
Virome in the Lungs: The Role of Anelloviruses in Childhood Respiratory Diseases



Abstract
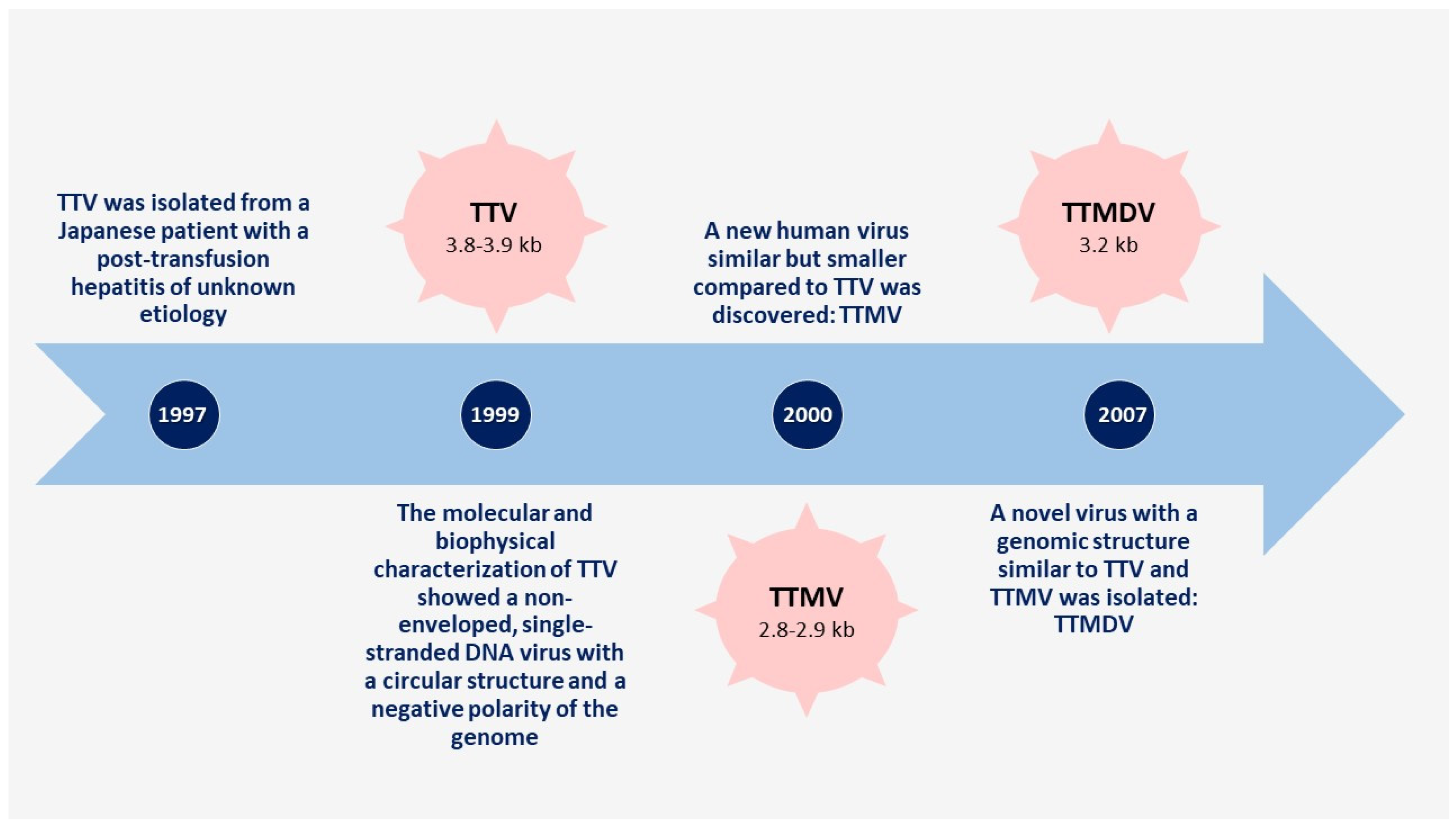
:1. Introduction
2. Search Strategy and Inclusion Criteria
3. Anelloviruses
4. Anelloviruses and Immunity
5. Anelloviruses and Chronic Respiratory Diseases
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Virgin, H.W. The Virome in Mammalian Physiology and Disease. Cell 2014, 157, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.B.; Glanville, A.R. Introduction to Techniques and Methodologies for Characterizing the Human Respiratory Virome. Methods Mol. Biol. 2018, 1838, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Wylie, K.M. The Virome of the Human Respiratory Tract. Clin. Chest Med. 2017, 38, 11–19. [Google Scholar] [CrossRef]
- Singanayagam, A.; Joshi, P.V.; Mallia, P.; Johnston, S.L. Viruses exacerbating chronic pulmonary disease: The role of immune modulation. BMC Med. 2012, 10, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willner, D.; Furlan, M.; Haynes, M.; Schmieder, R.; Angly, F.E.; Silva, J.; Tammadoni, S.; Nosrat, B.; Conrad, D.; Rohwer, F. Metagenomic Analysis of Respiratory Tract DNA Viral Communities in Cystic Fibrosis and Non-Cystic Fibrosis Individuals. PLoS ONE 2009, 4, e7370. [Google Scholar] [CrossRef] [PubMed]
- Vannella, K.M.; Moore, B.B. Viruses as co-factors for the initiation or exacerbation of lung fibrosis. Fibrogenesis Tissue Repair 2008, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Willner, D.; Furlan, M. Deciphering the role of phage in the cystic fibrosis airway. Virulence 2010, 1, 309–313. [Google Scholar] [CrossRef] [Green Version]
- Fancello, L.; Desnues, C.; Raoult, D.; Rolain, J.M. Bacteriophages and diffusion of genes encoding antimicrobial resistance in cystic fibrosis sputum microbiota. J. Antimicrob. Chemother. 2011, 66, 2448–2454. [Google Scholar] [CrossRef]
- Rolain, J.M.; Fancello, L.; Desnues, C.; Raoult, D. Bacteriophages as vehicles of the resistome in cystic fibrosis. J. Antimicrob. Chemother. 2011, 66, 2444–2447. [Google Scholar] [CrossRef] [Green Version]
- A Duerkop, B.; Hooper, L.V. Resident viruses and their interactions with the immune system. Nat. Immunol. 2013, 14, 654–659. [Google Scholar] [CrossRef]
- E James, C.; Davies, E.V.; Fothergill, J.; Walshaw, M.J.; Beale, C.M.; A Brockhurst, M.; Winstanley, C. Lytic activity by temperate phages of Pseudomonas aeruginosa in long-term cystic fibrosis chronic lung infections. ISME J. 2015, 9, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- Davies, E.V.; E James, C.; Kukavica-Ibrulj, I.; Levesque, R.C.; Brockhurst, M.; Winstanley, C. Temperate phages enhance pathogen fitness in chronic lung infection. ISME J. 2016, 10, 2553–2555. [Google Scholar] [CrossRef] [PubMed]
- Virgin, H.W.; Wherry, E.J.; Ahmed, R. Redefining Chronic Viral Infection. Cell 2009, 138, 30–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klumpp, J.; Fouts, D.E.; Sozhamannan, S. Next generation sequencing technologies and the changing landscape of phage genomics. Bacteriophage 2012, 2, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Wommack, K.E.; Bhavsar, J.; Polson, S.W.; Chen, J.; Dumas, M.; Srinivasiah, S.; Furman, M.; Jamindar, S.; Nasko, D.J. VIROME: A standard operating procedure for analysis of viral metagenome sequences. Stand. Genom. Sci. 2012, 6, 427–439. [Google Scholar] [CrossRef] [Green Version]
- Grinde, B. Viruses belonging to Anelloviridae or Circoviridae as a possible cause of chronic fatigue. J. Transl. Med. 2020, 18, 485. [Google Scholar] [CrossRef]
- Nishizawa, T.; Okamoto, H.; Konishi, K.; Yoshizawa, H.; Miyakawa, Y.; Mayumi, M. A novel DNA virus (TTV) associated with elevated transaminase levels in posttransfusion hepatitis of unknown etiology. Biochem. Biophys. Res. Commun. 1997, 241, 92–97. [Google Scholar] [CrossRef]
- Mushahwar, I.K.; Erker, J.C.; Muerhoff, A.S.; Leary, T.P.; Simons, J.N.; Birkenmeyer, L.G.; Chalmers, M.L.; Pilot-Matias, T.J.; Dexai, S.M. Molecular and biophysical characterization of TT virus: Evidence for a new virus family infecting humans. Proc. Natl. Acad. Sci. USA 1999, 96, 3177–3182. [Google Scholar] [CrossRef] [Green Version]
- Pisani, G.; Antigoni, I.; Bisso, G.; Wirz, M.; Iudicone, P.; Miceli, M.; Gentili, G. Prevalence of TT viral DNA in italian blood donors with and without elevated serum ALT levels: Molecular characterization of viral DNA isolates. Haematologica 2000, 85, 181–185. [Google Scholar]
- Takahashi, K.; Iwasa, Y.; Hijikata, M.; Mishiro, S. Identification of a new human DNA virus (TTV-like mini virus, TLMV) intermediately related to TT virus and chicken anemia virus. Arch. Virol. 2000, 145, 979–993. [Google Scholar] [CrossRef]
- Naoumov, N.V.; Petrova, E.P.; Thomas, M.G.; Williams, R. Presence of a newly described human DNA virus (TTV) in patients with liver disease. Lancet 1998, 352, 195–197. [Google Scholar] [CrossRef]
- Ninomiya, M.; Nishizawa, T.; Takahashi, M.; Lorenzo, F.R.; Shimosegawa, T.; Okamoto, H. Identification and genomic characterization of a novel human torque teno virus of 3.2 kb. J. Gen. Virol. 2007, 88, 1939–1944. [Google Scholar] [CrossRef] [PubMed]
- Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. (Eds.) Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- TeKippe, E.M.; Wylie, K.M.; Deych, E.; Sodergren, E.; Weinstock, G.; Storch, G.A. Increased Prevalence of Anellovirus in Pediatric Patients with Fever. PLoS ONE 2012, 7, e50937. [Google Scholar] [CrossRef]
- Blatter, J.A.; Sweet, S.C.; Conrad, C.; Danziger-Isakov, L.A.; Faro, A.; Goldfarb, S.B.; Hayes, D.; Melicoff, E.; Schecter, M.; Storch, G.; et al. Anellovirus loads are associated with outcomes in pediatric lung transplantation. Pediatr. Transplant. 2018, 22, e13069. [Google Scholar] [CrossRef]
- Maggi, F.; Pifferi, M.; Fornai, C.; Andreoli, E.; Tempestini, E.; Vatteroni, M.; Presciuttini, S.; Marchi, S.; Pietrobelli, A.; Boner, A.; et al. TT Virus in the Nasal Secretions of Children with Acute Respiratory Diseases: Relations to Viremia and Disease Severity. J. Virol. 2003, 77, 2418–2425. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, H. History of Discoveries and Pathogenicity of TT Viruses. TT Viruses 2009, 331, 1–20. [Google Scholar] [CrossRef]
- Ninomiya, M.; Takahashi, M.; Nishizawa, T.; Shimosegawa, T.; Okamoto, H. Development of PCR Assays with Nested Primers Specific for Differential Detection of Three Human Anelloviruses and Early Acquisition of Dual or Triple Infection during Infancy. J. Clin. Microbiol. 2008, 46, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spandole, S.; Cimponeriu, D.; Berca, L.M.; Mihăescu, G. Human anelloviruses: An update of molecular, epidemiological and clinical aspects. Arch. Virol. 2015, 160, 893–908. [Google Scholar] [CrossRef] [PubMed]
- Moen, E.M.; Sagedal, S.; Bjøro, K.; Degré, M.; Opstad, P.K.; Grinde, B. Effect of immune modulation on TT virus (TTV) and TTV-like-mini-virus (TLMV) viremia. J. Med. Virol. 2003, 70, 177–182. [Google Scholar] [CrossRef]
- Shibayama, T.; Masuda, G.; Ajisawa, A.; Takahashi, M.; Nishizawa, T.; Tsuda, F.; Okamoto, H. Inverse relationship between the titre of TT virus DNA and the CD4 cell count in patients infected with HIV. AIDS 2001, 15, 563–570. [Google Scholar] [CrossRef] [Green Version]
- Young, J.C.; Chehoud, C.; Bittinger, K.; Bailey, A.; Diamond, J.M.; Cantu, E.; Haas, A.R.; Abbas, A.; Frye, L.; Christie, J.; et al. Viral Metagenomics Reveal Blooms of Anelloviruses in the Respiratory Tract of Lung Transplant Recipients. Am. J. Transplant. 2015, 15, 200–209. [Google Scholar] [CrossRef] [Green Version]
- De Vlaminck, I.; Khush, K.K.; Strehl, C.; Kohli, B.; Luikart, H.; Neff, N.F.; Okamoto, J.; Snyder, T.M.; Cornfield, D.N.; Nicolls, M.R.; et al. Temporal Response of the Human Virome to Immunosuppression and Antiviral Therapy. Cell 2013, 155, 1178–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Görzer, I.; Jaksch, P.; Kundi, M.; Seitz, T.; Klepetko, W.; Puchhammer-Stöckl, E. Pre-Transplant Plasma Torque Teno Virus Load and Increase Dynamics after Lung Transplantation. PLoS ONE 2015, 10, e0122975. [Google Scholar] [CrossRef] [Green Version]
- Wylie, K.M.; Mihindukulasuriya, K.A.; Sodergren, E.; Weinstock, G.M.; Storch, G.A. Sequence Analysis of the Human Virome in Febrile and Afebrile Children. PLoS ONE 2012, 7, e27735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggi, F.; Pifferi, M.; Tempestini, E.; Lanini, L.; De Marco, E.; Fornai, C.; Andreoli, E.; Presciuttini, S.; Vatteroni, M.L.; Pistello, M.; et al. Correlation between Torque Tenovirus Infection and Serum Levels of Eosinophil Cationic Protein in Children Hospitalized for Acute Respiratory Diseases. J. Infect. Dis. 2004, 190, 971–974. [Google Scholar] [CrossRef] [Green Version]
- Thom, K.; Petrik, J. Progression towards AIDS leads to increased torque teno virus and torque teno minivirus titers in tissues of HIV infected individuals. J. Med. Virol. 2007, 79, 1–7. [Google Scholar] [CrossRef]
- Nasser, T.F.; de Oliveira, K.B.; Reiche, E.M.V.; Amarante, M.K.; Fungaro, M.H.P.; Watanabe, M.A.E. Detection of TT virus in HIV-1 exposed but uninfected individuals and in HIV-1 infected patients and its influence on CD4+ lymphocytes and viral load. Microb. Pathog. 2009, 47, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Moen, E.M.; Sleboda, J.; Grinde, B. Serum concentrations of TT virus and TT virus-like mini virus in patients developing AIDS. AIDS 2002, 16, 1679–1682. [Google Scholar] [CrossRef]
- Lemanske, R.F., Jr. Viral infections and asthma inception. J. Allergy Clin. Immunol. 2004, 114, 1023–1026. [Google Scholar] [CrossRef]
- Lemanske, R.F., Jr.; Jackson, D.J.; Gangnon, R.E.; Evans, M.D.; Li, Z.; Shult, P.A.; Kirk, C.J.; Reisdorf, E.; Roberg, K.A.; Anderson, E.L.; et al. Rhinovirus illnesses during infancy predict subsequent childhood wheezing. J. Allergy Clin. Immunol. 2005, 116, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Gern, J.E.; Rosenthal, L.A.; Sorkness, R.L.; Lemanske, R.F. Effects of viral respiratory infections on lung development and childhood asthma. J. Allergy Clin. Immunol. 2005, 115, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Vareille, M.; Kieninger, E.; Edwards, M.R.; Regamey, N. The Airway Epithelium: Soldier in the Fight against Respiratory Viruses. Clin. Microbiol. Rev. 2011, 24, 210–229. [Google Scholar] [CrossRef] [Green Version]
- Lejeune, S.; Deschildre, A.; Le Rouzic, O.; Engelmann, I.; Dessein, R.; Pichavant, M.; Gosset, P.; Stéphanie, L.; Antoine, D.; Olivier, L.R.; et al. Childhood asthma heterogeneity at the era of precision medicine: Modulating the immune response or the microbiota for the management of asthma attack. Biochem. Pharmacol. 2020, 179, 114046. [Google Scholar] [CrossRef]
- Durrani, S.R.; Montville, D.J.; Pratt, A.S.; Sahu, S.; DeVries, M.K.; Rajamanickam, V.; Gangnon, R.E.; Gill, M.A.; Gern, J.E.; Lemanske, R.F.; et al. Innate immune responses to rhinovirus are reduced by the high-affinity IgE receptor in allergic asthmatic children. J. Allergy Clin. Immunol. 2012, 130, 489–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holt, P.G.; Sly, P. Viral infections and atopy in asthma pathogenesis: New rationales for asthma prevention and treatment. Nat. Med. 2012, 18, 726–735. [Google Scholar] [CrossRef]
- De Heer, H.J.; Hammad, H.; Soullié, T.; Hijdra, D.; Vos, N.; Willart, M.A.; Hoogsteden, H.C.; Lambrecht, B.N. Essential Role of Lung Plasmacytoid Dendritic Cells in Preventing Asthmatic Reactions to Harmless Inhaled Antigen. J. Exp. Med. 2004, 200, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Troy, N.M.; Bosco, A. Respiratory viral infections and host responses; insights from genomics. Respir. Res. 2016, 17, 156. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, E.; Inoue, Y. MicroRNAs in Allergic Disease. Curr. Allergy Asthma Rep. 2016, 16, 67. [Google Scholar] [CrossRef]
- Rocchi, J.; Ricci, V.; Albani, M.; Lanini, L.; Andreoli, E.; Macera, L.; Pistello, M.; Ceccherini-Nelli, L.; Bendinelli, M.; Maggi, F. Torquetenovirus DNA drives proinflammatory cytokines production and secretion by immune cells via toll-like receptor 9. Virology 2009, 394, 235–242. [Google Scholar] [CrossRef]
- Freer, G.; Maggi, F.; Pistello, M. Virome and Inflammasomes, a Finely Tuned Balance with Important Consequences for the Host Health. Curr. Med. Chem. 2019, 26, 1027–1044. [Google Scholar] [CrossRef] [PubMed]
- Kincaid, R.P.; Burke, J.M.; Cox, J.C.; De Villiers, E.-M.; Sullivan, C.S. A Human Torque Teno Virus Encodes a MicroRNA That Inhibits Interferon Signaling. PLoS Pathog. 2013, 9, e1003818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieg, A.M. Cpg motifs inbacterialdnaandtheirimmuneeffects. Annu. Rev. Immunol. 2002, 20, 709–760. [Google Scholar] [CrossRef] [PubMed]
- Freer, G.; Maggi, F.; Pifferi, M.; Di Cicco, M.E.; Peroni, D.G.; Pistello, M. The Virome and Its Major Component, Anellovirus, a Convoluted System Molding Human Immune Defenses and Possibly Affecting the Development of Asthma and Respiratory Diseases in Childhood. Front. Microbiol. 2018, 9, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vignolini, T.; Macera, L.; Antonelli, G.; Pistello, M.; Maggi, F.; Giannecchini, S. Investigation on torquetenovirus (TTV) microRNA transcriptome in vivo. Virus Res. 2016, 217, 18–22. [Google Scholar] [CrossRef]
- Itoha, Y.; Takahashib, M.; Fukudaa, M.; Shibayamac, T.; Ishikawad, T.; Tsudae, F.; Tanakaf, T.; Nishizawab, T.; Okamoto, H. Visualization of TT Virus Particles Recovered from the Sera and Feces of Infected Humans. Biochem. Biophys. Res. Commun. 2000, 279, 718–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakkola, L.; Bondén, H.; Hedman, L.; Kivi, N.; Moisala, S.; Julin, J.; Ylä-Liedenpohja, J.; Miettinen, S.; Kantola, K.; Hedman, K.; et al. Expression of all six human Torque teno virus (TTV) proteins in bacteria and in insect cells, and analysis of their IgG responses. Virology 2008, 382, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Ye, L.; Fang, X.; Li, B.; Wang, Y.; Xiang, X.; Kong, L.; Wang, W.; Zeng, Y.; Ye, L.; et al. Torque Teno Virus (SANBAN Isolate) ORF2 Protein Suppresses NF-κB Pathways via Interaction with IκB Kinases. J. Virol. 2007, 81, 11917–11924. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, F. IgM-class antibodies to TT virus (TTV) in patients with acute TTV infection. Hepatol. Res. 2001, 19, 1–11. [Google Scholar] [CrossRef]
- Mankotia, D.S.; Irshad, M. Cloning and expression of N22 region of Torque Teno virus (TTV) genome and use of peptide in developing immunoassay for TTV antibodies. Virol. J. 2014, 11, 96. [Google Scholar] [CrossRef]
- Martelli, F.; Macera, L.; Spezia, P.G.; Medici, C.; Pistello, M.; Guasti, D.; Romagnoli, P.; Maggi, F.; Giannecchini, S. Torquetenovirus detection in exosomes enriched vesicles circulating in human plasma samples. Virol. J. 2018, 15, 145. [Google Scholar] [CrossRef]
- World Health Organization. Global Surveillance, Prevention and Control of Chronic Respiratory Diseases. A Comprehensive Approach; WHO: Geneva, Switzerland, 2007; Available online: http://www.who.int/gard/publications/GARD_Manual/en/ (accessed on 25 January 2020).
- Mukherjee, M.; Stoddart, A.; Gupta, R.P.; Nwaru, B.I.; Farr, A.; Heaven, M.; Fitzsimmons, D.; Bandyopadhyay, A.; Aftab, C.; Simpson, C.; et al. The epidemiology, healthcare and societal burden and costs of asthma in the UK and its member nations: Analyses of standalone and linked national databases. BMC Med. 2016, 14, 113. [Google Scholar] [CrossRef] [Green Version]
- Arrieta, M.-C.; Stiemsma, L.T.; Dimitriu, P.A.; Thorson, L.; Russell, S.; Yurist-Doutsch, S.; Kuzeljevic, B.; Gold, M.J.; Britton, H.M.; Lefebvre, D.L.; et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci. Transl. Med. 2015, 7, 307ra152. [Google Scholar] [CrossRef]
- Lal, C.V.; Travers, C.; Aghai, Z.H.; Eipers, P.; Jilling, T.; Halloran, B.; Carlo, W.A.; Keeley, J.; Rezonzew, G.; Kumar, R.; et al. The Airway Microbiome at Birth. Sci. Rep. 2016, 6, 31023. [Google Scholar] [CrossRef] [Green Version]
- Pearce, N.; Aït-Khaled, N.; Beasley, R.; Mallol, J.; Keil, U.; Mitchell, E.; Robertson, C. Worldwide trends in the prevalence of asthma symptoms: Phase III of the International Study of Asthma and Allergies in Childhood (ISAAC). Thorax 2007, 62, 758–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenney, W.; Bush, A.; Fitzgerald, D.A.; Fletcher, M.; Ostrem, A.; Pedersen, S.; Szefler, S.J.; Zar, H.J. Improving the global diagnosis and management of asthma in children. Thorax 2018, 73, 662–669. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, G.; La Grutta, S. The Burden of Pediatric Asthma. Front. Pediatr. 2018, 6, 186. [Google Scholar] [CrossRef] [Green Version]
- Global Initiative for Asthma. Global Strategy for Asthma Management and Prevention. 2021. Available online: www.ginasthma.org (accessed on 20 February 2021).
- Hossain, F.M.A.; Choi, J.Y.; Uyangaa, E.; Park, S.O.; Eo, S.K. The Interplay between Host Immunity and Respiratory Viral Infection in Asthma Exacerbation. Immune Netw. 2019, 19, e31. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.D.; Buchta, K.A.; Swenson, C.A.; Gern, J.E.; Busse, W.W. Rhinovirus-induced Interferon-γ and Airway Responsiveness in Asthma. Am. J. Respir. Crit. Care Med. 2003, 168, 1091–1094. [Google Scholar] [CrossRef] [PubMed]
- Chau-Etchepare, F.; Hoerger, J.L.; Kuhn, B.T.; A Zeki, A.; Haczku, A.; Louie, S.; Kenyon, N.J.; E Davis, C.; Schivo, M. Viruses and non-allergen environmental triggers in asthma. J. Investig. Med. 2019, 67, 1029–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jartti, T.; Bønnelykke, K.; Elenius, V.; Feleszko, W. Role of viruses in asthma. Semin. Immunopathol. 2020, 42, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jartti, T.; Gern, J.E. Role of viral infections in the development and exacerbation of asthma in children. J. Allergy Clin. Immunol. 2017, 140, 895–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, A.; Taylor, L.J.; Dothard, M.I.; Leiby, J.S.; Fitzgerald, A.S.; Khatib, L.A.; Collman, R.G.; Bushman, F.D. Redondoviridae, a Family of Small, Circular DNA Viruses of the Human Oro-Respiratory Tract Associated with Periodontitis and Critical Illness. Cell Host Microbe 2019, 25, 719–729.e4. [Google Scholar] [CrossRef]
- Bush, A. Cytokines and Chemokines as Biomarkers of Future Asthma. Front. Pediatr. 2019, 7, 72. [Google Scholar] [CrossRef] [Green Version]
- D’Anna, S.E.; Maniscalco, M.; Cappello, F.; Carone, M.; Motta, A.; Balbi, B.; Ricciardolo, F.L.M.; Caramori, G.; Di Stefano, A. Bacterial and viral infections and related inflammatory responses in chronic obstructive pulmonary disease. Ann. Med. 2021, 53, 135–150. [Google Scholar] [CrossRef] [PubMed]
- Budden, K.F.; Gellatly, S.L.; Wood, D.L.A.; Cooper, M.A.; Morrison, M.; Hugenholtz, P.; Hansbro, P.M. Emerging pathogenic links between microbiota and the gut–lung axis. Nat. Rev. Microbiol. 2017, 15, 55–63. [Google Scholar] [CrossRef]
- Langelier, C.; Kalantar, K.L.; Moazed, F.; Wilson, M.R.; Crawford, E.D.; Deiss, T.; Belzer, A.; Bolourchi, S.; Caldera, S.; Fung, M.; et al. Integrating host response and unbiased microbe detection for lower respiratory tract infection diagnosis in critically ill adults. Proc. Natl. Acad. Sci. USA 2018, 115, E12353–E12362. [Google Scholar] [CrossRef] [Green Version]
- Guttenberg, M.A.; Vose, A.T.; Tighe, R.M. Role of Innate Immune System in Environmental Lung Diseases. Curr. Allergy Asthma Rep. 2021, 21, 34. [Google Scholar] [CrossRef] [PubMed]
- Budden, K.F.; Shukla, S.; Rehman, S.F.; Bowerman, K.; Keely, S.; Hugenholtz, P.; Armstrong-James, D.; Adcock, I.M.; Chotirmall, S.H.; Chung, K.F.; et al. Functional effects of the microbiota in chronic respiratory disease. Lancet Respir. Med. 2019, 7, 907–920. [Google Scholar] [CrossRef]
- Zhou, Y.; Jackson, D.; Bacharier, L.B.; Mauger, D.; Boushey, H.; Castro, M.; Durack, J.; Huang, Y.; Jr, R.F.L.; Storch, G.A.; et al. The upper-airway microbiota and loss of asthma control among asthmatic children. Nat. Commun. 2019, 10, 5714. [Google Scholar] [CrossRef] [Green Version]
- Robledo Aceves, M.; Barrón Balderas, A.; Jaime Ornelas, M.L. Most commonly isolated viruses in asthma exacerbation and their correlation with eosinophil and total serum immunoglobulin E levels. Arch. Argent Pediatr. 2018, 116, 192–197. [Google Scholar]
- Megremis, S.; Constantinides, B.; Xepapadaki, P.; Bachert, C.; Neurath-Finotto, S.; Jartti, T.; Kowalski, M.L.; Sotiropoulos, A.G.; Tapinos, A.; Vuorinen, T.; et al. Bacteriophage deficiency characterizes respiratory virome dysbiosis in childhood asthma. bioRxiv 2020. [Google Scholar] [CrossRef]
- Görzer, I.; Haloschan, M.; Jaksch, P.; Klepetko, W.; Puchhammer-Stöckl, E. Plasma DNA levels of Torque teno virus and immunosuppression after lung transplantation. J. Heart Lung Transplant. 2014, 33, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Kaczorowska, J.; van der Hoek, L. Human anelloviruses: Diverse, omnipresent and commensal members of the virome. FEMS Microbiol. Rev. 2020, 44, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasetyo, A.A.; Desyardi, M.N.; Tanamas, J.; Kageyama, S.; Chikumi, H.; Shimizu, E. Respiratory Viruses and Torque Teno Virus in Adults with Acute Respiratory Infections. Intervirology 2015, 58, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Xue, Q.; Jiao, W.; Wu, J.; Yu, Y.; Zhao, L.; Xu, Y.; Deng, X.; Fang, G.; Zheng, Y.; et al. Associations Between Sputum Torque Teno Virus Load and Lung Function and Disease Severity in Patients With Chronic Obstructive Pulmonary Disease. Front. Med. 2021, 8, 618757. [Google Scholar] [CrossRef] [PubMed]
- Pifferi, M.; Maggi, F.; Andreoli, E.; Lanini, L.; De Marco, E.; Fornai, C.; Vatteroni, M.L.; Pistello, M.; Ragazzo, V.; Macchia, P.; et al. Associations between Nasal Torquetenovirus Load and Spirometric Indices in Children with Asthma. J. Infect. Dis. 2005, 192, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Maggi, F.; Bendinelli, M. Immunobiology of the Torque Teno Viruses and Other Anelloviruses. TT Viruses 2009, 331, 65–90. [Google Scholar] [CrossRef]
- Pifferi, M.; Maggi, F.; Caramella, D.; De Marco, E.; Andreoli, E.; Meschi, S.; Macchia, P.; Bendinelli, M.; Boner, A.L. High Torquetenovirus Loads Are Correlated With Bronchiectasis and Peripheral Airflow Limitation in Children. Pediatr. Infect. Dis. J. 2006, 25, 804–808. [Google Scholar] [CrossRef] [Green Version]
- Maggi, F.; Pifferi, M.; Tempestini, E.; Fornai, C.; Lanini, L.; Andreoli, E.; Vatteroni, M.; Presciuttini, S.; Pietrobelli, A.; Boner, A.; et al. TT Virus Loads and Lymphocyte Subpopulations in Children with Acute Respiratory Diseases. J. Virol. 2003, 77, 9081–9083. [Google Scholar] [CrossRef] [Green Version]
- Polavarapu, M.; Klonoff-Cohen, H. Serum ECP as a diagnostic marker for asthma in children less than 5 years: A systematic review and meta-analysis. Pediatr. Pulmonol. 2020, 55, 3243–3251. [Google Scholar] [CrossRef] [PubMed]
- Njå, F.; Røksund, O.D.; Carlsen, K.-H. Eosinophil cationic protein (ECP) in schoolchildren living in a mountainous area of Norway: A population-based study of ECP as a tool for diagnosing asthma in children with reference values. Allergy 2001, 56, 138–144. [Google Scholar] [CrossRef]
- Jiang, X.-G.; Yang, X.-D.; Lv, Z.; Zhuang, P.-H. Elevated serum levels of TNF-α, IL-8, and ECP can be involved in the development and progression of bronchial asthma. J. Asthma 2017, 55, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Linsuwanon, P.; Poovorawan, Y.; Li, L.; Deng, X.; Vongpunsawad, S.; Delwart, E. The Fecal Virome of Children with Hand, Foot, and Mouth Disease that Tested PCR Negative for Pathogenic Enteroviruses. PLoS ONE 2015, 10, e0135573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Koff, E.M.; De Winter-De Groot, K.M.; Bogaert, D. Development of the respiratory tract microbiota in cystic fibrosis. Curr. Opin. Pulm. Med. 2016, 22, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Willner, D.; Haynes, M.R.; Furlan, M.; Hanson, N.; Kirby, B.; Lim, Y.W.; Rainey, P.B.; Schmieder, R.; Youle, M.; Conrad, D.; et al. Case Studies of the Spatial Heterogeneity of DNA Viruses in the Cystic Fibrosis Lung. Am. J. Respir. Cell Mol. Biol. 2012, 46, 127–131. [Google Scholar] [CrossRef]


{kind=link}
| TTVs and Innate Immunity |
|
|
| TTVs and Adaptative Immunity |
|
|
|
| Immuno-Escape Mechanisms |
|---|
|
|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dodi, G.; Attanasi, M.; Di Filippo, P.; Di Pillo, S.; Chiarelli, F. Virome in the Lungs: The Role of Anelloviruses in Childhood Respiratory Diseases. Microorganisms 2021, 9, 1357. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071357
Dodi G, Attanasi M, Di Filippo P, Di Pillo S, Chiarelli F. Virome in the Lungs: The Role of Anelloviruses in Childhood Respiratory Diseases. Microorganisms. 2021; 9(7):1357. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071357
Chicago/Turabian StyleDodi, Giulia, Marina Attanasi, Paola Di Filippo, Sabrina Di Pillo, and Francesco Chiarelli. 2021. "Virome in the Lungs: The Role of Anelloviruses in Childhood Respiratory Diseases" Microorganisms 9, no. 7: 1357. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071357