
Escherichia coli Affects Expression of Circadian Clock Genes in Human Hepatoma Cells

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data and Code Availability
2.2. Experimental Model and Subject Details
2.3. Microbial Strains
2.4. Method Details
2.4.1. Bacterial Growth Curves
2.4.2. Stable Co-Culture Preparation
2.4.3. 72 h Sampling
2.4.4. RNA Isolation and RT-PCR Analysis
2.4.5. Apoptosis-Necrosis Assay
2.4.6. Quantification and Statistical Analysis
3. Results
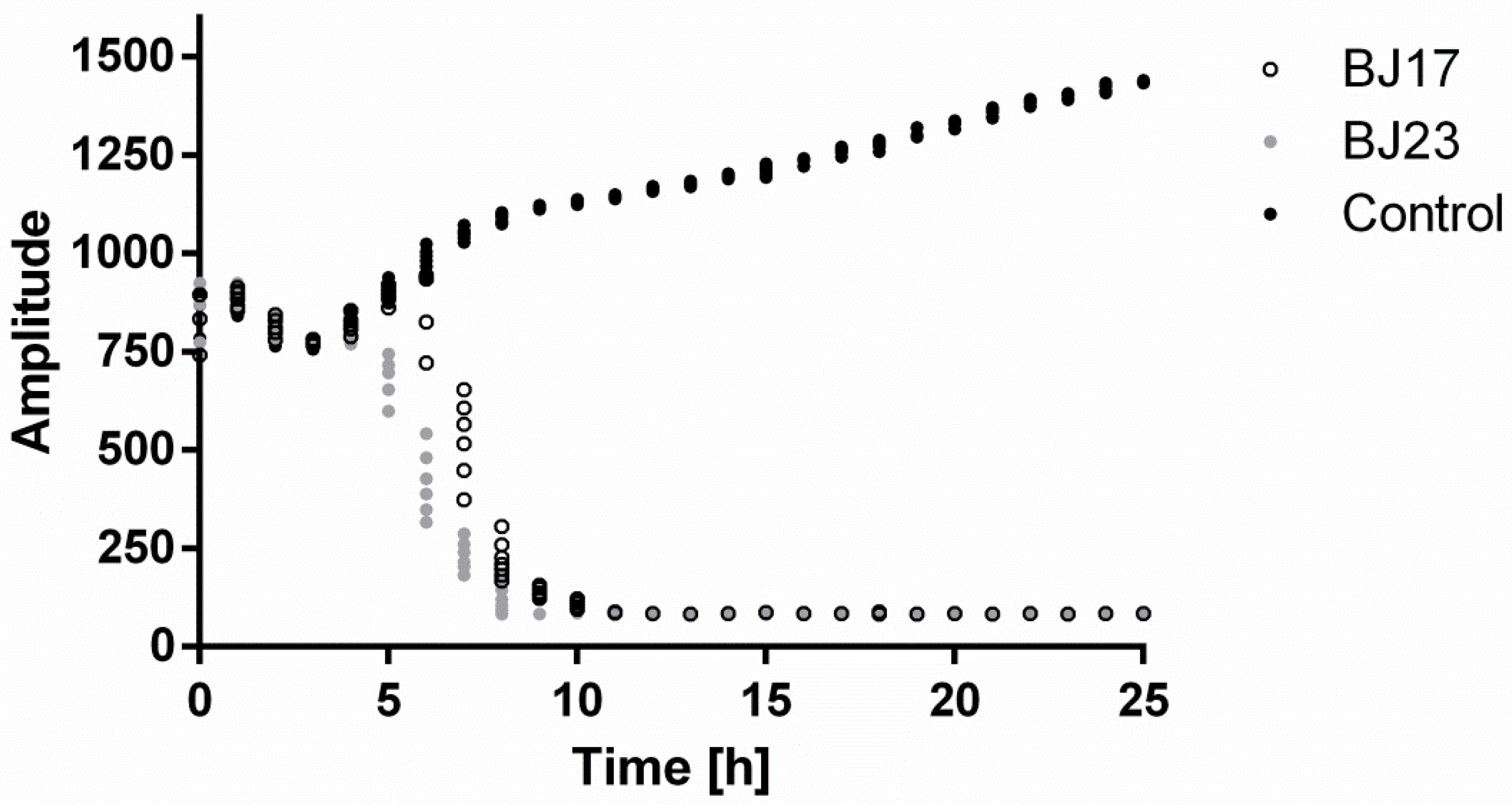
3.1. Assessing Growth Curves of E. coli Strains BJ17 and BJ23
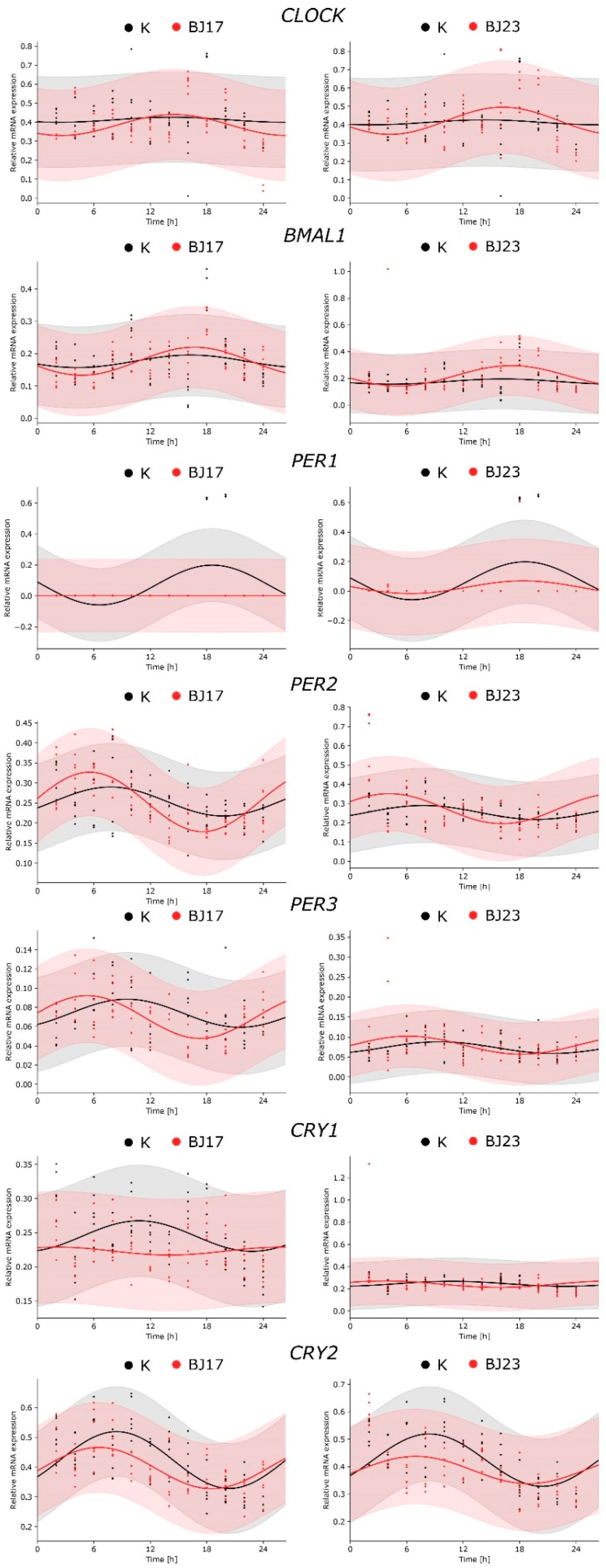
3.2. Co-Culture of HepG2 Cells with Bacterial Strains Is Stable for 4 h
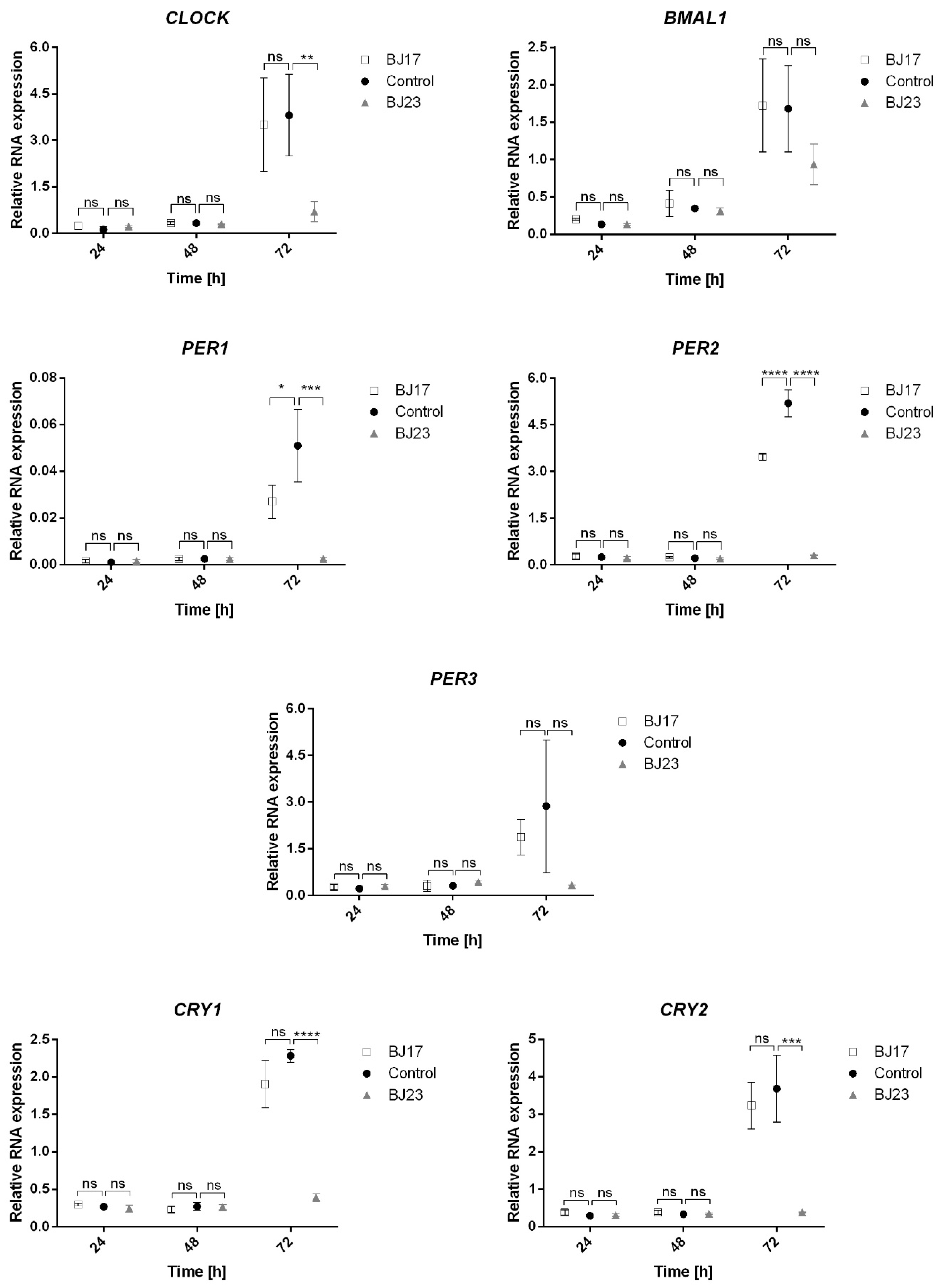
3.3. The 4 h Incubation of HepG2 Cells with E. coli Affects Expression of HepG2 Circadian Genes
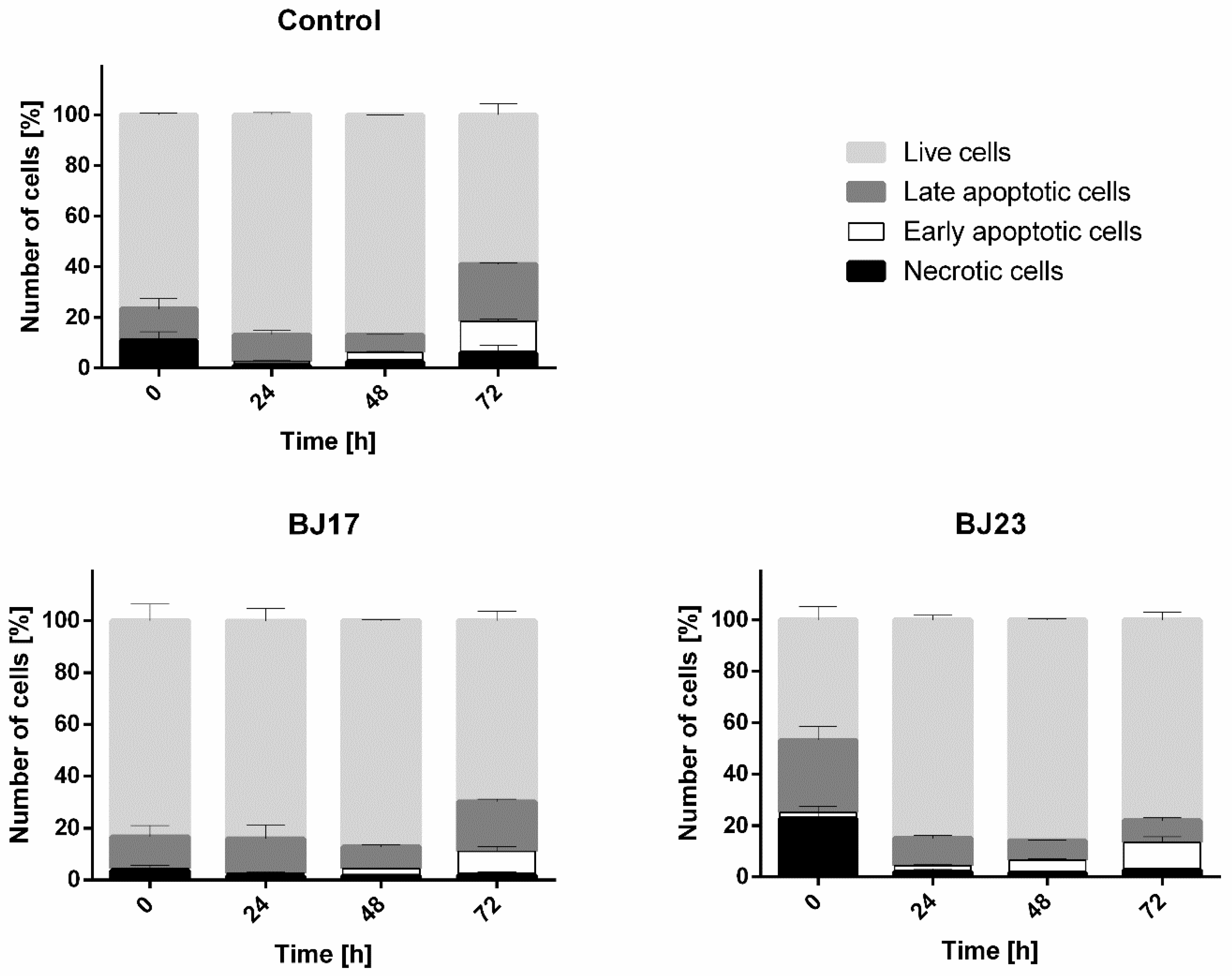
3.4. The 4 h Incubation of HepG2 Cells with the Bacterial Strain with More Virulence-Associated Genes Did Affect the HepG2 Cells Viability
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of diurnal variation of gut microbes and high-fat feeding on host circadian clock function and metabolism. Cell Host Microbe 2015, 17, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Parkar, S.G.; Kalsbeek, A.; Cheeseman, J.F. Potential role for the gut microbiota in modulating host circadian rhythms and metabolic health. Microorganisms 2019, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adlerberth, I.; Wold, A.E. Establishment of the gut microbiota in Western infants. Acta Paediatr. Int. J. Paediatr. 2009, 98, 229–238. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Butler, T.D.; Gibbs, J.E. Circadian host-microbiome interactions in immunity. Front. Immunol. 2020, 11, 1783. [Google Scholar] [CrossRef]
- Biedermann, L.; Rogler, G. The intestinal microbiota: Its role in health and disease. Eur. J. Pediatr. 2015, 174, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Tognini, P.; Murakami, M.; Sassone-Corsi, P. Interplay between microbes and the circadian clock. Cold Spring Harb. Perspect. Biol. 2018, 10, a028365. [Google Scholar] [CrossRef]
- Weger, B.D.; Gobet, C.; Yeung, J.; Martin, E.; Jimenez, S.; Betrisey, B.; Foata, F.; Berger, B.; Balvay, A.; Foussier, A.; et al. The mouse microbiome is required for sex-specific diurnal rhythms of gene expression and metabolism. Cell Metab. 2019, 29, 362–382.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota diurnal rhythmicity programs host transcriptome oscillations. Cell 2016, 167, 1495–1510.e12. [Google Scholar] [CrossRef] [Green Version]
- Silver, A.C. Pathogen-associated molecular patterns alter molecular clock gene expression in mouse splenocytes. PLoS ONE 2017, 12, 12–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, N.; Yoo, S.H.; Huang, H.C.; Kumar, V.; Lee, C.; Kim, T.K.; Takahashi, J.S. Transcriptional architecture and chromatin landscape of the core circadian clock in mammals. Science 2012, 338, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Russo, T.A. Extraintestinal pathogenic Escherichia coli: “The other bad E coli”. J. Lab. Clin. Med. 2002, 139, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Köhler, C.D.; Dobrindt, U. What defines extraintestinal pathogenic Escherichia coli? Int. J. Med. Microbiol. 2011, 301, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Brzuszkiewicz, E.; Gottschalk, G.; Ron, E.; Hacker, J.; Dobrindt, U. Adaptation of pathogenic E. coli to various niches: Genome flexibility is the key. Genome Dyn. 2009, 6, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Bast, A.; Schmidt, I.H.E.; Brauner, P.; Brix, B.; Breitbach, K.; Steinmetz, I. Defense mechanisms of hepatocytes against Burkholderia pseudomallei. Front. Microbiol. 2012, 2, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, B.; Zucoloto, A.Z.; Yu, I.L.; Burkhard, R.; Brown, K.; Geuking, M.B.; McCoy, K.D.; PMcDonald, B. Programing of an intravascular immune firewall by the gut microbiota protects against pathogen dissemination during infection. Cell Host Microbe 2020, 28, 660–668.e4. [Google Scholar] [CrossRef]
- Szabo, G.; Bala, S. Alcoholic liver disease and the gut-liver axis. World J. Gastroenterol. 2010, 16, 1321–1329. [Google Scholar] [CrossRef]
- Parlesak, A.; Schäfer, C.; Schütz, T.; Bode, J.C.; Bode, C. Increased intestinal permeability to macromolecules and endotoxemia in patients with chronic alcohol abuse in different stages of alcohol-induced liver disease. J. Hepatol. 2000, 32, 742–747. [Google Scholar] [CrossRef]
- Miele, L.; Valenza, V.; La Torre, G.; Montalto, M.; Cammarota, G.; Ricci, R.; Mascianà, R.; Forgione, A.; Gabrieli, M.L.; Perotti, G.; et al. Increased intestinal permeability and tight junction alterations in nonalcoholic fatty liver disease. Hepatology 2009, 49, 1877–1887. [Google Scholar] [CrossRef]
- Skubic, C.; Drakulić, Ž.; Rozman, D. Personalized therapy when tackling nonalcoholic fatty liver disease: A focus on sex, genes, and drugs. Expert Opin. Drug Metab. Toxicol. 2018, 14, 831–841. [Google Scholar] [CrossRef]
- Kosir, R.; Acimovic, J.; Golicnik, M.; Perse, M.; Majdic, G.; Fink, M.; Rozman, D. Determination of reference genes for circadian studies in different tissues and mouse strains. BMC Mol. Biol. 2010, 11, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozman, D. From nonalcoholic fatty liver disease to hepatocellular carcinoma: A systems understanding. Dig. Dis. Sci. 2014, 59, 238–241. [Google Scholar] [CrossRef] [Green Version]
- Sato, J.; Kanazawa, A.; Ikeda, F.; Yoshihara, T.; Goto, H.; Abe, H.; Komiya, K.; Kawaguchi, M.; Shimizu, T.; Ogihara, T.; et al. Gut dysbiosis and detection of “Live gut bacteria” in blood of Japanese patients with type 2 diabetes. Diabetes Care 2014, 37, 2343–2350. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, S.A.; deAndrade, D.R.; deAndrade, D.R. Rat hepatocyte invasion by Listeria monocytogenes and analysis of TNF-α role in apoptosis. Rev. Inst. Med. Trop. Sao Paulo 2005, 47, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camilleri, M. Leaky gut: Mechanisms, measurement and clinical implications in humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef]
- Micó-Carnero, M.; Rojano-Alfonso, C.; Álvarez-Mercado, A.I.; Gracia-Sancho, J.; Casillas-Ramírez, A.; Peralta, C. Effects of gut metabolites and microbiota in healthy and marginal livers submitted to surgery. Int. J. Mol. Sci. 2021, 22, 44. [Google Scholar] [CrossRef] [PubMed]
- Rowell, D.L.; Eckmann, L.; Dwinell, M.B.; Carpenter, S.P.; Raucy, J.L.; Yang, S.K.; Kagnoff, M.F. Human hepatocytes express an array of proinflammatory cytokines after agonist stimulation or bacterial invasion. Am. J. Physiol. Gastrointest. Liver Physiol. 1997, 273. [Google Scholar] [CrossRef]
- Tan, X.; Li, L.; Wang, J.; Zhao, B.; Pan, J.; Wang, L.; Liu, X.; Liu, X.; Liu, Z. Resveratrol prevents acrylamide-induced mitochondrial dysfunction and inflammatory responses via targeting circadian regulator Bmal1 and Cry1 in hepatocytes. J. Agric. Food Chem. 2019, 67, 8510–8519. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Guo, R.; Tian, H.; Li, L.; Liu, H.; Mi, Y.; Liu, X. Nobiletin protects against insulin resistance and disorders of lipid metabolism by reprogramming of circadian clock in hepatocytes. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 549–562. [Google Scholar] [CrossRef]
- Chen, L.; Li, S.; Nie, J.; Zhao, J.; Yu, S.; Li, Y.; Peng, J. Bmal1 regulates coagulation factor biosynthesis in mouse liver in Streptococcus oralis infection. Front. Cell. Infect. Microbiol. 2020, 10, 530190. [Google Scholar] [CrossRef] [PubMed]
- Nader, N.; Chrousos, G.P.; Kino, T. Circadian rhythm transcription factor CLOCK regulates the transcriptional activity of the glucocorticoid receptor by acetylating its hinge region lysine cluster: Potential physiological implications. FASEB J. 2009, 23, 1572–1583. [Google Scholar] [CrossRef] [Green Version]
- Chauvet, C.; Vanhoutteghem, A.; Duhem, C.; Saint-Auret, G.; Bois-Joyeux, B.; Djian, P.; Staels, B.; Danan, J.-L. Control of gene expression by the retinoic acid-related orphan receptor Alpha in HepG2 human hepatoma cells. PLoS ONE 2011, 6, e22545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Xu, M.J.; Gao, B. Hepatocytes: A key cell type for innate immunity. Cell. Mol. Immunol. 2016, 13, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Chiu, Y.H.; Lin, S.L.; Ou, C.C.; Lu, Y.C.; Huang, H.Y.; Lin, M.Y. Anti-inflammatory effect of lactobacilli bacteria on HepG2 cells is through cross-regulation of TLR4 and NOD2 signalling. J. Funct. Foods 2013, 5, 820–828. [Google Scholar] [CrossRef]
- Starčič Erjavec, M.; Jesenko, B.; Petkovšek, Ž.; Žgur-Bertok, D. Prevalence and associations of tcpC, a gene encoding a Toll/interleukin-1 receptor domain-containing protein, among Escherichia coli urinary tract infection, skin and soft tissue infection, and commensal isolates. J. Clin. Microbiol. 2010, 48, 966–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starčič Erjavec, M.; Petkovšek, Ž.; Predojević, L.; Žgur-Bertok, D. Iron uptake and bacteriocin genes among Escherichia coli strains from skin and soft-tissue infections. Apmis 2017, 125, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, G. Cosinor-based rhythmometry. Theor. Biol. Med. Model. 2014, 11, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bingham, C.; Arbogast, B.; Guillaume, G.C.; Lee, J.K.; Halberg, F. Inferential statistical methods for estimating and comparing cosinor parameters. Chronobiologia 1982, 9, 397–439. [Google Scholar]
- Moškon, M. CosinorPy: A python package for cosinor-based rhythmometry. BMC Bioinformatics 2020, 21, 485. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Tognini, P. The circadian clock as an essential molecular link between host physiology and microorganisms. Front. Cell. Infect. Microbiol. 2020, 9, 469. [Google Scholar] [CrossRef] [Green Version]
- Schmalle, V.; Lorentz, A. Role of the microbiota in circadian rhythms of the host. Chronobiol. Int. 2020, 37, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Shao, W.; Li, S.; Ma, L.; Zheng, L.; Shang, W.; Jia, X.; Sun, P.; Liang, X.; Jia, J. H. pylori infection induced BMAL1 expression and rhythm disorder aggravate gastric inflammation. EBioMedicine. 2019, 39, 301–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, K.; Yano, M.; Doki, Y.; Azama, T.; Iwanaga, H.; Miki, H.; Nakayama, M.; Miyata, H.; Takiguchi, S.; Fujiwara, Y.; et al. Injection of LPS causes transient suppression of biological clock genes in rats. J. Surg. Res. 2008, 145, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Watanabe, K.; Anayama, N.; Miyazaki, K. Effect of lipopolysaccharide on circadian clock genes Per2 and Bmal1 in mouse ovary. J. Physiol. Sci. 2017, 67, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Fuller, K.K.; Dunlap, J.C.; Loros, J.J. A pro- and anti-inflammatory axis modulates the macrophage circadian clock. Front. Immunol. 2020, 11, 867. [Google Scholar] [CrossRef]
- Lee, H.; Nah, S.S.; Chang, S.H.; Kim, H.K.; Kwon, J.T.; Lee, S.; Cho, I.H.; Lee, S.W.; Kim, Y.O.; Hong, S.J.; et al. PER2 is downregulated by the LPS-induced inflammatory response in synoviocytes in rheumatoid arthritis and is implicated in disease susceptibility. Mol. Med. Rep. 2017, 16, 422–428. [Google Scholar] [CrossRef]
- Yamamura, Y.; Yano, I.; Kudo, T.; Shibata, S. Time-dependent inhibitory effect of lipopolysaccharide injection on Per1 and Per2 gene expression in the mouse heart and liver. Chronobiol. Int. 2010, 27, 213–232. [Google Scholar] [CrossRef]
- Mossman, K.L.; Mian, M.F.; Lauzon, N.M.; Gyles, C.L.; Lichty, B.; Mackenzie, R.; Gill, N.; Ashkar, A.A. Cutting edge: FimH adhesin of type 1 fimbriae is a novel TLR4 ligand. J. Immunol. 2008, 181, 6702–6706. [Google Scholar] [CrossRef]
- Cavadini, G.; Petrzilka, S.; Kohler, P.; Jud, C.; Tobler, I.; Birchler, T.; Fontana, A. TNF-α suppresses the expression of clock genes by interfering with E-box-mediated transcription. Proc. Natl. Acad. Sci. USA 2007, 104, 12843–12848. [Google Scholar] [CrossRef] [Green Version]
- Coogan, A.N.; Wyse, C.A. Neuroimmunology of the circadian clock. Brain Res. 2008, 1232, 104–112. [Google Scholar] [CrossRef]
- dos Santos, S.A.; de Andrade Júnior, D.R.; de Andrade, D.R. TNF-α production and apoptosis in hepatocytes after Listeria monocytogenes and Salmonella typhimurium invasion. Rev. Inst. Med. Trop. Sao Paulo 2011, 53, 107–112. [Google Scholar] [CrossRef]
- Starčič Erjavec, M.; Žgur-Bertok, D. Virulence potential for extraintestinal infections among commensal Escherichia coli isolated from healthy humans-the Trojan horse within our gut. FEMS Microbiol. Lett. 2015, 362, fnu061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, S.; Honeywell, R.; Geletu, M.; Arulanandam, R.; Raptis, L. Housekeeping genes; expression levels may change with density of cultured cells. J. Immunol. Methods 2010, 355, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Baddela, V.S.; Baufeld, A.; Yenuganti, V.R.; Vanselow, J.; Singh, D. Suitable housekeeping genes for normalization of transcript abundance analysis by real-time RT-PCR in cultured bovine granulosa cells during hypoxia and differential cell plating density. Reprod. Biol. Endocrinol. 2014, 12, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, G.; Yoshida, K.; Narahara, S.; Uchikawa, M.; Kawamura, M.; Yamauchi, N.; Xi, Y.; Shigeyoshi, Y.; Hashimoto, S.; Hattori, M.A. Alterations of circadian clockworks during differentiation and apoptosis of rat ovarian cells. Chronobiol. Int. 2011, 28, 477–487. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovač, U.; Žužek, Z.; Raspor Dall’Olio, L.; Pohar, K.; Ihan, A.; Moškon, M.; Rozman, D.; Starčič Erjavec, M. Escherichia coli Affects Expression of Circadian Clock Genes in Human Hepatoma Cells. Microorganisms 2021, 9, 869. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040869
Kovač U, Žužek Z, Raspor Dall’Olio L, Pohar K, Ihan A, Moškon M, Rozman D, Starčič Erjavec M. Escherichia coli Affects Expression of Circadian Clock Genes in Human Hepatoma Cells. Microorganisms. 2021; 9(4):869. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040869
Chicago/Turabian StyleKovač, Urša, Zala Žužek, Lucija Raspor Dall’Olio, Katka Pohar, Alojz Ihan, Miha Moškon, Damjana Rozman, and Marjanca Starčič Erjavec. 2021. "Escherichia coli Affects Expression of Circadian Clock Genes in Human Hepatoma Cells" Microorganisms 9, no. 4: 869. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9040869