
Diversity and Geographical Structure of Xanthomonas citri pv. citri on Citrus in the South West Indian Ocean Region

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
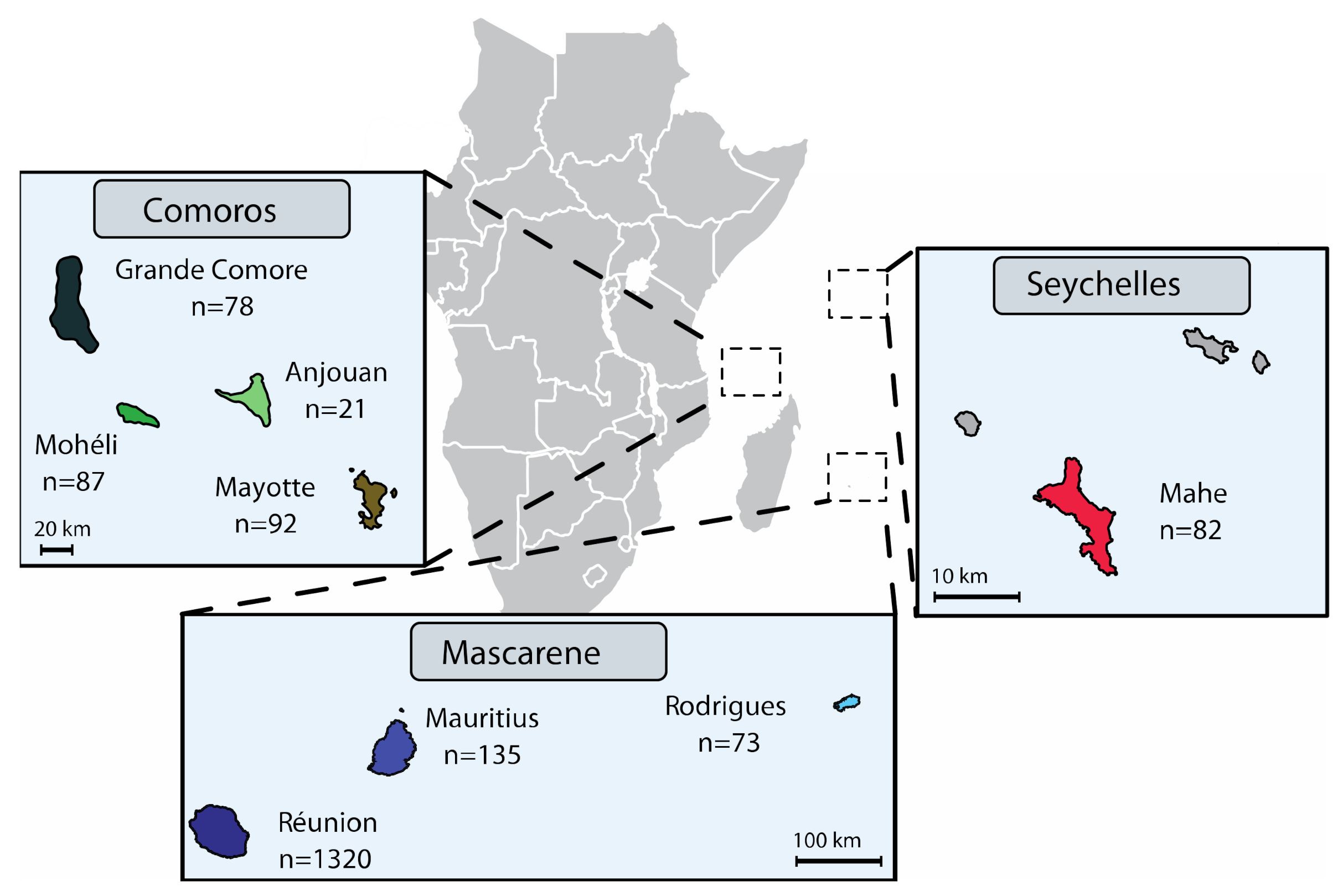
2.1. Bacterial Strains and Media
2.2. Microsatellite Genotyping (MLVA-14)
2.3. Minisatellite Typing (MLVA-31)
2.4. Typeability of SWIO Strains with XAC1051-qPCR
2.5. Detached Leaf Assay-Based Pathogenicity Tests
2.6. Attached Leaf Assay-Based Pathogenicity and in Planta Growth of Lineage 1 Strains
3. Results
3.1. Genetic Diversity among Strains from Different Islands in the SWIO Region Revealed by Microsatellite Typing and Cop-PCR
3.2. Minisatellite-Based Assignation of SWIO Strains to X. citri pv. citri Genetic Lineages
3.3. Pathogenicity Assays Confirm the Prevalence of Pathotype A and the Limited Presence of Pathotype A* in the SWIO Region
3.4. Relative Virulence of SWIO Strains from the Four Major Genetic Clusters on Three Citrus Species
4. Discussion
4.1. Genetic Lineage 1 is Markedly Prevalent in the SWIO Region, Displays a Geographic Structure and Sporadically Cohabits with Nonepidemic Lineage 4 Strains
4.2. The Seychelles Host Genetically Diverse X. citri pv. citri Pathotype A Strains Differing in Pathogenicity
4.3. Microsatellite Typing Suggests that the Pathogen’s Inter- and Intra-Island Movements Differ
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pimentel, D.; McNair, S.; Janecka, J.; Wightman, J.; Simmonds, C.; O’Connell, C.; Wong, E.; Russel, L.; Zern, J.; Aquino, T.; et al. Economic and environmental threats of alien plant, animal, and microbe invasions. Agric. Ecosyst. Environ. 2001, 84, 1–20. [Google Scholar] [CrossRef]
- Vurro, M.; Bonciani, B.; Vannacci, G. Emerging infectious diseases of crop plants in developing countries: Impact on agriculture and socio-economic consequences. Food Secur. 2010, 2, 113–132. [Google Scholar] [CrossRef]
- Engering, A.; Hogerwerf, L.; Slingenbergh, J. Pathogen-host-environment interplay and disease emergence. Emerg. Microbes Infect. 2013, 2, e5. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Bebber, D.P.; Holmes, T.; Gurr, S.J. The global spread of crop pests and pathogens. Glob. Ecol. Biogeogr. 2014, 23, 1398–1407. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.A.; Terol, J.; Ibanez, V.; Lopez-Garcia, A.; Perez-Roman, E.; Borreda, C.; Domingo, C.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of Citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.H.; Gottwald, T.R.; Cubero, J.; Achor, D.S. Xanthomonas axonopodis pvcitri: Factors affecting successful eradication of citrus canker. Mol. Plant Pathol. 2004, 5, 1–15. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Graham, J.H. A device for precise and nondisruptive stomatal inoculation of leaf tissue with bacterial pathogens. Phytopathology 1992, 82, 930–935. [Google Scholar] [CrossRef]
- Vernière, C.; Gottwald, T.R.; Pruvost, O. Disease development and symptom expression of Xanthomonas axonopodis pv. citri in various citrus plant tissues. Phytopathology 2003, 93, 832–843. [Google Scholar] [CrossRef] [Green Version]
- Vernière, C.; Hartung, J.S.; Pruvost, O.P.; Civerolo, E.L.; Alvarez, A.M.; Maestri, P.; Luisetti, J. Characterization of phenotypically distinct strains of Xanthomonas axonopodis pv. citri from Southwest Asia. Eur. J. Plant Pathol. 1998, 104, 477–487. [Google Scholar] [CrossRef]
- Sun, X.A.; Stall, R.E.; Jones, J.B.; Cubero, J.; Gottwald, T.R.; Graham, J.H.; Dixon, W.N.; Schubert, T.S.; Chaloux, P.H.; Stromberg, V.K.; et al. Detection and characterization of a new strain of citrus canker bacteria from key Mexican lime and Alemow in South Florida. Plant Dis. 2004, 88, 1179–1188. [Google Scholar] [CrossRef] [Green Version]
- Bull, C.T.; Koike, S.T. Practical benefits of knowing the enemy: Modern molecular tools for diagnosing the etiology of bacterial diseases and understanding the taxonomy and diversity of plant-pathogenic bacteria. Annu. Rev. Phytopathol. 2015, 53, 157–180. [Google Scholar] [CrossRef]
- Vernière, C.; Bui Thi Ngoc, L.; Jarne, P.; Ravigné, V.; Guérin, F.; Gagnevin, L.; Le Mai, N.; Chau, N.M.; Pruvost, O. Highly polymorphic markers reveal the establishment of an invasive lineage of the citrus bacterial pathogen Xanthomonas citri pv. citri in its area of origin. Environ. Microbiol. 2014, 16, 2226–2237. [Google Scholar] [CrossRef]
- Pruvost, O.; Magne, M.; Boyer, K.; Leduc, A.; Tourterel, C.; Drevet, C.; Ravigné, V.; Gagnevin, L.; Guérin, F.; Chiroleu, F.; et al. A MLVA genotyping scheme for global surveillance of the citrus pathogen Xanthomonas citri pv. citri suggests a worldwide geographical expansion of a single genetic lineage. PLoS ONE 2014, 9, e98129. [Google Scholar] [CrossRef]
- Leduc, A.; Traoré, Y.N.; Boyer, K.; Magne, M.; Grygiel, P.; Juhasz, C.; Boyer, C.; Guérin, F.; Wonni, I.; Ouedraogo, L.; et al. Bridgehead invasion of a monomorphic plant pathogenic bacterium: Xanthomonas citri pv. citri, an emerging citrus pathogen in Mali and Burkina Faso. Environ. Microbiol. 2015, 17, 4429–4442. [Google Scholar] [CrossRef]
- Pruvost, O.; Boyer, K.; Ravigné, V.; Richard, D.; Vernière, C. Deciphering how plant pathogenic bacteria disperse and meet: Molecular epidemiology of Xanthomonas citri pv. citri at microgeographic scales in a tropical area of Asiatic citrus canker endemicity. Evol. Appl. 2019, 12, 1523–1538. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.L.; Lefeuvre, P.; Escalon, A.; Barbe, V.; Cruveiller, S.; Gagnevin, L.; Pruvost, O. Comparative genomics of 43 strains of Xanthomonas citri pv. citri reveals the evolutionary events giving rise to pathotypes with different host ranges. BMC Genom. 2015, 16, 1098. [Google Scholar] [CrossRef] [Green Version]
- Bui Thi Ngoc, L.; Vernière, C.; Pruvost, O.; Kositcharoenkul, N.; Phawichit, S. First report in Thailand of Xanthomonas axonopodis pv. citri-A* causing citrus canker on lime. Plant Dis. 2007, 91, 771. [Google Scholar] [CrossRef]
- Derso, E.; Vernière, C.; Pruvost, O. First report of Xanthomonas citri pv. citri-A* causing citrus canker on lime in Ethiopia. Plant Dis. 2009, 93, 203. [Google Scholar] [CrossRef]
- Pruvost, O.; Goodarzi, T.; Boyer, K.; Soltaninejad, H.; Escalon, A.; Alavi, S.M.; Javegny, S.; Boyer, C.; Cottyn, B.; Gagnevin, L.; et al. Genetic structure analysis of strains causing citrus canker in Iran reveals the presence of two different lineages of Xanthomonas citri pv. citri pathotype A*. Plant Pathol. 2015, 64, 776–784. [Google Scholar] [CrossRef]
- Gottwald, T.R.; Graham, J.H.; Schubert, T.S. Citrus canker: The pathogen and its impact. Plant Health Prog. 2002, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Richard, D.; Ravigné, V.; Rieux, A.; Facon, B.; Boyer, C.; Boyer, K.; Grygiel, P.; Javegny, S.; Terville, M.; Canteros, B.I.; et al. Adaptation of genetically monomorphic bacteria: Evolution of copper resistance through multiple horizontal gene transfers of complex and versatile mobile genetic elements. Mol. Ecol. 2017, 26, 2131–2149. [Google Scholar] [CrossRef] [PubMed]
- Behlau, F.; Gochez, A.M.; Jones, J.B. Diversity and copper resistance of Xanthomonas affecting citrus. Trop. Plant Pathol. 2020, 45, 200–212. [Google Scholar] [CrossRef]
- Richard, D.; Tribot, N.; Boyer, C.; Terville, M.; Boyer, K.; Javegny, S.; Roux-Cuvelier, M.; Pruvost, O.; Moreau, A.; Chabirand, A.; et al. First report of copper-resistant Xanthomonas citri pv. citri pathotype A causing Asiatic citrus canker in Réunion, France. Plant Dis. 2017, 101, 503. [Google Scholar] [CrossRef]
- Aubert, B. Vergers de la Réunion et de l’Océan Indien. In Hommes et Fruits en Pays du Sud; CIRAD: Montpellier, France, 2014; pp. 111–166. [Google Scholar]
- Grygiel, P.; Seny-Couty, A.; Abdou Hassani, F.; Boyer, C.; Boyer, K.; Vernière, C.; Pruvost, O.; Hamza, A.A. First report of Xanthomonas citri pv. citri pathotype A causing Asiatic citrus canker in Grande Comore and Anjouan. Plant Dis. 2014, 98, 1739. [Google Scholar] [CrossRef] [Green Version]
- Pruvost, O.; Roumagnac, P.; Gaube, C.; Chiroleu, F.; Gagnevin, L. New media for the semi-selective isolation and enumeration of Xanthomonas campestris pv. mangiferaeindicae, the causal agent of mango bacterial black spot. J. Appl. Microbiol. 2005, 99, 803–815. [Google Scholar] [CrossRef]
- Behlau, F.; Canteros, B.I.; Jones, J.B.; Graham, J.H. Copper resistance genes from different xanthomonads and citrus epiphytic bacteria confer resistance to Xanthomonas citri subsp citri. Eur. J. Plant Pathol. 2012, 133, 949–963. [Google Scholar] [CrossRef]
- Da Silva, A.C.; Ferro, J.A.; Reinach, F.C.; Farah, C.S.; Furlan, L.R.; Quaggio, R.B.; Monteiro-Vitorello, C.B.; Van Sluys, M.A.; Almeida, N.F.; Alves, L.M.; et al. Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature 2002, 417, 459–463. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J. Hierfstat, a package for R to compute and test hierarchical F-statistics. Mol. Ecol. Notes 2005, 5, 184–186. [Google Scholar] [CrossRef] [Green Version]
- Francisco, A.P.; Vaz, C.; Monteiro, P.T.; Melo-Cristino, J.; Ramirez, M.; Carriço, J.A. PHYLOViZ: Phylogenetic inference and data visualization for sequence based typing methods. BMC Bioinform. 2012, 13, 87. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Robene, I.; Maillot-Lebon, V.; Chabirand, A.; Moreau, A.; Becker, N.; Moumene, A.; Rieux, A.; Campos, P.; Gagnevin, L.; Gaudeul, M.; et al. Development and comparative validation of genomic-driven PCR-based assays to detect Xanthomonas citri pv. citri in citrus plants. BMC Microbiol. 2020, 20, 296. [Google Scholar] [CrossRef]
- Favaro, M.A.; Micheloud, N.G.; Roeschlin, R.A.; Chiesa, M.A.; Castagnaro, A.P.; Vojnov, A.A.; Gmitter, F.G., Jr.; Gadea, J.; Rista, L.M.; Gariglio, N.F.; et al. Surface barriers of mandarin ‘Okitsu’ leaves make a major contribution to canker disease resistance. Phytopathology 2014, 104, 970–976. [Google Scholar] [CrossRef] [Green Version]
- Reed, H.S.; HIrano, E. The density of stomata in citrus leaves. J. Agric. Res. 1931, 43, 209–222. [Google Scholar]
- Simko, I.; Piepho, H.P. The Area Under the Disease Progress Stairs: Calculation, advantage, and application. Phytopathology 2012, 102, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Brooks, M.E.; Kristensen, K.; Van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2019; p. 608. [Google Scholar]
- Scherm, H.; Ojiambo, P.S. Applications of survival analysis in botanical epidemiology. Phytopathology 2004, 94, 1022–1026. [Google Scholar] [CrossRef] [Green Version]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; p. 495. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escalon, A.; Javegny, S.; Vernière, C.; Noël, L.D.; Vital, K.; Poussier, S.; Hajri, A.; Boureau, T.; Pruvost, O.; Arlat, M.; et al. Variations in type III effector repertoires, pathological phenotypes and host range of Xanthomonas citri pv. citri pathotypes. Mol. Plant Pathol. 2013, 14, 483–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruvost, O.; Boher, B.; Brocherieux, C.; Nicole, M.; Chiroleu, F. Survival of Xanthomonas axonopodis pv. citri in leaf lesions under tropical environmental conditions and simulated splash dispersal of inoculum. Phytopathology 2002, 92, 336–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmer, L.W.; Gottwald, T.R.; Zitko, S.E. Bacterial exudation from lesions of asiatic citrus canker and citrus bacterial spot. Plant Dis. 1991, 75, 192–195. [Google Scholar] [CrossRef]
- Richard, D.; Rieux, A.; Lefeuvre, P.; Hamza, A.; Lobin, K.K.; Naiken, M.; Stravens, R.; Boyer, C.; Boyer, K.; Javegny, S.; et al. Draft genome sequences of 284 Xanthomonas citri pv. citri strains causing Asiatic citrus canker. Microbiol. Resour. Announc. 2021, 10, 1. [Google Scholar] [CrossRef]
- Estoup, A.; Jarne, P.; Cornuet, J.M. Homoplasy and mutation model at microsatellite loci and their consequences for population genetics analysis. Mol. Ecol. 2002, 11, 1591–1604. [Google Scholar] [CrossRef]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef]
- Favaro, M.A.; Molina, M.C.; Roeschlin, R.A.; Gadea, J.; Gariglio, N.; Marano, M.R. Different responses in mandarin cultivars uncover a role of cuticular waxes in the resistance to citrus canker. Phytopathology 2020, 110, 1791–1801. [Google Scholar] [CrossRef]
- Newberry, E.A.; Bhandari, R.; Minsavage, G.V.; Timilsina, S.; Jibrin, M.O.; Kemble, J.; Sikora, E.J.; Jones, J.B.; Potnis, N. Independent evolution with the gene flux originating from multiple Xanthomonas species explains genomic heterogeneity in Xanthomonas perforans. Appl. Environ. Microbiol. 2019, 85, e00885-19. [Google Scholar] [CrossRef]
- Newberry, E.; Bhandari, R.; Kemble, J.; Sikora, E.; Potnis, N. Genome-resolved metagenomics to study co-occurrence patterns and intraspecific heterogeneity among plant pathogen metapopulations. Environ. Microbiol. 2020, 22, 2693–2708. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Island | N 1 | NH 2 | A 3 | AP 4 | HE 5 | Cluster 6 |
|---|---|---|---|---|---|---|
| The Comoros | ||||||
| Anjouan | 21 | 19 | 3.786 | 0.000 | 0.430 | GC1 (95%) |
| Grande Comore | 78 | 51 | 3.604 | 0.000 | 0.490 | GC1 (100%) |
| Mayotte | 92 | 56 | 3.117 | 0.185 | 0.439 | GC1 (99%) |
| Moheli | 86 | 78 | 5.644 | 0.082 | 0.666 | GC1 (74%) |
| The Mascarenes | ||||||
| Mauritius | 133 | 73 | 4.013 | 0.083 | 0.468 | GC1 (95%) |
| Réunion 7 | 1320 | 789 | 5.289 | 0.455 | 0.633 | GC1 (98 %) |
| Rodrigues | 73 | 63 | 5.265 | 0.014 | 0.649 | GC1 (63%) |
| The Seychelles | ||||||
| Mahé | 82 | 46 | 3.512 | 0.549 | 0.680 | GC2 (54%) GC3 (30%) GC4 (16%) |
| Cluster (Microsatellite Data) | Country (Archipelago) | Source | n | Genetic Lineage (Minisatellite Data) 1 |
|---|---|---|---|---|
| GC1 | Anjouan (The Comoros) | This study | 16 | 1 |
| GC1 | Grande Comore (The Comoros) | This study | 5 | 1 |
| GC1 | Mayotte (The Comoros) | This study | 56 | 1 |
| GC1 | Moheli (The Comoros) | This study | 22 | 1 |
| GC1 | Mauritius (The Mascarenes) | This study | 25 | 1 |
| GC1 | Rodrigues (The Mascarenes) | This study | 11 | 1 |
| GC1 | Réunion (The Mascarenes) | This study | 25 | 1 |
| GC1 | Réunion (The Mascarenes) | [23] | 162 | 1 |
| GC2 | Mahé (The Seychelles) | This study | 13 | 1 |
| GC3 | Mahé (The Seychelles) | This study | 6 | 1 |
| GC4 | Mahé (The Seychelles) | This study | 5 | 1 |
| minor/singleton | Anjouan (The Comoros) | This study | 1 | 1 |
| minor/singleton | Mayotte (The Comoros) | This study | 1 | 1 |
| minor/singleton | Moheli (The Comoros) | This study | 18 | 1 |
| minor/singleton | Moheli (The Comoros) | This study | 1 | 4 |
| minor/singleton | Mauritius (The Mascarenes) | This study | 4 | 1 |
| minor/singleton | Mauritius (The Mascarenes) | This study | 2 | 4 |
| minor/singleton | Rodrigues (The Mascarenes) | This study | 24 | 1 |
| minor/singleton | Réunion (The Mascarenes) | This study | 10 | 1 |
| minor/singleton | Réunion (The Mascarenes) | This study | 1 | 4 |
| minor/singleton | Réunion (The Mascarenes) | [23] | 5 | 1 |
| Citron | Mandarin | Sweet Orange | |||||
|---|---|---|---|---|---|---|---|
| Exuded | Total | Exuded | Total | Exuded | Total | ||
| ANOVA probability | 0.002 | 0.004 | 0.006 | < 0.001 | < 0.001 | 0.025 | |
| GC | Strain | Log-transformed cfu lesion−1 compared using Tukey’s tests | |||||
| 1 | LH241 | 6.69 ab | 7.43 a | 6.20 ab | 7.33 abc | 6.97 ab | 7.43 ab |
| 1 | LM089-41 | 7.00 a | 7.41 a | 6.63 a | 7.62 a | 6.88 abc | 7.27 ab |
| 1 | LN005-4 | 6.65 ab | 7.08 ab | 6.20 ab | 6.99 abc | 6.91 abc | 7.58 a |
| 1 | LN007-3 | 6.47 b | 7.30 ab | 6.30 ab | 7.21 abc | 6.79 abc | 7.47 ab |
| 2 | JZ092 | 6.45 b | 7.12 ab | 6.29 ab | 7.19 abc | 6.98 abc | 7.32 ab |
| 2 | LB100-1 | 6.56 ab | 7.14 ab | 5.93 ab | 6.79 bc | 6.52 c | 6.90 b |
| 2 | LJ001 | 6.54 b | 6.92 ab | 6.50 ab | 7.49 ab | 6.72 abc | 7.07 ab |
| 2 | LP029-15 | 6.80 ab | 7.09 ab | 5.65 ab | 6.60 c | 6.39 c | 7.19 ab |
| 3 | JZ094 | 6.16 b | 6.75 ab | 6.12 ab | 7.13 abc | 6.70 abc | 7.01 ab |
| 3 | LP027-3 | 6.75 ab | 7.28 ab | 5.64 ab | 6.49 c | 6.88 abc | 7.38 ab |
| 3 | LP027-5 | 6.32 b | 6.64 b | 6.49 ab | 7.07 abc | 6.60 bc | 7.23 ab |
| 3 | LP027-13 | 6.58 ab | 7.06 ab | 5.85 ab | 6.89 abc | 6.52 c | 7.12 ab |
| 4 | LP028-2 | 6.84 ab | 7.36 a | 5.60 b | 6.57 c | 6.46 bc | 7.17 ab |
| 4 | LP028-3 | 6.73 ab | 7.15 ab | 5.70 ab | 6.64 c | 7.02 a | 7.39 ab |
| 4 | LP028-5 | 6.66 ab | 7.00 ab | 5.92 ab | 6.63 bc | 6.63 abc | 7.34 ab |
| 4 | LP028-6 | 6.80 ab | 7.16 ab | 6.11 ab | 6.82 c | 6.69 abc | 7.31 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pruvost, O.; Richard, D.; Boyer, K.; Javegny, S.; Boyer, C.; Chiroleu, F.; Grygiel, P.; Parvedy, E.; Robène, I.; Maillot-Lebon, V.; et al. Diversity and Geographical Structure of Xanthomonas citri pv. citri on Citrus in the South West Indian Ocean Region. Microorganisms 2021, 9, 945. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050945
Pruvost O, Richard D, Boyer K, Javegny S, Boyer C, Chiroleu F, Grygiel P, Parvedy E, Robène I, Maillot-Lebon V, et al. Diversity and Geographical Structure of Xanthomonas citri pv. citri on Citrus in the South West Indian Ocean Region. Microorganisms. 2021; 9(5):945. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050945
Chicago/Turabian StylePruvost, Olivier, Damien Richard, Karine Boyer, Stéphanie Javegny, Claudine Boyer, Frédéric Chiroleu, Pierre Grygiel, Evelyne Parvedy, Isabelle Robène, Véronique Maillot-Lebon, and et al. 2021. "Diversity and Geographical Structure of Xanthomonas citri pv. citri on Citrus in the South West Indian Ocean Region" Microorganisms 9, no. 5: 945. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9050945