
Biological Properties and Genetic Characterization of Novel Low Pathogenic H7N3 Avian Influenza Viruses Isolated from Mallard Ducks in the Caspian Region, Dagestan, Russia

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animal and Human Rights Statement
2.2. Sample Collection
2.3. Influenza Virus Detection and Isolation
2.4. Pathogenicity Tests in Chickens
2.5. Experimental Infection of Mice
2.6. Sequencing
2.7. Molecular Characterization
2.7.1. Amino Acid Analysis
2.7.2. Nucleotide Analysis
2.7.3. Phylogenetic Analyses
2.7.4. Maximum Likelihood Phylogenetic Analysis
2.7.5. Bayesian Phylogenetic Analysis
3. Results
3.1. Isolation and Pathogenicity of Two Novel H7N3 LPAI Strains
3.2. Nucleotide and Protein Analyses
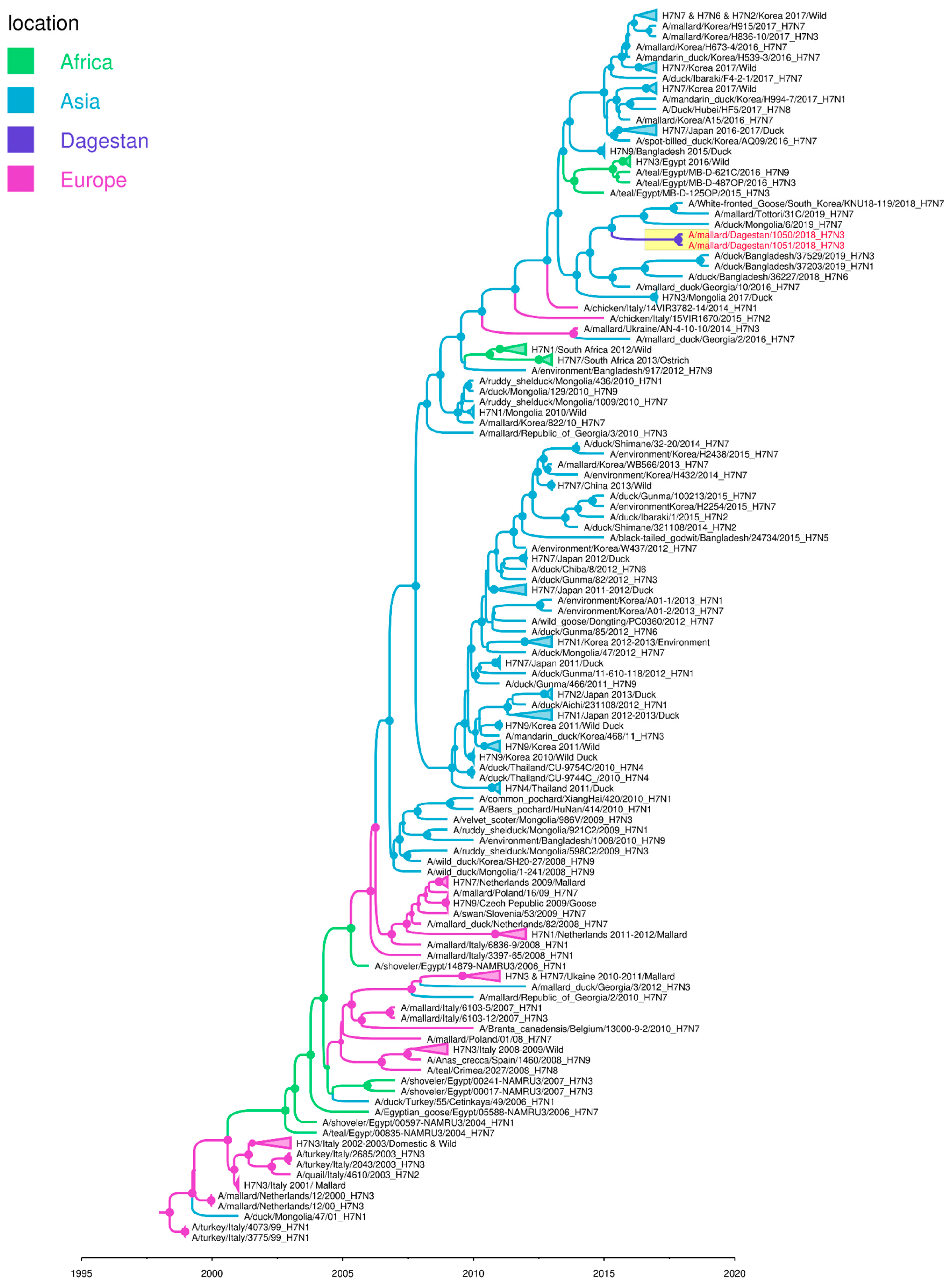
3.3. Phylogenetic Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| No | Segment | Best-Fit Model of Substitution According to BIC | Input Data |
|---|---|---|---|
| 1 | PB2 | GTR + F + G4 | 131 sequences with 2280 nt sites |
| 2 | PB1 | TIM + F + I + G4 | 136 sequences with 2274 nt sites |
| 3 | PA | TVM + F + G4 | 119 sequences with 2151 nt sites |
| 4 | HA | TVM + F + G4 | 104 sequences with 1683 nt sites |
| 5 | NP | K3Pu + F + R2 | 105 sequences with 1497 nt sites |
| 6 | NA | TIM + F + I + G4 | 116 sequences with 1410 nt sites |
| 7 | MP | TIMe + I + G4 | 119 sequences with 982 nt sites |
| 8 | NS | HKY + F + R2 | 112 sequences with 838 nt sites |

References
- Yoon, S.W.; Webby, R.J.; Webster, R.G. Evolution and ecology of influenza A viruses. Curr. Top. Microbiol. Immunol. 2014, 385, 359–375. [Google Scholar] [CrossRef] [Green Version]
- Olsen, B.; Munster, V.J.; Wallensten, A.; Waldenstrom, J.; Osterhaus, A.D.; Fouchier, R.A. Global patterns of influenza A virus in wild birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef] [Green Version]
- Latorre-Margalef, N.; Gunnarsson, G.; Munster, V.J.; Fouchier, R.A.; Osterhaus, A.D.; Elmberg, J.; Olsen, B.; Wallensten, A.; Haemig, P.D.; Fransson, T.; et al. Effects of influenza A virus infection on migrating mallard ducks. Proc. Biol. Sci. 2009, 276, 1029–1036. [Google Scholar] [CrossRef] [Green Version]
- Verhagen, J.H.; van Dijk, J.G.; Vuong, O.; Bestebroer, T.; Lexmond, P.; Klaassen, M.; Fouchier, R.A. Migratory birds reinforce local circulation of avian influenza viruses. PLoS ONE 2014. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, D.; Poen, M.J.; Bestebroer, T.M.; Scheuer, R.D.; Vuong, O.; Chkhaidze, M.; Machablishvili, A.; Mamuchadze, J.; Ninua, L.; Fedorova, N.B.; et al. Avian Influenza Viruses in Wild Birds: Virus Evolution in a Multihost Ecosystem. J. Virol. 2018, 92, 15. [Google Scholar] [CrossRef] [Green Version]
- Hill, N.J.; Ma, E.J.; Meixell, B.W.; Lindberg, M.S.; Boyce, W.M.; Runstadler, J.A. Transmission of influenza reflects seasonality of wild birds across the annual cycle. Ecol. Lett. 2016, 19, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Lycett, S.J.; Duchatel, F.; Digard, P. A brief history of bird flu. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374. [Google Scholar] [CrossRef] [PubMed]
- Naguib, M.M.; Verhagen, J.H.; Mostafa, A.; Wille, M.; Li, R.; Graaf, A.; Järhult, J.D.; Ellström, P.; Zohari, S.; Lundkvist, Å.; et al. Global patterns of avian influenza A(H7): Virus evolution and zoonotic threats. FEMS Microbiol. Rev. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO [Food and Agriculture Administration of the United Nations]. H7N9 Situation Update. 2019. Available online: http://www.fao.org/ag/againfo/programmes/en/empres/h7n9/situation_update.html] (accessed on 16 April 2021).
- Dhingra, M.S.; Artois, J.; Dellicour, S.; Lemey, P.; Dauphin, G.; Von Dobschuetz, S.; Van Boeckel, T.P.; Castellan, D.M.; Morzaria, S.; Gilbert, M. Geographical and Historical Patterns in the Emergences of Novel Highly Pathogenic Avian Influenza (HPAI) H5 and H7 Viruses in Poultry. Front. Vet. Sci. 2018, 5, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, M.; Uchida, Y.; Shibata, A.; Kobayashi, Y.; Mine, J.; Takemae, N.; Tsunekuni, R.; Tanikawa, T.; Harada, R.; Osaka, H.; et al. A novel H7N3 reassortant originating from the zoonotic H7N9 highly pathogenic avian influenza viruses that has adapted to ducks. Transbound. Emerg. Dis. 2019, 66, 2342–2352. [Google Scholar] [CrossRef]
- Danilenko, E.A.; Soldatov, M.S. Bird Migration in the Caspian Region. 2015. Available online: www.geogr.msu.ru/casp (accessed on 16 April 2021).
- Gulyaeva, M.; Sobolev, I.; Sharshov, K.; Kurskaya, O.; Alekseev, A.; Shestopalova, L.; Kovner, A.; Bi, Y.; Shi, W.; Shchelkanov, M.; et al. Characterization of Avian-like Influenza A (H4N6) Virus Isolated from Caspian Seal in 2012. Virol. Sin. 2018, 33, 449–452. [Google Scholar] [CrossRef]
- Vilkov, E.V. The Genesis and Evolution of the Caspian Sea Lagoons as Avifauna Refuges at the Transboundary Scale. Open J. Mar. Sci. 2016, 6, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Suttie, A.; Yann, S.; Tum, S.; Deng, Y.M.; Hul, V.; Horm, V.S.; Barr, I.; Greenhill, A.; Horwood, P.F.; Osbjer, K.; et al. Detection of Low Pathogenicity Influenza A(H7N3) Virus during Duck Mortality Event, Cambodia, 2017. Emerg. Infect Dis. 2018, 24, 1103–1107. [Google Scholar] [CrossRef] [Green Version]
- Swayne, D.E.; Glisson, J.R.; Jackwood, M.W.; Pearson, J.E.; Reed, W.M. A Laboratory Manual for the Isolation and Identification of Avian Pathogens, 4th ed.; Am. Assoc. Avian Pathol: Jacksonville, FL, USA, 1998; pp. 74–80. [Google Scholar]
- OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 2018. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.03.04_AI.pdf (accessed on 16 April 2021).
- Mo, I.P.; Brugh, M.; Fletcher, O.J.; Rowland, G.N.; Swayne, D.E. Comparative pathology of chickens experimentally inoculated with avian influenza viruses of low and high pathogenicity. Avian Dis. 1997, 41, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Suttie, A.; Deng, Y.M.; Greenhill, A.R.; Dussart, P.; Horwood, P.F.; Karlsson, E.A. Inventory of molecular markers affecting biological characteristics of avian influenza A viruses. Virus Genes 2019, 55, 739–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mänz, B.; Schwemmle, M.; Brunotte, L. Adaptation of avian influenza A virus polymerase in mammals to overcome the host species barrier. J. Virol. 2013, 87, 7200–7209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Song, M.S.; Marathe, B.M.; Kumar, G.; Wong, S.S.; Rubrum, R.; Zanin, M.; Choi, Y.K.; Webster, R.G.; Govorkova, E.A.; Webby, R.J. Unique Determinants of Neuraminidase Inhibitor Resistance among N3, N7, and N9 Avian Influenza Viruses. J. Virol. 2015, 89, 10891–10900. [Google Scholar] [CrossRef] [Green Version]
- Gubareva, L.V.; Hayden, F.G. M2 and neuraminidase inhibitors: Anti-influenza activity, mechanisms of resistance, and clinical effectiveness. In Influenza Virology: Current Topics; Kawaoka, Y., Ed.; Caister Academic Press: Norwich, UK, 2006; pp. 169–202. [Google Scholar]
- Kazutaka, K.; Standley, K. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar]
- Rambaut, A.; Lam, T.T.; Carvalho, L.M.; Pybus, O.G. Exploring the temporal structure of heterochronous sequences using TempEst. Virus Evol. 2016, 2. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, N.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Systematic. Biology 2018. [Google Scholar] [CrossRef] [Green Version]
- Banks, J.; Speidel, E.S.; Moore, E.; Plowright, L.; Piccirillo, A.; Capua, I.; Cordioli, P.; Fioretti, A.; Alexander, D.J. Changes in the haemagglutinin and the neuraminidase genes prior to the emergence of highly pathogenic H7N1 avian influenza viruses in Italy. Arch Virol. 2001, 146, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Campitelli, L.; Mogavero, E.; De Marco, M.A.; Delogu, M.; Puzelli, S.; Frezza, F.; Facchini, M.; Chiapponi, C.; Foni, E. Interspecies transmission of an H7N3 influenza virus from wild birds to intensively reared domestic poultry in Italy. Virology 2004, 323, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Holmes, E.C. Avian Influenza Virus Exhibits Rapid Evolutionary Dynamics. Mol. Biol. Evol. 2006, 23, 2336–2341. [Google Scholar] [CrossRef]
- Abdelwhab, E.M.; Veits, J.; Mettenleiter, T.C. Prevalence and control of H7 avian influenza viruses in birds and humans. Epidemiol. Infect. 2014, 142, 896–920. [Google Scholar] [CrossRef] [PubMed]
- Hiono, T.; Ohkawara, A.; Ogasawara, K.; Okamatsu, M.; Tamura, T.; Chu, D.H.; Suzuki, M.; Kuribayashi, S.; Shichinohe, S.; Takada, A.; et al. Genetic and antigenic characterization of H5 and H7 influenza viruses isolated from migratory water birds in Hokkaido, Japan and Mongolia from 2010 to 2014. Virus Genes 2015, 51, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Youk, S.; Lee, D.H.; Killian, M.L.; Pantin-Jackwood, M.J.; Swayne, D.E.; Torchetti, M.K. Highly Pathogenic Avian Influenza A(H7N3) Virus in Poultry, United States, 2020. Emerg. Infect. Dis. 2020, 26, 2966–2969. [Google Scholar] [CrossRef] [PubMed]
- Muzyka, D.; Pantin-Jackwood, M.; Spackman, E.; Smith, D.; Rula, O.; Muzyka, N.; Stegniy, B. Isolation and Genetic Characterization of Avian Influenza Viruses Isolated from Wild Birds in the Azov-Black Sea Region of Ukraine (2001–2012). Avian Dis. 2016, 60, 365–377. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gulyaeva, M.; De Marco, M.A.; Kovalenko, G.; Bortz, E.; Murashkina, T.; Yurchenko, K.; Facchini, M.; Delogu, M.; Sobolev, I.; Gadzhiev, A.; et al. Biological Properties and Genetic Characterization of Novel Low Pathogenic H7N3 Avian Influenza Viruses Isolated from Mallard Ducks in the Caspian Region, Dagestan, Russia. Microorganisms 2021, 9, 864. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040864
Gulyaeva M, De Marco MA, Kovalenko G, Bortz E, Murashkina T, Yurchenko K, Facchini M, Delogu M, Sobolev I, Gadzhiev A, et al. Biological Properties and Genetic Characterization of Novel Low Pathogenic H7N3 Avian Influenza Viruses Isolated from Mallard Ducks in the Caspian Region, Dagestan, Russia. Microorganisms. 2021; 9(4):864. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040864
Chicago/Turabian StyleGulyaeva, Marina, Maria Alessandra De Marco, Ganna Kovalenko, Eric Bortz, Tatiana Murashkina, Kseniya Yurchenko, Marzia Facchini, Mauro Delogu, Ivan Sobolev, Alimurad Gadzhiev, and et al. 2021. "Biological Properties and Genetic Characterization of Novel Low Pathogenic H7N3 Avian Influenza Viruses Isolated from Mallard Ducks in the Caspian Region, Dagestan, Russia" Microorganisms 9, no. 4: 864. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040864