
Gram-Negative Bacteria Holding Together in a Biofilm: The Acinetobacter baumannii Way

 , ,
, ,  ,
,  , and
, and
Abstract
:1. Introduction
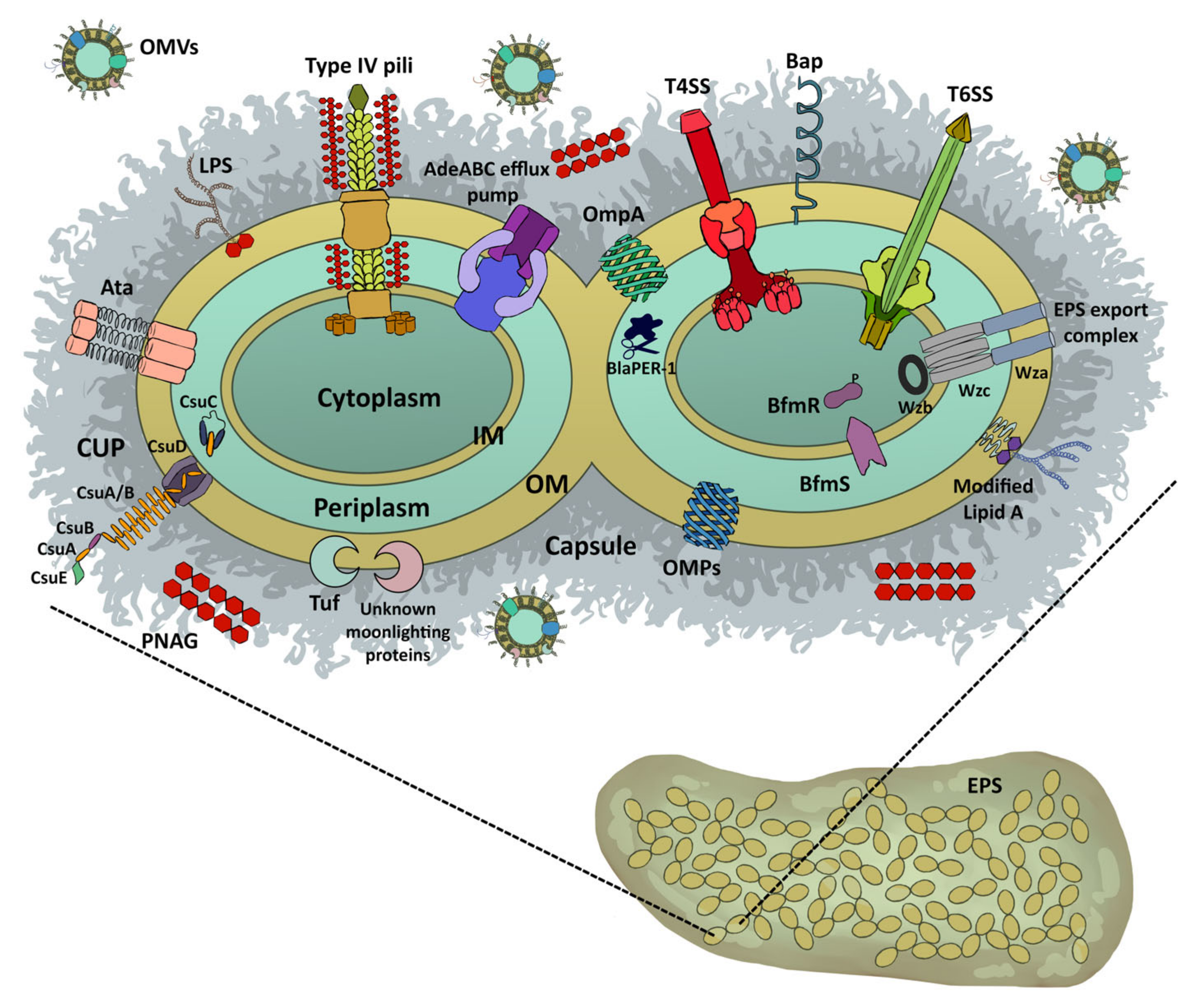
2. Mechanisms of Biofilm Formation
2.1. Role of Fimbrial Structures
2.2. Role of Afimbrial Structures
2.3. Role of Outer Membrane Vesicles (OMVs)
3. Are MDR Bacteria More Able to Form Biofilm?
3.1. The Success of A. baumannii Biofilms
3.2. Biofilms as a Source of Bacterial Dissemination
4. Clinical Impact of A. baumannii Biofilms
Device and Non-Device Related Biofilm Infections
5. Prevention and Treatment Strategies against A. baumannii Biofilm
5.1. Inhibition of Biofilm Formation
5.1.1. Modulation of Genes Involved in Biofilm Formation
5.1.2. Inhibition of QS Signals
5.1.3. Inhibition in EPS Production
5.1.4. Inhibition of Efflux Pumps
5.1.5. New Formulations of Antibiotics
5.1.6. Antibiofilm Antibodies
5.1.7. Carbon Monoxide Releasing Molecules (CORMs)
5.1.8. Iron Chelation
5.1.9. Antipersister Effects
5.2. Disrupting Preformed Biofilm
5.2.1. Peptides
5.2.2. Photodynamic Inactivation
5.2.3. Phage-Based Therapy
5.2.4. Potentiation of Antibiotics
5.2.5. Nanoparticles
5.2.6. Hydrogel-Based Formulations
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AFM | atomic force microscopy |
| AgNPs | silver nanoparticles |
| AgSD | silver sulfadiazine |
| AHL | N-acyl homoserine lactone |
| AMP | ampicillin |
| AMPs | antimicrobial peptides |
| AZM | azithromycin |
| Bap | biofilm-associated protein |
| CCCP | carbonyl cyanide 3-chlorophenylhydrazone |
| CFS | cell-free supernatant |
| CHs | chitosan hydrogels |
| CIP | ciprofloxacin |
| CLSM | confocal laser scanning microscopy |
| Col | colistin |
| CORMs | carbon monoxide releasing molecules |
| CSAs | ceragenins |
| CSH | cell surface hydrophobicity |
| CUP | chaperone-usher pili |
| CVC | central venous catheter |
| EPIs | efflux pump inhibitors |
| EPS | extracellular polymeric substance |
| ETEC | enterotoxigenic E. coli |
| FE-SEM | field-emission scanning electron microscopy |
| GN | gentamicin |
| HAIs | healthcare-acquired infections |
| haNPs | human albumin nanoparticles |
| IM | inner membrane |
| IMP | imipenem |
| MBEC-50 | minimum concentration that kills 50% of cells in preformed biofilm |
| MBEC-80 | minimum concentration that kills 80% of cells in preformed biofilm |
| MBEC | minimal biofilm eradication concentration |
| MBIC | minimal biofilm inhibitory concentration |
| MDR | multidrug resistant |
| MIC | minimal inhibitory concentration |
| MOI | multiplicity of infection |
| MRP | meropenem |
| NL | nanoliposomal |
| NMP | 1-(1-naphthylmethyl)-piperazine |
| NPs | nanoparticles |
| NS | not specified |
| 3-OH-C12-HSL | 3-hydroxy-dodecanoyl-L-homoserine lactone |
| OM | outer membrane |
| OMPs | outer membrane proteins |
| OMVs | outer membrane vesicles |
| PaβN | phenylalanine-arginine β-naphthylamide |
| PDI | photodynamic inactivation |
| PNAG | poly-β-(1-6)-N-acetylglucosamine |
| POL | polymyxin B |
| PS | photosensitizer |
| quorum quenching | |
| QS | quorum sensing |
| QSIs | QS inhibitors |
| RIF | rifampicin |
| RND | resistance nodulation-division |
| Sec | general secretory pathway |
| SEM | scanning electron microscopy |
| SXT | cotrimoxazole |
| T2SS | type II secretion system |
| T7SS | type VII secretion system |
| T8SS | type VIII secretion system |
| TIG | tigecycline |
| TOB | tobramycin |
| XDR | extensively drug resistant |
References
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Stewart, P.S.; Franklin, M.J. Physiological heterogeneity in biofilms. Nat. Rev. Microbiol. 2008, 6, 199–210. [Google Scholar] [CrossRef]
- Naseem, H.; Ahsan, M.; Shahid, M.A.; Khan, N. Exopolysaccharides producing rhizobacteria and their role in plant growth and drought tolerance. J. Basic Microbiol. 2018, 58, 1009–1022. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Besemer, K. Biodiversity, community structure and function of biofilms in stream ecosystems. Res. Microbiol. 2015, 166, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Van Houdt, R.; Michiels, C.W. Biofilm formation and the food industry, a focus on the bacterial outer surface. J. Appl Microbiol. 2010, 109, 1117–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, S.J.; Kjellerup, B.V. Applications of biofilms in bioremediation and biotransformation of persistent organic pollutants, pharmaceuticals/personal care products, and heavy metals. Appl. Microbiol. Biotechnol. 2013, 97, 9909–9921. [Google Scholar] [CrossRef] [PubMed]
- Karadag, D.; Köroğlu, O.E.; Özkaya, B.; Cakmakci, M. A review on anaerobic biofilm reactors for the treatment of dairy in-dustry wastewater. Process. Biochem. 2015, 50, 262–271. [Google Scholar] [CrossRef]
- Santoro, C.; Arbizzani, C.; Erable, B.; Ieropoulos, I. Microbial fuel cells: From fundamentals to applications. A review. J. Power Sources 2017, 356, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Moradali, M.F.; Rehm, B.H.A. Bacterial biopolymers: From pathogenesis to advanced materials. Nat. Rev. Microbiol. 2020, 18, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Shunmugaperumal, T. Microbial colonization of medical devices and novel preventive strategies. Recent Patents Drug Deliv. Formul. 2010, 4, 153–173. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.; Giri, P.A.; Saini, S.; Saini, S.R. Dental Plaque: A Complex Biofilm. Pravara Med. Rev. 2015, 7, 9–14. [Google Scholar]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as Complex Differentiated Communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [Green Version]
- Rossi, E.; Cimdins, A.; Lüthje, P.; Brauner, A.; Sjöling, Å.; Landini, P.; Romling, U. “It’s a gut feeling”—Escherichia coli biofilm formation in the gastrointestinal tract environment. Crit. Rev. Microbiol. 2018, 44, 1–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landini, P. Cross-talk mechanisms in biofilm formation and responses to environmental and physiological stress in Escherichia coli. Res. Microbiol. 2009, 160, 259–266. [Google Scholar] [CrossRef]
- Gunn, J.S.; Bakaletz, L.O.; Wozniak, D.J. What’s on the Outside Matters: The Role of the Extracellular Polymeric Substance of Gram-negative Biofilms in Evading Host Immunity and as a Target for Therapeutic Intervention. J. Biol. Chem. 2016, 291, 12538–12546. [Google Scholar] [CrossRef] [Green Version]
- Berne, C.; Ducret, A.; Hardy, G.; Brun, Y.V. Adhesins Involved in Attachment to Abiotic Surfaces by Gram-Negative Bacteria. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scribano, D.; Sarshar, M.; Prezioso, C.; Lucarelli, M.; Angeloni, A.; Zagaglia, C.; Palamara, A.T.; Ambrosi, C. D-Mannose Treatment neither Affects Uro-pathogenic Escherichia coli Properties nor Induces Stable FimH Modifications. Molecules 2020, 25, 316. [Google Scholar] [CrossRef] [Green Version]
- Sarshar, M.; Behzadi, P.; Ambrosi, C.; Zagaglia, C.; Palamara, A.T.; Scribano, D. FimH and Anti-Adhesive Therapeutics: A Disarming Strategy Against Uropathogens. Antibiotics 2020, 9, 397. [Google Scholar] [CrossRef] [PubMed]
- Ambrosi, C.; Scribano, D.; Aleandri, M.; Zagaglia, C.; Di Francesco, L.; Putignani, L.; Palamara, A.T. Acinetobacter baumannii Virulence Traits: A Comparative Study of a Novel Sequence Type with Other Italian Endemic International Clones. Front. Microbiol. 2017, 8, 1977. [Google Scholar] [CrossRef]
- Chagnot, C.; Zorgani, M.A.; Astruc, T.; Desvaux, M. Proteinaceous determinants of surface colonization in bacteria: Bacterial adhesion and biofilm formation from a protein secretion perspective. Front. Microbiol. 2013, 4, 303. [Google Scholar] [CrossRef] [Green Version]
- Zav’yalov, V.; Zavialov, A.; Zav’yalova, G.; Korpela, T. Adhesive organelles of Gram-negative pathogens assembled with the classical chaperone/usher machinery: Structure and function from a clinical standpoint. FEMS Microbiol. Rev. 2010, 34, 317–378. [Google Scholar] [CrossRef] [Green Version]
- Desvaux, M.; Hébraud, M.; Talon, R.; Henderson, I.R. Secretion and subcellular localizations of bacterial proteins: A semantic awareness issue. Trends Microbiol. 2009, 17, 139–145. [Google Scholar] [CrossRef]
- Nuccio, S.-P.; Baäumler, A.J. Evolution of the Chaperone/Usher Assembly Pathway: Fimbrial Classification Goes Greek. Microbiol. Mol. Biol. Rev. 2007, 71, 551–575. [Google Scholar] [CrossRef] [Green Version]
- Hospenthal, M.K.; Costa, T.R.D.; Waksman, G. A comprehensive guide to pilus biogenesis in Gram-negative bacteria. Nat. Rev. Microbiol. 2017, 15, 365–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarshar, M.; Behzadi, P.; Scribano, D.; Palamara, A.; Ambrosi, C. Acinetobacter baumannii: An Ancient Commensal with Weapons of a Pathogen. Pathogens 2021, 10, 387. [Google Scholar] [CrossRef] [PubMed]
- Hospenthal, M.K.; Waksman, G. The Remarkable Biomechanical Properties of the Type 1 Chaperone-Usher Pilus: A Structural and Molecular Perspective. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- St Geme, J.W., 3rd; Cutter, D. Influence of pili, fibrils, and capsule on in vitro adherence by Haemophilus influenzae type b. Mol. Microbiol. 1996, 21, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Vallet, I.; Olson, J.W.; Lory, S.; Lazdunski, A.; Filloux, A. The chaperone/usher pathways of Pseudomonas aeruginosa: Identifi-cation of fimbrial gene clusters (cup) and their involvement in biofilm formation. Proc. Natl. Acad. Sci. USA 2001, 98, 6911–6916. [Google Scholar] [CrossRef] [Green Version]
- Gohl, O.; Friedrich, A.; Hoppert, M.; Averhoff, B. The Thin Pili of Acinetobacter sp. Strain BD413 Mediate Adhesion to Biotic and Abiotic Surfaces. Appl. Environ. Microbiol. 2006, 72, 1394–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, X.; Zhu, W.; Jin, J.; Mao, X. Evidence that a chaperone–usher-like pathway of Myxococcus xanthus functions in spore coat formation. Microbiology 2011, 157, 1886–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inhülsen, S.; Aguilar, C.; Schmid, N.; Suppiger, A.; Riedel, K.; Eberl, L. Identification of functions linking quorum sensing with biofilm formation in Burkholderia cenocepacia H111. Microbiol. Open 2012, 1, 225–242. [Google Scholar] [CrossRef] [Green Version]
- Ormeño-Orrillo, E.; Menna, P.; Almeida, L.G.P.; Ollero, F.J.; Nicolás, M.F.; Rodrigues, E.P.; Nakatani, A.S.; Batista, J.S.S.; Chueire, L.M.O.; Souza, R.C.; et al. Genomic basis of broad host range and environmental adaptability of Rhizobium tropici CIAT 899 and Rhizobium sp. PRF 81 which are used in inoculants for common bean (Phaseolus vulgaris L.). BMC Genom. 2012, 13, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudirkienė, E.; Bager, R.J.; Johnson, T.J.; Bojesen, A.M. Chaperone-usher fimbriae in a diverse selection of Gallibacterium ge-nomes. BMC Genom. 2014, 15, 1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psonis, J.J.; Thanassi, D.G. Therapeutic Approaches Targeting the Assembly and Function of Chaperone-Usher Pili. EcoSal Plus 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Ageorges, V.; Monteiro, R.; Leroy, S.; Burgess, C.M.; Pizza, M.; Chaucheyras-Durand, F.; Desvaux, M. Molecular determinants of surface colonisation in diarrhoeagenic Escherichia coli (DEC): From bacterial adhesion to biofilm formation. FEMS Microbiol. Rev. 2020, 44, 314–350. [Google Scholar] [CrossRef]
- Kimkes, T.E.P.; Heinemann, M. How bacteria recognise and respond to surface contact. FEMS Microbiol. Rev. 2020, 44, 106–122. [Google Scholar] [CrossRef]
- Sarshar, M.; Scribano, D.; Marazzato, M.; Ambrosi, C.; Aprea, M.R.; Aleandri, M.; Pronio, A.; Longhi, C.; Nicoletti, M.; Zagaglia, C.; et al. Genetic diversity, phylogroup distribution and virulence gene profile of pks positive Escherichia coli colonizing human intestinal polyps. Microb. Pathog. 2017, 112, 274–278. [Google Scholar] [CrossRef]
- Ambrosi, C.; Sarshar, M.; Aprea, M.R.; Pompilio, A.; Di Bonaventura, G.; Strati, F.; Pronio, A.; Nicoletti, M.; Zagaglia, C.; Palamara, A.T.; et al. Colonic adenoma-associated Escherichia coli express specific phenotypes. Microbes Infect. 2019, 21, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Ayoub Moubareck, C.; Hammoudi Halat, D. Insights into Acinetobacter baumannii: A Review of Microbiological, Virulence, and Resistance Traits in a Threatening Nosocomial Pathogen. Antibiotics 2020, 9, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakharukova, N.; Tuittila, M.; Paavilainen, S.; Malmi, H.; Parilova, O.; Teneberg, S.; Knight, S.D.; Zavialov, A.V. Structural basis for Acinetobacter baumannii biofilm formation. Proc. Natl. Acad. Sci. USA 2018, 115, 5558–5563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaras, A.P.; Flagler, M.J.; Dorsey, C.W.; Gaddy, J.A.; Actis, L.A. Characterization of a two-component regulatory system from Aci-netobacter baumannii that controls biofilm formation and cellular morphology. Microbiology 2008, 154, 3398–3409. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-H.; Lee, H.-W.; Kim, S.-M.; Kim, J. Proteomic analysis of Acinetobacter baumannii in biofilm and planktonic growth mode. J. Microbiol. 2009, 47, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Liou, M.L.; Soo, P.C.; Ling, S.R.; Kuo, H.Y.; Tang, C.Y.; Chang, K.C. The sensor kinase BfmS mediates virulence in Acinetobacter baumannii. J. Microbiol. Immunol. Infect. 2014, 47, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef]
- Cerqueira, G.M.; Kostoulias, X.; Khoo, C.; Aibinu, I.; Qu, Y.; Traven, A.; Peleg, A. A Global Virulence Regulator in Acinetobacter baumannii and Its Control of the Phenylacetic Acid Catabolic Pathway. J. Infect. Dis. 2014, 210, 46–55. [Google Scholar] [CrossRef] [Green Version]
- De Silva, P.M.; Kumar, A. Signal Transduction Proteins in Acinetobacter baumannii: Role in Antibiotic Resistance, Virulence, and Potential as Drug Targets. Front. Microbiol. 2019, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Winterhalter, M.; Pagès, J.-M. Outer Membrane Porins. Subcell Biochem. 2019, 92, 79–123. [Google Scholar] [CrossRef]
- Confer, A.W.; Ayalew, S. The OmpA family of proteins: Roles in bacterial pathogenesis and immunity. Vet. Microbiol. 2013, 163, 207–222. [Google Scholar] [CrossRef]
- Scribano, D.; Damico, R.; Ambrosi, C.; Superti, F.; Marazzato, M.; Conte, M.P.; Longhi, C.; Palamara, A.T.; Zagaglia, C.; Nicoletti, M. The Shigella flexneri OmpA amino acid residues 188 EVQ 190 are essential for the interaction with the virulence factor PhoN2. Biochem. Biophys. Rep. 2016, 8, 168–173. [Google Scholar] [CrossRef] [Green Version]
- Scribano, D.; Petrucca, A.; Pompili, M.; Ambrosi, C.; Bruni, E.; Zagaglia, C.; Prosseda, G.; Nencioni, L.; Casalino, M.; Polticelli, F.; et al. Polar Localization of PhoN2, a Periplasmic Virulence-Associated Factor of Shigella flexneri, Is Required for Proper IcsA Exposition at the Old Bacterial Pole. PLoS ONE 2014, 9, e90230. [Google Scholar] [CrossRef]
- Ambrosi, C.; Pompili, M.; Scribano, D.; Zagaglia, C.; Ripa, S.; Nicoletti, M. Outer Membrane Protein A (OmpA): A New Player in Shigella flexneri Protrusion Formation and Inter-Cellular Spreading. PLoS ONE 2012, 7, e49625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaddy, J.A.; Tomaras, A.P.; Actis, L.A. The Acinetobacter baumannii 19606 OmpA Protein Plays a Role in Biofilm Formation on Abiotic Surfaces and in the Interaction of This Pathogen with Eukaryotic Cells. Infect. Immun. 2009, 77, 3150–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Wood, T.K. OmpA influences Escherichia coli biofilm formation by repressing cellulose production through the CpxRA two-component system. Environ. Microbiol. 2009, 11, 2735–2746. [Google Scholar] [CrossRef]
- Zeighami, H.; Valadkhani, F.; Shapouri, R.; Samadi, E.; Haghi, F. Virulence characteristics of multidrug resistant biofilm forming Acinetobacter baumannii isolated from intensive care unit patients. BMC Infect. Dis. 2019, 19, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flannery, A.; Le Berre, M.; Pier, G.B.; O’Gara, J.P.; Kilcoyne, M. Glycomics Microarrays Reveal Differential In Situ Presentation of the Biofilm Polysaccharide Poly-N-acetylglucosamine on Acinetobacter baumannii and Staphylococcus aureus Cell Surfaces. Int. J. Mol. Sci. 2020, 21, 2465. [Google Scholar] [CrossRef] [Green Version]
- Geisinger, E.; Mortman, N.J.; Vargas-Cuebas, G.; Tai, A.K.; Isberg, R.R. A global regulatory system links virulence and antibiotic re-sistance to envelope homeostasis in Acinetobacter baumannii. PLoS Pathog. 2018, 14, e1007030. [Google Scholar] [CrossRef]
- Geisinger, E.; Isberg, R.R. Antibiotic Modulation of Capsular Exopolysaccharide and Virulence in Acinetobacter baumannii. PLOS Pathog. 2015, 11, e1004691. [Google Scholar] [CrossRef] [Green Version]
- Goh, H.M.S.; Beatson, S.A.; Totsika, M.; Moriel, D.G.; Phan, M.-D.; Szubert, J.; Runnegar, N.; Sidjabat, H.E.; Paterson, D.L.; Nimmo, G.R.; et al. Molecular Analysis of the Acinetobacter baumannii Biofilm-Associated Protein. Appl. Environ. Microbiol. 2013, 79, 6535–6543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brossard, K.A.; Campagnari, A.A. The Acinetobacter baumannii Biofilm-Associated Protein Plays a Role in Adherence to Human Epithelial Cells. Infect. Immun. 2011, 80, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Loehfelm, T.W.; Luke, N.R.; Campagnari, A.A. Identification and Characterization of an Acinetobacter baumannii Bio-film-Associated Protein. J. Bacteriol. 2007, 190, 1036–1044. [Google Scholar] [CrossRef] [Green Version]
- Guérin, J.; Bigot, S.; Schneider, R.; Buchanan, S.K.; Jacob-Dubuisson, F. Two-Partner Secretion: Combining Efficiency and Simplicity in the Secretion of Large Proteins for Bacteria-Host and Bacteria-Bacteria Interactions. Front. Cell. Infect. Microbiol. 2017, 7, 148. [Google Scholar] [CrossRef]
- Bentancor, L.V.; Camacho-Peiro, A.; Bozkurt-Guzel, C.; Pier, G.B.; Maira-Litrán, T. Identification of Ata, a Multifunctional Trimeric Autotransporter of Acinetobacter baumannii. J. Bacteriol. 2012, 194, 3950–3960. [Google Scholar] [CrossRef] [Green Version]
- Pérez, A.; Merino, M.; Rumbo-Feal, S.; Álvarez-Fraga, L.; Vallejo, J.A.; Beceiro, A.; Ohneck, E.J.; Mateos, J.; Fernandez-Puente, P.; Actis, L.A.; et al. The FhaB/FhaC two-partner secretion system is involved in adhesion of Acinetobacter baumannii AbH12O-A2 strain. Virulence 2017, 8, 959–974. [Google Scholar] [CrossRef] [Green Version]
- Kishii, K.; Hamada, M.; Aoki, K.; Ito, K.; Onodera, J.; Ishii, Y.; Tateda, K. Differences in biofilm formation and transcription of bio-film-associated genes among Acinetobacter baumannii clinical strains belonging to the international clone II lineage. J. Infect. Chemother. 2020, 26, 693–698. [Google Scholar] [CrossRef]
- Saipriya, K.; Swathi, C.; Ratnakar, K.; Sritharan, V.; Kamaraju, S.; Ch, S. Quorum-sensing system in Acinetobacter baumannii: A potential target for new drug development. J. Appl. Microbiol. 2019, 128, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi Badi, S.; Moshiri, A.; Fateh, A.; Rahimi Jamnani, F.; Sarshar, M.; Vaziri, F.; Siadat, S.D. Microbiota-Derived Extracellular Vesicles as New Systemic Regulators. Front. Microbiol. 2017, 8, 1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarshar, M.; Scribano, D.; Ambrosi, C.; Palamara, A.T.; Masotti, A. Fecal microRNAs as Innovative Biomarkers of Intestinal Diseases and Effective Players in Host-Microbiome Interactions. Cancers 2020, 12, 2174. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, O.Y.; Gho, Y.S. Proteomic profiling of Gram-negative bacterial outer membrane vesicles: Current perspectives. Pro-teom. Clin. Appl. 2016, 10, 897–909. [Google Scholar] [CrossRef]
- Weber, B.S.; Kinsella, R.L.; Harding, C.M.; Feldman, M.F. The Secrets of Acinetobacter Secretion. Trends Microbiol. 2017, 25, 532–545. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, M.H.; Kim, S.I.; Son, J.H.; Kim, S.; Lee, Y.C.; Shin, M.; Oh, M.H.; Lee, J.C. The sensor kinase BfmS controls production of outer membrane vesicles in Acinetobacter baumannii. BMC Microbiol. 2019, 19, 301. [Google Scholar] [CrossRef]
- Ahmad, I.; Karah, N.; Nadeem, A.; Wai, S.N.; Uhlin, B.E. Analysis of colony phase variation switch in Acinetobacter baumannii clinical isolates. PLoS ONE 2019, 14, e0210082. [Google Scholar] [CrossRef]
- Ebner, P.; Götz, F. Bacterial Excretion of Cytoplasmic Proteins (ECP): Occurrence, Mechanism, and Function. Trends Microbiol. 2019, 27, 176–187. [Google Scholar] [CrossRef]
- Koenigs, A.; Zipfel, P.F.; Kraiczy, P. Translation Elongation Factor Tuf of Acinetobacter baumannii Is a Plasminogen-Binding Protein. PLoS ONE 2015, 10, e0134418. [Google Scholar]
- Kainulainen, V.; Korhonen, T.K. Dancing to Another Tune—Adhesive Moonlighting Proteins in Bacteria. Biology 2014, 3, 178–204. [Google Scholar] [CrossRef] [Green Version]
- Balaban, N.Q.; Merrin, J.; Chait, R.; Kowalik, L.; Leibler, S. Bacterial Persistence as a Phenotypic Switch. Science 2004, 305, 1622–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittershaus, E.S.; Baek, S.-H.; Sassetti, C.M. The Normalcy of Dormancy: Common Themes in Microbial Quiescence. Cell Host Microbe 2013, 13, 643–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.X.; Wang, C.Y.; Li, Y.Y.; Li, J.; Wan, Q.Q.; Chen, J.H.; Tay, F.R.; Niu, L.-N. Considerations and Caveats in Combating ESKAPE Pathogens against Nosocomial Infections. Adv. Sci. 2020, 7, 1901872. [Google Scholar] [CrossRef] [Green Version]
- Ambrosi, C.; Aleandri, M.; Giordano, A.; Scribano, D.; Marazzato, M.; Zagaglia, C.; Conte, M.P.; Palamara, A.T. Molecular characterisation of extensively drug-resistant Acinetobacter baumannii: First report of a new sequence type in Italy. J. Glob. Antimicrob. Resist. 2016, 7, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Gayoso, C.M.; Mateos, J.; Méndez, J.A.; Fernández-Puente, P.; Rumbo, C.; Tomás, M.; de Ilarduya, O.M.; Bou, G. Molecular mechanisms involved in the re-sponse to desiccation stress and persistence in Acinetobacter baumannii. J. Proteome Res. 2014, 13, 460–476. [Google Scholar] [CrossRef]
- Richmond, G.E.; Evans, L.P.; Anderson, M.J.; Wand, M.E.; Bonney, L.C.; Ivens, A.; Chua, K.L.; Webber, M.A.; Sutton, J.M.; Peterson, M.L.; et al. Erratum for Richmond et al., The Acinetobacter baumannii Two-Component System AdeRS Regulates Genes Required for Multidrug Efflux, Biofilm Formation, and Virulence in a Strain-Specific Manner. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boll, J.M.; Tucker, A.T.; Klein, D.R.; Beltran, A.M.; Brodbelt, J.S.; Davies, B.; Trent, M.S. Reinforcing Lipid A Acylation on the Cell Surface of Acinetobacter baumannii Promotes Cationic Antimicrobial Peptide Resistance and Desiccation Survival. mBio 2015, 6, e00478-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalabaev, S.; Chauhan, A.; Novikov, A.; Iyer, P.; Szczesny, M.; Beloin, C.; Caroff, M.; Ghigo, J.-M. Biofilms Formed by Gram-Negative Bacteria Undergo Increased Lipid A Palmitoylation, Enhancing In Vivo Survival. mBio 2014, 5, e01116-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farshadzadeh, Z.; Taheri, B.; Rahimi, S.; Shoja, S.; Pourhajibagher, M.; Haghighi, M.A.; Bahador, A. Growth Rate and Biofilm Formation Ability of Clinical and Laboratory-Evolved Colistin-Resistant Strains of Acinetobacter baumannii. Front. Microbiol. 2018, 9, 153. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, G.J.; Domingues, S. Interplay between Colistin Resistance, Virulence and Fitness in Acinetobacter baumannii. Antibiotics 2017, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Lu, F.; Yuan, F.; Jiang, D.; Zhao, P.; Zhu, J.; Cheng, H.; Cao, J.; Lu, G. Biofilm Formation Caused by Clinical Acinetobacter baumannii Isolates Is Associated with Overexpression of the AdeFGH Efflux Pump. Antimicrob. Agents Chemother. 2015, 59, 4817–4825. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Unno, Y.; Ubagai, T.; Ono, Y. Sub-minimum inhibitory concentrations of colistin and polymyxin B promote Acineto-bacter baumannii biofilm formation. PLoS ONE 2018, 13, e0194556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, C.; Clemmer, K.M.; Bonomo, R.A.; Rather, P.N. Isolation and Characterization of an Autoinducer Synthase from Aci-netobacter baumannii. J. Bacteriol. 2008, 190, 3386–3392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.H.; Park, E.C.; Lee, S.-Y.; Lee, H.; Choi, C.-W.; Yi, Y.-S.; Ro, H.-J.; Lee, J.C.; Jun, S.; Kim, H.-Y.; et al. Antibiotic treatment modulates protein components of cytotoxic outer membrane vesicles of multidrug-resistant clinical strain, Acinetobacter bau-mannii DU202. Clin. Proteom. 2018, 15, 28. [Google Scholar] [CrossRef] [PubMed]
- Scribano, D.; Marzano, V.; Mortera, S.L.; Sarshar, M.; Vernocchi, P.; Zagaglia, C.; Putignani, L.; Palamara, A.T.; Ambrosi, C. Insights into the Periplasmic Proteins of Acinetobacter baumannii AB5075 and the Impact of Imipenem Exposure: A Proteomic Approach. Int. J. Mol. Sci. 2019, 20, 3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, C.; Vadlamudi, G.; Newton, D.; Foxman, B.; Xi, C. The influence of biofilm formation and multidrug resistance on envi-ronmental survival of clinical and environmental isolates of Acinetobacter baumannii. Am. J. Infect. Control. 2016, 44, e65–e71. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Baño, J.; Marti, S.; Soto, S.M.; Fernández-Cuenca, F.; Cisneros, J.; Pachón, J.; Pascual, A.; Martínez, L.M.; McQueary, C.; Actis, L.; et al. Biofilm formation in Acinetobacter baumannii: Associated features and clinical implications. Clin. Microbiol. Infect. 2008, 14, 276–278. [Google Scholar] [CrossRef] [Green Version]
- Ambrosi, C.; Scribano, D.; Sarshar, M.; Zagaglia, C.; Singer, B.B.; Palamara, A.T. Acinetobacter baumannii Targets Human Carci-noembryonic Antigen-Related Cell Adhesion Molecules (CEACAMs) for Invasion of Pneumocytes. mSystems 2020, 5, e00604-20. [Google Scholar] [CrossRef] [PubMed]
- Azizi, O.; Shahcheraghi, F.; Salimizand, H.; Modarresi, F.; Shakibaie, M.R.; Mansouri, S.; Ramazanzadeh, R.; Badmasti, F.; Nikbin, V. Molecular Analysis and Expression of bap Gene in Biofilm-Forming Multi-Drug-Resistant Acinetobacter baumannii. Rep. Biochem. Mol. Biol. 2016, 5, 62–72. [Google Scholar]
- Thummeepak, R.; Kongthai, P.; Leungtongkam, U.; Sitthisak, S. Distribution of virulence genes involved in biofilm formation in multi-drug resistant Acinetobacter baumannii clinical isolates. Int. Microbiol. 2016, 19, 121–129. [Google Scholar]
- Aliramezani, A.; Douraghi, M.; Hajihasani, A.; Mohammadzadeh, M.; Rahbar, M. Clonal relatedness and biofilm formation of OXA-23-producing carbapenem resistant Acinetobacter baumannii isolates from hospital environment. Microb. Pathog. 2016, 99, 204–208. [Google Scholar] [CrossRef]
- Amin, M.; Navidifar, T.; Shooshtari, F.S.; Rashno, M.; Savari, M.; Jahangirmehr, F.; Arshadi, M. Association Between Biofilm Formation, Structure, and the Expression Levels of Genes Related to biofilm formation and Biofilm-Specific Resistance of Acinetobacter bau-mannii Strains Isolated from Burn Infection in Ahvaz, Iran. Infect. Drug Resist. 2019, 12, 3867–3881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-H.; Su, P.-W.; Moi, S.-H.; Chuang, L.-Y. Biofilm Formation in Acinetobacter Baumannii: Genotype-Phenotype Cor-relation. Molecules 2019, 24, 1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahdouh, E.; Orgaz, B.; Gomez-Gil, R.; Mingorance, J.; Daoud, Z.; Suárez, M.; José, C.S. Patterns of biofilm structure and formation kinetics among Acinetobacter baumannii clinical isolates with different antibiotic resistance profiles. MedChemComm 2016, 7, 157–163. [Google Scholar] [CrossRef]
- Moscoso, M.; García, E. Pneumococcal biofilms. Int. Microbiol. 2009, 1, 77–85. [Google Scholar]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial Biofilms: Development, Dispersal, and Therapeutic Strategies in the Dawn of the Postantibiotic Era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef] [Green Version]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Rumbaugh, K.P.; Sauer, K. Biofilm dispersion. Nat. Rev. Microbiol. 2020, 18, 571–586. [Google Scholar] [CrossRef]
- Berlanga, M.; Guerrero, R. Living together in biofilms: The microbial cell factory and its biotechnological implications. Microb. Cell Fact. 2016, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Yamamoto, N.; Kino, Y.; Yamamoto, N.; Kamei, S.; Mori, H.; Kurokawa, K.; Nakashima, N. Establishment of a multi-species biofilm model and metatranscriptomic analysis of biofilm and planktonic cell communities. Appl. Microbiol. Biotechnol. 2016, 100, 7263–7279. [Google Scholar] [CrossRef]
- Caro-Astorga, J.; Frenzel, E.; Perkins, J.R.; Álvarez-Mena, A.; de Vicente, A.; Ranea, J.A.G.; Kuipers, O.P.; Romero, D. Biofilm formation displays intrinsic offensive and defensive features of Bacillus cereus. NPJ Biofilms Microbiomes 2020, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Chapartegui-González, I.; Lázaro-Díez, M.; Bravo, Z.; Navas, J.; Icardo, J.M.; Ramos-Vivas, J. Acinetobacter baumannii maintains its virulence after long-time starvation. PLoS ONE 2018, 13, e0201961. [Google Scholar] [CrossRef]
- Ivanković, T.; Goić-Barišić, I.; Hrenović, J. Reduced susceptibility to disinfectants of Acinetobacter baumannii biofilms on glass and ceramic. Arh. Hig. Rada Toksikol. 2017, 68, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Chiang, S.R.; Jung, F.; Tang, H.J.; Chen, C.H.; Chen, C.C.; Chou, H.Y.; Chuang, Y.C. Desiccation and ethanol resistances of multidrug resistant Acinetobacter baumannii embedded in biofilm: The favorable antiseptic efficacy of combination chlorhexidine gluconate and ethanol. J. Microbiol. Immunol. Infect. 2018, 51, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.; Johani, K.; Melo, D.; Lopes, L.; Lima, L.L.; Tipple, A.; Hu, H.; Vickery, K. Biofilm contamination of high-touched surfaces in intensive care units: Epidemiology and potential impacts. Lett. Appl. Microbiol. 2019, 68, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asati, S.; Chaudhary, U. Prevalence of biofilm producing aerobic bacterial isolates in burn wound infections at a tertiary care hospital in northern India. Ann. Burn. Fire Disasters 2017, 30, 39–42. [Google Scholar]
- Szczypta, A.; Talaga-Ćwiertnia, K.; Kielar, M.; Krzyściak, P.; Gajewska, A.; Szura, M.; Bulanda, M.; Chmielarczyk, A. Inves-tigation of Acinetobacter baumannii Activity in Vascular Surgery Units through Epidemiological Management Based on the Analysis of Antimicrobial Resistance, Biofilm Formation and Genotyping. Int. J. Environ. Res. Public Health 2021, 18, 1563. [Google Scholar] [CrossRef] [PubMed]
- Vuotto, C.; Grosso, F.; Longo, F.; Balice, M.P.; De Barros, M.C.; Peixe, L.; Donelli, G. Biofilm-Forming Ability and Clonality in Acinetobacter baumannii Strains Isolated from Urine Samples and Urinary Catheters in Different European Hospitals. Adv. Exp. Med. Biol. 2017, 1057, 73–83. [Google Scholar] [CrossRef]
- Kim, M.; Park, J.; Park, W. Genomic and phenotypic analyses of multidrug-resistant Acinetobacter baumannii NCCP 16007 iso-lated from a patient with a urinary tract infection. Virulence 2021, 12, 150–164. [Google Scholar] [CrossRef]
- Zhang, Y.; Brackman, G.; Coenye, T. Pitfalls associated with evaluating enzymatic quorum quenching activity: The case of MomL and its effect on Pseudomonas aeruginosa and Acinetobacter baumannii biofilms. PeerJ 2017, 5, e3251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alamri, A.M.; Alsultan, A.A.; Ansari, M.A.; Alnimr, A.M. Biofilm-Formation in Clonally Unrelated Multidrug-Resistant Aci-netobacter baumannii Isolates. Pathogens 2020, 9, 630. [Google Scholar] [CrossRef] [PubMed]
- Asaad, A.M.; Ansari, S.; Ajlan, S.E.; Awad, S.M. Epidemiology of Biofilm Producing Acinetobacter baumannii Nosocomial Iso-lates from a Tertiary Care Hospital in Egypt: A Cross-Sectional Study. Infect. Drug Resist. 2021, 14, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.Y.; Baek, W.-K.; Kim, H.A. Association of biofilm production with colonization among clinical isolates of Acinetobacter baumannii. Korean J. Intern. Med. 2017, 32, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, D.P.; Wang, Q.; Weinberg, A.; Lamont, R.J. Transcriptome analysis of Porphyromonas gingivalis and Acinetobacter bau-mannii in polymicrobial communities. Mol. Oral Microbiol. 2018, 33, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; He, L.; Tao, X.; Meng, F.; Zhang, J. Biofilm may not be Necessary for the Epidemic Spread of Acinetobacter baumannii. Sci. Rep. 2016, 6, 32066. [Google Scholar] [CrossRef] [PubMed]
- Sechi, L.A.; Karadenizli, A.; Deriu, A.; Zanetti, S.; Kolayli, F.; Balikci, E.; Vahaboglu, H. PER-1 type beta-lactamase production in Acinetobacter baumannii is related to cell adhesion. Med. Sci. Monit. 2004, 10, BR180–BR184. [Google Scholar] [PubMed]
- Kaliterna, V.; Goic-Barisic, I. The ability of biofilm formation in clinical isolates of Acinetobacter baumannii belonging to two different European clones causing outbreaks in the Split University Hospital, Croatia. J. Chemother. 2013, 25, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.J.; Mende, K.; Beckius, M.L.; Akers, K.S.; Romano, D.R.; Wenke, J.C.; Murray, C.K. Biofilm formation by clinical isolates and the implications in chronic infections. BMC Infect. Dis. 2013, 13, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talreja, D.; Kaye, K.S.; Yu, F.-S.; Walia, S.K.; Kumar, A. Pathogenicity of Ocular Isolates of Acinetobacter baumannii in a Mouse Model of Bacterial Endophthalmitis. Investig. Opthalmol. Vis. Sci. 2014, 55, 2392–2402. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.G.; Black, C.C.; Pavlicek, R.L.; Honnold, C.L.; Wise, M.C.; Alamneh, Y.A.; Moon, J.K.; Kessler, J.L.; Si, Y.; Williams, R.; et al. Validation of a Novel Murine Wound Model of Acinetobacter baumannii Infection. Antimicrob. Agents Chemother. 2013, 58, 1332–1342. [Google Scholar] [CrossRef] [Green Version]
- Djeribi, R.; Bouchloukh, W.; Jouenne, T.; Menaa, B. Characterization of bacterial biofilms formed on urinary catheters. Am. J. Infect. Control. 2012, 40, 854–859. [Google Scholar] [CrossRef]
- McQueary, C.N.; Actis, L.A. Acinetobacter baumannii biofilms: Variations among strains and correlations with other cell prop-erties. J. Microbiol. 2011, 49, 243–250. [Google Scholar] [CrossRef]
- Lin, M.-F.; Lin, Y.-Y.; Lan, C.-Y. A method to assess influence of different medical tubing on biofilm formation by Acinetobacter baumannii. J. Microbiol. Methods 2019, 160, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Gil-Perotin, S.; Ramirez, P.; Marti, V.; Sahuquillo, J.M.; Gonzalez, E.; Calleja, I.; Menendez, R.; Bonastre, J. Implications of endotracheal tube biofilm in ventilator-associated pneumonia response: A state of concept. Crit. Care 2012, 16, R93. [Google Scholar] [CrossRef] [Green Version]
- Ferreira Tde, O.; Koto, R.Y.; Leite, G.F.; Klautau, G.B.; Nigro, S.; da Silva, C.B.; Idalgo da Fonseca Souza, A.P.; Mimica, M.J.; Grigolli, R.; Costa Salles, C.M.S. Microbial investigation of biofilms recovered from endotracheal tubes using sonication in intensive care unit pediatric patients. Braz. J. Infect. Dis. 2016, 20, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Pour, N.K.; Dusane, D.H.; Dhakephalkar, P.K.; Zamin, F.R.; Zinjarde, S.S.; Chopade, B.A. Biofilm formation by Acinetobacter baumannii strains isolated from urinary tract infection and urinary catheters. FEMS Immunol. Med. Microbiol. 2011, 62, 328–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asadian, M.; Azimi, L.; Alinejad, F.; Ostadi, Y.; Lari, A.R. Molecular Characterization of Acinetobacter baumannii Isolated from Ventilator-Associated Pneumonia and Burn Wound Colonization by Random Amplified Polymorphic DNA Polymerase Chain Reaction and the Relationship between Antibiotic Susceptibility and Biofilm Production. Adv. Biomed. Res. 2019, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Yeo, H.J.; Yoon, S.H.; Lee, S.E.; Cho, W.H.; Kim, D.; Jeon, D.; Shin, K.-H.; Kim, Y.S. Bacterial Biofilms on Extracorporeal Membrane Oxygenation Catheters. ASAIO J. 2018, 64, e48–e54. [Google Scholar] [CrossRef] [PubMed]
- Abirami, G.; Durgadevi, R.; Velmurugan, P.; Ravi, A.V. Gene expressing analysis indicates the role of Pyrogallol as a novel anti-biofilm and antivirulence agent against Acinetobacter baumannii. Arch. Microbiol. 2021, 203, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, A.; Valliammai, A.; Sivasankar, C.; Suba, M.; Sakthivel, G.; Pandian, S.K. Antibiofilm and antivirulence efficacy of myrtenol enhances the antibiotic susceptibility of Acinetobacter baumannii. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Raorane, C.J.; Lee, J.H.; Kim, Y.G.; Rajasekharan, S.K.; García-Contreras, R.; Lee, J. Antibiofilm and Antivirulence Efficacies of Fla-vonoids and Curcumin Against Acinetobacter baumannii. Front. Microbiol. 2019, 10, 990. [Google Scholar] [CrossRef]
- Russo, T.A.; Manohar, A.; Beanan, J.M.; Olson, R.; MacDonald, U.; Graham, J.; Umland, T.C. The Response Regulator BfmR Is a Potential Drug Target for Acinetobacter baumannii. mSphere 2016, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayakumar, K.; Thirunanasambandham, R. 5-Hydroxymethylfurfural inhibits Acinetobacter baumannii biofilms: An in vitro study. Arch. Microbiol. 2021, 203, 673–682. [Google Scholar] [CrossRef]
- Liu, W.; Wu, Z.; Mao, C.; Guo, G.; Zeng, Z.; Fei, Y.; Wan, S.; Peng, J.; Wu, J. Antimicrobial Peptide Cec4 Eradicates the Bacteria of Clinical Car-bapenem-Resistant Acinetobacter baumannii Biofilm. Front. Microbiol. 2020, 11, 1532. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.-S.; Eom, Y.-B. Antimicrobial and antibiofilm activities of Clostridium butyricum supernatant against Acinetobacter baumannii. Arch. Microbiol. 2020, 202, 1059–1068. [Google Scholar] [CrossRef]
- Stacy, D.M.; Welsh, M.; Rather, P.N.; Blackwell, H.E. Attenuation of Quorum Sensing in the PathogenAcinetobacter bau-manniiUsing Non-nativeN-Acyl Homoserine Lactones. ACS Chem. Biol. 2012, 7, 1719–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, P.; Alqahtani, A.S.; Husain, F.M.; Rehman, T.; Alajmi, M.F.; Noman, O.M.; El Gamal, A.A.; Al-Massarani, S.M.; Khan, M.S. Siphonocholin isolated from red sea sponge Siphonochalina siphonella attenuates quorum sensing controlled virulence and biofilm formation. Saudi Pharm. J. 2020, 28, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, N.; Singh, S.P.; Sharma, A.; Sharma, P.; Capalash, N. Attenuation of quorum sensing-mediated virulence of Aci-netobacter baumannii by Glycyrrhiza glabra flavonoids. Future Microbiol. 2015, 10, 1953–1968. [Google Scholar] [CrossRef]
- Alves, S.; Duarte, A.; Sousa, S.; Domingues, F.C. Study of the major essential oil compounds of Coriandrum sativum against Aci-netobacter baumannii and the effect of linalool on adhesion, biofilms and quorum sensing. Biofouling 2016, 32, 155–165. [Google Scholar] [CrossRef]
- Paul Bhattacharya, S.; Mitra, A.; Bhattacharya, A.; Sen, A. Quorum quenching activity of pentacyclic triterpenoids leads to inhi-bition of biofilm formation by Acinetobacter baumannii. Biofouling 2020, 36, 922–937. [Google Scholar] [CrossRef]
- Mayer, C.; Muras, A.; Martínez, A.P.; Romero, M.; Rumbo-Feal, S.; Poza, M.; Ramos-Vivas, J.; Otero, A. Quorum Sensing as a Target for Controlling Surface Associated Motility and Biofilm Formation in Acinetobacter baumannii ATCC® 17978TM. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Nicol, M.; Alexandre, S.; Luizet, J.-B.; Skogman, M.; Jouenne, T.; Salcedo, S.P.; Dé, E. Unsaturated Fatty Acids Affect Quorum Sensing Communication System and Inhibit Motility and Biofilm Formation of Acinetobacter baumannii. Int. J. Mol. Sci. 2018, 19, 214. [Google Scholar] [CrossRef] [Green Version]
- Muzammil, S.; Khurshid, M.; Nawaz, I.; Siddique, M.H.; Zubair, M.; Nisar, M.A.; Imran, M.; Hayat, S. Aluminium oxide nanoparticles inhibit EPS production, adhesion and biofilm formation by multidrug resistant Acinetobacter baumannii. Biofouling 2020, 36, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Panwar, S.; Kothidar, A.; Tiwari, V. Rational targeting of Wzb phosphatase and Wzc kinase interaction inhibits ex-tracellular polysaccharides synthesis and biofilm formation in Acinetobacter baumannii. Carbohydr. Res. 2020, 492, 108025. [Google Scholar] [CrossRef]
- Scutera, S.; Argenziano, M.; Sparti, R.; Bessone, F.; Bianco, G.; Bastiancich, C.; Castagnoli, C.; Stella, M.; Musso, T.; Cavalli, R. Enhanced Antimicrobial and Antibiofilm Effect of New Colistin-Loaded Human Albumin Nanoparticles. Antibiotics 2021, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Zeng, P.; Yi, L.; Xu, J.; Gao, W.; Xu, C.; Chen, S.; Chan, K.-F.; Wong, K.-Y. Investigation of antibiofilm activity, antibacterial activity, and mechanistic studies of an amphiphilic peptide against Acinetobacter baumannii. Biochim. Biophys. Acta (BBA) Biomembr. 2021, 1863, 183600. [Google Scholar] [CrossRef]
- Krishnan, M.; Choi, J.; Jang, A.; Kim, Y. A Novel Peptide Antibiotic, Pro10–1D, Designed from Insect Defensin Shows Anti-bacterial and Anti-Inflammatory Activities in Sepsis Models. Int. J. Mol. Sci. 2020, 21, 6216. [Google Scholar] [CrossRef]
- Raorane, C.J.; Lee, J.H.; Lee, J. Rapid Killing and Biofilm Inhibition of Multidrug-Resistant Acinetobacter baumannii Strains and Other Microbes by Iodoindoles. Biomolecules 2020, 10, 1186. [Google Scholar] [CrossRef]
- Pourhajibagher, M.; Hosseini, N.; Boluki, E.; Chiniforush, N.; Bahador, A. Photoelimination Potential of Chitosan Nanopar-ti-cles-Indocyanine Green Complex Against the Biological Activities of Acinetobacter baumannii Strains: A Preliminary In Vitro Study in Burn Wound Infections. J. Lasers Med. Sci. 2020, 11, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Pourhajibagher, M.; Partoazar, A.; Alaeddini, M.; Etemad-Moghadam, S.; Bahador, A. Photodisinfection effects of silver sul-fadiazine nanoliposomes doped-curcumin on Acinetobacter baumannii: A mouse model. Nanomedicine 2020, 15, 437–452. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Li, X.; Wang, L.; Li, G.; Cong, C.; Li, R.; Cui, H.; Murtaza, B.; Xu, Y. The endolysin of the Acinetobacter baumannii phage vB_AbaP_D2 shows broad antibacterial activity. Microb. Biotechnol. 2021, 14, 403–418. [Google Scholar] [CrossRef]
- Peng, Q.; Lin, F.; Ling, B. In vitro activity of biofilm inhibitors in combination with antibacterial drugs against extensively drug-resistant Acinetobacter baumannii. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Toscani, A.; Hind, C.; Clifford, M.; Kim, S.-H.; Gucic, A.; Woolley, C.; Saeed, N.; Rahman, K.M.; Sutton, J.M.; Castagnolo, D. Development of photoactivable phenanthroline-based manganese(I) CO-Releasing molecules (PhotoCORMs) active against ESKAPE bacteria and bacterial biofilms. Eur. J. Med. Chem. 2021, 213, 113172. [Google Scholar] [CrossRef]
- Zhang, J.; Liang, X.; Zhang, S.; Song, Z.; Wang, C.; Xu, Y. Maipomycin A, a Novel Natural Compound With Promising An-ti-biofilm Activity Against Gram-Negative Pathogenic Bacteria. Front. Microbiol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, J.; Yin, Y.; Wang, G.; Yang, M.; Li, Y.; Zhang, Z.; Lai, R. The antimicrobial peptide ZY4 combats multidrug-resistant Pseu-domonas aeruginosa and Acinetobacter baumannii infection. Proc. Natl. Acad. Sci. USA 2019, 116, 26516–26522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Carrizales, M.; Mendoza-Mendoza, E.; Peralta-Rodriguez, R.D.; Pérez-Díaz, M.A.; Portales-Pérez, D.; Magaña-Aquino, M.; Aragón-Piña, A.; Infante-Martínez, R.; Barriga-Castro, E.D.; Sánchez-Sánchez, R. Characterization, antibiofilm and biocompatibility properties of chitosan hydrogels loaded with silver nanoparticles and ampicillin: An alternative protection to central venous catheters. Colloids Surf. B Biointerfaces 2020, 196, 111292. [Google Scholar] [CrossRef] [PubMed]
- Krasowska, A.; Sigler, K. How microorganisms use hydrophobicity and what does this mean for human needs? Front. Cell. Infect. Microbiol. 2014, 4, 112. [Google Scholar] [CrossRef] [Green Version]
- Smani, Y.; McConnell, M.J.; Pachón, J. Role of Fibronectin in the Adhesion of Acinetobacter baumannii to Host Cells. PLoS ONE 2012, 7, e33073. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural Anti-biofilm Agents: Strategies to Control Bio-film-Forming Pathogens. Front. Microbiol. 2020, 11, 566325. [Google Scholar] [CrossRef]
- Reid, G.; Jass, J.; Sebulsky, M.T.; McCormick, J. Potential Uses of Probiotics in Clinical Practice. Clin. Microbiol. Rev. 2003, 16, 658–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila-Farrés, X.; Parra-Millán, R.; Sánchez-Encinales, V.; Varese, M.; Ayerbe-Algaba, R.; Bayo, N.; Guardiola, S.; Pachón-Ibáñez, M.E.; Kotev, M.; García, J.; et al. Combating virulence of Gram-negative bacilli by OmpA inhibition. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra-Millán, R.; Vila-Farrés, X.; Ayerbe-Algaba, R.; Varese, M.; Sánchez-Encinales, V.; Bayo, N.; Pachón-Ibáñez, M.E.; Teixidó, M.; Vila, J.; Pachón, J.; et al. Synergistic activity of an OmpA inhibitor and colistin against colistin-resistant Acinetobacter bau-mannii: Mechanistic analysis and in vivo efficacy. J. Antimicrob. Chemother. 2018, 73, 3405–3412. [Google Scholar] [CrossRef]
- Lin, M.F.; Tsai, P.W.; Chen, J.Y.; Lin, Y.Y.; Lan, C.Y. OmpA Binding Mediates the Effect of Antimicrobial Peptide LL-37 on Acineto-bacter baumannii. PLoS ONE 2015, 10, e0141107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, R.H.; Dijkshoorn, L.; Barselaar, M.V.D.; Nudel, C. Quorum sensing signal profile of Acinetobacter strains from nosocomial and environmental sources. Revista Argentina de Microbiología 2009, 41, 73–78. [Google Scholar] [PubMed]
- LaSarre, B.; Federle, M.J. Exploiting Quorum Sensing To Confuse Bacterial Pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saroj, S.D.; Rather, P.N. Streptomycin Inhibits Quorum Sensing in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2013, 57, 1926–1929. [Google Scholar] [CrossRef] [Green Version]
- Ismail, M.M.; Samir, R.; Saber, F.R.; Ahmed, S.R.; Farag, M.A. Pimenta Oil as A Potential Treatment for Acinetobacter Baumannii Wound Infection: In Vitro and In Vivo Bioassays in Relation to Its Chemical Composition. Antibiotics 2020, 9, 679. [Google Scholar] [CrossRef]
- Pourhajibagher, M.; Mahmoudi, H.; Rezaei-Soufi, L.; Alikhani, M.Y.; Bahador, A. Potentiation effects of antimicrobial pho-todynamic therapy on quorum sensing genes expression: A promising treatment for multi-species bacterial biofilms in burn wound infections. Photodiagn. Photodyn. Ther. 2020, 30, 101717. [Google Scholar] [CrossRef]
- Paluch, E.; Rewak-Soroczyńska, J.; Jędrusik, I.; Mazurkiewicz, E.; Jermakow, K. Prevention of biofilm formation by quorum quenching. Appl. Microbiol. Biotechnol. 2020, 104, 1871–1881. [Google Scholar] [CrossRef] [Green Version]
- Chow, J.Y.; Yang, Y.; Tay, S.B.; Chua, K.L.; Yew, W.S. Disruption of biofilm formation by the human pathogen Acinetobacter bau-mannii using engineered quorum-quenching lactonases. Antimicrob. Agents Chemother. 2014, 58, 1802–1805. [Google Scholar] [CrossRef] [Green Version]
- Bales, P.M.; Renke, E.M.; May, S.L.; Shen, Y.; Nelson, D.C. Purification and Characterization of Biofilm-Associated EPS Ex-opolysac-charides from ESKAPE Organisms and Other Pathogens. PLoS ONE 2013, 8, e67950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Cheng, C.; Xu, G.-B.; Tang, L.; Chua, K.-L.; Yang, Y.-Y. Synthesis and antibiofilm evaluation of 3-hydroxy-2,3-dihydroquinazolin-4(1H)-one derivatives against opportunistic pathogen Acinetobacter baumannii. Bioorgan. Med. Chem. 2020, 28, 115606. [Google Scholar] [CrossRef]
- Nakar, D.; Gutnick, D.L. Involvement of a Protein Tyrosine Kinase in Production of the Polymeric Bioemulsifier Emulsan from the Oil-Degrading Strain Acinetobacter lwoffii RAG-1. J. Bacteriol. 2003, 185, 1001–1009. [Google Scholar] [CrossRef] [Green Version]
- Reza, A.; Sutton, J.M.; Rahman, K.M. Effectiveness of Efflux Pump Inhibitors as Biofilm Disruptors and Resistance Breakers in Gram-Negative (ESKAPEE) Bacteria. Antibiotics 2019, 8, 229. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, C.; Barnett, P.; Perlmutter, J.; Dunman, P.M. Identification of Acinetobacter baumannii Serum-Associated Antibiotic Efflux Pump Inhibitors. Antimicrob. Agents Chemother. 2014, 58, 6360–6370. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.Q.; Estellés, A.; Li, L.; Abdelhady, W.; Gonzales, R.; Bayer, A.S.; Tenorio, E.; Leighton, A.; Ryser, S.; Kauvar, L.M. A Human Biofilm-Disrupting Monoclonal Antibody Potentiates Antibiotic Efficacy in Rodent Models of both Staphylococcus aureus and Acinetobacter baumannii Infections. Antimicrob. Agents Chemother. 2017, 61, e00904-17. [Google Scholar] [CrossRef] [Green Version]
- Eijkelkamp, B.A.; Hassan, K.A.; Paulsen, I.T.; Brown, M.H. Investigation of the human pathogen Acinetobacter baumannii under iron limiting conditions. BMC Genom. 2011, 12, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacioglu, M.; Oyardi, O.; Bozkurt-Guzel, C.; Savage, P.B. Antibiofilm activities of ceragenins and antimicrobial peptides against fungal-bacterial mono and multispecies biofilms. J. Antibiot. 2020, 73, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Fekrirad, Z.; Darabpour, E.; Kashef, N. Eradication of Acinetobacter baumannii Planktonic and Biofilm Cells Through Erythro-sine-Mediated Photodynamic Inactivation Augmented by Acetic Acid and Chitosan. Curr Microbiol. 2021, 78, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Anane, Y.A.; Apalata, T.; Vasaikar, S.; Okuthe, G.E.; Songca, S.P. In vitro antimicrobial photodynamic inactivation of multi-drug-resistant Acinetobacter baumannii biofilm using Protoporphyrin IX and Methylene blue. Photodiagnosis Photodyn. Ther. 2020, 30, 101752. [Google Scholar] [CrossRef]
- Vukotic, G.; Obradovic, M.; Novovic, K.; Di Luca, M.; Jovcic, B.; Fira, D.; Neve, H.; Kojic, M.; McAuliffe, O. Characterization, Antibiofilm, and Depolymerizing Activity of Two Phages Active on Carbapenem-Resistant Acinetobacter baumannii. Front. Med. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Grygorcewicz, B.; Wojciuk, B.; Roszak, M.; Łubowska, N.; Błażejczak, P.; Jursa-Kulesza, J.; Rakoczy, R.; Masiuk, H.; Dołęgowska, B. Environmental Phage-Based Cocktail and Antibiotic Combination Effects on Acinetobacter baumannii Biofilm in a Human Urine Model. Microb. Drug Resist. 2021, 27, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Neethu, S.; Midhun, S.J.; Radhakrishnan, E.K.; Jyothis, M. Surface functionalization of central venous catheter with myco-fabricated silver nanoparticles and its antibiofilm activity on multidrug resistant Acinetobacter baumannii. Microb. Pathog. 2020, 138, 103832. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.L.; Kawaguchi, B.; Taylor, N.B.; Allyn, G.; Badham, M.A.; Rogers, J.C.; Peterson, B.R.; Sebahar, P.R.; Haussener, T.J.; Reddy Kanna Reddy, H. In vivo efficacy of a unique first-in-class antibiofilm antibiotic for biofilm-related wound infections caused by Acinetobacter baumannii. Biofilm 2020, 2, 100032. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente-Núñez, C.; Cardoso, M.H.; de Souza Cândido, E.; Franco, O.L.; Hancock, R.E. Synthetic antibiofilm peptides. Biochim. Biophys. Acta 2016, 1858, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, K.D.; Pham, T.X.; Farnsworth, R.W.; Williams, D.L.; Loc-Carrillo, C.; Horne, L.A.; Ingebretsen, S.H.; Bloebaum, R.D. Development of a broad spectrum polymer-released antimicrobial coating for the prevention of resistant strain bacterial in-fections. J. Biomed. Mater. Res. Part A 2012, 100, 2732–2738. [Google Scholar] [CrossRef] [Green Version]
- Frei, R.; Breitbach, A.S.; Blackwell, H.E. 2-Aminobenzimidazole Derivatives Strongly Inhibit and Disperse Pseudomonas aeruginosa Biofilms. Angew. Chem. Int. Ed. 2012, 51, 5226–5229. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Chen, H.; Zhang, M.; Zhao, Y.; Jiang, Y.; Liu, X.; Huang, W.; Ma, Y. Clinical Experience of Personalized Phage Therapy Against Car-bapenem-Resistant Acinetobacter baumannii Lung Infection in a Patient With Chronic Obstructive Pulmonary Disease. Front. Cell Infect. Microbiol. 2021, 11, 631585. [Google Scholar] [CrossRef]
- Wu, N.; Dai, J.; Guo, M.; Li, J.; Zhou, X.; Li, F.; Gao, Y.; Qu, H.; Lu, H.; Jin, J.; et al. Pre-optimized phage therapy on secondary Acinetobacter baumannii infection in four critical COVID-19 patients. Emerg. Microbes Infect. 2021, 10, 612–618. [Google Scholar] [CrossRef]
- Blasco, L.; Ambroa, A.; Lopez, M.; Fernandez-Garcia, L.; Bleriot, I.; Trastoy, R.; Ramos-Vivas, J.; Coenye, T.; Fernandez-Cuenca, F.; Vila, J.; et al. Combined Use of the Ab105–2φΔCI Lytic Mutant Phage and Different Antibiotics in Clinical Isolates of Mul-ti-Resistant Acinetobacter baumannii. Microorganisms 2019, 7, 556. [Google Scholar] [CrossRef] [Green Version]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [Green Version]
- Briers, Y.; Walmagh, M.; Lavigne, R. Use of bacteriophage endolysin EL188 and outer membrane permeabilizers against Pseu-domonas aeruginosa. J. Appl. Microbiol. 2011, 110, 778–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furiga, A.; Lajoie, B.; El Hage, S.; Baziard, G.; Roques, C. Impairment of Pseudomonas aeruginosa Biofilm Resistance to Antibiotics by Combining the Drugs with a New Quorum-Sensing Inhibitor. Antimicrob. Agents Chemother. 2016, 60, 1676–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosler, S.; Karaaslan, E. Inhibition and destruction of Pseudomonas aeruginosa biofilms by antibiotics and antimicrobial peptides. Peptides 2014, 62, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Tak, Y.K.; Song, J.M. Does the antibacterial activity of silver nanoparticles depend on the shape of the nanoparticle? A study of the Gram-negative bacterium Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 1712–1720. [Google Scholar] [CrossRef] [Green Version]
- Prashanth, P.A.; Raveendra, R.S.; Hari Krishna, R.; Ananda, S.; Bhagya, N.P.; Nagabhushana, B.M.; Lingaraju, K.; Naika, H.R. Synthesis, characterizations, antibacterial and photoluminescence studies of solution combustion-derived α-Al2O3 nanopar-ticles. J. Asian Ceram. Soc. 2015, 3, 345–351. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Strategy | Strains/Isolates | Antibiofilm Activity | Antibiofilm Mechanisms | Reference |
|---|---|---|---|---|
| Pyrogallol (polyphenolic organic compound from galls and barks of various trees) | MTCC 9829 reference strain | MBIC: 20 μg/mL | Reduced CSH Reduced motility (swarming) Reduced EPS production Downregulation of adhesion-related genes (ompA, csuA/B) Downregulation of bap gene involved in biofilm formation and stabilization | [138] |
| Myrtenol (bicyclic monoterpene derived from various plants) | ATCC 19606 and MTCC 9826 reference strains Clinical isolates (n = 2) | MBIC: 200 μg/mL CLSM showed reduced biomass, maximum thickness, and surface-to-volume ratio | Reduced CSH Reduced motility (swarming, twitching) Downregulation of biofilm-associated genes (bfmR, csuA/B, bap, ompA, pgaA, pgaC) | [139] |
| Curcumin (flavonoid) | A. baumannii ATCC 17,978 reference strainClinical isolates (n = 8) | Biofilm inhibition by: 46% at 20 μg/mL 93% at 100 μ/mL | Reduced motility (swimming, swarming) Interaction with the biofilm response regulator BfmR | [140,141] |
| 5-hydroxymethylfurfural (furan organic compound derived from the dehydration of reducing sugars) | ATCC 19606 reference strain | MBIC: 100 μg/mL | Reduced CSH Reduced motility (swarming, twitching) Reduced EPS production Downregulation of biofilm-related genes (bap, csuA/B, ompA, bfmR, katE) | [142] |
| Synthetic peptide Cec4 | Carbapenem-resistant isolates (n = 200) | MBIC: 64–128 µg/mL | Reduced motility (twitching) Downregulation of biofilm-related genes (csuE, bfmR and bfmS, bap) | [143] |
| CFS from Clostridium butyricum | ATCC 19606 reference strain MDR clinical isolates (n = 2) | Biofilm inhibition by: 24.4–33.9%, at 12.5% CFS 28.2–43.1%, at 25% CFS 93.6–99.6%, at 50% CFS | Reduced motility Downregulation of RND-type efflux pump-related adeABC genes | [144] |
| Non-native AbaR antagonists | M2 abaI::lacZ (ΔabaI reporter) and M2 wild-type | Biofilm inhibition by 40% | QS inhibition | [145] |
| Siphonocholin (from marine sponge Siphonochalina siphonella) | ATCC BAA747 reference strain | Biofilm inhibition by 70% | QS inhibition Reduced motility (swarming) Reduced EPS production | [146] |
| Flavonoid-rich active fraction F1 (from Glycyrrhiza glabra) | ATCC 19,606 and ATCC 17,978 reference strains Clinical isolates (n = 5) | Concentration-dependent effect Maximum biofilm inhibition by 30–70%, at 2 mg/mL | QS inhibition by abaI downregulation Reduced motility (twitching) | [147] |
| Linalool (oil compounds from Coriandrum sativum) | LMG 1025 and LMG 1041 reference strains Clinical isolates (n = 3) | Concentration-dependent effect: 1–18%, at 0.25 × MIC 75–97.1%, at 4 × MIC | Reduced adhesion QS inhibition | [148] |
| Pentacyclic triterpenoids (betulinic acid, glycyrrhetinic acid, ursolic acid) | ATCC 19606 reference strain | Biofilm inhibition (respectively at 50, 100 and 200 µg/mL): 36, 56, 80% (glycyrrhetinic acid) 31, 63, 88% (ursolic acid) 45, 62, 88% (betulinic acid) | QS inhibition (at AHL synthase and AHL dependent transcriptional activator) Reduced EPS production | [149] |
| MomL (AHL lactonase belonging to the metallo-β-lactamase superfamily) | LMG10520, LMG10531 and AB5075 reference strains | Concentration-dependent effect Maximum biofilm inhibition by 42% at 5 µg/mL | AHL degrading activity | [119] |
| Purified QQ enzyme Aii20J | ATCC17978 reference strain MDR clinical strains (n = 5) | Biofilm inhibition by 80% The effect is strain-dependent and improved when QQ enzyme is combined with DNase | Decreased the number of surface short pili | [150] |
| Palmitoleic acid, myristoleic acid (unsaturated fatty acids) | ATCC17978 reference strain Clinical isolates (n = 22) | Biofilm inhibition (at 0.02 and 0.05 mg/mL, respectively) by: 37 and 39% (palmitoleic acid) 28 and 42% (mirystoleic acid) Significant biofilm reduction in: 13 isolates (palmitoleic acid) 8 isolates (mirystoleic acid) | Inhibition of abaR gene expressionAccumulation of fatty acids at the air−liquid interface, due to their amphiphilic nature | [151] |
| Al2O3 synthetic NPs | MDR strains (n = 3) | Biofilm inhibition by 11.6 to 70.2% at 0.5xMIC | Reduced EPS production | [152] |
| Labetalol hydrochloride (Wzb-Wzc interaction inhibitor) | RS 307 reference strain | MBIC: 1 mM | Reduced EPS production | [153] |
| Chitosan-coated human albumin nanoparticles for the delivery of colistin (Col/haNPs) | ATCC 19,606 reference strain Colistin-susceptible (n = 1) and -resistant (n = 3) clinical isolates | Significant biofilm inhibition at 1/2x and 1/4xMIC Col/haNPs > 4–60-fold vs. free colistin | Positively charged NPs might adsorb and accumulate on the negatively charged bacterial surface and EPS by electrostatic interactions Prolonged release of colistin Chitosan−colistin synergistic effect | [154] |
| Polyclonal antibodies vs. self-complemented CsuA/B subunit (αA/B) and CsuENTD (αEN) | Clinical strains (n = 5) | αEN inhibits biofilm formation more efficiently than αA/B: αEN diluted up to several thousand times completely blocked biofilm formation αA/B inhibited biofilm formation only at high concentration | Inhibition of the binding to hydrophobic plastics by blocking the three hydrophobic fingers at the tip of CsuE N-terminal domain (CsuENTD) | [43] |
| Cationic amphiphilic peptide zp3 (GIIAGIIIKIKK-NH2) | ATCC 19606 reference strain | Biofilm inhibition by: 20%, at 0.5 μM 100%, at >4 μM | Destabilization of cell membranes with pore formation and consequent biofilm collapse | [155] |
| Pro10-1D (a short peptide from insect defensin) | KCCM 40203, CCARM 12010 and CCARM 12220 reference strains | Concentration-dependent effect: 20% inhibition at 2 µM >99.9% inhibition at 64 µM | Reduced EPS production | [156] |
| 24 indole derivatives (including 16 halogenated indoles) | ATCC 17978 and ATCC BAA-1709 reference strains MDR clinical isolates (n = 7) | Biofilm inhibition at 50 µg/mL: 62% (4-bromoindole) 75% (4-chloroindole) 60% (4-iodoindole) 94% (6-iodoindole) 96% (5-iodoindole) | Reduced surface motility Induced reactive oxygen species, resulting in loss of cell membrane integrity and cell shrinkage | [157] |
| PDI mediated by indocyanine green encapsulated in chitosan nanoparticles (NCs@ICG-aPDT) | Isolates from burn wounds (n = 50) | Biofilm inhibition by: 55.3% after exposure to NCs@ICG-aPDT No inhibition after exposure to NCs@ICG, ICG, and the diode laser alone | Bactericidal effect | [158] |
| PDI of nanoliposomal silver sulfadiazine doped with curcumin (AgSD-NLs@Cur) | Isolates from burn wounds (n = 100) | Biofilm inhibition by 76.4% after exposure to AgSD-NLs@Cur at MIC90 and light-emitting diode Photoexcited AgSD and AgSD-NLs at MIC90 are more effective than either group without LED irradiation (38.1 vs. 44.8%, respectively) | Ag in AgSD-NLs interacts with DNA and sulfhydryl groups of microbial enzymes, leading to bacterial growth inhibition Downregulation of luxI gene | [159] |
| Endolysin Abtn-4 from phage vB_AbaP_D2 (isolated from hospital wastewater) | Clinical MDR isolates (n = 15) AB9 host strain | Biofilm inhibition > 30% following exposure in the early (12 h post-incubation) or pre-maturation phase (36 h post-incubation) | EPS disruption Bacterial cell wall degradation | [160] |
| Pentacyclic triterpenoids, (glycyrrhetinic acid, ursolic acid, betulinic acid) combined with a conventional antibiotic (doxycycline, roxithromycin or ciprofloxacin) | ATCC 19606 reference strain | Glycyrrhetinic acid and betulinic acid increase antibiofilm activity of doxycycline and roxithromycin Ursolic acid improves the effect of ciprofloxacin | Increased antibiotic diffusion through biofilm mediated by the triterpenoids | [149] |
| Biofilm inhibitors (zinc lactate, stannous fluoride, furanone, AZM, and RIF) combined with a conventional antibiotic (IMP, MRP, TIG, POL) | XDR clinical isolates (n = 9) | Biofilm inhibition: 16 to 50%, at sub-MICs lactate > stannous fluoride > furanone > RIF > AZM Synergistic effects of: zinc lactate, stannous fluoride and furanone combined with TIG (22, 56 and 11% of the isolates, respectively) zinc lactate and stannous fluoride each used with a carbapenem (IMP or MRP), in 33% of the isolates | Zinc compounds inhibit EPS synthesis and the formation of matrix networks Stannous fluoride destroys the biofilm structure by loosening the structure of the biofilm matrix Furanone replaces the binding sites of QS signal molecules Azithromycin inhibits EPS production, leading to the formation of channels that favor antibiotic diffusion through the biofilm | [161] |
| Phenanthroline-based visible-light-activated manganese (I) carbon-monoxide-releasing molecules (PhotoCORMs) | ATCC BAA 1710 and ATCC 17978 reference strains | Biofilm formation inhibition only at high concentrations (>128 mg/mL) in the dark Compounds 1-2 reveal remarkable activity at 4–8 mg/mL when irradiated with blue LED light Compound 2 shows higher activity than ciprofloxacin vs. MDR ATCC BAA 1710 strain | Antibacterial activity due to the combination of CO release as well as the production of photo-byproducts | [162] |
| Maipomycin A (from the metabolites of the marine actinomycete Kibdelosporangium phytohabitans XY-R10) | ATCC 19606 reference strain | Biofilm inhibition by 84.3% at MBIC (8 μg/mL) Concentration-dependent effect Inhibition of biofilm formed on medical materials, such as catheters (silicone) and endotracheal tubes (polyvinyl chloride) | Fe(II) and Fe(III) ions chelation The chelation of Maipomycin A and iron ions may be negatively affected by other metal as competitors | [163] |
| ZY4 cyclic synthetic peptide (designed on cathelicidin-BF15 and stabilized by a disulfide bridge) | ATCC 22933 reference strain MDR clinical isolates (n = 5) | Concentration-dependent effect 22%, at 0.5xMIC 46%, at 2xMIC 66%, at 8xMIC | Bactericidal effect by permeabilizing the cell membrane | [164] |
| Chitosan hydrogels loaded with AgNPs and AMP | Carbapenem-resistant isolate from CVC | Biofilm viability inhibition on CVC: 1 log10 (chitosan) 10 log10 (chitosan with 25 ppm AgNPs and 50 ppm AMP) | NS | [165] |
| Strategy | Strains/Isolates | Antibiofilm Activity | Antibiofilm Mechanisms | Reference |
|---|---|---|---|---|
| Myrtenol (bicyclic monoterpene derived from various plants) | ATCC-19606 and MTCC-9826 reference strains Clinical isolates (n = 2) | Significant biofilm dispersion at MBIC for all strains tested | Reduced CSH Reduced motility (swarming, twitching) Downregulation of biofilm-associated genes (bfmR, csuA/B, bap, ompA, pgaA, pgaC) | [139] |
| Synthetic peptide Cec4 | Carbapenem-resistant isolates (n = 200) | Biofilm dispersion > 20% at 1xMIC (4 µg/mL) MBEC50: 16 µg/mL MBEC80: 128 µg/mL MBEC: 256–512 µg/mL | Reduced motility (twitching) Downregulation of biofilm-related genes (csuE, bfmR and bfmS, bap) | [143] |
| CFS from Clostridium butyricum | ATCC 19,606 reference strain MDR clinical isolates (n = 2) | Biofilm dispersion by: 24–63.2%, at 12.5% CFS 28.4–84%, at 25% CFS 80.5–92.6%, at 50% CFS Decreased metabolic activity by 92.9–100% at 50% CFS | Reduced motility Downregulation of RND-type efflux pump-related adeABC genes | [144] |
| Linalool (oil compounds from Coriandrum sativum) | LMG 1025 and LMG 1041 reference strains Clinical isolates (n = 3) | Concentration-dependent biofilm dispersion: 1–28%, at 0.25xMIC 55–86%, at 4xMIC | Reduced adhesion QS inhibition | [148] |
| Pentacyclic triterpenoids (betulinic acid, glycyrrhetinic acid, ursolic acid) | ATCC 19606 reference strain | Reduction of biofilm viability at 200 µg/mL by: 26% (glycyrrhetinic acid) 23% (ursolic acid) 23% (betulinic acid) | QS inhibition (at AHL synthase and AHL dependent transcriptional activator) Reduced EPS production | [149] |
| PDI mediated by indocyanine green | ATCC 19606 reference strain | Biofilm viability reduction by 3 log10 at 1000 µg/mL (laser irradiation for 1 min, with an estimated average output light energy 31.2 J/cm2) | QS inhibition by abaI downregulation | [177] |
| Thermostable QQ phosphotriesterase-like lactonase (from Geobacillus kaustophilus) | Clinical isolate | Biofilm dispersion by 75% CLSM confirms reduction in biofilm biomass, thickness, and surface area | Hydrolysis of two biologically relevant C-3-hydroxylated AHLs (3-OH-C10-HSL, 3-OH-C12-HSL) | [179] |
| Palmitoleic acid, myristoleic acid (unsaturated fatty acids) | ATCC 17978 reference strain Clinical isolates (n = 22) | Biofilm dispersion by 24% for both fatty acids at 0.05 mg/mL AFM shows merging microcolonies with cell elongation | Inhibition of abaR gene expression Accumulation of fatty acids at the air−liquid interface, due to their amphiphilic nature | [151] |
| Al2O3 synthetic NPs | MDR strains (n = 3) | Biofilm dispersion by 11.4 to 56.8% at 0.5xMIC Efficacy reduced over time, due to the loss of affinity of the NPs with treated cells: 11.4 to 20.9% reduction of 120 h-old biofilm 33.8 to 56.8% reduction of 48 h-old biofilm | Reduced EPS production | [152] |
| TRL1068 (native human monoclonal antibody) | MDR clinical isolate | Biofilm disruption at 1.2 µg/mL for 12 h Murine skin and soft tissue infection model: imipenem + TRL1068 > imipenem + isotype control (reduction by 0.8 and 0.5 log10 CFU/catheter, respectively) | Binding to the biofilm component DNABII protein family | [185] |
| Cationic amphiphilic peptide zp3 (GIIAGIIIKIKK-NH2) | ATCC 19606 reference strain | Concentration-dependent effect: 30% at 2 μM (0.5xMIC) >50% at 64 μM (16xMIC) | Destabilization of cell membranes with pore formation and consequent biofilm collapse | [155] |
| Ceragenins (CSA-13, CSA-44, CSA-90, CSA-131, CSA-138, CSA-142, CSA-144, and CSA-192) synthesized from a cholic acid scaffold technique | ATCC 19606 reference strain | CSA-13 is the most effective, causing 1 log10 reduction of biofilm viability at 100 µg/mL All but CSA-144 are effective vs. A. baumannii-C. albicans mixed biofilms | NS | [187] |
| Indole derivatives (including halogenated indoles) | ATCC 17,978 and ATCC BAA-1709 reference strains MDR clinical isolates (n = 7) | 5-iodoindole at 250 µg/mL causes: 62% biofilm dispersion 95% biofilm killing CLSM shows that 5-iodoindole significantly reduces biofilm biomass, thickness, and substrate coverage | Reduced surface motility Induced reactive oxygen species, resulting in loss of cell membrane integrity and cell shrinkage | [157] |
| PDI mediated by erythrosine B | ATCC BAA 747 reference strain Isolates from burn wounds (n = 2) | PDI mediated by erythrosine B + acetic acid does not cause lethal effect (≥3 log10 reduction in CFU) on biofilm A lethal effect is observed after adding chitosan at 1/2xMIC to erythrosine B | Chitosan improves the antibiofilm efficacy of erythrosine B-mediated PDI by: disruption of biofilm structure, thus helping the PDI to act on the cells released from the biofilm permeabilization of the bacterial outer membrane acting as a drug carrier for delivery of erythrosine to biofilm | [188] |
| PDI mediated by methylene blue and protoporphyrin IX | MDR isolates from clinical (n = 1), abattoir (n = 1) and aquatic (n = 1) sources | Dose-dependent biofilm viability reduction by: 6 log10 for protoporphyrin IX 7 log10 for methylene blue | Cationic methylene blue is attracted to the negative EPS and cell walls, thus increasing its intracellular concentration following antimicrobial PDI; this causes biofilm detachment secondary to disintegration of the interaction between bacteria, due to reactive oxygen species exposure The negatively charged protoporphyrin IX is electrostatically repulsed from EPS and also from the lipopolysaccharide layer of the bacterial cell wall | [189] |
| ISTD depolymerase producing phage, isolated from wastewaters | Clinical isolates (89 carbapenem-resistant, 14 carbapenem-sensitive) 6077/12 host strain | Biofilm viability reduction after 6 h exposure to ISTD phages: 0.6 log10, at MOI 0.1 2 log10, at MOI 100 No reduction after 24 h exposure | NS | [190] |
| Environmental phages, tested both alone or combined with a conventional antibiotic (CIP, SXT, GN, TOB, IMP, MRP) | MDR isolates (n = 25) from urinary tract infections | Biofilm biomass reduction in a human urine model: by 35 to 67.4% after single phage exposure dose-dependent effect of phage cocktail + antibiotic (CIP, SXT, GN, TOB, IMP, or MRP) combination SXT-phage cocktail the most active for biofilm dispersion (94.3 and 98.6% at 1/4× and ½ × MIC, respectively), and bacterial regrowth limitation (to 18% and 6% for 1/4 and ½ × MIC values, respectively) | NS | [191] |
| Surface functionalization of central venous catheter with mycofabricated silver nanoparticles (AgNPs) | MDR isolate | MBEC: 31.2 μg/mL SEM and FE-SEM analyses confirm eradication of preformed biofilm on the AgNPs-coated surface catheter | The direct interaction of AgNPs to the bacterial cell wall and the subsequent penetration resulted in infiltration of AgNPs through the biofilm structure causing physical dispersion of biofilms and bacterial lysis | [192] |
| ZY4 cyclic synthetic peptide (designed on cathelicidin-BF15 and stabilized by a disulfide bridge) | ATCC 22933 reference strain MDR clinical isolates (n = 5) | Concentration-dependent effect More than half of the persisters eliminated at 1 × MIC | Bactericidal effect by permeabilizing the cell membrane | [164] |
| Hydrogel formulation of first-in-class series of antibiofilm antibiotic (CZ-01179) | ATCC BAA-1605 reference strain CDC clinical isolates (n = 11) | CZ-01179 is more effective than clinical standard (silver sulfadiazine) at eradicating biofilms both in vitro and up to 6 days faster at eradicating biofilms in a pig wound model | NS | [193] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pompilio, A.; Scribano, D.; Sarshar, M.; Di Bonaventura, G.; Palamara, A.T.; Ambrosi, C. Gram-Negative Bacteria Holding Together in a Biofilm: The Acinetobacter baumannii Way. Microorganisms 2021, 9, 1353. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071353
Pompilio A, Scribano D, Sarshar M, Di Bonaventura G, Palamara AT, Ambrosi C. Gram-Negative Bacteria Holding Together in a Biofilm: The Acinetobacter baumannii Way. Microorganisms. 2021; 9(7):1353. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071353
Chicago/Turabian StylePompilio, Arianna, Daniela Scribano, Meysam Sarshar, Giovanni Di Bonaventura, Anna Teresa Palamara, and Cecilia Ambrosi. 2021. "Gram-Negative Bacteria Holding Together in a Biofilm: The Acinetobacter baumannii Way" Microorganisms 9, no. 7: 1353. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071353