
Bovine Milk Oligosaccharides and Human Milk Oligosaccharides Modulate the Gut Microbiota Composition and Volatile Fatty Acid Concentrations in a Preclinical Neonatal Model

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Care and Dietary Treatments
2.2. Sample Collection
2.3. Dry Matter, pH, and VFA Concentrations
2.4. DNA Extraction, PCR Amplification and Sequencing of 16S rRNA Genes
2.5. Sequence Processing
2.6. Real-Time PCR
2.7. Statistics
3. Results
3.1. DM, pH, and VFA Concentrations
3.2. Sequencing of 16S rRNA Amplicons


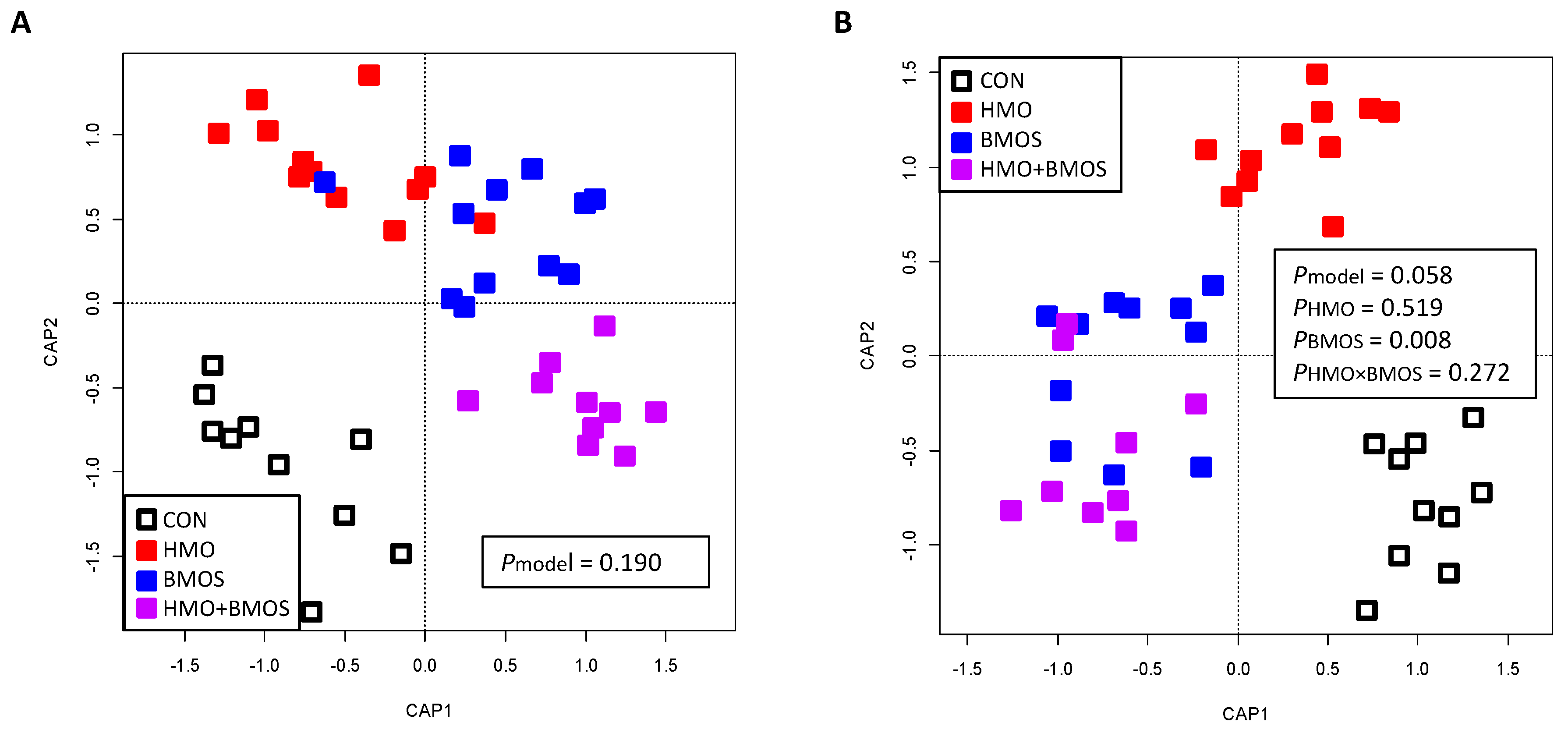{kind=link}
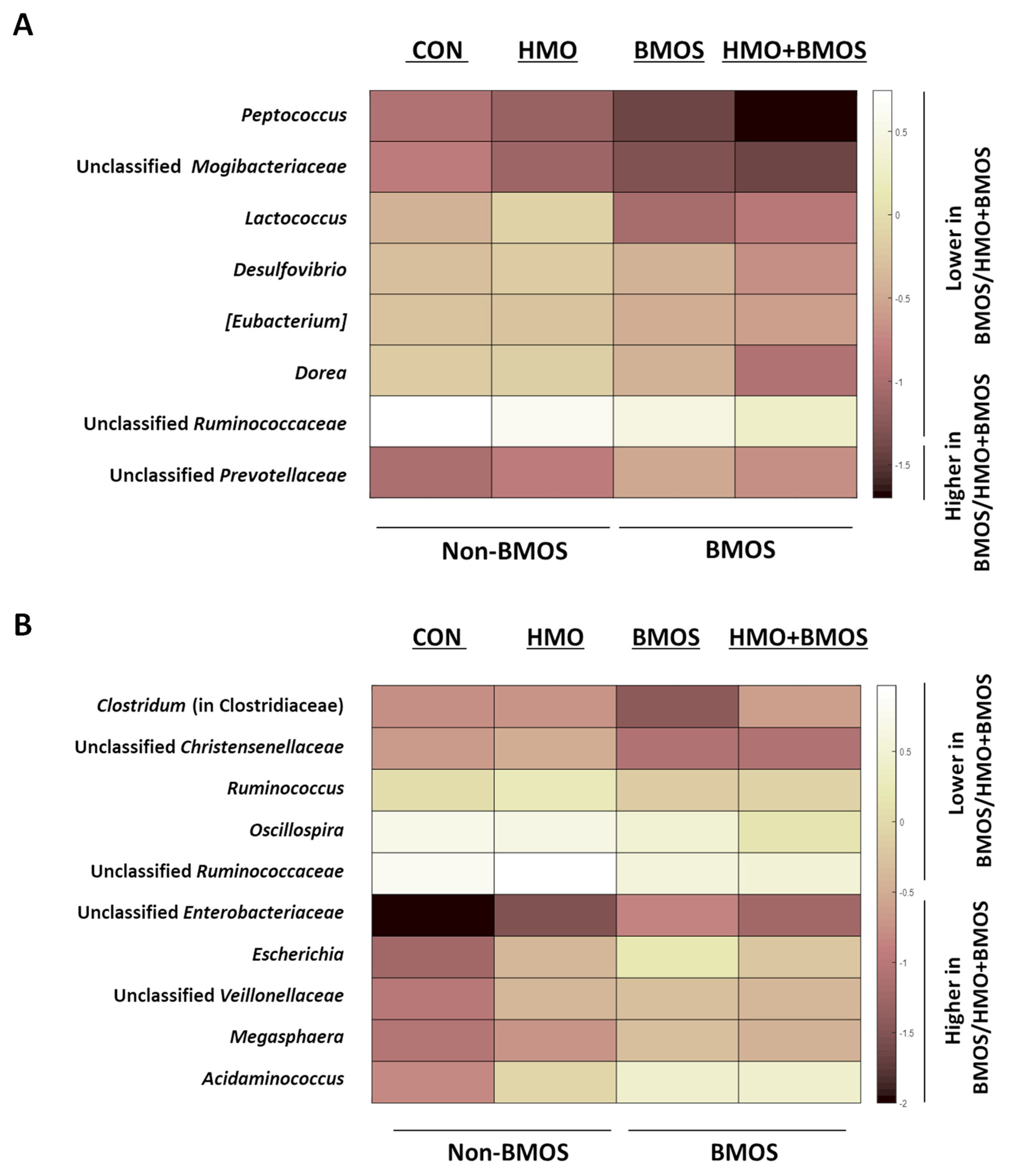{kind=link}
{kind=link}
{kind=link}
| CON (n = 10) | HMO (n = 10–11) | BMOS (n = 8–11) | HMO + BMOS (n = 6–8) | p-Values | |||
|---|---|---|---|---|---|---|---|
| HMO | BMOS | HMO × BMOS | |||||
| Ascending colon | |||||||
| pH | 6.67 ± 0.16 | 6.86 ± 0.08 | 6.80 ± 0.10 | 6.51 ± 0.18 | 0.7063 | 0.4082 | 0.0558 |
| DM (%) | 20.1 ± 1.82 | 19.5 ± 1.69 | 16.6 ± 1.17 | 16.3 ± 1.31 | 0.3431 | 0.0025 | 0.4405 |
| Acetate (μmol/g of DM) | 439.6 ± 99.8 | 423.3 ± 98.2 | 511.4 ± 90.2 | 512.4 ± 113.5 | 0.3873 | 0.0473 | 0.4746 |
| Propionate (μmol/g of DM) | 138.1 ± 31.4 | 125.2 ± 30.5 | 128.4 ± 21.2 | 128.0 ± 24.2 | 0.5545 | 0.5030 | 0.6591 |
| Butyrate (μmol/g of DM) | 98.4 ± 28.5 | 59.2 ± 9.79 | 71.3 ± 12.2 | 69.7 ± 14.9 | 0.5667 | 0.9412 | 0.8236 |
| Isobutyrate (μmol/g of DM) | 9.34 ± 1.52 | 10.1 ± 2.22 | 9.30 ± 1.23 | 8.91 ± 1.29 | 0.4518 | 0.8059 | 0.2631 |
| Valerate (μmol/g of DM) | 11.5 ± 1.66 | 12.3 ± 2.45 | 10.8 ± 1.04 | 10.3 ± 0.82 | 0.6290 | 0.1346 | 0.1890 |
| Isovalerate (μmol/g of DM) | 17.8 ± 3.95 | 16.4 ± 3.25 | 16.2 ± 2.13 | 17.2 ± 3.16 | 0.5977 | 0.5838 | 0.4524 |
| Feces | |||||||
| pH | 6.89 ± 0.12 | 6.99 ± 0.09 | 7.13 ± 0.08 | 7.07 ± 0.07 | 0.7818 | 0.0587 | 0.3066 |
| DM (%) | 32.3 ± 2.56 | 36.5 ± 1.97 | 28.7 ± 2.13 | 27.9 ± 3.30 | 0.4698 | 0.0136 | 0.3040 |
| Acetate (μmol/g of DM) | 146.1 ± 21.0 a | 95.4 ± 9.59 b | 132.6 ± 19.5 ab | 180.8 ± 43.5 a | 0.5653 | 0.1044 | 0.0394 |
| Propionate (μmol/g of DM) | 42.4 ± 5.60 | 29.9 ± 4.10 | 40.2 ± 7.17 | 49.5 ± 10.3 | 0.7676 | 0.1521 | 0.0831 |
| Butyrate (μmol/g of DM) | 20.0 ± 2.62 ab | 13.0 ± 2.46 b | 14.9 ± 3.07 ab | 36.7 ± 19.0 a | 0.9446 | 0.3624 | 0.0291 |
| Isobutyrate (μmol/g of DM) | 5.81 ± 1.01 | 4.85 ± 0.77 | 5.61 ± 0.65 | 6.24 ± 1.30 | 0.7980 | 0.4364 | 0.3902 |
| Valerate (μmol/g of DM) | 8.49 ± 1.85 | 7.20 ± 1.41 | 7.70 ± 0.85 | 9.31 ± 2.30 | 0.8394 | 0.3726 | 0.4515 |
| Isovalerate (μmol/g of DM) | 5.57 ± 0.93 | 4.67 ± 0.98 | 5.32 ± 0.93 | 6.66 ± 1.38 | 0.8564 | 0.1345 | 0.2303 |
| Phylum | CON (n = 10) | HMO (n = 12) | BMOS (n = 12) | HMO + BMOS (n = 10) | p-Values | ||
|---|---|---|---|---|---|---|---|
| HMO | BMOS | HMO × BMOS | |||||
| Actinobacteria | 0.81 ± 0.23 | 1.84 ± 0.89 | 0.84 ± 0.37 | 0.42 ± 0.15 | 0.7297 | 0.1451 | 0.2349 |
| Bacteroidetes | 42.8 ± 8.73 | 37.8 ± 6.88 | 47.7 ± 5.73 | 55.5 ± 5.73 | 0.7864 | 0.0432 | 0.3107 |
| Cyanobacteria | 0.01 ± 0.01 | 0.31 ± 0.30 | 0 ± 0 | 0 ± 0 | 0.3223 | 0.2774 | 0.3181 |
| Deferribacteres | 0.06 ± 0.04 a | 0.01 ± 0.01 b | 0.01 ± 0 b | 0.04 ± 0.03 ab | 0.2511 | 0.394 | 0.0158 |
| Elusimicrobia | 1.62 ± 0.66 | 1.10 ± 0.58 | 2.04 ± 1.21 | 3.14 ± 2.24 | 0.9516 | 0.6119 | 0.5118 |
| Firmicutes | 46.4 ± 8.70 | 53.1 ± 6.89 | 50.0 ± 5.51 | 36.4 ± 5.90 | 0.8138 | 0.1056 | 0.2419 |
| Fusobacteria | 2.58 ± 2.38 | 0.03 ± 0.02 | 0.04 ± 0.02 | 0.08 ± 0.07 | 0.1553 | 0.1809 | 0.1444 |
| Lentisphaerae | 0.01 ± 0.01 | 0 ± 0 | 0.01 ± 0 | 0.01 ± 0 | 0.1712 | 0.5105 | 0.2735 |
| Proteobacteria | 1.93 ± 0.53 | 2.08 ± 0.41 | 1.67 ± 0.41 | 2.37 ± 1.32 | 0.6862 | 0.6198 | 0.9967 |
| Synergistetes | 0.05 ± 0.03 | 0.09 ± 0.07 | 0.03 ± 0.02 | 0.04 ± 0.02 | 0.5248 | 0.416 | 0.9753 |
| Tenericutes | 0.38 ± 0.22 | 0.07 ± 0.02 | 0.11 ± 0.09 | 0.02 ± 0.02 | 0.0170 | 0.0032 | 0.3194 |
| Verrucomicrobia | 0.07 ± 0.05 | 1.04 ± 1.03 | 0.26 ± 0.17 | 0.02 ± 0.01 | 0.9169 | 0.8424 | 0.3211 |
| Unclassified | 1.36 ± 0.17 | 1.30 ± 0.13 | 1.07 ± 0.06 | 1.09 ± 0.11 | 0.8083 | 0.0266 | 0.7509 |

| Phylum | CON (n = 10) | HMO (n = 11) | BMOS (n = 11) | HMO + BMOS (n = 9) | p-Values | ||
|---|---|---|---|---|---|---|---|
| HMO | BMOS | HMO × BMOS | |||||
| Actinobacteria | 0.92 ± 0.32 | 2.56 ± 0.93 | 1.10 ± 0.50 | 1.06 ± 0.45 | 0.1788 | 0.2220 | 0.1324 |
| Bacteroidetes | 49.5 ± 5.73 ab | 39.6 ± 5.47 b | 44.2 ± 4.0 ab | 54.11 ± 3.60 a | 0.9400 | 0.2825 | 0.0317 |
| Cyanobacteria | 0.01 ± 0.01 | 0.04 ± 0.04 | 0 ± 0 | 0 ± 0 | 0.2377 | 0.1500 | 0.4995 |
| Deferribacteres | 0.02 ± 0.01 | 0 ± 0 | 0.02 ± 0.01 | 0 ± 0 | 0.1123 | 0.9305 | 0.8006 |
| Elusimicrobia | 4.02 ± 2.31 | 6.58 ± 3.19 | 4.13 ± 1.69 | 5.93 ± 2.59 | 0.5138 | 0.8268 | 0.8141 |
| Firmicutes | 37.1 ± 4.43 | 44.1 ± 5.69 | 41.3 ± 4.88 | 33.6 ± 3.40 | 0.8313 | 0.3986 | 0.0711 |
| Fusobacteria | 2.38 ± 2.00 | 0.60 ± 0.38 | 1.03 ± 0.75 | 0.11 ± 0.05 | 0.1906 | 0.2594 | 0.8083 |
| Lentisphaerae | 0.09 ± 0.06 | 0.08 ± 0.03 | 0.04 ± 0.02 | 0.02 ± 0.01 | 0.8342 | 0.0778 | 0.5231 |
| Proteobacteria | 2.39 ± 0.94 | 1.73 ± 0.35 | 3.54 ± 0.96 | 1.77 ± 0.44 | 0.1124 | 0.3798 | 0.3321 |
| Synergistetes | 0.45 ± 0.19 | 1.31 ± 0.87 | 0.34 ± 0.12 | 0.76 ± 0.47 | 0.2246 | 0.3505 | 0.5413 |
| Tenericutes | 0.26 ± 0.16 | 0.03 ± 0.01 | 0.05 ± 0.03 | 0.02 ± 0.01 | 0.0313 | 0.0292 | 0.0789 |
| Verrucomicrobia | 1.06 ± 0.50 | 1.51 ± 1.35 | 2.02 ± 1.16 | 0.49 ± 0.36 | 0.3031 | 0.9871 | 0.5431 |
| Unclassified | 1.50 ± 0.18 | 1.47 ± 0.13 | 1.83 ± 0.17 | 1.46 ± 0.16 | 0.1817 | 0.3248 | 0.2708 |

3.3. RT PCR
| CON (n = 10) | HMO (n = 11–12) | BMOS (n = 11–12) | HMO + BMOS (n = 9–10) | p-Values | |||
|---|---|---|---|---|---|---|---|
| HMO | BMOS | HMO × BMOS | |||||
| Ascending colon | |||||||
| Observed OTUs | 2123.2 ± 124.1 | 2014.2 ± 100.6 | 1922.3 ± 105.6 | 1777.0 ± 84.0 | 0.1358 | 0.0148 | 0.8187 |
| Shannon | 7.24 ± 0.37 | 7.10 ± 0.25 | 6.96 ± 0.31 | 6.54 ± 0.25 | 0.2829 | 0.1172 | 0.5843 |
| Simpson reciprocal | 27.12 ± 7.90 | 18.47 ± 2.48 | 20.74 ± 2.97 | 14.98 ± 2.69 | 0.2674 | 0.4494 | 0.6410 |
| Chao1 | 5527.6 ± 310.8 | 5003.3 ± 227.5 | 5019.3 ± 314.6 | 4521.6 ± 179.8 | 0.0275 | 0.0321 | 0.9991 |
| Faith PD | 153.98 ± 8.00 | 150.70 ± 7.18 | 141.99 ± 5.47 | 132.47 ± 6.45 | 0.2075 | 0.0066 | 0.5543 |
| Feces | |||||||
| Observed OTUs | 2230.3 ± 77.0 | 2187.3 ± 119.6 | 2354.7 ± 73.3 | 2048.7 ± 149.8 | 0.0793 | 0.9205 | 0.2156 |
| Shannon | 7.66 ± 0.17 | 7.72 ± 0.20 | 7.96 ± 0.19 | 7.36 ± 0.32 | 0.2037 | 0.8925 | 0.1217 |
| Simpson reciprocal | 24.68 ± 2.49 b | 31.74 ± 4.19 ab | 37.47 ± 4.70 a | 27.35 ± 4.94 ab | 0.7041 | 0.2999 | 0.0377 |
| Chao1 | 6051.6 ± 248.6 | 5841.7 ± 389.1 | 6315.6 ± 245.9 | 5527.4 ± 406.6 | 0.0948 | 0.9096 | 0.3951 |
| Faith PD | 165.19 ± 4.69 | 166.45 ± 7.48 | 174.55 ± 5.58 | 152.40 ± 9.73 | 0.0903 | 0.6571 | 0.0832 |
| CON (n = 10) | HMO (n = 11–12) | BMOS (n = 11–12) | HMO + BMOS (n = 9–10) | p-Values | |||
|---|---|---|---|---|---|---|---|
| HMO | BMOS | HMO × BMOS | |||||
| Log10 16S rRNA gene copies/g of AC contents or feces | |||||||
| Ascending colon | |||||||
| Total bacteria | 11.0 ± 0.07 | 11.1 ± 0.07 | 11.0 ± 0.05 | 11.2 ± 0.09 | 0.1612 | 0.2765 | 0.5490 |
| Enterobacteriaceae | 8.82 ± 0.28 | 8.91 ± 0.32 | 9.38 ± 0.18 | 9.12 ± 0.30 | 0.7559 | 0.1306 | 0.4818 |
| Bifidobacterium spp. | 6.73 ± 0.27 | 7.07 ± 0.17 | 7.23 ± 0.18 | 7.02 ± 0.15 | 0.5418 | 0.1384 | 0.1283 |
| Lactobacillus spp. | 9.69 ± 0.15 | 9.64 ± 0.12 | 9.54 ± 0.18 | 9.81 ± 0.17 | 0.4833 | 0.9444 | 0.1796 |
| Prevotella spp. | 10.6 ± 0.25 | 10.6 ± 0.20 | 10.9 ± 0.20 | 11.1 ± 0.16 | 0.5011 | 0.0210 | 0.4291 |
| Bacteroides fragilis | 6.57 ± 0.52 | 6.53 ± 0.38 | 6.40 ± 0.32 | 6.31 ± 0.31 | 0.7790 | 0.5282 | 0.9274 |
| Clostridium difficile | 4.89 ± 0.07 | 4.88 ± 0.08 | 4.92 ± 0.13 | 5.14 ± 0.20 | 0.4131 | 0.2323 | 0.3541 |
| Clostridium perfringens | 6.81 ± 0.31 | 6.47 ± 0.29 | 6.32 ± 0.30 | 6.10 ± 0.40 | 0.3708 | 0.1840 | 0.8425 |
| Escherichia coli | 8.14 ± 0.32 | 8.29 ± 0.35 | 8.80 ± 0.20 | 8.45 ± 0.32 | 0.7617 | 0.1365 | 0.3564 |
| Feces | |||||||
| Total bacteria | 10.4 ± 0.17 | 10.3 ± 0.24 | 10.6 ± 0.10 | 10.6 ± 0.11 | 0.8045 | 0.2313 | 0.4963 |
| Enterobacteriaceae | 8.31 ± 0.16 | 8.51 ± 0.40 | 9.42 ± 0.24 | 8.97 ± 0.25 | 0.6544 | 0.0039 | 0.2356 |
| Bifidobacterium spp. | 6.02 ± 0.20 | 6.47 ± 0.24 | 6.46 ± 0.26 | 6.62 ± 0.24 | 0.1849 | 0.2021 | 0.5273 |
| Lactobacillus spp. | 8.98 ± 0.17 | 9.51 ± 0.31 | 9.48 ± 0.15 | 9.13 ± 0.30 | 0.5456 | 0.1502 | 0.1827 |
| Prevotella spp. | 10.4 ± 0.22 | 10.3 ± 0.16 | 10.6 ± 0.16 | 10.6 ± 0.15 | 0.8392 | 0.2374 | 0.6492 |
| Bacteroides fragilis | 6.45 ± 0.45 | 6.38 ± 0.35 | 5.99 ± 0.31 | 5.92 ± 0.36 | 0.8413 | 0.2076 | 0.9962 |
| Clostridium difficile | 4.91 ± 0.08 | 4.89 ± 0.09 | 5.09 ± 0.17 | 4.88 ± 0.06 | 0.2542 | 0.3929 | 0.3663 |
| Clostridium perfringens | 5.82 ± 0.39 | 5.93 ± 0.44 | 5.74 ± 0.36 | 5.18 ± 0.20 | 0.5237 | 0.2476 | 0.3579 |
| Escherichia coli | 7.51 ± 0.16 | 7.69 ± 0.46 | 8.75 ± 0.26 | 8.26 ± 0.27 | 0.6217 | 0.0035 | 0.2863 |

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Huërou-Luron, I.; Blat, S.; Boudry, G. Breast-v. formula-feeding: Impacts on the digestive tract and immediate and long-term health effects. Nutr. Res. Rev. 2010, 23, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Guaraldi, F.; Salvatori, G. Effect of Breast and Formula Feeding on Gut Microbiota Shaping in Newborns. Front. Cell. Infect. Microbiol. 2012, 2, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Academy of Pediatrics, Section on Breastfeeding. Breastfeeding and the Use of Human Milk. Pediatrics 2012, 129, e827–e841. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Breastfeeding Report Card–United States. 2018. Available online: https://www.cdc.gov/breastfeeding/pdf/2018breastfeedingreportcard.pdf (accessed on 18 October 2020).
- Hernell, O. Human milk vs. cow’s milk and the evolution of infant formulas. Milk Milk Prod. Hum. Nutr. 2011, 67, 17–28. [Google Scholar]
- Ahern, G.J.; Hennessy, A.A.; Ryan, C.A.; Ross, R.P.; Stanton, C. Advances in Infant Formula Science. Annu. Rev. Food Sci. Technol. 2019, 10, 75–102. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [Green Version]
- Gnoth, M.J.; Kunz, C.; Kinne-Saffran, E.; Rudloff, S. Human Milk Oligosaccharides Are Minimally Digested In Vitro. J. Nutr. 2000, 130, 3014–3020. [Google Scholar] [CrossRef]
- Ninonuevo, M.R.; Park, Y.; Yin, H.; Zhang, J.; Ward, R.E.; Clowers, B.H.; German, J.B.; Freeman, S.L.; Killeen, K.; Grimm, R.; et al. A Strategy for Annotating the Human Milk Glycome. J. Agric. Food Chem. 2006, 54, 7471–7480. [Google Scholar] [CrossRef]
- Robinson, R.C. Structures and Metabolic Properties of Bovine Milk Oligosaccharides and Their Potential in the Development of Novel Therapeutics. Front. Nutr. 2019, 6, 50. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; Lebrilla, C.B.; Mills, D.A.; German, J.B.; Freeman, S.L. Breast Milk Oligosaccharides: Structure-Function Relationships in the Neonate. Annu. Rev. Nutr. 2014, 34, 143–169. [Google Scholar] [CrossRef] [Green Version]
- Kuntz, S.; Rudloff, S.; Kunz, C. Oligosaccharides from human milk influence growth-related characteristics of intestinally transformed and non-transformed intestinal cells. Br. J. Nutr. 2008, 99, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Holscher, H.D.; Bode, L.; Tappenden, K.A. Human Milk Oligosaccharides Influence Intestinal Epithelial Cell Maturation In Vitro. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 296–301. [Google Scholar] [CrossRef]
- Lane, J.A.; O’Callaghan, J.; Carrington, S.D.; Hickey, R.M. Transcriptional response of HT-29 intestinal epithelial cells to human and bovine milk oligosaccharides. Br. J. Nutr. 2013, 110, 2127–2137. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; Elderman, M.; Cheng, L.; de Haan, B.J.; Nauta, A.; de Vos, P. Modulation of Intestinal Epithelial Glycocalyx Development by Human Milk Oligosaccharides and Non-Digestible Carbohydrates. Mol. Nutr. Food Res. 2019, 63, e1900303. [Google Scholar] [CrossRef] [PubMed]
- Natividad, J.M.; Rytz, A.; Keddani, S.; Bergonzelli, G.; Garcia-Rodenas, C.L. Blends of Human Milk Oligosaccharides Confer Intestinal Epithelial Barrier Protection in Vitro. Nutrients 2020, 12, 3047. [Google Scholar] [CrossRef]
- Duncan, P.I.; Aitio, O.; Heiskanen, A.; Niemelä, R.; Saarinen, J.; Helin, J.; Porta, N.; Fiaux, M.; Moënnoz, D.; Golliard, M.; et al. Structure and Function of Bovine Whey Derived Oligosaccharides Showing Synbiotic Epithelial Barrier Protective Properties. Nutrients 2020, 12, 2007. [Google Scholar] [CrossRef] [PubMed]
- Comstock, S.S.; Wang, M.; Hester, S.N.; Li, M.; Donovan, S.M. Select human milk oligosaccharides directly modulate peripheral blood mononuclear cells isolated from 10-d-old pigs. Br. J. Nutr. 2014, 111, 819–828. [Google Scholar] [CrossRef] [Green Version]
- Goehring, K.C.; Marriage, B.J.; Oliver, J.S.; Wilder, J.A.; Barrett, E.G.; Buck, R.H. Similar to Those Who Are Breastfed, Infants Fed a Formula Containing 2′-Fucosyllactose Have Lower Inflammatory Cytokines in a Randomized Controlled Trial. J. Nutr. 2016, 146, 2559–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudd, A.T.; Fleming, S.A.; Labhart, B.; Chichlowski, M.; Berg, B.M.; Donovan, S.M.; Dilger, R.N. Dietary Sialyllactose Influences Sialic Acid Concentrations in the Prefrontal Cortex and Magnetic Resonance Imaging Measures in Corpus Callosum of Young Pigs. Nutrients 2017, 9, 1297. [Google Scholar] [CrossRef] [Green Version]
- Fleming, S.A.; Mudd, A.T.; Hauser, J.; Yan, J.; Metairon, S.; Steiner, P.; Donovan, S.M.; Dilger, R.N. Dietary Oligofructose Alone or in Combination with 2′-Fucosyllactose Differentially Improves Recognition Memory and Hippocampal mRNA Expression. Nutrients 2020, 12, 2131. [Google Scholar] [CrossRef]
- Hegar, B.; Wibowo, Y.; Basrowi, R.W.; Ranuh, R.G.; Sudarmo, S.M.; Munasir, Z.; Atthiyah, A.F.; Widodo, A.D.; Supriatmo; Kadim, M.; et al. The Role of Two Human Milk Oligosaccharides, 2′-Fucosyllactose and Lacto-N-Neotetraose, in Infant Nutrition. Pediatr. Gastroenterol. Hepatol. Nutr. 2019, 22, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, M.; Wu, S.; Lebrilla, C.B.; Chapkin, R.S.; Ivanov, I.; Donovan, S.M. Fecal Microbiota Composition of Breast-Fed Infants Is Correlated With Human Milk Oligosaccharides Consumed. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 825–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcobal, A.; Barboza, M.; Froehlich, J.W.; Block, D.E.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Consumption of Human Milk Oligosaccharides by Gut-Related Microbes. J. Agric. Food Chem. 2010, 58, 5334–5340. [Google Scholar] [CrossRef] [Green Version]
- Newburg, D.S.; Ruiz-Palacios, G.M.; Morrow, A.L. Human milk glycans protect infants against enteric pathogens. Annu. Rev. Nutr. 2005, 25, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.L.; Ruiz-Palacios, G.M.; Jiang, X.; Newburg, D.S. Human-Milk Glycans That Inhibit Pathogen Binding Protect Breast-feeding Infants against Infectious Diarrhea. J. Nutr. 2005, 135, 1304–1307. [Google Scholar] [CrossRef] [Green Version]
- Wiciński, M.; Sawicka, E.; Gębalski, J.; Kubiak, K.; Malinowski, B. Human Milk Oligosaccharides: Health Benefits, Potential Applications in Infant Formulas, and Pharmacology. Nutrients 2020, 12, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puccio, G.; Alliet, P.; Cajozzo, C.; Janssens, E.; Corsello, G.; Sprenger, N.; Wernimont, S.; Egli, D.; Gosoniu, L.; Steenhout, P. Effects of Infant Formula With Human Milk Oligosaccharides on Growth and Morbidity: A randomized multicenter trial. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 624–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, B.; Porta, N.; Foata, F.; Grathwohl, D.; Delley, M.; Moine, D.; Charpagne, A.; Siegwald, L.; Descombes, P.; Alliet, P.; et al. Linking Human Milk Oligosaccharides, Infant Fecal Community Types, and Later Risk To Require Antibiotics. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Escalante, E.; Alatorre-Santamaría, S.; Castañeda-Ovando, A.; Salazar-Pereda, V.; Bautista-Ávila, M.; Cruz-Guerrero, A.E.; Flores-Aguilar, J.F.; González-Olivares, L.G. Human milk oligosaccharides as bioactive compounds in infant formula: Recent advances and trends in synthetic methods. Crit. Rev. Food Sci. Nutr. 2020, 1–34. [Google Scholar] [CrossRef]
- Barile, D.; Tao, N.; Lebrilla, C.B.; Coisson, J.-D.; Arlorio, M.; German, J.B. Permeate from cheese whey ultrafiltration is a source of milk oligosaccharides. Int. Dairy J. 2009, 19, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Yang, H.; Li, J.; Li, Y.; Ding, X.; Wu, G.; Yin, Y. Pig models on intestinal development and therapeutics. Amino Acids 2017, 49, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Mudd, A.T.; Dilger, R.N. Early-Life Nutrition and Neurodevelopment: Use of the Piglet as a Translational Model. Adv. Nutr. 2017, 8, 92–104. [Google Scholar] [CrossRef] [Green Version]
- Fleming, S.A.; Mudd, A.T.; Hauser, J.; Yan, J.; Metairon, S.; Steiner, P.; Donovan, S.M.; Dilger, R.N. Human and Bovine Milk Oligosaccharides Elicit Improved Recognition Memory Concurrent With Alterations in Regional Brain Volumes and Hippocampal mRNA Expression. Front. Neurosci. 2020, 14, 770. [Google Scholar] [CrossRef]
- Wang, M.; Radlowski, E.C.; Monaco, M.H.; Fahey, G.C., Jr.; Gaskins, H.R.; Donovan, S.M. Mode of delivery and early nutrition modulate microbial colonization and fermentation products in neonatal piglets. J. Nutr. 2013, 143, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Reznikov, E.A.; Comstock, S.S.; Hoeflinger, J.L.; Wang, M.; Miller, M.J.; Donovan, S.M. Dietary Bovine Lactoferrin Reduces Staphylococcus aureus in the Tissues and Modulates the Immune Response in Piglets Systemically Infected with S. aureus. Curr. Dev. Nutr. 2017, 2, nzy001. [Google Scholar] [CrossRef] [Green Version]
- Monaco, M.H.; Wang, M.; Pan, X.; Li, Q.; Richards, J.D.; Chichlowski, M.; Berg, B.M.; Dilger, R.N.; Donovan, S.M. Evaluation of Sialyllactose Supplementation of a Prebiotic-Containing Formula on Growth, Intestinal Development, and Bacterial Colonization in the Neonatal Piglet. Curr. Dev. Nutr. 2018, 2, nzy067. [Google Scholar] [CrossRef] [PubMed]
- Meli, F.; Puccio, G.; Cajozzo, C.; Ricottone, G.L.; Pecquet, S.; Sprenger, N.; Steenhout, P. Growth and safety evaluation of infant formulae containing oligosaccharides derived from bovine milk: A randomized, double-blind, noninferiority trial. BMC Pediatr. 2014, 14, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, P.; Bolton, K.D.; Velaphi, S.; De Groot, N.; Emady-Azar, S.; Pecquet, S.; Steenhout, P. Early Benefits of a Starter Formula Enriched in Prebiotics and Probiotics on the Gut Microbiota of Healthy Infants Born to HIV+ Mothers: A Randomized Double-Blind Controlled Trial. Clin. Med. Insights Pediatr. 2016, 10, CMPed-S40134. [Google Scholar] [CrossRef] [Green Version]
- Radke, M.; Picaud, J.-C.; Loui, A.; Cambonie, G.; Faas, D.; Lafeber, H.N.; de Groot, N.; Pecquet, S.S.; Steenhout, P.G.; Hascoet, J.-M. Starter formula enriched in prebiotics and probiotics ensures normal growth of infants and promotes gut health: A randomized clinical trial. Pediatr. Res. 2017, 81, 622–631. [Google Scholar] [CrossRef] [Green Version]
- Castanet, M.; Costalos, C.; Haiden, N.; Hascoet, J.-M.; Berger, B.; Sprenger, N.; Grathwohl, D.; Brüssow, H.; De Groot, N.; Steenhout, P.; et al. Early Effect of Supplemented Infant Formulae on Intestinal Biomarkers and Microbiota: A Randomized Clinical Trial. Nutriemts 2020, 12, 1481. [Google Scholar] [CrossRef]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.-Z.; et al. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell 2018, 174, 1388–1405.e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.V.; Le Van, P.; Le Huy, C.; Gia, K.N.; Weintraub, A. Etiology and epidemiology of diarrhea in children in Hanoi, Vietnam. Int. J. Infect. Dis. 2006, 10, 298–308. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, R.; Tsuji, H.; Takahashi, T.; Nomoto, K.; Kawashima, K.; Nagata, S.; Yamashiro, Y. Gut dysbiosis following C-section instigates higher colonisation of toxigenic Clostridium perfringens in infants. Benef. Microbes 2017, 8, 353–365. [Google Scholar] [CrossRef] [PubMed]
- GBD Diarrhoeal Diseases Collaborators. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef] [Green Version]
- Thongaram, T.; Hoeflinger, J.L.; Chow, J.; Miller, M.J. Human milk oligosaccharide consumption by probiotic and human-associated bifidobacteria and lactobacilli. J. Dairy Sci. 2017, 100, 7825–7833. [Google Scholar] [CrossRef] [Green Version]
- Wright, D.P.; I Rosendale, D.; Roberton, A.M. Prevotella enzymes involved in mucin oligosaccharide degradation and evidence for a small operon of genes expressed during growth on mucin. FEMS Microbiol. Lett. 2000, 190, 73–79. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.4-6. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 22 February 2018).
- Ramette, A.N. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [Green Version]
- Rautava, S.; Luoto, R.; Salminen, S.; Isolauri, E. Microbial contact during pregnancy, intestinal colonization and human disease. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 565–576. [Google Scholar] [CrossRef]
- Rook, G.A.; Raison, C.L.; Lowry, C.A. Microbiota, immunoregulatory old friends and psychiatric disorders. Adv. Exp. Med. Biol. 2014, 817, 319–356. [Google Scholar]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2010, 108 (Suppl. 1), 4578–4585. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Monaco, M.H.; Donovan, S.M. Impact of early gut microbiota on immune and metabolic development and function. Semin. Fetal Neonatal Med. 2016, 21, 380–387. [Google Scholar] [CrossRef]
- Davis, E.C.; Dinsmoor, A.M.; Wang, M.; Donovan, S.M. Microbiome Composition in Pediatric Populations from Birth to Adolescence: Impact of Diet and Prebiotic and Probiotic Interventions. Dig. Dis. Sci. 2020, 65, 706–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Marriage, B.J.; Buck, R.H.; Goehring, K.C.; Oliver, J.S.; Williams, J.A. Infants Fed a Lower Calorie Formula With 2′FL Show Growth and 2′FL Uptake Like Breast-Fed Infants. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 649–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, E.M.; Joshi, L.; Hickey, R.M. Symposium review: Dairy-derived oligosaccharides—Their influence on host–microbe interactions in the gastrointestinal tract of infants. J. Dairy Sci. 2020, 103, 3816–3827. [Google Scholar] [CrossRef] [PubMed]
- Mehra, R.; Barile, D.; Marotta, M.; Lebrilla, C.B.; Chu, C.; German, J.B. Novel High-Molecular Weight Fucosylated Milk Oligosaccharides Identified in Dairy Streams. PLoS ONE 2014, 9, e96040. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Gibson, G.R.; Cummings, J.H. Comparison of fermentation reactions in different regions of the human colon. J. Appl. Bacteriol. 1992, 72, 57–64. [Google Scholar]
- Van Limpt, C.; Crienen, A.; Vriesema, A.; Knol, J. Effect of Colonic Short Chain Fatty Acids, Lactate and PH on The Growth of Common Gut Pathogens. Pediatr. Res. 2004, 56, 487. [Google Scholar] [CrossRef] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulet, M.; Phothirath, P.; Allais, L.; Schilter, B. Pre-clinical safety evaluation of the synthetic human milk, nature-identical, oligosaccharide 2′-O-Fucosyllactose (2′FL). Regul. Toxicol. Pharmacol. 2014, 68, 59–69. [Google Scholar] [CrossRef]
- Wang, M.; Radlowski, E.C.; Li, M.; Monaco, M.H.; Donovan, S.M. Feeding Mode, but Not Prebiotics, Affects Colonic Microbiota Composition and Volatile Fatty Acid Concentrations in Sow-Reared, Formula-Fed, and Combination-Fed Piglets. J. Nutr. 2019, 149, 2156–2163. [Google Scholar] [CrossRef]
- Berding, K.; Wang, M.; Monaco, M.H.; Alexander, L.S.; Mudd, A.T.; Chichlowski, M.; Waworuntu, R.V.; Berg, B.M.; Miller, M.J.; Dilger, R.N.; et al. Prebiotics and Bioactive Milk Fractions Affect Gut Development, Microbiota, and Neurotransmitter Expression in Piglets. J. Pediatr. Gastroenterol. Nutr. 2016, 63, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Haarman, M.; Knol, J. Quantitative Real-Time PCR Assays To Identify and Quantify Fecal Bifidobacterium Species in Infants Receiving a Prebiotic Infant Formula. Appl. Environ. Microbiol. 2005, 71, 2318–2324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinritz, S.N.; Mosenthin, R.; Weiss, E. Use of pigs as a potential model for research into dietary modulation of the human gut microbiota. Nutr. Res. Rev. 2013, 26, 191–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakanaka, M.; Gotoh, A.; Yoshida, K.; Odamaki, T.; Koguchi, H.; Xiao, J.-Z.; Kitaoka, M.; Katayama, T. Varied Pathways of Infant Gut-Associated Bifidobacterium to Assimilate Human Milk Oligosaccharides: Prevalence of the Gene Set and Its Correlation with Bifidobacteria-Rich Microbiota Formation. Nutrients 2019, 12, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Donovan, S.M. Human Microbiota-Associated Swine: Current Progress and Future Opportunities. ILAR J. 2015, 56, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Finegold, S.M.; Song, Y.; Lawson, P.A. Reclassification of Clostridium coccoides, Ruminococcus hansenii, Ruminococcus hydrogenotrophicus, Ruminococcus luti, Ruminococcus productus and Ruminococcus schinkii as Blautia coccoides gen. nov., comb. nov., Blautia hansenii comb. nov., Blautia hydrogenotrophica comb. nov., Blautia luti comb. nov., Blautia producta comb. nov., Blautia schinkii comb. nov. and description of Blautia wexlerae sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2008, 58, 1896–1902. [Google Scholar] [PubMed]
- Kurakawa, T.; Ogata, K.; Matsuda, K.; Tsuji, H.; Kubota, H.; Takada, T.; Kado, Y.; Asahara, T.; Takahashi, T.; Nomoto, K. Diversity of Intestinal Clostridium coccoides Group in the Japanese Population, as Demonstrated by Reverse Transcription-Quantitative PCR. PLoS ONE 2015, 10, e0126226. [Google Scholar] [CrossRef] [Green Version]
- Eren, A.M.; Sogin, M.L.; Morrison, H.G.; Vineis, J.H.; Fisher, J.C.; Newton, R.J.; McLellan, S.L. A single genus in the gut microbiome reflects host preference and specificity. ISME J. 2015, 9, 90–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, R.; Sakaue, Y.; Sawai, C.; Sawai, T.; Ozeki, M.; Romero-Pérez, G.A.; Tsukahara, T. A preliminary investigation on the relationship between gut microbiota and gene expressions in peripheral mononuclear cells of infants with autism spectrum disorders. Biosci. Biotechnol. Biochem. 2016, 80, 2450–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murri, M.; Leiva, I.; Gomez-Zumaquero, J.M.; Tinahones, F.J.; Cardona, F.; Soriguer, F.; Queipo-Ortuño, M.I. Gut microbiota in children with type 1 diabetes differs from that in healthy children: A case-control study. BMC Med. 2013, 11, 46. [Google Scholar] [CrossRef] [Green Version]
- Jost, T.; Lacroix, C.; Braegger, C.P.; Chassard, C. New Insights in Gut Microbiota Establishment in Healthy Breast Fed Neonates. PLoS ONE 2012, 7, e44595. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; Barboza, M.; Sonnenburg, E.D.; Pudlo, N.; Martens, E.C.; Desai, P.; Lebrilla, C.B.; Weimer, B.C.; Mills, D.A.; German, J.B.; et al. Bacteroides in the Infant Gut Consume Milk Oligosaccharides via Mucus-Utilization Pathways. Cell Host Microbe 2011, 10, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, E.C.; Wang, M.; Donovan, S.M. The role of early life nutrition in the establishment of gastrointestinal microbial composition and function. Gut Microbes 2017, 8, 143–171. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Palacio, S.D.; Montes, S.A.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Monaco, M.H.; Hauser, J.; Yan, J.; Dilger, R.N.; Donovan, S.M. Bovine Milk Oligosaccharides and Human Milk Oligosaccharides Modulate the Gut Microbiota Composition and Volatile Fatty Acid Concentrations in a Preclinical Neonatal Model. Microorganisms 2021, 9, 884. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050884
Wang M, Monaco MH, Hauser J, Yan J, Dilger RN, Donovan SM. Bovine Milk Oligosaccharides and Human Milk Oligosaccharides Modulate the Gut Microbiota Composition and Volatile Fatty Acid Concentrations in a Preclinical Neonatal Model. Microorganisms. 2021; 9(5):884. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050884
Chicago/Turabian StyleWang, Mei, Marcia H. Monaco, Jonas Hauser, Jian Yan, Ryan N. Dilger, and Sharon M. Donovan. 2021. "Bovine Milk Oligosaccharides and Human Milk Oligosaccharides Modulate the Gut Microbiota Composition and Volatile Fatty Acid Concentrations in a Preclinical Neonatal Model" Microorganisms 9, no. 5: 884. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050884