
First Record of a Suspected Human-Pathogenic Borrelia Species in Populations of the Bat Tick Carios vespertilionis in Sweden

Abstract
:1. Introduction
2. Materials and Methods
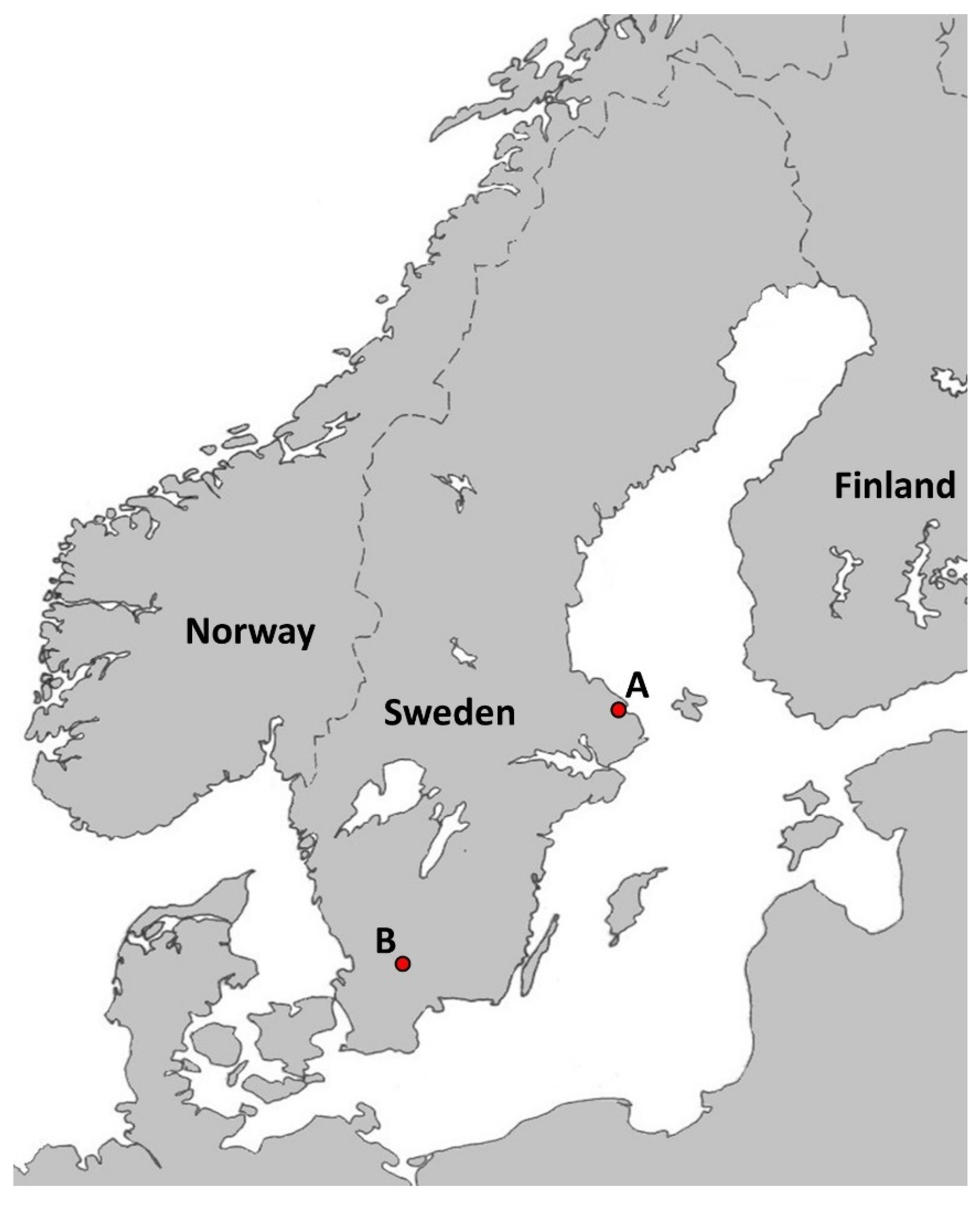
2.1. Tick Collection
2.2. Tick Identification
2.3. Nucleic Acid Extraction and cDNA Synthesis from Ticks
2.4. Detection of Borrelia Bacteria and Determination of Species
2.5. Detection of Borrelia miyamotoi
2.6. Nucleotide Sequencing of PCR-Products and Phylogenetic Analysis
2.7. Statistical Analyses
3. Results
3.1. Tick Collection and Tick Blood Engorgement
3.2. Prevalence of Borrelia Bacteria in the Ticks
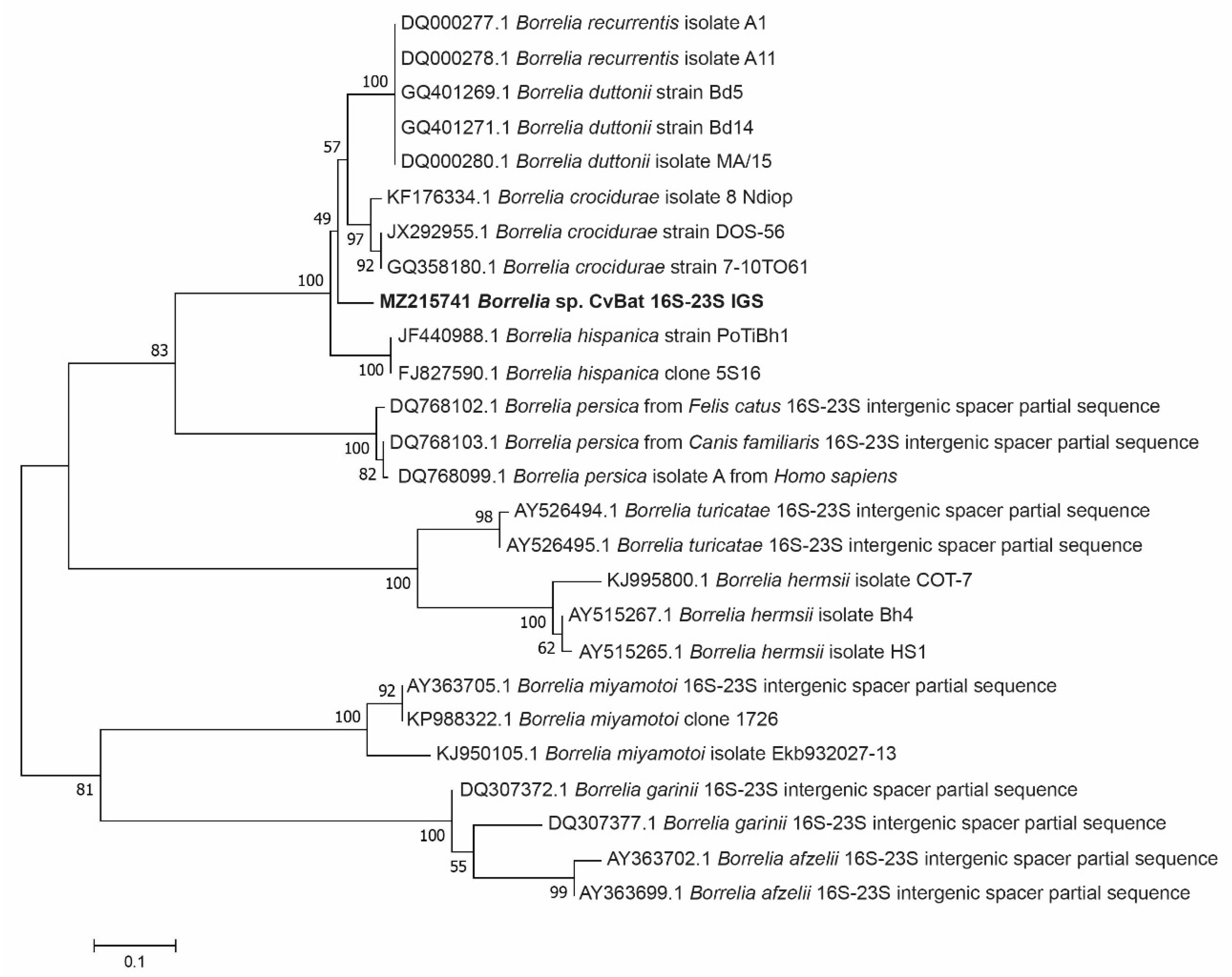
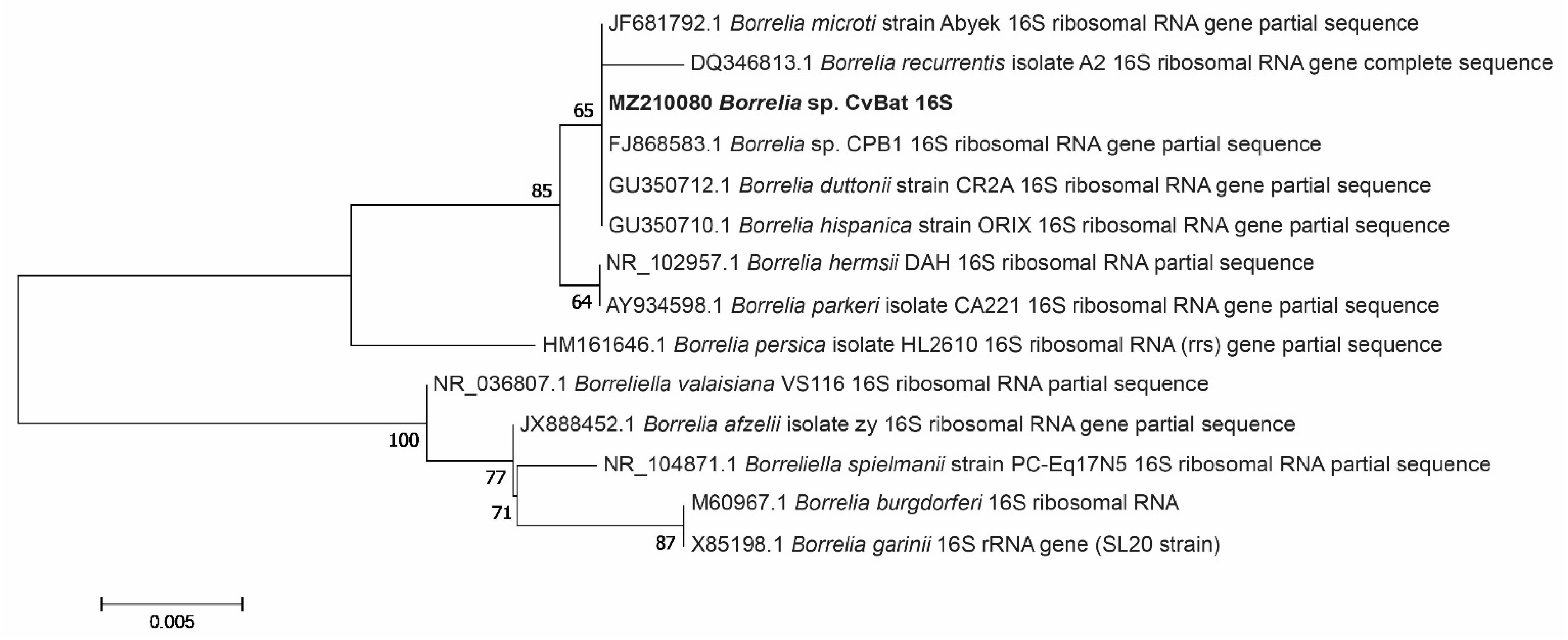
3.3. Phylogenetic Analysis of the Borrelia bacteria Detected in Ticks
3.4. Prevalence of B. miyamotoi in the Ticks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brook, C.E.; Dobson, A.P. Bats as ‘special’ reservoirs for emerging zoonotic pathogens. Trends Microbiol. 2015, 23, 172–180. [Google Scholar] [CrossRef]
- Leroy, E.M.; Kumulungui, B.; Pourrut, X.; Rouquet, P.; Hassanin, A.; Yaba, P.; Délicat, A.; Paweska, J.T.; Gonzalez, J.-P.; Swanepoel, R. Fruit bats as reservoirs of Ebola virus. Nature 2005, 438, 575–576. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Shi, Z.; Yu, M.; Ren, W.; Smith, C.; Epstein, J.H.; Wang, H.; Crameri, G.; Hu, Z.; Zhang, H.; et al. Bats are natural reservoirs of SARS-like coronaviruses. Science 2005, 310, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Dobson, A.P. Virology: What links bats to emerging infectious diseases? Science 2005, 310, 628–629. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.J. Bats as the reservoir for outbreaks of emerging infectious diseases. Eurosurveillance 2005, 10, E051110.4. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Hu, Z.; Deng, F.; Shen, S. Tick-Borne Viruses. Virol. Sin. 2018, 33, 21–43. [Google Scholar] [CrossRef] [Green Version]
- Streicker, D.G.; Gilbert, A.T. Contextualizing bats as viral reservoirs. Science 2020, 370, 172–173. [Google Scholar] [CrossRef]
- Wilson, D.E.; Mittermeier, R.A. Handbook of Mammals of the World, Volume 9—Bats; Lynx Edicions: Barcelona, Spain, 2019; 1008p, ISBN 9788416728190. [Google Scholar]
- Swedish Environmental Protection Agency. Fladdermusarter i Sverige [Bat Species in Sweden]. Available online: http://www.naturvardsverket.se/Var-natur/Djur-och-vaxter/Rad/Fladdermossen-i-Sverige/Fladdermusarter-i-Sverige/ (accessed on 28 April 2021).
- Lourenço, S.I.; Palmeirim, J.M. Influence of temperature in roost selection by Pipistrellus pygmaeus (Chiroptera): Relevance for the design of bat boxes. Biol. Conserv. 2004, 119, 237–243. [Google Scholar] [CrossRef]
- Ahlén, I. Fladdermusfaunan i Sverige—Arternas utbredning och Status. Kunskapsläget 2011. [Summary: The Bat fauna in Sweden. Present knowledge on distribution and status]. Fauna Flora 2011, 106, 2–19. [Google Scholar]
- Hoogstraal, H. Argasid and Nuttalliellid ticks as parasites and vectors. Adv. Parasitol. 1985, 24, 135–238. [Google Scholar] [CrossRef]
- Arthur, D.A. British Ticks; Butterworths: London, UK, 1963. [Google Scholar]
- Estrada-Peña, A.; Mihalca, A.; Petney, T. Ticks of Europe and North Africa: A Guide to Species Identification, 1st ed.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Hornok, S.; Szőke, K.; Tu, V.T.; Kontschán, J.; Takács, N.; Sándor, A.D.; Halajian, A.; Földvári, G.; Estók, P.; Plantard, O.; et al. Mitochondrial gene heterogeneity of the bat soft tick Argas vespertilionis (Ixodida: Argasidae) in the Palaearctic. Parasites Vectors 2017, 10, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaenson, T.G.; Tälleklint, L.; Lundqvist, L.; Olsen, B.; Chirico, J.; Mejlon, H. Geographical distribution, host associations, and vector roles of ticks (Acari: Ixodidae, Argasidae) in Sweden. J. Med. Èntomol. 1994, 31, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Péter, Á.; Barti, L.; Corduneanu, A.; Hornok, S.; Mihalca, A.D.; Sándor, A.D. First record of Ixodes simplex found on a human host, with a review of cases of human infestation by bat tick species occurring in Europe. Ticks Tick-Borne Dis. 2021, 12, 101722. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Szőke, K.; Görföl, T.; Foldvari, G.; Tu, V.T.; Takács, N.; Kontschán, J.; Sándor, A.D.; Estók, P.; Epis, S.; et al. Molecular investigations of the bat tick Argas vespertilionis (Ixodida: Argasidae) and Babesia vesperuginis (Apicomplexa: Piroplasmida) reflect “bat connection” between Central Europe and Central Asia. Exp. Appl. Acarol. 2017, 72, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Hoogstraal, H. African Ixodoidea. Ticks of the Sudan (with special reference to Equatoria Province and with preliminary reviews of the genera Boophilus, Margaropus, and Hyalomma). In African Ixodoidea; United States Naval Medical Research Unit no. 3: Cairo, Egypt, 1956; Volume 1, pp. 1–1105. [Google Scholar]
- Shah, A.; Khan, M.; Iqbal, Z.; Sajid, M.; Akhtar, M. Some epidemiological aspects and vector role of tick infestation on layers in the Faisalabad district (Pakistan). World’s Poult. Sci. J. 2006, 62, 145–157. [Google Scholar] [CrossRef]
- Hubbard, M.J.; Baker, A.S.; Cann, K.J. Distribution of Borrelia burgdorferi s.l. spirochaete DNA in British ticks (Argasidae and Ixodidae) since the 19th Century, assessed by PCR. Med. Vet. Èntomol. 1998, 12, 89–97. [Google Scholar] [CrossRef]
- Zabashta, M.V.; Orlova, M.V.; Pichurina, N.L.; Khametova, A.P.; Romanova, L.V.; Borodina, T.N.; Zabashta, A.V. Participation of bats (Chiroptera, Mammalia) and their ectoparasites in circulation of pathogens of natural focal infections in the south of Russia. Èntomol. Rev. 2019, 99, 513–521. [Google Scholar] [CrossRef]
- Socolovschi, C.; Kernif, T.; Raoult, D.; Parola, P. Borrelia, Rickettsia, and Ehrlichia species in bat ticks, France, 2010. Emerg. Infect. Dis. 2012, 18, 1966–1975. [Google Scholar] [CrossRef]
- Evans, N.J.; Bown, K.; Timofte, D.; Simpson, V.R.; Birtles, R. Fatal borreliosis in bat caused by relapsing fever spirochete, United Kingdom. Emerg. Infect. Dis. 2009, 15, 1331–1333. [Google Scholar] [CrossRef]
- Lvov, D.K.; Karas, F.R.; Timofeev, E.M.; Tsyrkin, Y.M.; Vargina, S.G.; Veselovskaya, O.V.; Osipova, N.Z.; Grebenyuk, Y.I.; Gromashevski, V.L.; Steblyanko, S.N.; et al. “Issyk-Kul” virus, a new arbovirus isolated from bats and argas (Carios) vespertilionis (Latr., 1802) in the Kirghiz S.S.R. Arch. Gesamte Virusforsch. 1973, 42, 207–209. [Google Scholar] [CrossRef]
- Zhmaeva, Z.M.; Pchelkina, A.A.; Belashova, V.S. Spontaneous infection of Argas vespertilionis with Rickettsia burnetii in the south of Kazakhstan. Med. Parazitol. Mosk. 1966, 35, 595–596. [Google Scholar] [PubMed]
- Lv, J.; Fernández de Marco, M.D.M.; Goharriz, H.; Phipps, L.P.; McElhinney, L.M.; Hernández-Triana, L.M.; Wu, S.; Lin, X.; Fooks, A.R.; Johnson, N. Detection of tick-borne bacteria and babesia with zoonotic potential in Argas (Carios) vespertilionis (Latreille, 1802) ticks from British bats. Sci. Rep. 2018, 8, 1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yan, B.; Wang, Q.; Jiang, M.; Tu, C.; Chen, C.; Hornok, S.; Wang, Y. Babesia vesperuginis in Common Pipistrelle (Pipistrellus pipistrellus) and the bat soft tick Argas vespertilionis in the People’s Republic of China. J. Wildl. Dis. 2018, 54, 419–421. [Google Scholar] [CrossRef]
- Hornok, S.; Szőke, K.; Meli, M.L.; Sándor, A.D.; Görföl, T.; Estók, P.; Wang, Y.; Tu, V.T.; Kováts, D.; Boldogh, S.A.; et al. Molecular detection of vector-borne bacteria in bat ticks (Acari: Ixodidae, Argasidae) from eight countries of the Old and New Worlds. Parasites Vectors 2019, 12, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Arthur, D.R.; Nuttall, G.H.F.; Robinson, L.E.; Warburton, C. Ticks, a Monograph of the Ixodoidea; Cambridge [England] University Press: Cambridge, UK, 1908. [Google Scholar]
- Hoogstraal, H. Bat ticks of the genus Argas (Ixodoidea, Argasidae), 3. The Subgenus Carios, A Redescription of A. (C.) vespertilionis (Latreille, 1802), and variation within an Egyptian population. Ann. Èntomol. Soc. Am. 1958, 51, 19–26. [Google Scholar] [CrossRef]
- Filippova, N.A. Argasid ticks (Argasidae). Fauna of the USSR; Paukoobraznye, Nauka: Moscow/Leningrad, Russia, 1966; Volume 4. [Google Scholar]
- Yamaguti, N. Ticks of Japan, Korea, and the Ryukyu Islands; Brigham Young University: Provo, UT, USA, 1971. [Google Scholar]
- Hillyard, P.D. Ticks of North-West Europe; Field Studies Council: Shrewsbury, UK, 1996. [Google Scholar]
- Mans, B.J.; Kelava, S.; Pienaar, R.; Featherston, J.; de Castro, M.H.; Quetglas, J.; Reeves, W.K.; Durden, L.A.; Miller, M.M.; Laverty, T.M.; et al. Nuclear (18S-28S rRNA) and mitochondrial genome markers of Carios (Carios) vespertilionis (Argasidae) support Carios Latreille, 1796 as a lineage embedded in the Ornithodorinae: Re-classification of the Carios sensu Klompen and Oliver (1993) clade into its respective subgenera. Ticks Tick-borne Dis. 2021, 12, 101688. [Google Scholar] [CrossRef]
- Gyllemark, P.; Wilhelmsson, P.; Elm, C.; Hoornstra, D.; Hovius, J.W.; Johansson, M.; Tjernberg, I.; Lindgren, P.E.; Henningsson, J.H.; Sjöwall, J. Are other tick-borne infections overlooked in patients investigated for Lyme neuroborreliosis? A large retrospective study from south-eastern Sweden. Ticks Tick Borne Dis. 2021. under review. [Google Scholar]
- Wilhelmsson, P.; Fryland, L.; Börjesson, S.; Nordgren, J.; Bergström, S.; Ernerudh, J.; Forsberg, P.; Lindgren, P.-E. Prevalence and diversity of Borrelia species in ticks that have bitten humans in Sweden. J. Clin. Microbiol. 2010, 48, 4169–4176. [Google Scholar] [CrossRef] [Green Version]
- Postic, D.; Assous, M.V.; Grimont, P.A.D.; Baranton, G. Diversity of Borrelia burgdorferi sensu lato evidenced by restriction fragment length polymorphism of rrf (5S)-rrl (23S) intergenic spacer amplicons. Int. J. Syst. Bacteriol. 1994, 44, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Bunikis, J.; Garpmo, U.; Tsao, J.; Berglund, J.; Fish, D.; Barbour, A.G. Sequence typing reveals extensive strain diversity of the Lyme borreliosis agents Borrelia burgdorferi in North America and Borrelia afzelii in Europe. Microbiology 2004, 150, 1741–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunikis, J.; Tsao, J.; Garpmo, U.; Berglund, J.; Fish, D.; Barbour, A.G. Typing of Borrelia relapsing fever group strains. Emerg. Infect. Dis. 2004, 10, 1661–1664. [Google Scholar] [CrossRef]
- Lambert, J.S.; Cook, M.J.; Healy, J.E.; Murtagh, R.; Avramovic, G.; Lee, S.H. Metagenomic 16S rRNA gene sequencing survey of Borrelia species in Irish samples of Ixodes ricinus ticks. PLoS ONE 2019, 14, e0209881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaenson, T.G.T.; Wilhelmsson, P. First records of tick-borne pathogens in populations of the taiga tick Ixodes persulcatus in Sweden. Parasites Vectors 2019, 12, 559. [Google Scholar] [CrossRef] [Green Version]
- Hovius, J.W.; de Wever, B.; Sohne, M.; Brouwer, M.C.; Coumou, J.; Wagemakers, A.; Oei, A.; Knol, H.; Narasimhan, S.; Hodiamont, C.J.; et al. A case of meningoencephalitis by the relapsing fever spirochaete Borrelia miyamotoi in Europe. Lancet 2013, 382, 658. [Google Scholar] [CrossRef] [Green Version]
- Fraenkel, C.-J.; Garpmo, U.; Berglund, J. Determination of novel Borrelia genospecies in Swedish Ixodes ricinus ticks. J. Clin. Microbiol. 2002, 40, 3308–3312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelmsson, P.; Lindblom, P.; Fryland, L.; Ernerudh, J.; Forsberg, P.; Lindgren, P.-E. Prevalence, diversity, and load of Borrelia species in ticks that have fed on humans in regions of Sweden and Åland Islands, Finland with different Lyme borreliosis incidences. PLoS ONE 2013, 8, e81433. [Google Scholar] [CrossRef] [Green Version]
- Fukunaga, M.; Koreki, Y. The flagellin gene of Borrelia miyamotoi sp. nov. and its phylogenetic relationship among Borrelia species. FEMS Microbiol. Lett. 1995, 134, 255–258. [Google Scholar] [CrossRef]
- Platonov, A.E.; Karan, L.S.; Kolyasnikova, N.M.; Makhneva, N.A.; Toporkova, M.G.; Maleev, V.V.; Fish, D.; Krause, P.J. Humans infected with relapsing fever spirochete Borrelia miyamotoi, Russia. Emerg. Infect. Dis. 2011, 17, 1816–1823. [Google Scholar] [CrossRef] [PubMed]
- Henningsson, A.J.; Asgeirsson, H.; Hammas, B.; Karlsson, E.; Parke, Å.; Hoornstra, D.; Wilhelmsson, P.; Hovius, J.W. Two cases of Borrelia miyamotoi meningitis, Sweden, 2018. Emerg. Infect. Dis. 2019, 25, 1965–1968. [Google Scholar] [CrossRef] [Green Version]
- Cutler, S.; Vayssier-Taussat, M.; Estrada-Peña, A.; Potkonjak, A.; Mihalca, A.D.; Zeller, H. A new Borrelia on the block: Borrelia miyamotoi—A human health risk? Eurosurveillance 2019, 24, 1800170. [Google Scholar] [CrossRef] [Green Version]
- Wilhelmsson, P.; Jaenson, T.G.T.; Olsen, B.; Waldenström, J.; Lindgren, P.-E. Migratory birds as disseminators of ticks and the tick-borne pathogens Borrelia bacteria and tick-borne encephalitis (TBE) virus: A seasonal study at Ottenby Bird Observatory in South-eastern Sweden. Parasites Vectors 2020, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ornstein, K.; Barbour, A.G. A reverse transcriptase–polymerase chain reaction assay of Borrelia burgdorferi 16S rRNA for highly sensitive quantification of pathogen load in a vector. Vector-Borne Zoonotic Dis. 2006, 6, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Lager, M.; Faller, M.; Wilhelmsson, P.; Kjelland, V.; Andreassen, Å.; Dargis, R.; Quarsten, H.; Dessau, R.; Fingerle, V.; Margos, G.; et al. Molecular detection of Borrelia burgdorferi sensu lato—An analytical comparison of real-time PCR protocols from five different Scandinavian laboratories. PLoS ONE 2017, 12, e0185434. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Target | Oligo Name | Sequence (5’→3’) | Amplicon Length (bp) | Reference |
|---|---|---|---|---|---|
| Borrelia spp. | 16S rRNA | Borrelia-F | GCTGAGTCACGAAAGCGTAG | 116 | [36] |
| Borrelia-R | CACTTAACACGTTAGCTTCGGTA | ||||
| Borrelia-P | FAM-CGCTGTAAACGATGCACACTTGGT-MGB | ||||
| 5S-23S rRNA IGS | B5S-23S_F | CTGCGAGTTCGCGGGAGA | 225–266 a | [38] | |
| B5S-23S_R | TCCTAGGCATTCACCATA | ||||
| B5S-23S_Fn | GAGTTCGCGGGAGAGTAA | [37] | |||
| B5S-23S_Rn | TAGGCATTCACCATAGACTCTT | ||||
| 16S-23S rRNA IGS | B16S-23S_F | GTATGTTTAGTGAGGGGGGTG | 388–685 a | [39,40] | |
| B16S-23S_R | GGATCATAGCTCAGGTGGTTAG | ||||
| B16S-23S_Fn | AGGGGGGTGAAGTCGTAACAAG | ||||
| B16S-23S_Rn | GTCTGATAAACCTGAGGTCGGA | ||||
| flaB | flaB-F | CATCTGATGATGCTGCTGGT | 699 | This study | |
| flaB-R | TGTTTTGGAAAGCACCAAGA | ||||
| flaB-Fn | GGGTGTTGCTGGGAAAATTA | 672 | |||
| flaB-Rn | TGGAAAGCACCAAGATTTGC | ||||
| 16S rRNA | M1 | ACGATGCACACTTGGTGTTAA | 357–358 a | [41] | |
| M2 | TCCGACTTATCACCGGCAGTC | ||||
| B. miyamotoi | flaB | Bm_F | AGAAGGTGCTCAAGCAG | 156 | [42] |
| Bm_R | TCGATCTTTGAAAGTGACATAT | ||||
| Bm_P | FAM-AGCACAACAGGAGGGAGTTCAAGC-BHQ1 |
| Tick Developmental Stage | No. of Ticks Examined | No. (%) of Borrelia-Positive Ticks by Real-Time PCR | No. of Specimens with Successful Amplification of Gene Targets by Conventional PCR | |||
|---|---|---|---|---|---|---|
| 5S-23S IGS | 16S-23S IGS | flaB | 16S rRNA | |||
| Larva | 31 | 12 (38.7) | 0 | 4 | 3 | 2 |
| Nymph | 48 | 7 (14.6) | 0 | 4 | 1 | 1 |
| Adult | 13 a | 3 (23.1) a | 0 | 3 a | 2 a | 1 |
| Total | 92 | 22 (23.9) | 0 | 11 | 6 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaenson, T.G.T.; Wilhelmsson, P. First Record of a Suspected Human-Pathogenic Borrelia Species in Populations of the Bat Tick Carios vespertilionis in Sweden. Microorganisms 2021, 9, 1100. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051100
Jaenson TGT, Wilhelmsson P. First Record of a Suspected Human-Pathogenic Borrelia Species in Populations of the Bat Tick Carios vespertilionis in Sweden. Microorganisms. 2021; 9(5):1100. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051100
Chicago/Turabian StyleJaenson, Thomas G. T., and Peter Wilhelmsson. 2021. "First Record of a Suspected Human-Pathogenic Borrelia Species in Populations of the Bat Tick Carios vespertilionis in Sweden" Microorganisms 9, no. 5: 1100. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051100