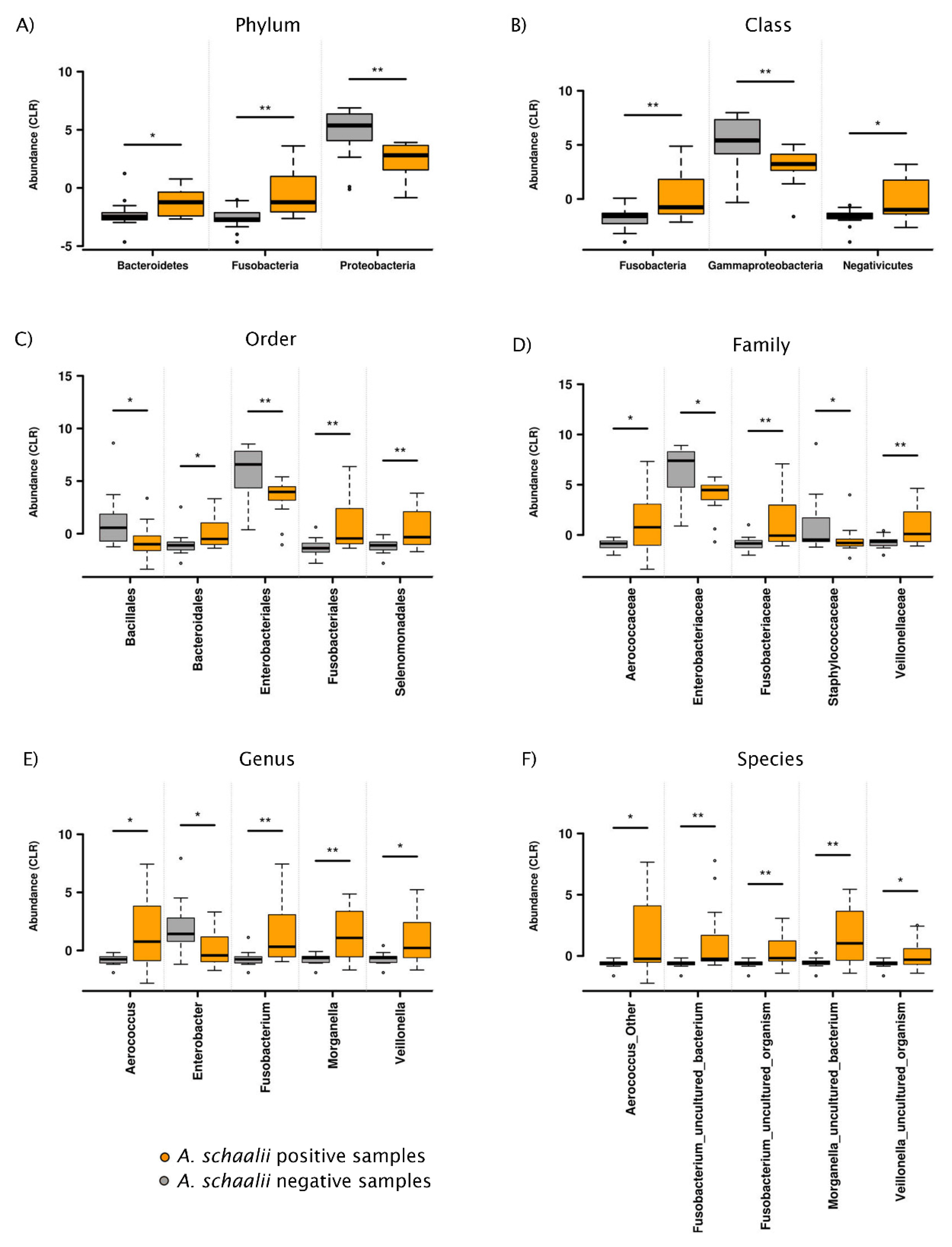
3.2.1. Exploratory Analyses
3.2.2. Alpha Diversity
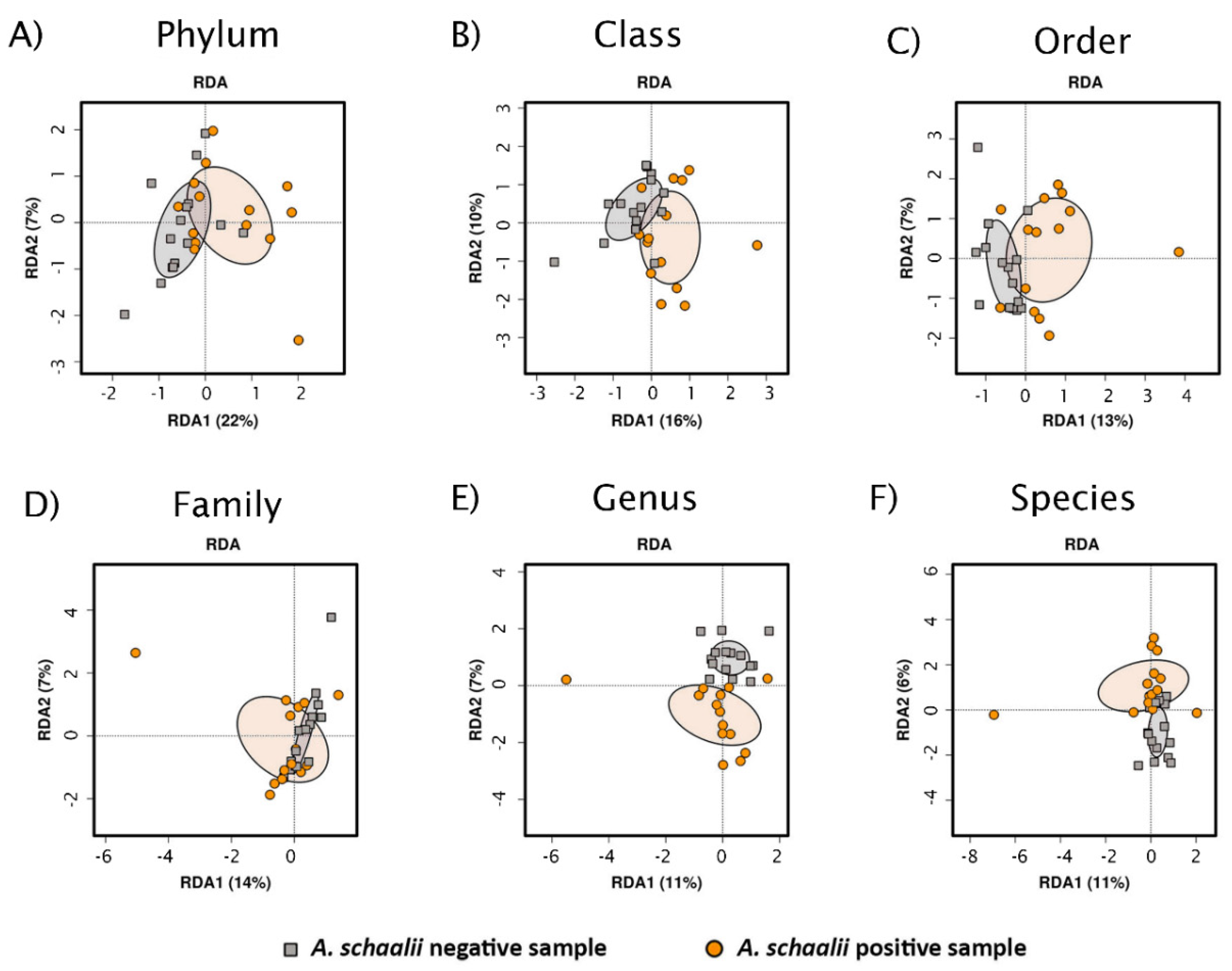
3.2.3. Beta Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lotte, L.; Lotte, R.; Durand, M.; Degand, N.; Ambrosetti, D.; Michiels, J.-F.; Amiel, J.; Cattoir, V.; Ruimy, R. Infections related to Actinotignum schaalii (formerly Actinobaculum schaalii): A 3-year prospective observational study on 50 cases. Clin. Microbiol. Infect. 2016, 22, 388–390. [Google Scholar] [CrossRef] [Green Version]
- Yassin, A.F.; Langenberg, S.; Huntemann, M.; Clum, A.; Pillay, M.; Palaniappan, K.; Varghese, N.; Mikhailova, N.; Mukherjee, S.; Reddy, T.B.K.; et al. Draft genome sequence of Actinotignum schaalii DSM 15541T: Genetic insights into the lifestyle, cell fitness and virulence. PLoS ONE 2017, 12, e0188914. [Google Scholar] [CrossRef] [Green Version]
- Olsen, A.B.; Andersen, P.K.; Bank, S.; Søby, K.M.; Lund, L.; Prag, J. Actinobaculum schaalii, a commensal of the urogenital area. BJU Int. 2013, 112, 394–397. [Google Scholar] [CrossRef]
- Lotte, R.; Lotte, L.; Ruimy, R. Actinotignum schaalii (formerly Actinobaculum schaalii): A newly recognized pathogen—Review of the literature. Clin. Microbiol. Infect. 2016, 22, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, M.; Kobayashi, T.; Suzuki, T.; Wakabayashi, Y.; Ohama, Y.; Maekawa, S.; Takahashi, S.; Homma, Y.; Tatsuno, K.; Sato, T.; et al. Propionimicrobium lymphophilum and Actinotignum schaalii bacteraemia: A case report. New Microbes New Infect. 2017, 18, 18–21. [Google Scholar] [CrossRef]
- Sturm, P.D.J.; Van Eijk, J.; Veltman, S.; Meuleman, E.; Schulin, T. Urosepsis with Actinobaculum schaalii and Aerococcus urinae. J. Clin. Microbiol. 2006, 44, 652–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanikolaou, A.; Brugger, J.; Sendi, P.; Olariu, R. An unusual clinical presentation of necrotizing fasciitis. Infection 2020, 48, 655–656. [Google Scholar] [CrossRef]
- Panganiban, C.M.; Gupta, S. Actinotignum schaalii Abscess in a Patient with Common Variable Immunodeficiency. Pathogens 2020, 9, 494. [Google Scholar] [CrossRef] [PubMed]
- Sandlund, J.; Glimåker, M.; Svahn, A.; Brauner, A. Bacteraemia caused by Actinobaculum schaalii: An overlooked pathogen? Scand. J. Infect. Dis. 2014, 46, 605–608. [Google Scholar] [CrossRef]
- Bryan, A.; Kirkpatrick, L.M.; Manaloor, J.J.; Salipante, S.J. 16S rRNA deep sequencing identifies Actinotignum schaalii as the major component of a polymicrobial intra-abdominal infection and implicates a urinary source. JMM Case Rep. 2017, 4, e005091. [Google Scholar] [CrossRef]
- Beguelin, C.; Genne, D.; Varca, A.; Tritten, M.-L.; Siegrist, H.H.; Jaton, K.; Lienhard, R. Actinobaculum schaalii: Clinical observation of 20 cases. Clin. Microbiol. Infect. 2011, 17, 1027–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschudin-Sutter, S.; Frei, R.; Weisser, M.; Goldenberger, D.; Widmer, A.F. Actinobaculum schaalii-invasive pathogen or innocent bystander? A retrospective observational study. BMC Infect. Dis. 2011, 11, 289. [Google Scholar] [CrossRef] [Green Version]
- Prigent, G.; Perillaud, C.; Amara, M.; Coutard, A.; Blanc, C.; Pangon, B. Actinobaculum schaalii: A truly emerging pathogen? Actinobaculum schaalii: Un pathogène réellement émergent? New Microbes New Infect. 2016, 11, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Bank, S.; Søby, K.M.; Kristensen, L.H.; Voldstedlund, M.; Prag, J. A validation of the Danish microbiology database (MiBa) and incidence rate of Actinotignum schaalii (Actinobaculum schaalii) bacteraemia in Denmark. Clin. Microbiol. Infect. 2015, 21, 1097.e1–1097.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.; Nurjadi, D.; Horner, S.; Heeg, K.; Zimmermann, S.; Burckhardt, I. Significant increase in cultivation of Gardnerella vaginalis, Alloscardovia omnicolens, Actinotignum schaalii, and Actinomyces spp. in urine samples with total laboratory automation. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1305–1311. [Google Scholar] [CrossRef] [Green Version]
- Barraud, O.; Ravry, C.; François, B.; Daix, T.; Ploy, M.-C.; Vignon, P. Shotgun metagenomics for microbiome and resistome detection in septic patients with urinary tract infection. Int. J. Antimicrob. Agents 2019, 54, 803–808. [Google Scholar] [CrossRef] [PubMed]
- Stevens, R.P.; Taylor, P.C. Actinotignum (formerly Actinobaculum) schaalii: A review of MALDI-TOF for identification of clinical isolates, and a proposed method for presumptive phenotypic identification. Pathology 2016, 48, 367–371. [Google Scholar] [CrossRef]
- Nielsen, H.L.; Søby, K.M.; Christensen, J.J.; Prag, J. Actinobaculum schaalii: A common cause of urinary tract infection in the elderly population. Bacteriological and clinical characteristics. Scand. J. Infect. Dis. 2009, 42, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Larios, O.E.; Bernard, K.A.; Manickam, K.; Ng, B.; Alfa, M.; Ronald, A. First report of Actinobaculum schaalii urinary tract infection in North America. Diagn. Microbiol. Infect. Dis. 2010, 67, 282–285. [Google Scholar] [CrossRef] [PubMed]
- Loiez, C.; Pilato, R.; Mambie, A.; Hendricx, S.; Faure, K.; Wallet, F. Native aortic endocarditis due to an unusual pathogen: Actinotignum schaalii. APMIS 2018, 126, 171–173. [Google Scholar] [CrossRef]
- Horton, L.E.; Mehta, S.R.; Aganovic, L.; Fierer, J. Actinotignum schaalii Infection: A Clandestine Cause of Sterile Pyuria? Open Forum Infect. Dis. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhard, M.; Prag, J.; Kemp, M.; Andresen, K.; Klemmensen, B.; Højlyng, N.; Sørensen, S.H.; Christensen, J.J. Ten Cases of Actinobaculum schaalii Infection: Clinical Relevance, Bacterial Identification, and Antibiotic Susceptibility. J. Clin. Microbiol. 2005, 43, 5305–5308. [Google Scholar] [CrossRef] [Green Version]
- Gomez, E.; Gustafson, D.R.; Rosenblatt, J.E.; Patel, R. Actinobaculum bacteremia: A Report of 12 Cases. J. Clin. Microbiol. 2011, 49, 4311–4313. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, E.; White, J.R.; Yu, S.-H.; Kulac, I.; Ertunc, O.; De Marzo, A.M.; Yegnasubramanian, S.; Mangold, L.A.; Partin, A.W.; Sfanos, K.S. Profiling the Urinary Microbiome in Men with Positive versus Negative Biopsies for Prostate Cancer. J. Urol. 2018, 199, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, S.A.; Razvi, H.; Dave, S.; Reid, G.; Burton, J.P. The microbiome of the urinary tract—A role beyond infection. Nat. Rev. Urol. 2015, 12, 81–90. [Google Scholar] [CrossRef]
- Barberis, C.; Cittadini, R.; Del Castillo, M.; Acevedo, P.; Roig, C.G.; Ramirez, M.S.; Perez, S.; Almuzara, M.; Vay, C. Actinobaculum schaalii causing urinary tract infections: Report of four cases from Argentina. J. Infect. Dev. Ctries. 2014, 8, 240–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popović, V.B.; Šitum, M.; Chow, C.-E.T.; Chan, L.S.; Roje, B.; Terzić, J. The urinary microbiome associated with bladder cancer. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pearce, M.M.; Hilt, E.E.; Rosenfeld, A.B.; Zilliox, M.J.; Thomas-White, K.; Fok, C.S.; Kliethermes, S.; Schreckenberger, P.C.; Brubaker, L.; Gai, X.; et al. The Female Urinary Microbiome: A Comparison of Women with and without Urgency Urinary Incontinence. mBio 2014, 5, e01283-14. [Google Scholar] [CrossRef] [Green Version]
- Bossa, L.; Kline, K.; McDougald, D.; Lee, B.B.; Rice, S.A. Urinary catheter-associated microbiota change in accordance with treatment and infection status. PLoS ONE 2017, 12, e0177633. [Google Scholar] [CrossRef] [Green Version]
- Kotaskova, I.; Obrucova, H.; Malisova, B.; Videnska, P.; Zwinsova, B.; Peroutkova, T.; Dvorackova, M.; Kumstat, P.; Trojan, P.; Ruzicka, F.; et al. Molecular Techniques Complement Culture-Based Assessment of Bacteria Composition in Mixed Biofilms of Urinary Tract Catheter-Related Samples. Front. Microbiol. 2019, 10, 462. [Google Scholar] [CrossRef] [PubMed]
- Kotásková, I.; Mališová, B.; Obručová, H.; Holá, V.; Peroutková, T.; Růžička, F.; Freiberger, T. Contribution of PCR Denaturing Gradient Gel Electrophoresis Combined with Mixed Chromatogram Software Separation for Complex Urinary Sample Analysis. Microb. Physiol. 2017, 27, 350–355. [Google Scholar] [CrossRef]
- Obručová, H.; Kotásková, I.; Tihelková, R.; Holá, V.; Růžička, F.; Freiberger, T. Fluorescent Capillary Electrophoresis Is Superior to Culture in Detecting Candida Species from Samples of Urinary Catheters and Ureteral Stents with Mono- or Polyfungal Biofilm Growth. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [Green Version]
- Leal, S.M.; Jones, M.; Gilligan, P.H. Clinical Significance of Commensal Gram-Positive Rods Routinely Isolated from Patient Samples. J. Clin. Microbiol. 2016, 54, 2928–2936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvadó, M.; Plasencia, V.; Segura, C.; Gómez, J.; Medina, M.J.; Sáez-Nieto, J.A.; Castellanos, S.; Horcajada, J.P. Infection due to Actinobaculum spp: Report of 12 patients in Spain. J. Infect. 2013, 66, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.; Senneby, E.; Rasmussen, M. Clinical and microbiological features of Actinotignum bacteremia: A retrospective observational study of 57 cases. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 36, 791–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bank, S.; Jensen, A.; Hansen, T.M.; Søby, K.M.; Prag, J. Actinobaculum schaalii, a Common Uropathogen in Elderly Patients, Denmark. Emerg. Infect. Dis. 2010, 16, 76–80. [Google Scholar] [CrossRef]
- Buhmann, M.T.; Abt, D.; Nolte, O.; Neu, T.R.; Strempel, S.; Albrich, W.C.; Betschart, P.; Zumstein, V.; Neels, A.; Maniura-Weber, K.; et al. Encrustations on ureteral stents from patients without urinary tract infection reveal distinct urotypes and a low bacterial load. Microbiome 2019, 7, 60. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2012, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Mrázek, J.; Štrosová, L.; Fliegerova, K.; Kott, T.; Kopečný, J. Diversity of insect intestinal microflora. Folia Microbiol. 2008, 53, 229–233. [Google Scholar] [CrossRef]
- Nadkarni, M.A.; Martin, F.E.; Jacques, N.A.; Hunter, N. Determination of bacterial load by real-time PCR using a broad-range (universal) probe and primers set. Microbiology 2002, 148, 257–266. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 20 January 2020).
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.-J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome–environment interactions. Bioinformatics 2016, 33, 782–783. [Google Scholar] [CrossRef] [Green Version]
- Hayes, B.W.; Abraham, S.N. Innate Immune Responses to Bladder Infection. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, A.C.; Goldstein, D.R.; Montgomery, R.R. Age-dependent dysregulation of innate immunity. Nat. Rev. Immunol. 2013, 13, 875–887. [Google Scholar] [CrossRef] [Green Version]
- Adebayo, A.; Ackermann, G.; Bowyer, R.; Wells, P.; Humphreys, G.; Knight, R.; Spector, T.; Steves, C. The Urinary Tract Microbiome in Older Women Exhibits Host Genetic and Environmental Influences. Cell Host Microbe 2020, 28, 298–305.e3. [Google Scholar] [CrossRef]
- Horwitz, D.; McCue, T.; Mapes, A.C.; Ajami, N.J.; Petrosino, J.F.; Ramig, R.F.; Trautner, B.W. Decreased microbiota diversity associated with urinary tract infection in a trial of bacterial interference. J. Infect. 2015, 71, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Oresta, B.; Braga, D.; Lazzeri, M.; Frego, N.; Saita, A.; Faccani, C.; Fasulo, V.; Colombo, P.; Guazzoni, G.; Hurle, R.; et al. The Microbiome of Catheter Collected Urine in Males with Bladder Cancer According to Disease Stage. J. Urol. 2021, 205, 86–93. [Google Scholar] [CrossRef]
- Imirzalioglu, C.; Hain, T.; Chakraborty, T.; Domann, E. Hidden pathogens uncovered: Metagenomic analysis of urinary tract infections. Andrologia 2008, 40, 66–71. [Google Scholar] [CrossRef]
- Williams, G.D. Two Cases of Urinary Tract Infection Caused by Propionimicrobium lymphophilum. J. Clin. Microbiol. 2015, 53, 3077–3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lainhart, W.; Gonzalez, M.D. Aerococcus urinae, Alloscardovia omnicolens, and Actinotignum schaalii: The AAA Minor League Team of Urinary Tract Infection Pathogens. Clin. Microbiol. Newsl. 2018, 40, 77–82. [Google Scholar] [CrossRef]
- Moustafa, A.; Li, W.; Singh, H.; Moncera, K.J.; Torralba, M.G.; Yu, Y.; Manuel, O.; Biggs, W.; Venter, J.C.; Nelson, K.E.; et al. Microbial metagenome of urinary tract infection. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tsitrin, T.; Singh, H.; Doerfert, S.N.; Sizova, M.V.; Epstein, S.S. Actinobaculum massiliense Proteome Profiled in Polymicrobial Urethral Catheter Biofilms. Proteomes 2018, 6, 52. [Google Scholar] [CrossRef] [Green Version]
- Brennan, C.A.; Garrett, W.S. Fusobacterium nucleatum—Symbiont, opportunist and oncobacterium. Nat. Rev. Genet. 2018, 17, 156–166. [Google Scholar] [CrossRef]
- He, X.; Hu, W.; Kaplan, C.W.; Guo, L.; Shi, W.; Lux, R. Adherence to Streptococci Facilitates Fusobacterium nucleatum Integration into an Oral Microbial Community. Microb. Ecol. 2011, 63, 532–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, C.W.; Lux, R.; Haake, S.K.; Shi, W. The Fusobacterium nucleatumouter membrane protein RadD is an arginine-inhibitable adhesin required for inter-species adherence and the structured architecture of multispecies biofilm. Mol. Microbiol. 2008, 71, 35–47. [Google Scholar] [CrossRef] [Green Version]
- De, A.; Raj, H.J.; Maiti, P.K. Biofilm in Osteomyelitis caused by a Rare Pathogen, Morganella morganii: A Case Report. J. Clin. Diagn. Res. 2016, 10, DD06–DD08. [Google Scholar] [CrossRef]
- Chen, Y.-T.; Peng, H.-L.; Shia, W.-C.; Hsu, F.-R.; Ken, C.-F.; Tsao, Y.-M.; Chen, C.-H.; Liu, C.-E.; Hsieh, M.-F.; Chen, H.-C.; et al. Whole-genome sequencing and identification of Morganella morganii KT pathogenicity-related genes. BMC Genom. 2012, 13, S4. [Google Scholar] [CrossRef] [Green Version]
- Periasamy, S.; Kolenbrander, P.E. Aggregatibacter actinomycetemcomitans Builds Mutualistic Biofilm Communities with Fusobacterium nucleatum and Veillonella Species in Saliva. Infect. Immun. 2009, 77, 3542–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, O.; Mörgelin, M.; Rasmussen, M. Platelet Activation and Biofilm Formation by Aerococcus urinae, an Endocarditis-Causing Pathogen. Infect. Immun. 2010, 78, 4268–4275. [Google Scholar] [CrossRef] [Green Version]
- Hilt, E.E.; Putonti, C.; Thomas-White, K.; Lewis, A.L.; Visick, K.L.; Gilbert, N.M.; Wolfe, A.J. Aerococcus urinae Isolated from Women with Lower Urinary Tract Symptoms: In Vitro Aggregation and Genome Analysis. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, S.; Kolenbrander, P.E.; Zeng, L.; Das, S.; Burne, R.A. Central Role of the Early Colonizer Veillonella sp. in Establishing Multispecies Biofilm Communities with Initial, Middle, and Late Colonizers of Enamel. J. Bacteriol. 2010, 192, 2965–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Toit, M.; Huch, M.; Cho, G.-S.; Franz, C.M. The genus Streptococcus. In Lactic Acid Bacteria; John Wiley & Sons: Chichester, UK, 2014; pp. 457–505. [Google Scholar] [CrossRef]
- Tomich, M.; Planet, P.J.; Figurski, D.H. The tad locus: Postcards from the widespread colonization island. Nat. Rev. Microbiol. 2007, 5, 363–375. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Urinary Catheter-Related (UC) | Double-J Catheter-Related (DJC) 1 | In Total (% of All Samples) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| A. schaalii Positive (% of Subtotal) | A. schaalii Negative (% of Subtotal) | Subtotal (% of All Samples) | p-Value 2 | A. schaalii Positive (% of Subtotal) | A. schaalii Negative (% of Subtotal) | Subtotal (% of All Samples) | p-Value 2 | ||
| Urine samples | 15 (23.9) | 48 (76.1) | 63 (46.7) | 0.340 | 10 (18.9) | 43 (81.1) | 53 (49.1) | 0.813 | 116 (47.8) |
| Catheter sonicates 1 | 23 (31.9) | 49 (68.1) | 72 (53.3) | 12 (21.8) | 43 (78.2) | 55 (50.9) | 127 (52.3) | ||
| All samples | 38 (28.1) | 97 (71.9) | 135 (100) | 22 (20.4) | 86 (79.6) | 108 (100) | 243 | ||
| A. schaalii Positive 1, n = 43, (% of Positive Patients) | A. schaalii Negative, n = 85, (% of Positive Patients) | In Total (% of All Patients) | p-Value 2 | |
|---|---|---|---|---|
| Sex | ||||
| Males (median age ± SD) | 31 (74.0 ± 15.4) (72.1) | 65 (75.0 ± 15.3) (76.5) | 96 (75 ± 15.3) (75.0) | 0.667 |
| Females (median age ± SD) | 12 (64.5 ± 14.6) (27.9) | 20 (66.5 ± 12.1) (23.5) | 32 (66 ± 13.1) (25.0) | |
| Type of Catheter in Patients | ||||
| Double-J catheter | 17 (39.5) | 38 (44.7) | 55 (43.0) | 0.706 |
| Urinary Catheter | 26 (60.5) | 47 (55.3) | 73 (57.0) | |
| Patients’ Diagnoses | Significance 2 | |||
| Renal colic | 5 (11.6) | 19 (22.4) | 24 (18.8) | n.s. |
| Hydronephrosis | 15 (34.9) | 28 (32.9) | 43 (33.6) | n.s. |
| Hydronephrosis with ureter stricture | 8 (18.6) | 4 (4.7) | 12 (9.4) | * |
| Urolithiasis | 13 (30.2) | 31 (36.5) | 44 (34.4) | n.s. |
| Prostatic or urinary tract cancer 3 | 10 (23.3) | 23 (27.1) | 33 (26.8) | n.s. |
| Bladder cancer | 2 (4.7) | 5 (5.9) | 7 (5.5) | n.s. |
| Prostate cancer 4 | 7 (22.6) | 13 (20) | 20 (15.6) | n.s. |
| Other | 2 (4.7) | 5 (9.4) | 7 (5.5) | n.s. |
| Other (none of mentioned above) | 16 (37.2) | 23 (27.1) | 39 (30.5) | n.s. |
| All Patients (% of 128 Patients) | 43 (33.6) | 85 (66.4) | 128 (100) |
| Concomitant Species 1 | A. schaalii Positive, n = 43, (% of Positive Patients) | A. schaalii Negative, n = 85, (% of Negative Patients) | In Total (% of All Patients) | Statistical Significance 2 |
|---|---|---|---|---|
| P. lymphophilum | 19 (44.2) | 10 (11.8) | 29 (22.7) | *** |
| F. nucleatum | 7 (16.3) | 3 (3.5) | 10 (7.8) | * |
| Streptococcus spp. | 9 (20.9) | 10 (11.8) | 19 (14.8) | n.s. |
| Alcaligenes faecalis | 3 (6.9) | 2 (2.4) | 5 (3.9) | n.s. |
| P. lymphophilum, F. nucleatum | 24 (55.8) | 11 (12.9) | 35 (27.3) | *** |
| P. lymphophilum, Streptococcus spp. | 26 (60.6) | 18 (21.2) | 44 (34.4) | *** |
| P. lymphophilum, A. faecalis | 21 (48.8) | 12 (14.1) | 33 (25.8) | *** |
| F. nucleatum, Streptococcus spp. | 12 (27.9) | 11 (12.9) | 23 (18.0) | n.s. |
| F. nucleatum, A. faecalis | 10 (23.3) | 5 (5.9) | 15 (11.8) | ** |
| Streptococcus spp., A. faecalis | 12 (27.9) | 12 (14.1) | 24 (18.7) | n.s. |
| P. lymphophilum, F. nucleatum, A. faecalis | 26 (60.5) | 13 (15.3) | 39 (30.5) | *** |
| P. lymphophilum, F. nucleatum, Streptococcus spp. | 27 (63.8) | 18 (21.2) | 45 (35.2) | *** |
| P. lymphophilum, A. faecalis, Streptococcus spp. | 28 (65.1) | 20 (23.5) | 48 (37.5) | *** |
| F. nucleatum, A. faecalis, Streptococcus spp. | 15 (34.9) | 13 (15.3) | 28 (21.9) | * |
| Any of indicator species | 29 (67.4) | 20 (23.5) | 49 (38.3) | *** |
| None of indicator species | 14 (32.6) | 65 (76.5) | 79 (61.7) | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotásková, I.; Syrovátka, V.; Obručová, H.; Vídeňská, P.; Zwinsová, B.; Holá, V.; Blaštíková, E.; Růžička, F.; Freiberger, T. Actinotignum schaalii: Relation to Concomitants and Connection to Patients’ Conditions in Polymicrobial Biofilms of Urinary Tract Catheters and Urines. Microorganisms 2021, 9, 669. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030669
Kotásková I, Syrovátka V, Obručová H, Vídeňská P, Zwinsová B, Holá V, Blaštíková E, Růžička F, Freiberger T. Actinotignum schaalii: Relation to Concomitants and Connection to Patients’ Conditions in Polymicrobial Biofilms of Urinary Tract Catheters and Urines. Microorganisms. 2021; 9(3):669. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030669
Chicago/Turabian StyleKotásková, Iva, Vít Syrovátka, Hana Obručová, Petra Vídeňská, Barbora Zwinsová, Veronika Holá, Eva Blaštíková, Filip Růžička, and Tomáš Freiberger. 2021. "Actinotignum schaalii: Relation to Concomitants and Connection to Patients’ Conditions in Polymicrobial Biofilms of Urinary Tract Catheters and Urines" Microorganisms 9, no. 3: 669. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9030669