
Current Status of Putative Animal Sources of SARS-CoV-2 Infection in Humans: Wildlife, Domestic Animals and Pets

 ,
,  ,
,
Abstract
:1. Introduction
2. Potentially Favourable Conditions for the Emergence of SARS-CoV-2
2.1. Viral Genetic Variation
2.2. Interactions of Viral Spike with ACE2 and Other Possible Cell Receptors
2.3. Host Body Temperature
2.4. Human and Animal Population Density
2.5. Group-Living and Grooming Habits
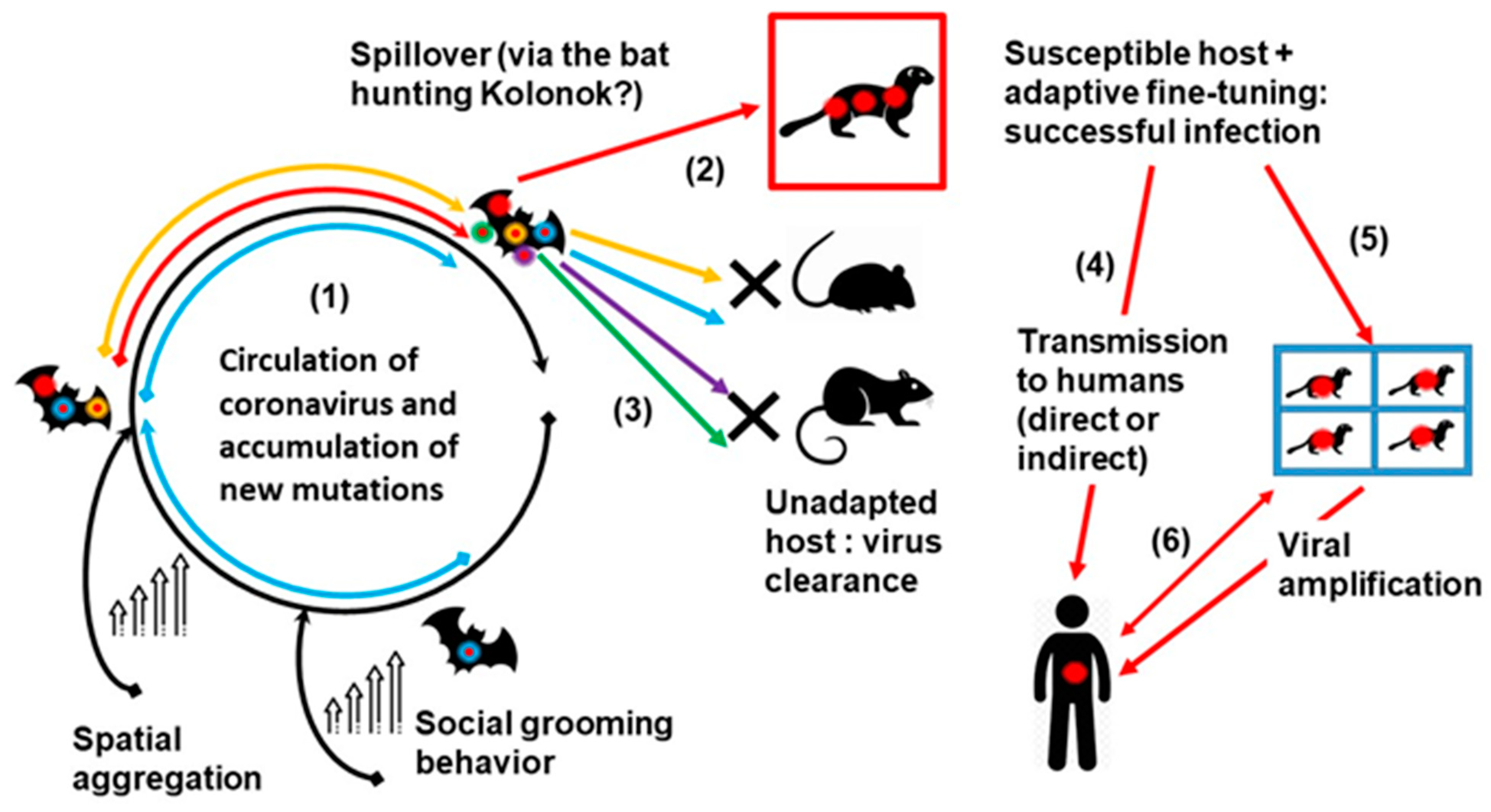
2.6. The Spillover versus Circulation Model
3. Experimental Models for SARS-CoV-2 Infection
3.1. Non-Human Primates
3.1.1. Callithrix Jacchus versus Macaca
3.1.2. Macaca Mullata (Rhesus Macaque)
3.1.3. African Green Monkey (Chlorocebus Sabaeus)
3.2. Bats
3.3. Pangolins
3.4. Dogs
3.5. Cats
3.6. Rabbits
3.7. Mink and Ferrets
3.8. Mice
3.9. Hamsters
3.10. Pigs
3.11. Tree Shrews
3.12. Poultry
4. Animal Species Susceptible to SARS-CoV-2 Infection, Viral Replication and Viral Spread
4.1. Domestic Animals
4.1.1. Pets
4.1.2. Other Domestic Animals
4.2. Captive Wild Animals in Farms
4.3. Captive wild Animals in Zoos
4.4. Non-Captive Wild Animals
4.5. World Organisation for Animal Health (OIE) and SARS-CoV-2 in Animals
5. Potential Interspecies Transmission of SARS-CoV-2
5.1. Transmission between Animals
5.2. Transmission between Humans and Domestic, Farm, or Zoo Animals
5.3. Transmission between Humans and Wild Animals
6. Spatial Aggregation of Susceptible Hosts Increases the Risk of SARS-CoV-2 Variant Selection
7. What Remains Unexplored
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdel-Moneim, A.S.; Abdelwhab, E.M. Evidence for SARS-CoV-2 Infection of Animal Hosts. Pathogens 2020, 9, 529. [Google Scholar] [CrossRef]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The Species Severe Acute Respiratory Syndrome-Related Coronavirus: Classifying 2019-NCoV and Naming It SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Molecular Evolution of Human Coronavirus Genomes. Trends Microbiol. 2017, 25, 35–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afelt, A.; Frutos, R.; Devaux, C. Bats, Coronaviruses, and Deforestation: Toward the Emergence of Novel Infectious Diseases? Front. Microbiol. 2018, 9, 702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, A.; Li, X.; Lau, S.; Woo, P. Global Epidemiology of Bat Coronaviruses. Viruses 2019, 11, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, D.L. SARS-CoV-2 and Bats: From Flight to Fighting COVID-19. JACC Basic Transl. Sci. 2020, 5, 545–546. [Google Scholar] [CrossRef]
- Frutos, R.; Serra-Cobo, J.; Pinault, L.; Lopez-Roig, M.; Devaux, C. Emergence of Bat-Related Betacoronaviruses: Hazard and Risks. Front. Microbiol. 2021, 12, 437. [Google Scholar] [CrossRef]
- Corman, V.M.; Muth, D.; Niemeyer, D.; Drosten, C. Hosts and Sources of Endemic Human Coronaviruses. Adv. Virus Res. 2018, 100, 163–188. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Liu, W.; Gan, M.; Zhang, L.; Wang, J.; Zhang, Z.; Zhu, A.; Li, F.; Sun, J.; et al. Discovery of a Subgenotype of Human Coronavirus NL63 Associated with Severe Lower Respiratory Tract Infection in China, 2018. Emerg. Microbes Infect. 2020, 9, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and Evolution of Pathogenic Coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Corman, V.M.; Eckerle, I.; Memish, Z.A.; Liljander, A.M.; Dijkman, R.; Jonsdottir, H.; Juma Ngeiywa, K.J.Z.; Kamau, E.; Younan, M.; Al Masri, M.; et al. Link of a Ubiquitous Human Coronavirus to Dromedary Camels. Proc. Natl. Acad. Sci. USA 2016, 113, 9864–9869. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19—11 March 2020. 2020. Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 2 February 2021).
- AlSamman, M.; Caggiula, A.; Ganguli, S.; Misak, M.; Pourmand, A. Non-Respiratory Presentations of COVID-19, a Clinical Review. Am. J. Emerg. Med. 2020, 38, 2444–2454. [Google Scholar] [CrossRef]
- Kordzadeh-Kermani, E.; Khalili, H.; Karimzadeh, I. Pathogenesis, Clinical Manifestations and Complications of Coronavirus Disease 2019 (COVID-19). Future Microbiol. 2020, 15, 1287–1305. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Frutos, R.; Lopez Roig, M.; Serra-Cobo, J.; Devaux, C.A. COVID-19: The Conjunction of Events Leading to the Coronavirus Pandemic and Lessons to Learn for Future Threats. Front. Med. 2020, 7, 223. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Luk, H.K.H.; Wong, A.C.P.; Li, K.S.M.; Zhu, L.; He, Z.; Fung, J.; Chan, T.T.Y.; Fung, K.S.C.; Woo, P.C.Y. Possible Bat Origin of Severe Acute Respiratory Syndrome Coronavirus 2. Emerg. Infect. Dis. 2020, 26, 1542–1547. [Google Scholar] [CrossRef]
- Yuen, K.-S.; Ye, Z.-W.; Fung, S.-Y.; Chan, C.-P.; Jin, D.-Y. SARS-CoV-2 and COVID-19: The Most Important Research Questions. Cell Biosci. 2020, 10, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, T.T.-Y.; Jia, N.; Zhang, Y.-W.; Shum, M.H.-H.; Jiang, J.-F.; Zhu, H.-C.; Tong, Y.-G.; Shi, Y.-X.; Ni, X.-B.; Liao, Y.-S.; et al. Identifying SARS-CoV-2-Related Coronaviruses in Malayan Pangolins. Nature 2020, 583, 282–285. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.; Zhai, J.; Feng, Y.; Zhou, N.; Zhang, X.; Zou, J.-J.; Li, N.; Guo, Y.; Li, X.; Shen, X.; et al. Isolation of SARS-CoV-2-Related Coronavirus from Malayan Pangolins. Nature 2020, 583, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The Proximal Origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef] [Green Version]
- Frutos, R.; Serra-Cobo, J.; Chen, T.; Devaux, C.A. COVID-19: Time to Exonerate the Pangolin from the Transmission of SARS-CoV-2 to Humans. Infect. Genet. Evol. 2020, 84, 104493. [Google Scholar] [CrossRef]
- Snijder, E.J.; Decroly, E.; Ziebuhr, J. The Nonstructural Proteins Directing Coronavirus RNA Synthesis and Processing. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2016; Volume 96, pp. 59–126. ISBN 978-0-12-804736-1. [Google Scholar]
- Naqvi, A.A.T.; Fatima, K.; Mohammad, T.; Fatima, U.; Singh, I.K.; Singh, A.; Atif, S.M.; Hariprasad, G.; Hasan, G.M.; Hassan, M.I. Insights into SARS-CoV-2 Genome, Structure, Evolution, Pathogenesis and Therapies: Structural Genomics Approach. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165878. [Google Scholar] [CrossRef]
- Chang, T.-J.; Yang, D.-M.; Wang, M.-L.; Liang, K.-H.; Tsai, P.-H.; Chiou, S.-H.; Lin, T.-H.; Wang, C.-T. Genomic Analysis and Comparative Multiple Sequences of SARS-CoV2. J. Chin. Med. Assoc. 2020, 83, 537–543. [Google Scholar] [CrossRef]
- Koma, T.; Adachi, S.; Doi, N.; Adachi, A.; Nomaguchi, M. Toward Understanding Molecular Bases for Biological Diversification of Human Coronaviruses: Present Status and Future Perspectives. Front. Microbiol. 2020, 11, 2016. [Google Scholar] [CrossRef]
- Flores-Alanis, A.; Sandner-Miranda, L.; Delgado, G.; Cravioto, A.; Morales-Espinosa, R. The Receptor Binding Domain of SARS-CoV-2 Spike Protein Is the Result of an Ancestral Recombination between the Bat-CoV RaTG13 and the Pangolin-CoV MP789. BMC Res. Notes 2020, 13, 398. [Google Scholar] [CrossRef] [PubMed]
- Coutard, B.; Valle, C.; de Lamballerie, X.; Canard, B.; Seidah, N.G.; Decroly, E. The Spike Glycoprotein of the New Coronavirus 2019-NCoV Contains a Furin-like Cleavage Site Absent in CoV of the Same Clade. Antivir. Res. 2020, 176, 104742. [Google Scholar] [CrossRef] [PubMed]
- Frutos, R.; Gavotte, L.; Devaux, C.A. Understanding the Origin of COVID-19 Requires to Change the Paradigm on Zoonotic Emergence from the Spillover Model to the Viral Circulation Model. Infect. Genet. Evol. 2021, 104812. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.; Bosch, B.; Li, F.; Li, W.; Lee, K.H.; Ghiran, S.; Vasilieva, N.; Dermody, T.S.; Harrison, S.C.; Dormitzer, P.R.; et al. SARS Coronavirus, but Not Human Coronavirus NL63, Utilizes Cathepsin L to Infect ACE2-Expressing Cells. J. Biol. Chem. 2006, 281, 3198–3203. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Liu, D.X. Proteolytic Activation of the Spike Protein at a Novel RRRR/S Motif Is Implicated in Furin-Dependent Entry, Syncytium Formation, and Infectivity of Coronavirus Infectious Bronchitis Virus in Cultured Cells. J. Virol. 2009, 83, 8744–8758. [Google Scholar] [CrossRef] [Green Version]
- Wacharapluesadee, S.; Tan, C.W.; Maneeorn, P.; Duengkae, P.; Zhu, F.; Joyjinda, Y.; Kaewpom, T.; Chia, W.N.; Ampoot, W.; Lim, B.L.; et al. Evidence for SARS-CoV-2 Related Coronaviruses Circulating in Bats and Pangolins in Southeast Asia. Nat. Commun. 2021, 12, 972. [Google Scholar] [CrossRef]
- Wrobel, A.G.; Benton, D.J.; Xu, P.; Calder, L.J.; Borg, A.; Roustan, C.; Martin, S.R.; Rosenthal, P.B.; Skehel, J.J.; Gamblin, S.J. Structure and Binding Properties of Pangolin-CoV Spike Glycoprotein Inform the Evolution of SARS-CoV-2. Nat. Commun. 2021, 12, 837. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Z.; Wang, F.-S. SARS-Associated Coronavirus Quasispecies in Individual Patients. N. Engl. J. Med. 2004, 350, 1366–1367. [Google Scholar] [CrossRef]
- Park, D.; Huh, H.J.; Kim, Y.J.; Son, D.-S.; Jeon, H.-J.; Im, E.-H.; Kim, J.-W.; Lee, N.Y.; Kang, E.-S.; Kang, C.I.; et al. Analysis of Intrapatient Heterogeneity Uncovers the Microevolution of Middle East Respiratory Syndrome Coronavirus. Cold Spring Harb. Mol. Case Stud. 2016, 2, a001214. [Google Scholar] [CrossRef] [Green Version]
- Karamitros, T.; Papadopoulou, G.; Bousali, M.; Mexias, A.; Tsiodras, S.; Mentis, A. SARS-CoV-2 Exhibits Intra-Host Genomic Plasticity and Low-Frequency Polymorphic Quasispecies. J. Clin. Virol. 2020, 131, 104585. [Google Scholar] [CrossRef]
- Chaudhry, M.Z.; Eschke, K.; Grashoff, M.; Abassi, L.; Kim, Y.; Brunotte, L.; Ludwig, S.; Šafranko, Ž.M.; Kurolt, I.-C.; Markotić, A.; et al. SARS-CoV-2 Quasispecies Mediate Rapid Virus Evolution and Adaptation. bioRxiv 2020. [Google Scholar] [CrossRef]
- Luan, J.; Lu, Y.; Jin, X.; Zhang, L. Spike Protein Recognition of Mammalian ACE2 Predicts the Host Range and an Optimized ACE2 for SARS-CoV-2 Infection. Biochem. Biophys. Res. Commun. 2020, 526, 165–169. [Google Scholar] [CrossRef]
- Luan, J.; Jin, X.; Lu, Y.; Zhang, L. SARS-CoV-2 Spike Protein Favors ACE2 from Bovidae and Cricetidae. J. Med. Virol. 2020, 92, 1649–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damas, J.; Hughes, G.M.; Keough, K.C.; Painter, C.A.; Persky, N.S.; Corbo, M.; Hiller, M.; Koepfli, K.-P.; Pfenning, A.R.; Zhao, H.; et al. Broad Host Range of SARS-CoV-2 Predicted by Comparative and Structural Analysis of ACE2 in Vertebrates. Proc. Natl. Acad. Sci. USA 2020, 117, 22311–22322. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Hernandez-Allas, X.; Cianferoni, D.; Serrano, L. In Silico Mutagenesis of Human ACE2 with S Protein and Translational Efficiency Explain SARS-CoV-2 Infectivity in Different Species. PLoS Comput. Biol. 2020, 16, e1008450. [Google Scholar]
- Qiu, Y.; Zhao, Y.-B.; Wang, Q.; Li, J.-Y.; Zhou, Z.-J.; Liao, C.-H.; Ge, X.-Y. Predicting the Angiotensin Converting Enzyme 2 (ACE2) Utilizing Capability as the Receptor of SARS-CoV-2. Microbes Infect. 2020, 22, 221–225. [Google Scholar] [CrossRef]
- Lam, S.D.; Bordin, N.; Waman, V.P.; Scholes, H.M.; Ashford, P.; Sen, N.; van Dorp, L.; Rauer, C.; Dawson, N.L.; Pang, C.S.M.; et al. SARS-CoV-2 Spike Protein Predicted to Form Complexes with Host Receptor Protein Orthologues from a Broad Range of Mammals. Sci. Rep. 2020, 10, 16471. [Google Scholar] [CrossRef]
- Wang, L.; Mitchell, P.K.; Calle, P.P.; Bartlett, S.L.; McAloose, D.; Killian, M.L.; Yuan, F.; Fang, Y.; Goodman, L.B.; Fredrickson, R.; et al. Complete Genome Sequence of SARS-CoV-2 in a Tiger from a U.S. Zoological Collection. Microbiol. Resour. Announc. 2020, 9. [Google Scholar] [CrossRef]
- Devaux, C.; Pinault, L.; Omar Osman, I.; Raoult, D. Can ACE2 Receptor Polymorphism Predict Species Susceptibility to SARS-CoV-2? Front. Public Health 2020, 8, 608765. [Google Scholar] [CrossRef]
- Bouricha, E.M.; Hakmi, M.; Akachar, J.; Belyamani, L.; Ibrahimi, A. In Silico Analysis of ACE2 Orthologues to Predict Animal Host Range with High Susceptibility to SARS-CoV-2. 3 Biotech 2020, 10, 483. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, G.; Wang, Y.; Zhao, X.; Ji, F.; Ren, W.; Gong, M.; Ju, X.; Zhu, Y.; Cai, X.; et al. Functional and Genetic Analysis of Viral Receptor ACE2 Orthologs Reveals a Broad Potential Host Range of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2021. [Google Scholar] [CrossRef]
- Chu, H.; Chan, J.F.-W.; Yuen, T.T.-T.; Shuai, H.; Yuan, S.; Wang, Y.; Hu, B.; Yip, C.C.-Y.; Tsang, J.O.-L.; Huang, X.; et al. Comparative Tropism, Replication Kinetics, and Cell Damage Profiling of SARS-CoV-2 and SARS-CoV with Implications for Clinical Manifestations, Transmissibility, and Laboratory Studies of COVID-19: An Observational Study. Lancet Microbe 2020, 1, e14–e23. [Google Scholar] [CrossRef]
- Guan, W.-J.; Ni, Z.-Y.; Hu, Y.; Liang, W.-H.; Ou, C.-Q.; He, J.-X.; Liu, L.; Shan, H.; Lei, C.-L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Munster, V.J.; Feldmann, F.; Williamson, B.N.; van Doremalen, N.; Pérez-Pérez, L.; Schulz, J.; Meade-White, K.; Okumura, A.; Callison, J.; Brumbaugh, B.; et al. Respiratory Disease in Rhesus Macaques Inoculated with SARS-CoV-2. Nature 2020, 585, 268–272. [Google Scholar] [CrossRef]
- Rockx, B.; Kuiken, T.; Herfst, S.; Bestebroer, T.; Lamers, M.M.; Oude Munnink, B.B.; de Meulder, D.; van Amerongen, G.; van den Brand, J.; Okba, N.M.A.; et al. Comparative Pathogenesis of COVID-19, MERS, and SARS in a Nonhuman Primate Model. Science 2020, 368, 1012–1015. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-I.; Kim, S.-G.; Kim, S.-M.; Kim, E.-H.; Park, S.-J.; Yu, K.-M.; Chang, J.-H.; Kim, E.J.; Lee, S.; Casel, M.A.B.; et al. Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe 2020, 27, 704–709.e2. [Google Scholar] [CrossRef]
- Richard, M.; Kok, A.; de Meulder, D.; Bestebroer, T.M.; Lamers, M.M.; Okba, N.M.A.; Fentener van Vlissingen, M.; Rockx, B.; Haagmans, B.L.; Koopmans, M.P.G.; et al. SARS-CoV-2 Is Transmitted via Contact and via the Air between Ferrets. Nat. Commun. 2020, 11, 3496. [Google Scholar] [CrossRef] [PubMed]
- Schlottau, K.; Rissmann, M.; Graaf, A.; Schön, J.; Sehl, J.; Wylezich, C.; Höper, D.; Mettenleiter, T.C.; Balkema-Buschmann, A.; Harder, T.; et al. SARS-CoV-2 in Fruit Bats, Ferrets, Pigs, and Chickens: An Experimental Transmission Study. Lancet Microbe 2020, 1, e218–e225. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of Ferrets, Cats, Dogs, and Other Domesticated Animals to SARS-Coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beer, M. COVID-19: Experimental Infection of Fruit Bats, Ferrets, Pigs, and Chicken with SARS-CoV-2. ProMed International Society for Infectious Diseases. Update, 2020-04-07 2020. Available online: https://scholar.harvard.edu/files/kleelerner/files/20200407_promed_-_coronavirus_disease_2019_update_88_germany_animals_research_pig_chicken_bat_ferret_.pdf (accessed on 5 February 2021).
- Molenaar, R.J.; Vreman, S.; Hakze-van der Honing, R.W.; Zwart, R.; de Rond, J.; Weesendorp, E.; Smit, L.A.M.; Koopmans, M.; Bouwstra, R.; Stegeman, A.; et al. Clinical and Pathological Findings in SARS-CoV-2 Disease Outbreaks in Farmed Mink (Neovison Vison). Vet. Pathol. 2020, 57, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Oreshkova, N.; Molenaar, R.J.; Vreman, S.; Harders, F.; Oude Munnink, B.B.; Hakze-van der Honing, R.W.; Gerhards, N.; Tolsma, P.; Bouwstra, R.; Sikkema, R.S.; et al. SARS-CoV-2 Infection in Farmed Minks, The Netherlands, April and May 2020. Eurosurveillance 2020, 25. [Google Scholar] [CrossRef]
- Oude Munnink, B.B.; Sikkema, R.S.; Nieuwenhuijse, D.F.; Molenaar, R.J.; Munger, E.; Molenkamp, R.; van der Spek, A.; Tolsma, P.; Rietveld, A.; Brouwer, M.; et al. Transmission of SARS-CoV-2 on Mink Farms between Humans and Mink and Back to Humans. Science 2021, 371, 172–177. [Google Scholar] [CrossRef]
- Freuling, C.M.; Breithaupt, A.; Müller, T.; Sehl, J.; Balkema-Buschmann, A.; Rissmann, M.; Klein, A.; Wylezich, C.; Höper, D.; Wernike, K.; et al. Susceptibility of Raccoon Dogs for Experimental SARS-CoV-2 Infection. Emerg. Infect. Dis. 2020, 26, 2982–2985. [Google Scholar] [CrossRef]
- McAloose, D.; Laverack, M.; Wang, L.; Killian, M.L.; Caserta, L.C.; Yuan, F.; Mitchell, P.K.; Queen, K.; Mauldin, M.R.; Cronk, B.D.; et al. From People to Panthera: Natural SARS-CoV-2 Infection in Tigers and Lions at the Bronx Zoo. mBio 2020, 11. [Google Scholar] [CrossRef]
- Halfmann, P.J.; Hatta, M.; Chiba, S.; Maemura, T.; Fan, S.; Takeda, M.; Kinoshita, N.; Hattori, S.-I.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; et al. Transmission of SARS-CoV-2 in Domestic Cats. N. Engl. J. Med. 2020, 383, 592–594. [Google Scholar] [CrossRef]
- Patterson, E.I.; Elia, G.; Grassi, A.; Giordano, A.; Desario, C.; Medardo, M.; Smith, S.L.; Anderson, E.R.; Prince, T.; Patterson, G.T.; et al. Evidence of Exposure to SARS-CoV-2 in Cats and Dogs from Households in Italy. Nat. Commun. 2020, 11, 6231. [Google Scholar] [CrossRef]
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of Dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.-W.; Zhang, A.J.; Yuan, S.; Poon, V.K.-M.; Chan, C.C.-S.; Lee, A.C.-Y.; Chan, W.-M.; Fan, Z.; Tsoi, H.-W.; Wen, L.; et al. Simulation of the Clinical and Pathological Manifestations of Coronavirus Disease 2019 (COVID-19) in a Golden Syrian Hamster Model: Implications for Disease Pathogenesis and Transmissibility. Clin. Infect. Dis. 2020, 71, 2428–2446. [Google Scholar] [CrossRef] [PubMed]
- Sia, S.F.; Yan, L.-M.; Chin, A.W.H.; Fung, K.; Choy, K.-T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.A.P.M.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and Transmission of SARS-CoV-2 in Golden Hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Bao, L.; Yu, P.; Qi, F.; Gong, S.; Wang, J.; Zhao, B.; Liu, M.; Han, Y.; Deng, W.; et al. SARS-CoV-2 Causes a Systemically Multiple Organs Damages and Dissemination in Hamsters. Front. Microbiol. 2020, 11, 618891. [Google Scholar] [CrossRef] [PubMed]
- Mykytyn, A.Z.; Lamers, M.M.; Okba, N.M.A.; Breugem, T.I.; Schipper, D.; van den Doel, P.B.; van Run, P.; van Amerongen, G.; de Waal, L.; Koopmans, M.P.G.; et al. Susceptibility of Rabbits to SARS-CoV-2. Emerg. Microbes Infect. 2021, 10, 1–7. [Google Scholar] [CrossRef]
- Ulrich, L.; Wernike, K.; Hoffmann, D.; Mettenleiter, T.C.; Beer, M. Experimental Infection of Cattle with SARS-CoV-2. Emerg. Infect. Dis. 2020, 26, 2979–2981. [Google Scholar] [CrossRef]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F.; et al. The Pathogenicity of SARS-CoV-2 in HACE2 Transgenic Mice. Nature 2020, 583, 830–833. [Google Scholar] [CrossRef]
- Sun, S.-H.; Chen, Q.; Gu, H.-J.; Yang, G.; Wang, Y.-X.; Huang, X.-Y.; Liu, S.-S.; Zhang, N.-N.; Li, X.-F.; Xiong, R.; et al. A Mouse Model of SARS-CoV-2 Infection and Pathogenesis. Cell Host Microbe 2020, 28, 124–133.e4. [Google Scholar] [CrossRef]
- Lu, S.; Zhao, Y.; Yu, W.; Yang, Y.; Gao, J.; Wang, J.; Kuang, D.; Yang, M.; Yang, J.; Ma, C.; et al. Comparison of Nonhuman Primates Identified the Suitable Model for COVID-19. Signal. Transduct. Target. 2020, 5, 157. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Yao, Y.-F.; Yang, X.-L.; Zhou, Y.-W.; Gao, G.; Peng, Y.; Yang, L.; Hu, X.; Xiong, J.; Jiang, R.-D.; et al. Infection with Novel Coronavirus (SARS-CoV-2) Causes Pneumonia in Rhesus Macaques. Cell Res. 2020, 30, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Li, H.; Guo, L.; Liang, Y.; Li, J.; Wang, X.; Hu, Y.; Wang, L.; Liao, Y.; Yang, F.; et al. Virulence and Pathogenesis of SARS-CoV-2 Infection in Rhesus Macaques: A Nonhuman Primate Model of COVID-19 Progression. PloS Pathog. 2020, 16, e1008949. [Google Scholar] [CrossRef]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.-E.; Williamson, M.K.; Antón-Plágaro, C.; Shoemark, D.K.; Simón-Gracia, L.; Bauer, M.; Hollandi, R.; et al. Neuropilin-1 Is a Host Factor for SARS-CoV-2 Infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Uzoigwe, C.E. Body Temperature and Host Species Preferences of SARS-CoV-2. Clin. Microbiol. Infect. 2020, 26, 1709–1710. [Google Scholar] [CrossRef] [PubMed]
- Smit, L.A.M.; van der Sman-de Beer, F.; Opstal-van Winden, A.W.J.; Hooiveld, M.; Beekhuizen, J.; Wouters, I.M.; Yzermans, J.; Heederik, D. Q Fever and Pneumonia in an Area with a High Livestock Density: A Large Population-Based Study. PLoS ONE 2012, 7, e38843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huijskens, E.G.W.; Smit, L.A.M.; Rossen, J.W.A.; Heederik, D.; Koopmans, M. Evaluation of Patients with Community-Acquired Pneumonia Caused by Zoonotic Pathogens in an Area with a High Density of Animal Farms. Zoonoses Public Health 2016, 63, 160–166. [Google Scholar] [CrossRef]
- Dub, T.; Ollgren, J.; Huusko, S.; Uusitalo, R.; Siljander, M.; Vapalahti, O.; Sane, J. Game Animal Density, Climate, and Tick-Borne Encephalitis in Finland, 2007-2017. Emerg. Infect. Dis. 2020, 26, 2899–2906. [Google Scholar] [CrossRef]
- Wong, D.W.S.; Li, Y. Spreading of COVID-19: Density Matters. PLoS ONE 2020, 15, e0242398. [Google Scholar] [CrossRef]
- Kulu, H.; Dorey, P. Infection Rates from Covid-19 in Great Britain by Geographical Units: A Model-Based Estimation from Mortality Data. Health Place 2021, 67, 102460. [Google Scholar] [CrossRef]
- Tran, P.; Tran, L.; Tran, L. The Influence of Social Distancing on COVID-19 Mortality in US Counties: Cross-Sectional Study. JMIR Public Health Surveill 2021. [Google Scholar] [CrossRef]
- Hess, G. Disease in Metapopulation Models: Implications for Conservation. Ecology 1996, 77, 1617–1632. [Google Scholar] [CrossRef]
- North, A.R.; Godfray, H.C.J. The Dynamics of Disease in a Metapopulation: The Role of Dispersal Range. J. Biol. 2017, 418, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Fulford, G.R.; Roberts, M.G.; Heesterbeek, J.A.P. The Metapopulation Dynamics of an Infectious Disease: Tuberculosis in Possums. Popul Biol. 2002, 61, 15–29. [Google Scholar] [CrossRef] [Green Version]
- Craft, M.; Hawthorne, P.; Packer, C.; Dobson, A. Dynamics of a Multihost Pathogen in a Carnivore Community. J. Anim. Ecol. 2008, 77, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Stopka, P.; Graciasová, R. Conditional Allogrooming in the Herb-Field Mouse. Behav. Ecol. 2001, 12, 584–589. [Google Scholar] [CrossRef] [Green Version]
- Dunbar, R.I.M. Functional Significance of Social Grooming in Primates. Folia Primatol. 1991, 57, 121–131. [Google Scholar] [CrossRef]
- MacIntosh, A.J.J.; Jacobs, A.; Garcia, C.; Shimizu, K.; Mouri, K.; Huffman, M.A.; Hernandez, A.D. Monkeys in the Middle: Parasite Transmission through the Social Network of a Wild Primate. PLoS ONE 2012, 7, e51144. [Google Scholar] [CrossRef]
- Carter, G.; Leffer, L. Social Grooming in Bats: Are Vampire Bats Exceptional? PLoS ONE 2015, 10, e0138430. [Google Scholar] [CrossRef] [Green Version]
- Walsh, M.G.; Wiethoelter, A.; Haseeb, M.A. The Impact of Human Population Pressure on Flying Fox Niches and the Potential Consequences for Hendra Virus Spillover. Sci. Rep. 2017, 7, 8226. [Google Scholar] [CrossRef] [PubMed]
- Frutos, R.; Gavotte, L.; Devaux, C.A. Understanding the Origin of COVID-19 Requires to Change the Paradigm on Zoonotic Emergence. SSRN J. 2020. [Google Scholar] [CrossRef]
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; Lloyd-Smith, J.O. Pathways to Zoonotic Spillover. Nat. Rev. Microbiol. 2017, 15, 502–510. [Google Scholar] [CrossRef] [Green Version]
- Younes, S.; Younes, N.; Shurrab, F.; Nasrallah, G.K. Severe Acute Respiratory Syndrome Coronavirus-2 Natural Animal Reservoirs and Experimental Models: Systematic Review. Rev. Med. Virol. 2020, e2196. [Google Scholar] [CrossRef]
- Woolsey, C.; Borisevich, V.; Prasad, A.N.; Agans, K.N.; Deer, D.J.; Dobias, N.S.; Heymann, J.C.; Foster, S.L.; Levine, C.B.; Medina, L.; et al. Establishment of an African Green Monkey Model for COVID-19 and Protection against Re-Infection. Nat. Immunol. 2021, 22, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Cross, R.W.; Agans, K.N.; Prasad, A.N.; Borisevich, V.; Woolsey, C.; Deer, D.J.; Dobias, N.S.; Geisbert, J.B.; Fenton, K.A.; Geisbert, T.W. Intranasal Exposure of African Green Monkeys to SARS-CoV-2 Results in Acute Phase Pneumonia with Shedding and Lung Injury Still Present in the Early Convalescence Phase. Virol. J. 2020, 17, 125. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.-C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef]
- Hewitt, J.A.; Lutz, C.; Florence, W.C.; Pitt, M.L.M.; Rao, S.; Rappaport, J.; Haigwood, N.L. ACTIVating Resources for the COVID-19 Pandemic: In Vivo Models for Vaccines and Therapeutics. Cell Host Microbe 2020, 28, 646–659. [Google Scholar] [CrossRef]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian Hamsters as a Small Animal Model for SARS-CoV-2 Infection and Countermeasure Development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef]
- Bertzbach, L.D.; Vladimirova, D.; Dietert, K.; Abdelgawad, A.; Gruber, A.D.; Osterrieder, N.; Trimpert, J. SARS-CoV-2 Infection of Chinese Hamsters (Cricetulus Griseus) Reproduces COVID-19 Pneumonia in a Well-Established Small Animal Model. Transbound Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Deng, W.; Bao, L.; Gao, H.; Xiang, Z.; Qu, Y.; Song, Z.; Gong, S.; Liu, J.; Liu, J.; Yu, P.; et al. Ocular Conjunctival Inoculation of SARS-CoV-2 Can Cause Mild COVID-19 in Rhesus Macaques. Nat. Commun. 2020, 11, 4400. [Google Scholar] [CrossRef]
- Deng, W.; Bao, L.; Liu, J.; Xiao, C.; Liu, J.; Xue, J.; Lv, Q.; Qi, F.; Gao, H.; Yu, P.; et al. Primary Exposure to SARS-CoV-2 Protects against Reinfection in Rhesus Macaques. Science 2020, 369, 818–823. [Google Scholar] [CrossRef]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Porter, S.M.; Gordy, P.W.; Nehring, M.; Byas, A.D.; VandeWoude, S.; Ragan, I.K.; Maison, R.M.; Bowen, R.A. Experimental Infection of Domestic Dogs and Cats with SARS-CoV-2: Pathogenesis, Transmission, and Response to Reexposure in Cats. Proc. Natl. Acad. Sci. USA 2020, 117, 26382–26388. [Google Scholar] [CrossRef]
- Ryan, K.A.; Bewley, K.R.; Fotheringham, S.A.; Slack, G.S.; Brown, P.; Hall, Y.; Wand, N.I.; Marriott, A.C.; Cavell, B.E.; Tree, J.A.; et al. Dose-Dependent Response to Infection with SARS-CoV-2 in the Ferret Model and Evidence of Protective Immunity. Nat. Commun. 2021, 12, 81. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.O.; Case, J.B.; Winkler, E.S.; Thackray, L.B.; Kafai, N.M.; Bailey, A.L.; McCune, B.T.; Fox, J.M.; Chen, R.E.; Alsoussi, W.B.; et al. A SARS-CoV-2 Infection Model in Mice Demonstrates Protection by Neutralizing Antibodies. Cell 2020, 182, 744–753.e4. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, K.; Wang, T.; Deng, F.; Gong, W.; Hui, X.; Zhao, Y.; He, X.; Li, C.; Zhang, Q.; et al. SARS-CoV-2 Rapidly Adapts in Aged BALB/c Mice and Induces Typical Pneumonia. J. Virol. 2021. [Google Scholar] [CrossRef]
- Major, J.; Wack, A. An Ace Model for SARS-CoV-2 Infection. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Sun, J.; Zhuang, Z.; Zheng, J.; Li, K.; Wong, R.L.-Y.; Liu, D.; Huang, J.; He, J.; Zhu, A.; Zhao, J.; et al. Generation of a Broadly Useful Model for COVID-19 Pathogenesis, Vaccination, and Treatment. Cell 2020, 182, 734–743.e5. [Google Scholar] [CrossRef]
- Golden, J.W.; Cline, C.R.; Zeng, X.; Garrison, A.R.; Carey, B.D.; Mucker, E.M.; White, L.E.; Shamblin, J.D.; Brocato, R.L.; Liu, J.; et al. Human Angiotensin-Converting Enzyme 2 Transgenic Mice Infected with SARS-CoV-2 Develop Severe and Fatal Respiratory Disease. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Winkler, E.S.; Bailey, A.L.; Kafai, N.M.; Nair, S.; McCune, B.T.; Yu, J.; Fox, J.M.; Chen, R.E.; Earnest, J.T.; Keeler, S.P.; et al. SARS-CoV-2 Infection of Human ACE2-Transgenic Mice Causes Severe Lung Inflammation and Impaired Function. Nat. Immunol. 2020, 21, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.-D.; Liu, M.-Q.; Chen, Y.; Shan, C.; Zhou, Y.-W.; Shen, X.-R.; Li, Q.; Zhang, L.; Zhu, Y.; Si, H.-R.; et al. Pathogenesis of SARS-CoV-2 in Transgenic Mice Expressing Human Angiotensin-Converting Enzyme 2. Cell 2020, 182, 50–58.e8. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Blair, R.V.; Iwanaga, N.; Liu, F.; Russell-Lodrigue, K.E.; Qin, Z.; Midkiff, C.C.; Golden, N.A.; Doyle-Meyers, L.A.; Kabir, M.E.; et al. Lung Expression of Human Angiotensin-Converting Enzyme 2 Sensitizes the Mouse to SARS-CoV-2 Infection. Am. J. Respir Cell Mol. Biol. 2021, 64, 79–88. [Google Scholar] [CrossRef]
- Oladunni, F.S.; Park, J.-G.; Pino, P.A.; Gonzalez, O.; Akhter, A.; Allué-Guardia, A.; Olmo-Fontánez, A.; Gautam, S.; Garcia-Vilanova, A.; Ye, C.; et al. Lethality of SARS-CoV-2 Infection in K18 Human Angiotensin-Converting Enzyme 2 Transgenic Mice. Nat. Commun. 2020, 11, 6122. [Google Scholar] [CrossRef]
- Yinda, C.K.; Port, J.R.; Bushmaker, T.; Offei Owusu, I.; Purushotham, J.N.; Avanzato, V.A.; Fischer, R.J.; Schulz, J.E.; Holbrook, M.G.; Hebner, M.J.; et al. K18-HACE2 Mice Develop Respiratory Disease Resembling Severe COVID-19. PloS Pathog. 2021, 17, e1009195. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Chen, Q.; Yang, G.; He, L.; Fan, H.; Deng, Y.-Q.; Wang, Y.; Teng, Y.; Zhao, Z.; Cui, Y.; et al. Adaptation of SARS-CoV-2 in BALB/c Mice for Testing Vaccine Efficacy. Science 2020, 369, 1603–1607. [Google Scholar] [CrossRef] [PubMed]
- Dinnon, K.H.; Leist, S.R.; Schäfer, A.; Edwards, C.E.; Martinez, D.R.; Montgomery, S.A.; West, A.; Yount, B.L.; Hou, Y.J.; Adams, L.E.; et al. A Mouse-Adapted SARS-CoV-2 Model for the Evaluation of COVID-19 Medical Countermeasures. bioRxiv 2020. [Google Scholar] [CrossRef]
- Leist, S.R.; Dinnon, K.H.; Schäfer, A.; Tse, L.V.; Okuda, K.; Hou, Y.J.; West, A.; Edwards, C.E.; Sanders, W.; Fritch, E.J.; et al. A Mouse-Adapted SARS-CoV-2 Induces Acute Lung Injury and Mortality in Standard Laboratory Mice. Cell 2020, 183, 1070–1085.e12. [Google Scholar] [CrossRef] [PubMed]
- Sefik, E.; Israelow, B.; Zhao, J.; Qu, R.; Song, E.; Mirza, H.; Kaffe, E.; Halene, S.; Meffre, E.; Kluger, Y.; et al. A Humanized Mouse Model of Chronic COVID-19 to Evaluate Disease Mechanisms and Treatment Options. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Lee, A.C.-Y.; Zhang, A.J.; Chan, J.F.-W.; Li, C.; Fan, Z.; Liu, F.; Chen, Y.; Liang, R.; Sridhar, S.; Cai, J.-P.; et al. Oral SARS-CoV-2 Inoculation Establishes Subclinical Respiratory Infection with Virus Shedding in Golden Syrian Hamsters. Cell Rep. Med. 2020, 1, 100121. [Google Scholar] [CrossRef]
- Osterrieder, N.; Bertzbach, L.D.; Dietert, K.; Abdelgawad, A.; Vladimirova, D.; Kunec, D.; Hoffmann, D.; Beer, M.; Gruber, A.D.; Trimpert, J. Age-Dependent Progression of SARS-CoV-2 Infection in Syrian Hamsters. Viruses 2020, 12, 779. [Google Scholar] [CrossRef]
- Vergara-Alert, J.; Rodon, J.; Carrillo, J.; Te, N.; Izquierdo-Useros, N.; Rodríguez de la Concepción, M.L.; Ávila-Nieto, C.; Guallar, V.; Valencia, A.; Cantero, G.; et al. Pigs Are Not Susceptible to SARS-CoV-2 Infection but Are a Model for Viral Immunogenicity Studies. Transbound Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Meekins, D.A.; Morozov, I.; Trujillo, J.D.; Gaudreault, N.N.; Bold, D.; Carossino, M.; Artiaga, B.L.; Indran, S.V.; Kwon, T.; Balaraman, V.; et al. Susceptibility of Swine Cells and Domestic Pigs to SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 2278–2288. [Google Scholar] [CrossRef]
- Pickering, B.S.; Smith, G.; Pinette, M.M.; Embury-Hyatt, C.; Moffat, E.; Marszal, P.; Lewis, C.E. Susceptibility of Domestic Swine to Experimental Infection with Severe Acute Respiratory Syndrome Coronavirus 2. Emerg. Infect. Dis. 2021, 27, 104–112. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, J.; Kuang, D.; Xu, J.; Yang, M.; Ma, C.; Zhao, S.; Li, J.; Long, H.; Ding, K.; et al. Susceptibility of Tree Shrew to SARS-CoV-2 Infection. Sci. Rep. 2020, 10, 16007. [Google Scholar] [CrossRef]
- Suarez, D.L.; Pantin-Jackwood, M.J.; Swayne, D.E.; Lee, S.A.; DeBlois, S.M.; Spackman, E. Lack of Susceptibility to SARS-CoV-2 and MERS-CoV in Poultry. Emerg. Infect. Dis. 2020, 26, 3074–3076. [Google Scholar] [CrossRef] [PubMed]
- Barrs, V.R.; Peiris, M.; Tam, K.W.S.; Law, P.Y.T.; Brackman, C.J.; To, E.M.W.; Yu, V.Y.T.; Chu, D.K.W.; Perera, R.A.P.M.; Sit, T.H.C. SARS-CoV-2 in Quarantined Domestic Cats from COVID-19 Households or Close Contacts, Hong Kong, China. Emerg. Infect. Dis. 2020, 26, 3071–3074. [Google Scholar] [CrossRef]
- Garigliany, M.; Van Laere, A.-S.; Clercx, C.; Giet, D.; Escriou, N.; Huon, C.; van der Werf, S.; Eloit, M.; Desmecht, D. SARS-CoV-2 Natural Transmission from Human to Cat, Belgium, March 2020. Emerg. Infect. Dis. 2020, 26, 3069–3071. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Jin, Y.; Liu, Y.; Sun, J.; Hao, L.; Bai, J.; Huang, T.; Lin, D.; Jin, Y.; Tian, K. Serological Survey of SARS-CoV-2 for Experimental, Domestic, Companion and Wild Animals Excludes Intermediate Hosts of 35 Different Species of Animals. Transbound Emerg. Dis. 2020, 67, 1745–1749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, H.; Gao, J.; Huang, K.; Yang, Y.; Hui, X.; He, X.; Li, C.; Gong, W.; Zhang, Y.; et al. A Serological Survey of SARS-CoV-2 in Cat in Wuhan. Emerg. Microbes Infect. 2020, 9, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, Y.; Gao, J.; Huang, K.; Hu, C.; Hui, X.; He, X.; Li, C.; Gong, W.; Lv, C.; et al. A Serological Survey of Severe Acute Respiratory Syndrome Coronavirus 2 in Wuhan. Transbound Emerg. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Stevanovic, V.; Vilibic-Cavlek, T.; Tabain, I.; Benvin, I.; Kovac, S.; Hruskar, Z.; Mauric, M.; Milasincic, L.; Antolasic, L.; Skrinjaric, A.; et al. Seroprevalence of SARS-CoV-2 Infection among Pet Animals in Croatia and Potential Public Health Impact. Transbound Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Fritz, M.; Rosolen, B.; Krafft, E.; Becquart, P.; Elguero, E.; Vratskikh, O.; Denolly, S.; Boson, B.; Vanhomwegen, J.; Gouilh, M.A.; et al. High Prevalence of SARS-CoV-2 Antibodies in Pets from COVID-19+ Households. One Health 2021, 11, 100192. [Google Scholar] [CrossRef] [PubMed]
- Larsen, C.S.; Paludan, S.R. Corona’s New Coat: SARS-CoV-2 in Danish Minks and Implications for Travel Medicine. Travel Med. Infect. Dis. 2020, 38, 101922. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lau, C.C.Y.; Tsang, A.K.L.; Lau, J.H.N.; Bai, R.; Teng, J.L.L.; Tsang, C.C.C.; Wang, M.; et al. Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Ge, X.; Wang, L.-F.; Shi, Z. Bat Origin of Human Coronaviruses. Virol. J. 2015, 12, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, T.S.; Liu, D.X. Human Coronavirus: Host-Pathogen Interaction. Annu. Rev. Microbiol. 2019, 73, 529–557. [Google Scholar] [CrossRef] [Green Version]
- King, A. An Uncommon Cold. New Sci. 2020, 246, 32–35. [Google Scholar] [CrossRef]
- Vijgen, L.; Keyaerts, E.; Moës, E.; Thoelen, I.; Wollants, E.; Lemey, P.; Vandamme, A.-M.; Van Ranst, M. Complete Genomic Sequence of Human Coronavirus OC43: Molecular Clock Analysis Suggests a Relatively Recent Zoonotic Coronavirus Transmission Event. J. Virol. 2005, 79, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- Durães-Carvalho, R.; Caserta, L.C.; Barnabé, A.C.S.; Martini, M.C.; Ferreira, H.L.; Felippe, P.A.N.; Santos, M.B.; Arns, C.W. Coronaviruses Detected in Brazilian Wild Birds Reveal Close Evolutionary Relationships with Beta- and Deltacoronaviruses Isolated From Mammals. J. Mol. Evol. 2015, 81. [Google Scholar] [CrossRef]
- Masry, I.E. Exposure of humans or animals to SARS-CoV-2 from wild, livestock, companion and aquatic animals: Qualitative exposure assessment. In FAO animal production and health, Paper, 181; FAO: Rome, Italy, 2020; Volume 38. [Google Scholar]
- Boinski, S.; Timm, R.M. Predation by Squirrel Monkeys and Double-Toothed Kites on Tent-Making Bats. Am. J. Primatol. 1985, 9, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Souza, L.L.; Ferrari, S.F.; Pina, A.L.U.; Cia, C.B. Feeding Behaviour and Predation of a Bat by Saimiri Sciureus in a Semi-Natural Amazonian Environment. Available online: https://eurekamag.com/research/067/774/067774856.php (accessed on 6 February 2021).
- Milano, M.Z.; Monteiro-Filho, E.L.A. Predation on Small Mammals by Capuchin Monkeys, Cebus Cay. Neotrop. Primates 2009, 16, 78–80. [Google Scholar] [CrossRef] [Green Version]
- Tapanes, E.; Detwiler, K.M.; Cords, M. Bat Predation by Cercopithecus Monkeys: Implications for Zoonotic Disease Transmission. EcoHealth 2016, 13, 405–409. [Google Scholar] [CrossRef]
- Sparks, D.W.; Simmons, M.T.; Gummer, C.L.; Duchamp, J.E. Disturbance of Roosting Bats by Woodpeckers and Raccoons. Northeast. Nat. 2003, 10, 105–108. [Google Scholar] [CrossRef]
- McAlpine, D.; Vanderwolf, K.; Forbes, G.; Malloch, D. Consumption of Bats (Myotis spp.) by Raccoons (Procyon Lotor) during an Outbreak of White-Nose Syndrome in New Brunswick, Canada: Implications for Estimates of Bat Mortality. Can. Field Nat. 2011, 125, 157–160. [Google Scholar] [CrossRef] [Green Version]
- Forman, D. Evidence of Lesser Horseshoe Bat (Rhinolophus Hipposideros) Predation by Otter (Lutra Lutra) in a Welsh Cave System. Lutra 2007, 47, 53–55. [Google Scholar]
- Goodpaster, W.; Hoffmeister, D.F. Bats as Prey for Mink in Kentucky Cave. J. Mammal. 1950, 31, 457. [Google Scholar] [CrossRef]
- Tiunov, M.; Yudin, V. Sable predation at bats wintering (Chiroptera). Ecology 1986, 5, 84–85. [Google Scholar]
- Quick, H.F. Notes on the Ecology of Weasels in Gunnison County, Colorado. J. Mammal. 1951, 32, 281–290. [Google Scholar] [CrossRef]
- Zhigalin, A. Siberian Weasel Mustela Sibirica Pallas, 1773 Predatism on Bats during Winter Period. Subterr. Biol. 2019, 32, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Trebbien, R.; Chriel, M.; Struve, T.; Hjulsager, C.K.; Larsen, G.; Larsen, L.E. Wildlife Reservoirs of Canine Distemper Virus Resulted in a Major Outbreak in Danish Farmed Mink (Neovison Vison). PLoS ONE 2014, 9, e85598. [Google Scholar] [CrossRef] [Green Version]
- World Organisation for Animal Health (OIE). Infection with SARS-COV-2 in Animals: Aetiology Epidemiology Diagnosis Prevention and Control References (OIE Technical Factsheet); OIE: Paris, France, 2021.
- DeLiberto, T. Coronavirus Disease 2019 Update (536): Animal, USA (UTAH), Wild Mink, First Case. ProMed. 2020. Available online: https://promedmail.org/promed-post/?id=8015608 (accessed on 1 February 2021).
- Aguiló-Gisbert, J.; Padilla-Blanco, M.; Lizana, V.; Maiques, E.; Baquero, M.M.; Chillida-Martínez, E.; Cardells, J.; Rubio-Guerri, C. First Description of Natural SARS-CoV-2 Infection in Two Wild American Minks (Neovison Vison). Preprints 2021. [Google Scholar] [CrossRef]
- Huang, Q.; Zhan, X.; Zeng, X.-T. COVID-19 Pandemic: Stop Panic Abandonment of Household Pets. J. Travel Med. 2020, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, H.J.; Reifman, J. Quantitative Modeling of Virus Evolutionary Dynamics and Adaptation in Serial Passages Using Empirically Inferred Fitness Landscapes. J. Virol. 2014, 88, 1039–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepin, K.M.; Lass, S.; Pulliam, J.R.C.; Read, A.F.; Lloyd-Smith, J.O. Identifying Genetic Markers of Adaptation for Surveillance of Viral Host Jumps. Nat. Rev. Microbiol. 2010, 8, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, X.; Hu, T.; Li, J.; Song, H.; Liu, Y.; Wang, P.; Liu, D.; Yang, J.; Holmes, E.C.; et al. A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. Curr. Biol. 2020, 30, 2196–2203.e3. [Google Scholar] [CrossRef]
- Hon, C.-C.; Lam, T.-Y.; Shi, Z.-L.; Drummond, A.J.; Yip, C.-W.; Zeng, F.; Lam, P.-Y.; Leung, F.C.-C. Evidence of the Recombinant Origin of a Bat Severe Acute Respiratory Syndrome (SARS)-Like Coronavirus and Its Implications on the Direct Ancestor of SARS Coronavirus. JVI 2008, 82, 1819–1826. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Species (Human ACE2 and ACE2 Orthologs) | In Silico, Data from [39,40,41,42,43,44,45,46,47,48] 1 | In Vitro (in Cells), Data from [48,49], S = Susceptible, NS = Non Susceptible, to SARS-CoV-2 Infection | In Vivo, S = Susceptible, NS = Non Susceptible, to SARS-CoV-2 Infection |
|---|---|---|---|
| Human (Homo sapiens) | Yes (+++) | S for Calu3 cell sand Caco2 | COVID-19 outbreak [15,50,51] |
| Monkeys (Gorilla gorilla gorilla, Macaca mulatta; Pan troglodytes, Pongo abelii, Papio Anubis) | Yes (+++) | S for VeroE6 cells and FRhK4 cells, and for HEK293 cells expressing the monkey (M. mulatta) ACE2 | S (COVID-19-like signs) [52,53] |
| Monkeys (Callithrix jacchus/marmoset, tufted capuchin, squirrel monkey)) | Undetermined (− to ++) | S for HeLa cells expressing the monkey (marmoset) ACE2 | |
| Ferret (Mustela putorius furo) | Yes (++) | S (COVID-19-like signs) [54,55,56,57,58] | |
| Mink (Mustela lutreola; Neovison vison) | S (COVID-19-like signs) Mink-to-mink transmission and mink-to-human transmission reported [59,60,61] | ||
| Ermine/short tailed weasel (Mustela erminea) | Yes (++) | ||
| Raccoon dog (Nyctereutes procyonoides) | S (with minor clinical signs) Raccoon dog to raccoon dog transmission [62] | ||
| Civet (Paguma larvata) | Undetermined (− to ++) | ||
| Pangolin (Manis javanica) | Yes (+++) | ||
| Pangolins (Manis pentadactyla, Smutsia temminckii; Phataginus tricuspis) | No (−) | ||
| Bats (Rhinolophus sinicus; Rhinolophus pearsonii; Rhinolophus macrotis) | Yes (+++) | S [56] | |
| Bats (Rhinolophus ferrumequinum, Myotis) | No (−) | ||
| Bat (Desmodus rotundus) | No (−) | ||
| Camel (Camelus dromedarius) | Undetermined (− to ++) | ||
| Lion (Panthera leo) | S [63] | ||
| Tiger (Panthera tigris) | Yes (++) | S [45,63] | |
| Cat (Felis catus) | Yes (+++) | S for CRFK cells and HEK293 cells expressing the cat (F. catus) ACE2 | S (COVID-19-like signs) Cat-to-cat transmission and Human -to- cat transmission have been reported [57,64,65] |
| Dog (Canis lupus familiaris, Canis lupus dingo) | Yes (++) | S for HEK293 cells expressing the dog (C. lupus) ACE2 | S, yet the virus replicates very poorly (Human -to- dog transmission has been reported) [57,65,66] |
| Hamster (Mesocricetus auratus) | Yes (++) | S (COVID-19-like signs) [67,68,69] | |
| Rabbit (Oryctolagus cuniculus) | Yes (++) | S for HEK293 cells expressing the rabbit (O. cuniculus) ACE2 | S. Infected animals produce virus [70] |
| Pig (Sus scrofa domesticus) | Yes (++) | S for PK-15 cells and HeLa cells expressing the pig (S. scrofa) ACE2 | S, yet the virus replicates very poorly [56,57] |
| Boar (Sus scrofa) | Yes (++) | ||
| Cow (Bos taurus) | Yes (++) | S for HeLa cells expressing the cow (B. taurus) ACE2 | S, yet the virus replicates very poorly Cow-to-cow transmission [71] |
| Buffalo (Bubalus bubalus) | Yes (++) | ||
| Goat (Capra hircus) | Yes (++) | ||
| Sheep (Ovis aries) | Yes (++) | ||
| Rats (Rattus rattus, Rattus norvegicus) | Undetermined (− to +) | NS for HEK293 cells expressing the rat (R. norvegicus) ACE2 | |
| Mouse (Mus musculus) | No (-) | NS for HeLa cells expressing the mouse (M. musculus) ACE2 | NS, (hACE2 humanized mice are susceptible to infection and show (COVID-19-like signs) [72,73] |
| Pigeon (Columbia livia) | Undetermined (− to +) | ||
| Hen (Gallus gallus) | Undetermined (− to +) | ||
| Chiken | S, yet the virus replicates very poorly [57] | ||
| Duck | S, yet the virus replicates very poorly [57] | ||
| Turtle (Pelodiscus sinensis; Chrysemys picta bellii, Chelonia mydas) | Undetermined (− to ++) | ||
| Snake (Ophiophagus hannah) | Undetermined (− to +) | ||
| Snake/Pallas pit viper (Protobothrops mucrosquamatus) | Yes (++) | ||
| Frog (Xenopus tropicalis) | No (−) | ||
| Whale/Yangtze finless porpoise (Neophocaena asiaeorientalis asiaeorientalis) | Yes (++) |
| Animal | Clinical Symptoms | Viral RNA Detection | Infectious Virus Detection | Pathological Lung Lesions | Other Organs Involved | Specific Antibody Response | Transmission to Contact Animals | References |
|---|---|---|---|---|---|---|---|---|
| Callithrix jacchus Macaca fasicularis Macaca mulatta | Fever, body weight loss | Nose, lower viral load in C. jacchus | Lung, for Macaca only | Interstitial pneumonia, more severe in M. mulatta | Spleen and lymph nodes for Macaca only | Only for Macaca | ND | [74] |
| Rhesus macaque (M. mullata) | Fever, loss of appetite and reduced activity | Nose and oropharynx, than rectal swabs, lungs, lymph nodes | Rectal swabs | Severe interstitial pneumonia | Brain, spinal cords, kidney, liver, spleen, heart, intestine and testicle | [76] | ||
| Rhesus macaque (M. mullata) IT | Fever, bodyweight loss, dehydration, tachypnea | Nose, oropharynx, anal swab, lungs, gut, lymphoid tissues, and rarely other tissues | Nose, oropharynx, anal swab, trachea, bronchus, lungs | Severe interstitial pneumonia | Gut, lymphoid tissues, spinal cord, heart, skeletal muscles and bladder | Yes | ND | [75] |
| African green monkey (IT and IN; or IN with MAD) | Fever, loss of appetite, pneumonia, and coagulation disorders | Nose, rectal swab, BAL fluid, lungs | Nose, rectal swab | Multifocal chronic interstitial pneumonia | Lymphoid tissue, heart, gut, bladder, brain, and eyes | Yes | ND | [98,99] |
| Egyptian fruit bats (Rousettus aegyptiacus) | None | Oral cavity, trachea, lungs, lymph nodes, heart, skin, duodenum, adrenal gland tissues | Nose, trachea | Yes | Yes | [56] | ||
| Dogs | None | Rectal swabs at 2 dpi only | No | No | No | Yes | No | [57] |
| Raccoon dogs | None | Nose, oropharynx | Nose, oropharynx | No | No | Yes | Yes | [62] |
| Cats | Mild or no symptoms | Nose, soft palates, tonsils, trachea, lungs, small intestine | Nose | Severe lung lesions | Yes | Yes | [57,64] | |
| Rabbits | No symptoms | Nose, throat, feces | Nose | Mild to moderate phagocytic cells infiltration | No | Yes | ND | [70] |
| Ferrets | Fever, reduced activity, occasional cough | Nose, saliva, urine, feces, and rarely the lungs, kidney, and intestine | Nose only | Acute bronchiolitis, mild multifocal bronchopneumonia, and severe lung lesions | Yes | Yes | [54,57,100,101] | |
| Syrian and Chinese hamsters | Body weight loss | Nose, oropharynx, trachea, and many other tissues | Nose, oropharynx, trachea | Severe lung lesions (milder but more prolonged in Chinese hamsters) | Yes | Yes | [67,68,102,103] |
| Start Date of the Outbreak | Zoo Location | Animals | Clinical Symptoms | Sources |
|---|---|---|---|---|
| 03/27/20 | WCS Bronx zoo, New York, USA | 4 tigers 1 (Panthera tigris) out of 5 | Respiratory signs | https://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=33885 (accessed on 5 February 2021) https://promedmail.org/promed-post/?id=7191352 (accessed on 5 February 2021) |
| 03/27/20 | WCS Bronx zoo, New York, USA | 3 lions 1 (Panthera leo) out of 3 | Respiratory signs | https://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=33885 (accessed on 5 February 2021) https://promedmail.org/promed-post/?id=7191352 (accessed on 5 February 2021) |
| 10/12/20 | Knoxville, Tennessee, USA | 3 tigers (Panthera tigris) out of 3 | Respiratory signs | https://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=36433https://promedmail.org/promed-post/?id=7915683 (accessed on 5 February 2021) |
| 11/27/20 | Jefferson Kentucky, USA | 3 snow leopards (Panthera uncia) out of | Respiratory signs | https://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=37147 (accessed on 5 February 2021) |
| 07/17/20 | Johannesburg, South Africa | 1 cougar (Puma concolor) out of 2 | NA | https://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=35399 (accessed on 5 February 2021) |
| 12/10/20 | Barcelona, Spain | 4 lions (Panthera leo) | Respiratory signs | https://promedmail.org/promed-post/?id=8002466 (accessed on 5 February 2021) |
| 01/06/21 | San Diego, California, USA | 3 gorilla (Gorilla gorilla gorilla) out of 8 | Respiratory signs for 2 of them | https://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=37553 (accessed on 5 February 2021) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maurin, M.; Fenollar, F.; Mediannikov, O.; Davoust, B.; Devaux, C.; Raoult, D. Current Status of Putative Animal Sources of SARS-CoV-2 Infection in Humans: Wildlife, Domestic Animals and Pets. Microorganisms 2021, 9, 868. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040868
Maurin M, Fenollar F, Mediannikov O, Davoust B, Devaux C, Raoult D. Current Status of Putative Animal Sources of SARS-CoV-2 Infection in Humans: Wildlife, Domestic Animals and Pets. Microorganisms. 2021; 9(4):868. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040868
Chicago/Turabian StyleMaurin, Max, Florence Fenollar, Oleg Mediannikov, Bernard Davoust, Christian Devaux, and Didier Raoult. 2021. "Current Status of Putative Animal Sources of SARS-CoV-2 Infection in Humans: Wildlife, Domestic Animals and Pets" Microorganisms 9, no. 4: 868. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040868