
Climatic Zone and Soil Properties Determine the Biodiversity of the Soil Bacterial Communities Associated to Native Plants from Desert Areas of North-Central Algeria

 ,
,  , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
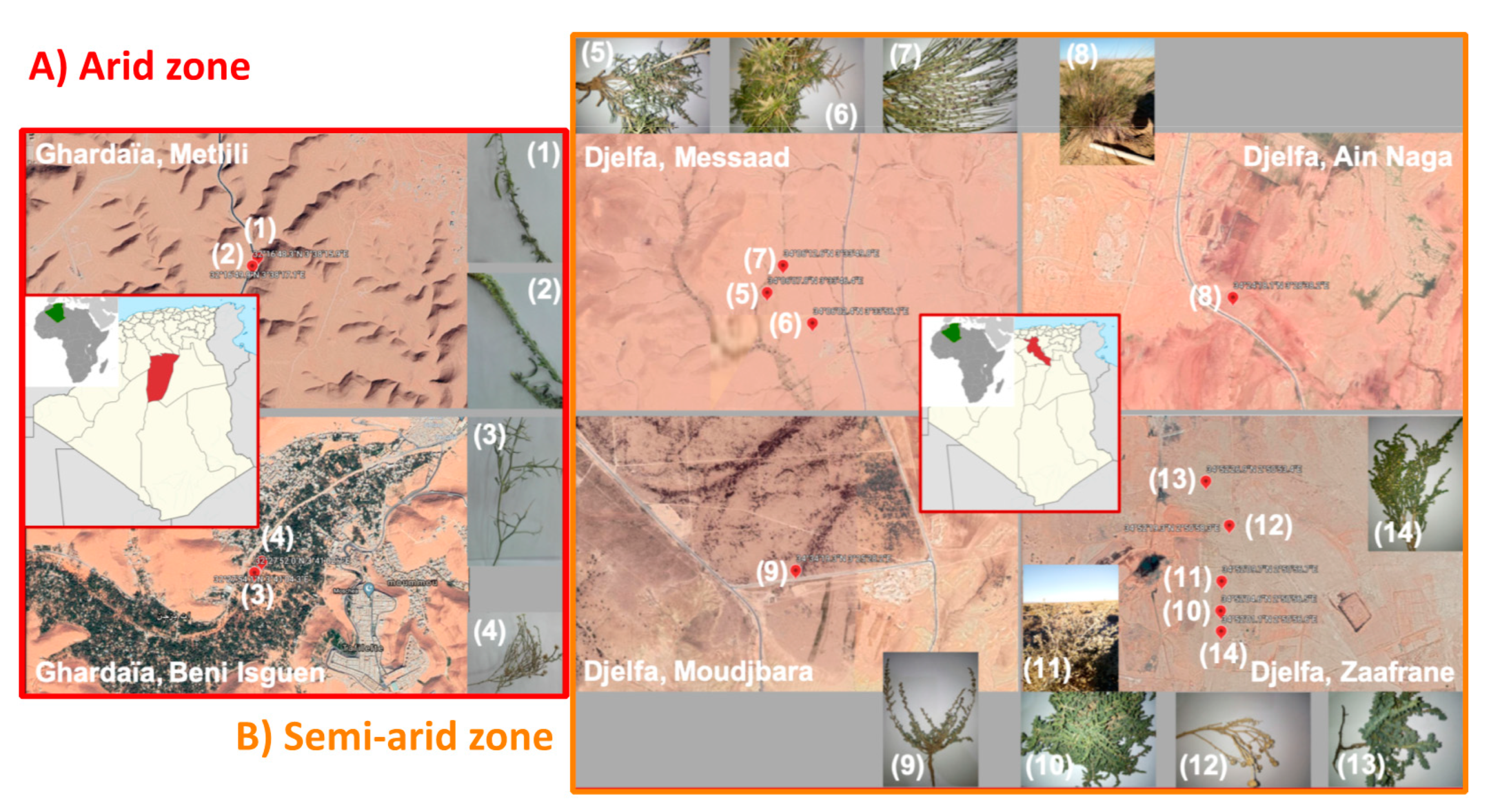
2.1. Soil Sampling
2.2. Microbiome Characterization
2.3. Bioinformatic and Statistical Analyses
3. Results
3.1. Soil Properties
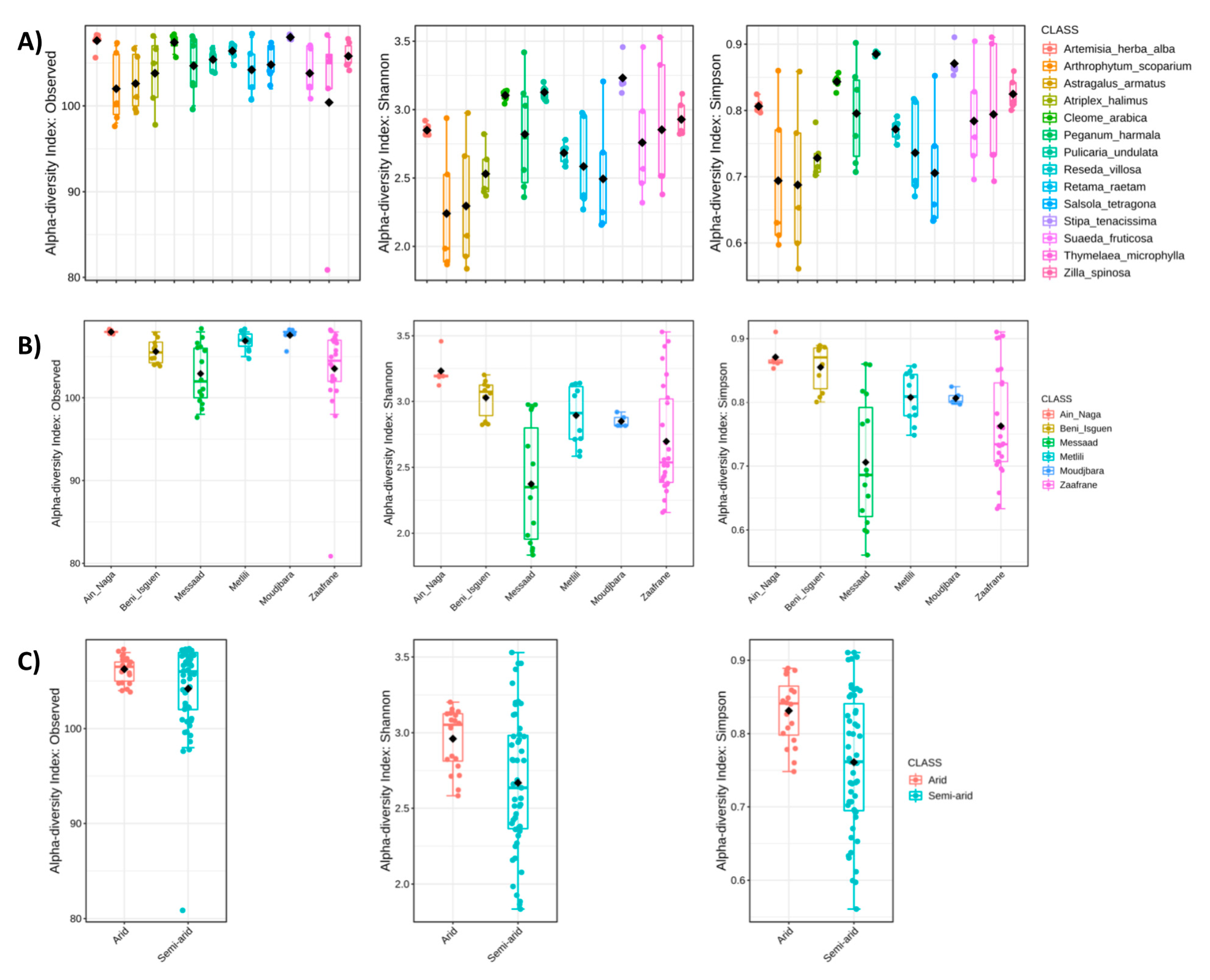
3.2. Biodiversity
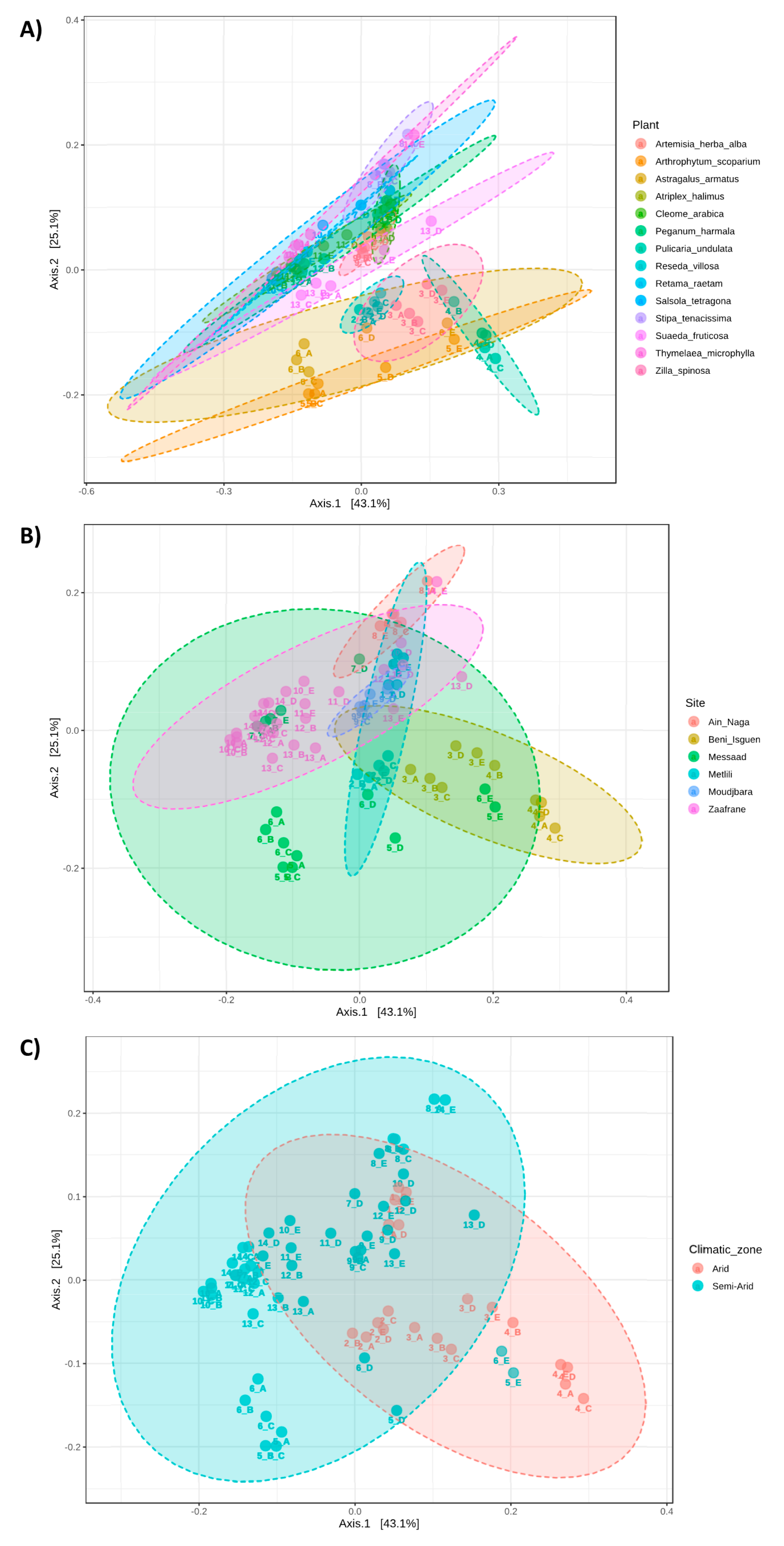
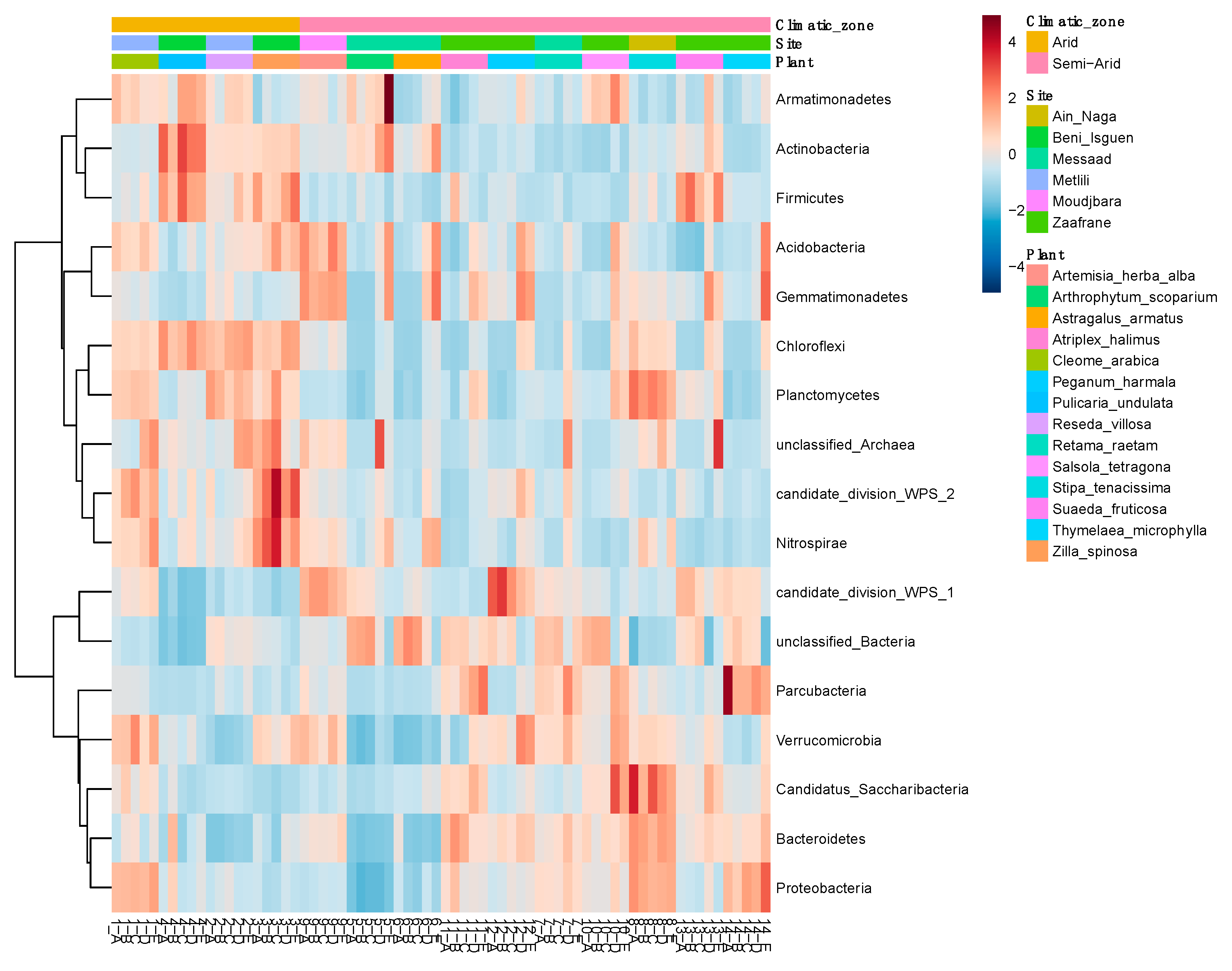
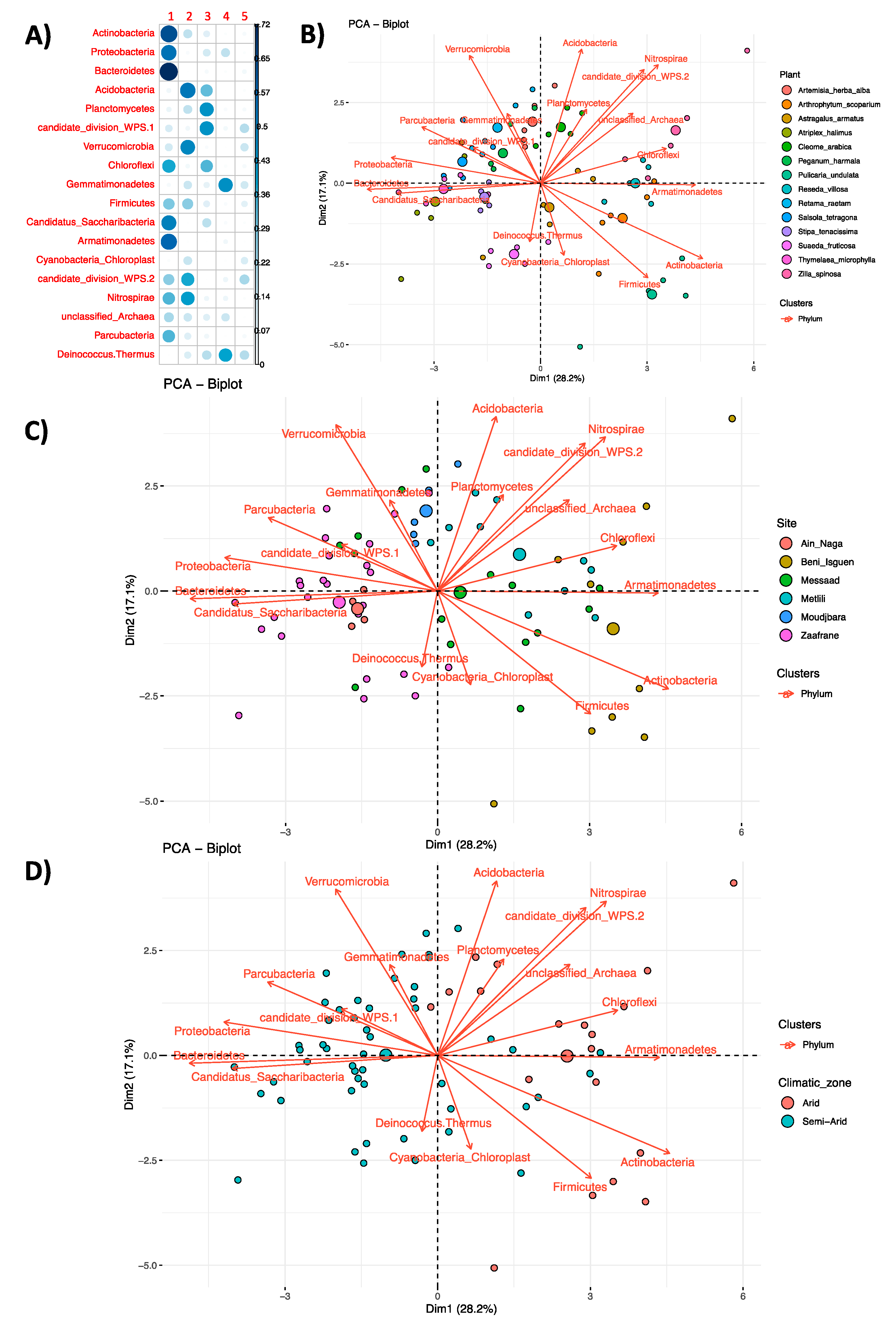
3.3. Community Profiling
3.4. Microbiota Comparison
3.5. Signature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van der Esch, S.; ten Brink, B.; Stehfest, E.; Bakkenes, M.; Sewell, A.; Bouwman, A.; Meijer, J.; Westhoek, H.; van den Berg, M. Exploring Future Changes in Land Use and Land Condition and the Impacts on Food, Water, Climate Change and Biodiversity; PBL Netherlands Environmental Assessment Agency: The Hague, The Netherlands, 2017.
- Djinni, I.; Defant, A.; Kecha, M.; Mancini, I. Actinobacteria Derived from Algerian Ecosystems as a Prominent Source of Antimicrobial Molecules. Antibiotics 2019, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Benarba, B. Medicinal Plants Used by Traditional Healers from South-West Algeria: An Ethnobotanical Study. J. Intercult. Ethnopharmacol. 2016, 5, 320. [Google Scholar] [CrossRef] [PubMed]
- Ouelbani, R.; Bensari, S.; Mouas, T.N.; Khelifi, D. Ethnobotanical Investigations on Plants Used in Folk Medicine in the Regions of Constantine and Mila (North-East of Algeria). J. Ethnopharmacol. 2016, 194, 196–218. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Berta, G.; Copetta, A.; Gamalero, E.; Bona, E.; Cesaro, P.; Scarafoni, A.; D’Agostino, G. Maize Development and Grain Quality Are Differentially Affected by Mycorrhizal Fungi and a Growth-Promoting Pseudomonad in the Field. Mycorrhiza 2014, 24, 161–170. [Google Scholar] [CrossRef]
- Bona, E.; Lingua, G.; Manassero, P.; Cantamessa, S.; Marsano, F.; Todeschini, V.; Copetta, A.; D’Agostino, G.; Massa, N.; Avidano, L.; et al. AM Fungi and PGP Pseudomonads Increase Flowering, Fruit Production, and Vitamin Content in Strawberry Grown at Low Nitrogen and Phosphorus Levels. Mycorrhiza 2015, 25, 181–193. [Google Scholar] [CrossRef]
- Bona, E.; Scarafoni, A.; Marsano, F.; Boatti, L.; Copetta, A.; Massa, N.; Gamalero, E.; D’Agostino, G.; Cesaro, P.; Cavaletto, M.; et al. Arbuscular Mycorrhizal Symbiosis Affects the Grain Proteome of Zea mays: A Field Study. Sci. Rep. 2016, 6, 26439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bona, E.; Lingua, G.; Todeschini, V. Effect of Bioinoculants on the Quality of Crops. In Bioformulations: For Sustainable Agriculture; Arora, N.K., Mehnaz, S., Balestrini, R., Eds.; Springer: New Delhi, India, 2016; pp. 93–124. ISBN 978-81-322-2777-9. [Google Scholar]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Pseudomonads Improve Yield, Quality and Nutritional Value of Tomato: A Field Study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bona, E.; Todeschini, V.; Cantamessa, S.; Cesaro, P.; Copetta, A.; Lingua, G.; Gamalero, E.; Berta, G.; Massa, N. Combined Bacterial and Mycorrhizal Inocula Improve Tomato Quality at Reduced Fertilization. Sci. Hortic. 2018, 234, 160–165. [Google Scholar] [CrossRef]
- Todeschini, V.; AitLahmidi, N.; Mazzucco, E.; Marsano, F.; Gosetti, F.; Robotti, E.; Bona, E.; Massa, N.; Bonneau, L.; Marengo, E.; et al. Impact of Beneficial Microorganisms on Strawberry Growth, Fruit Production, Nutritional Quality, and Volatilome. Front. Plant Sci. 2018, 9, 1611. [Google Scholar] [CrossRef]
- Massa, N.; Cesaro, P.; Todeschini, V.; Capraro, J.; Scarafoni, A.; Cantamessa, S.; Copetta, A.; Anastasia, F.; Gamalero, E.; Lingua, G.; et al. Selected Autochthonous Rhizobia, Applied in Combination with AM Fungi, Improve Seed Quality of Common Bean Cultivated in Reduced Fertilization Condition. Appl. Soil Ecol. 2020, 148, 103507. [Google Scholar] [CrossRef]
- Novello, G.; Cesaro, P.; Bona, E.; Massa, N.; Gosetti, F.; Scarafoni, A.; Todeschini, V.; Berta, G.; Lingua, G.; Gamalero, E. The Effects of Plant Growth-Promoting Bacteria with Biostimulant Features on the Growth of a Local Onion Cultivar and a Commercial Zucchini Variety. Agronomy 2021, 11, 888. [Google Scholar] [CrossRef]
- Santoyo, G.; Gamalero, E.; Glick, B.R. Mycorrhizal-Bacterial Amelioration of Plant Abiotic and Biotic Stress. Front. Sustain. Food Syst. 2021, 5, 18. [Google Scholar] [CrossRef]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of Microorganisms in the Evolution of Animals and Plants: The Hologenome Theory of Evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The Importance of the Microbiome of the Plant Holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial Interactions within the Plant Holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Ladhari, A.; Omezzine, F.; DellaGreca, M.; Zarrelli, A.; Zuppolini, S.; Haouala, R. Phytotoxic Activity of Cleome arabica L. and Its Principal Discovered Active Compounds. S. Afr. J. Bot. 2013, 88, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Habbachi, S.; Amri, N.; Habbachi, W.; Benhissen, S.; Tahraoui, A. Rebbas Toxic Effects of Cleome arabica L. (Capparidaceae) Aqueous Extracts on Mortality and Sexual Behavior of Drosophila melanogaster (Diptera: Drosophilidae). JABB 2019, 7, 137–143. [Google Scholar] [CrossRef]
- Berrehal, D.; Khalfallah, A.; Bencharif-Betina, S.; Kabouche, Z.; Kacem, N.; Kabouche, A.; Calliste, C.-A.; Duroux, J.-L. Comparative Antioxidant Activity of Two Algerian Reseda Species. Chem. Nat. Compd. 2010, 46, 456–458. [Google Scholar] [CrossRef]
- Chermat, S.; Gharzouli, R. Ethnobotanical Study of Medicinal Flora in the North East of Algeria—An Empirical Knowledge in Djebel Zdimm (Setif). J. Mater. Sci. Eng. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Shawky, R.A.; Alzamel, N.M. Survey on Medicinal Plants in the Flora of Al Riyadh Region, Saudi Arabia. EurAsian J. Biosci. 2020, 14, 3795–3800. [Google Scholar]
- Suleiman, M.H.A.; Ateeg, A.A. Antimicrobial and Antioxidant Activities of Different Extracts from Different Parts of Zilla spinosa (L.) Prantl. Evid. Based Complementary Altern. Med. 2020, 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- EL-Kamali, H.H.; Yousif, M.O.; Ahmed, O.I.; Sabir, S.S. Phytochemical Analysis of the Essential Oil from Aerial Parts of Pulicaria undulata (L.) Kostel from Sudan. Ethnobot. Leafl. 2009, 13, 467–471. [Google Scholar]
- Ravandeh, M.; Valizadeh, J.; Noroozifar, M.; Khorasani-Motlagh, M. Screening of Chemical Composition of Essential Oil, Mineral Elements and Antioxidant Activity in Pulicaria undulata (L.) C. A. Mey from Iran. J. Med. Plants Res. 2011, 5, 2035–2040. [Google Scholar]
- Ali, N.A.A.; Sharopov, F.S.; Alhaj, M.; Hill, G.M.; Porzel, A.; Arnold, N.; Setzer, W.N.; Schmidt, J.; Wessjohann, L. Chemical Composition and Biological Activity of Essential Oil from Pulicaria undulata from Yemen. Nat. Prod. Commun. 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Ajaib, M.; Mati-ur-Rehman, A.; Khan, K.M.; Perveen, S.; Shah, S. Pulicaria undulata: A Potential Phytochemical, Antimicrobial and Antioxidant Source. J. Chem. Soc. Pak. 2015, 37, 559–566. [Google Scholar]
- Boumaraf, M.; Mekkiou, R.; Benyahia, S.; Chalchat, J.; Benayache, F.; Benayache, S. Essential Oil Composition of Pulicaria undulata (L.) DC. (Asteraceae) Growing in Algeria. Int. J. Pharmacogn. Phytochem. Res. 2016, 8, 746–749. [Google Scholar]
- Chao, H.-C.; Najjaa, H.; Villareal, M.O.; Ksouri, R.; Han, J.; Neffati, M.; Isoda, H. Arthrophytum scoparium Inhibits Melanogenesis through the down-Regulation of Tyrosinase and Melanogenic Gene Expressions in B16 Melanoma Cells. Exp. Dermatol. 2013, 22, 131–136. [Google Scholar] [CrossRef]
- Smach, M.; Hafsa, J.; Chrfeddine, B.; Dridi, H.; Limem, K.; Jihene, B. Arthrophytum scoparium Extract Improves Memory Impairment and Affects Acetylcholinesterase Activity in Mice Brain. Curr. Pharm. Biotechnol. 2020, 21, 480–487. [Google Scholar] [CrossRef]
- Labed, A.; Ferhat, M.; Labed-Zouad, I.; Kaplaner, E.; Zerizer, S.; Voutquenne-Nazabadioko, L.; Alabdul Magid, A.; Semra, Z.; Kabouche, A.; Kabouche, Z.; et al. Compounds from the Pods of Astragalus armatus with Antioxidant, Anticholinesterase, Antibacterial and Phagocytic Activities. Pharm. Biol. 2016, 54, 3026–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edziri, H.; Mastouri, M.; Mahjoub, M.A.; Mighri, Z.; Mahjoub, A.; Verschaeve, L. Antibacterial, Antifungal and Cytotoxic Activities of Two Flavonoids from Retama raetam Flowers. Molecules 2012, 17, 7284–7293. [Google Scholar] [CrossRef] [Green Version]
- Khenblouche, A.; Bechki, D.; Gouamid, M.; Charradi, K.; Segni, L.; Hadjadj, M.; Boughali, S. Extraction and Characterization of Cellulose Microfibers from Retama raetam Stems. Polímeros 2019, 29, e2019011. [Google Scholar] [CrossRef]
- Hattalli, S.; Benaboura, A.; Ham-Pichavant, F.; Nourmamode, A.; Castellan, A. Adding Value to Alfa Grass (Stipa tenacissima L.) Soda Lignin as Phenolic Resins 1. Lignin Characterization. Polym. Degrad. Stab. 2002, 76, 259–264. [Google Scholar] [CrossRef]
- Nigam, M.; Atanassova, M.; Mishra, A.P.; Pezzani, R.; Devkota, H.P.; Plygun, S.; Salehi, B.; Setzer, W.N.; Sharifi-Rad, J. Bioactive Compounds and Health Benefits of Artemisia Species. Nat. Prod. Commun. 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Lakhdari, K.; Belhamra, M.; Mayouf, R. Forage Species Preferred by Dromedaries and Their Chemical Composition in Arid Rangelands of Algeria. Livest. Res. Rural. Dev. 2015, 27, 1. [Google Scholar]
- Lakhdari, W.; Dehliz, A.; Acheuk, F.; Mlik, R.; Hammi, H.; Doumandji-Mitiche, B.; Gheriani, S.; Berrekbia, M.; Guernit, K.; Chergui, S. Ethnobotanical Study of Some Plants Used in Traditional Medicine in the Region of Oued Righ (Algerian Sahara). J. Med. Plants Stud. 2016, 4, 204–211. [Google Scholar]
- Tapia, Y.; Diaz, O.; Pizarro, C.; Segura, R.; Vines, M.; Zúñiga, G.; Moreno-Jiménez, E. Atriplex atacamensis and Atriplex halimus Resist As Contamination in Pre-Andean Soils (Northern Chile). Sci. Total Environ. 2013, 450–451, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Ramdane, F.; Hadj Mahammed, M.; Didi Ould Hadj, M.; Hammoudi, R.; Hillali, N.; Mesrouk, H.; Bouafia, I.; Bahaz, C. Ethnobotanical Study of Some Medicinal Plants from Hoggar, Algeria. J. Med. Plants Res. 2015, 9, 820–827. [Google Scholar] [CrossRef]
- Herraiz, T.; González, D.; Ancín-Azpilicueta, C.; Arán, V.J.; Guillén, H. β-Carboline Alkaloids in Peganum harmala and Inhibition of Human Monoamine Oxidase (MAO). Food Chem. Toxicol. 2010, 48, 839–845. [Google Scholar] [CrossRef]
- Mzoughi, Z.; Abdelhamid, A.; Rihouey, C.; Le Cerf, D.; Bouraoui, A.; Majdoub, H. Optimized Extraction of Pectin-like Polysaccharide from Suaeda fruticosa Leaves: Characterization, Antioxidant, Anti-Inflammatory and Analgesic Activities. Carbohydr. Polym. 2018, 185, 127–137. [Google Scholar] [CrossRef]
- Kerbab, K.; Mekhelfi, T.; Zaiter, L.; Benayache, S.; Benayache, F.; Picerno, P.; Mencherini, T.; Sansone, F.; Aquino, R.P.; Rastrelli, L. Chemical Composition and Antioxidant Activity of a Polar Extract of Thymelaea microphylla Coss. et Dur. Nat. Prod. Res. 2015, 29, 671–675. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, Z.S.L.; Sharpton, T.J.; Grünwald, N.J. Metacoder: An R Package for Visualization and Manipulation of Community Taxonomic Diversity Data. PLoS Comput. Biol. 2017, 13, e1005404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Soft. 2008, 25. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Package “Facto Extra”. 2016. Available online: http://www.sthda.com/english/rpkgs/factoextra (accessed on 13 May 2021).
- Segata, N.; Boernigen, D.; Tickle, T.L.; Morgan, X.C.; Garrett, W.S.; Huttenhower, C. Computational Meta’omics for Microbial Community Studies. Mol. Syst. Biol. 2013, 9, 666. [Google Scholar] [CrossRef]
- Gamalero, E.; Bona, E.; Todeschini, V.; Lingua, G. Saline and Arid Soils: Impact on Bacteria, Plants, and Their Interaction. Biology 2020, 9, 116. [Google Scholar] [CrossRef]
- Giandon, P.; Bortolami, P. L’interpretazione Delle Analisi Del Terreno; ARPAV: Padova, Italy, 2007; ISBN 88-7504-115-6. [Google Scholar]
- Khan, A.L.; Asaf, S.; Abed, R.M.; Ning Chai, Y.; Al-Rawahi, A.N.; Mohanta, T.K.; Al-Rawahi, A.; Schachtman, D.P.; Al-Harrasi, A. Rhizosphere Microbiome of Arid Land Medicinal Plants and Extra Cellular Enzymes Contribute to Their Abundance. Microorganisms 2020, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Lee, S.-W. Bioprospecting Potential of the Soil Metagenome: Novel Enzymes and Bioactivities. Genom. Inform. 2013, 11, 114. [Google Scholar] [CrossRef]
- Rasche, F.; Marco-Noales, E.; Velvis, H.; van Overbeek, L.S.; López, M.M.; van Elsas, J.D.; Sessitsch, A. Structural Characteristics and Plant-Beneficial Effects of Bacteria Colonizing the Shoots of Field Grown Conventional and Genetically Modified T4-Lysozyme Producing Potatoes. Plant Soil 2006, 289, 123–140. [Google Scholar] [CrossRef]
- Khan, A.L.; Al-Harrasi, A.; Al-Rawahi, A.; Al-Farsi, Z.; Al-Mamari, A.; Waqas, M.; Asaf, S.; Elyassi, A.; Mabood, F.; Shin, J.-H.; et al. Endophytic Fungi from Frankincense Tree Improves Host Growth and Produces Extracellular Enzymes and Indole Acetic Acid. PLoS ONE 2016, 11, e0158207. [Google Scholar] [CrossRef]
- Goudjal, Y.; Toumatia, O.; Sabaou, N.; Barakate, M.; Mathieu, F.; Zitouni, A. Endophytic Actinomycetes from Spontaneous Plants of Algerian Sahara: Indole-3-Acetic Acid Production and Tomato Plants Growth Promoting Activity. World J. Microbiol. Biotechnol. 2013, 29, 1821–1829. [Google Scholar] [CrossRef] [Green Version]
- Basil, A.; Strap, J.L.; Knotek-Smith, H.M.; Krawford, D.L. Studies on the Microbial Population of the Rhizosphere of Big Sagebrush (Artemisia triridentata). J. Ind. Microbiol. Biotechnol. 2004, 31, 278–288. [Google Scholar] [CrossRef]
- Toumatia, O.; Compant, S.; Yekkour, A.; Goudjal, Y.; Sabaou, N.; Mathieu, F.; Sessitsch, A.; Zitouni, A. Biocontrol and Plant Growth Promoting Properties of Streptomyces mutabilis Strain IA1 Isolated from a Saharan Soil on Wheat Seedlings and Visualization of Its Niches of Colonization. S. Afr. J. Bot. 2016, 105, 234–239. [Google Scholar] [CrossRef]
- Vásquez-Dean, J.; Maza, F.; Morel, I.; Pulgar, R.; González, M. Microbial Communities from Arid Environments on a Global Scale. A Systematic Review. Biol. Res. 2020, 53, 29. [Google Scholar] [CrossRef]
- Fierer, N.; Leff, J.W.; Adams, B.J.; Nielsen, U.N.; Bates, S.T.; Lauber, C.L.; Owens, S.; Gilbert, J.A.; Wall, D.H.; Caporaso, J.G. Cross-Biome Metagenomic Analyses of Soil Microbial Communities and Their Functional Attributes. Proc. Natl. Acad. Sci. USA 2012, 109, 21390–21395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neilson, J.W.; Quade, J.; Ortiz, M.; Nelson, W.M.; Legatzki, A.; Tian, F.; LaComb, M.; Betancourt, J.L.; Wing, R.A.; Soderlund, C.A.; et al. Life at the Hyperarid Margin: Novel Bacterial Diversity in Arid Soils of the Atacama Desert, Chile. Extremophiles 2012, 16, 553–566. [Google Scholar] [CrossRef]
- Torres-Cortés, G.; Millán, V.; Fernández-González, A.J.; Aguirre-Garrido, J.F.; Ramírez-Saad, H.C.; Fernández-López, M.; Toro, N.; Martínez-Abarca, F. Bacterial Community in the Rhizosphere of the Cactus Species Mammillaria carnea during Dry and Rainy Seasons Assessed by Deep Sequencing. Plant Soil 2012, 357, 275–288. [Google Scholar] [CrossRef]
- Tkacz, A.; Cheema, J.; Chandra, G.; Grant, A.; Poole, P.S. Stability and Succession of the Rhizosphere Microbiota Depends upon Plant Type and Soil Composition. ISME J. 2015, 9, 2349–2359. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Luo, Y.; Wei, Y.; Huang, Y.; Zhang, H.; He, W.; Sheng, H.; An, L. Effect of Aridity and Dune Type on Rhizosphere Soil Bacterial Communities of Caragana microphylla in Desert Regions of Northern China. PLoS ONE 2019, 14, e0224195. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.B.; Andrade, G.S.; Meneghin, S.P.; Vicentini, R.; Ottoboni, L.M.M. Prospecting Plant Growth-Promoting Bacteria Isolated from the Rhizosphere of Sugarcane Under Drought Stress. Curr. Microbiol. 2019, 76, 1345–1354. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Phylum | p-Values | LDA Score | Arid/Semi-Arid % |
|---|---|---|---|---|
| Unclassified Acidobacteria Gp4 | Acidobacteria | 0.0270070 | 3.95 | 64% |
| unclassified Rubrobacter | Actinobacteria | 0.0003125 | 4.65 | 56% |
| unclassified Microbacteriaceae | Actinobacteria | 0.0151520 | 3.63 | 45% |
| unclassified Ilumatobacter | Actinobacteria | 0.0162540 | 3.67 | 42% |
| unclassified Armatimonas1Armatimonadetes gp1 | Armatimonadetes | 0.0019786 | 3.64 | 54% |
| unclassified Ohtaekwangia | Bacteroidetes | 0.0026650 | 4.64 | 48% |
| unclassified Cytophagales | Bacteroidetes | 0.0270060 | 3.51 | 40% |
| unclassified Bacteroidetes | Bacteroidetes | 0.0000008 | 4.81 | 24% |
| unclassified Flavobacteriaceae | Bacteroidetes | 0.0000008 | 4.25 | 2% |
| unclassified Saccharibacteria genera incertae sedis | Candidatus Saccharibacteria | 0.0006425 | 4.34 | 49% |
| unclassified Parcubacteria genera incertae sedis | Parcubacteria | 0.0014554 | 3.44 | 31% |
| unclassified Steroidobacter | Proteobacteria | 0.0055396 | 3.45 | 70% |
| unclassified Alphaproteobacteria | Proteobacteria | 0.0007742 | 4.51 | 65% |
| unclassified Gammaproteobacteria | Proteobacteria | 0.0053258 | 4.14 | 62% |
| unclassified Sphingomonadaceae | Proteobacteria | 0.0019786 | 4.43 | 61% |
| gamma proteobacterium SA29 B | Proteobacteria | 0.0183520 | 3.38 | 48% |
| unclassified Phaselicystis | Proteobacteria | 0.0005251 | 3.27 | 42% |
| unclassified Sphingomonadales | Proteobacteria | 0.0000782 | 3.94 | 41% |
| unclassified Phyllobacteriaceae | Proteobacteria | 0.0040260 | 3.30 | 39% |
| unclassified Caulobacteraceae | Proteobacteria | 0.0000018 | 3.72 | 25% |
| unclassified Xanthomonadaceae | Proteobacteria | 0.0003989 | 4.15 | 24% |
| unclassified Erythrobacteraceae | Proteobacteria | 0.0000002 | 4.07 | 11% |
| unclassified Bacteria | unclassified Bacteria | 0.0038659 | 5.60 | 83% |
| unclassified WPS 1 genera incertae sedis | unclassified Bacteria | 0.0000185 | 4.89 | 52% |
| unclassified Spartobacteria genera incertae sedis | Verrucomicrobia | 0.0006130 | 3.83 | 52% |
| unclassified Verrucomicrobia | Verrucomicrobia | 0.0001712 | 4.22 | 52% |
| unclassified Acidobacteria Gp3 | Acidobacteria | 0.0000050 | −4.09 | 282% |
| unclassified Acidobacteria | Acidobacteria | 0.0075492 | −3.78 | 134% |
| unclassified Gaiella | Actinobacteria | 0.0000037 | −4.99 | 446% |
| unclassified Aciditerrimonas | Actinobacteria | 0.0000029 | −4.31 | 369% |
| unclassified Solirubrobacter | Actinobacteria | 0.0000050 | −3.89 | 284% |
| unclassified Thermoleophilum | Actinobacteria | 0.0000103 | −3.69 | 278% |
| unclassified Actinobacteria | Actinobacteria | 0.0000047 | −5.42 | 200% |
| unclassified Euzebya | Actinobacteria | 0.0244220 | −3.50 | 180% |
| unclassified Solirubrobacterales | Actinobacteria | 0.0000536 | −4.31 | 159% |
| unclassified Pseudonocardiaceae | Actinobacteria | 0.0156950 | −3.64 | 142% |
| unclassified Actinomycetales | Actinobacteria | 0.0015237 | −4.63 | 131% |
| unclassified Micromonosporaceae | Actinobacteria | 0.0081443 | −3.89 | 130% |
| unclassified Acidimicrobiales | Actinobacteria | 0.0081443 | −4.00 | 127% |
| unclassified Archaea | Archea | 0.0019786 | −3.57 | 184% |
| unclassified Armatimonadetes gp4 | Armatimonadetes | 0.0000000 | −3.86 | 314% |
| unclassified Adhaeribacter | Bacteroidetes | 0.0189940 | −3.69 | 242% |
| Adhaeribacter aquaticus (T) | Bacteroidetes | 0.0213760 | −3.45 | 203% |
| unclassified Anaerolineaceae | Chloroflexi | 0.0000000 | −4.84 | 630% |
| unclassified Chloroflexi | Chloroflexi | 0.0000220 | −4.59 | 194% |
| unclassified Sphaerobacter | Chloroflexi | 0.0003611 | −3.52 | 175% |
| unclassified Litorilinea | Chloroflexi | 0.0022080 | −3.07 | 147% |
| unclassified Bacillales | Firmicutes | 0.0000010 | −3.92 | 215% |
| unclassified Pirellula | Planctomycetes | 0.0000001 | −4.26 | 309% |
| unclassified Planctomycetes | Planctomycetes | 0.0000405 | −3.40 | 205% |
| unclassified Gemmata | Planctomycetes | 0.0031487 | −3.29 | 166% |
| unclassified Planctomycetaceae | Planctomycetes | 0.0069934 | −4.56 | 129% |
| unclassified Rubellimicrobium | Proteobacteria | 0.0000050 | −4.34 | 583% |
| unclassified Burkholderiales | Proteobacteria | 0.0000042 | −4.27 | 273% |
| unclassified Cystobacteraceae | Proteobacteria | 0.0018147 | −3.82 | 253% |
| unclassified Microvirga | Proteobacteria | 0.0006848 | −4.09 | 225% |
| unclassified Nitrospira | Proteobacteria | 0.0000246 | −3.58 | 223% |
| unclassified Deltaproteobacteria | Proteobacteria | 0.0000233 | −3.87 | 181% |
| unclassified Rhodospirillales | Proteobacteria | 0.0004393 | −3.49 | 153% |
| unclassified Sphingomonas | Proteobacteria | 0.0147010 | −3.66 | 140% |
| unclassified Betaproteobacteria | Proteobacteria | 0.0307510 | −4.16 | 125% |
| unclassified WPS 2 genera incertae sedis | unclassified Bacteria | 0.0000246 | −4.09 | 216% |
| unclassified Gp3 | unclassified Bacteria | 0.0000308 | −3.79 | 199% |
| unclassified Subdivision3 genera incertae sedis | unclassified Bacteria | 0.0001994 | −4.49 | 175% |
| unclassified Gp7 | unclassified Bacteria | 0.0008490 | −4.09 | 171% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bona, E.; Massa, N.; Toumatia, O.; Novello, G.; Cesaro, P.; Todeschini, V.; Boatti, L.; Mignone, F.; Titouah, H.; Zitouni, A.; et al. Climatic Zone and Soil Properties Determine the Biodiversity of the Soil Bacterial Communities Associated to Native Plants from Desert Areas of North-Central Algeria. Microorganisms 2021, 9, 1359. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071359
Bona E, Massa N, Toumatia O, Novello G, Cesaro P, Todeschini V, Boatti L, Mignone F, Titouah H, Zitouni A, et al. Climatic Zone and Soil Properties Determine the Biodiversity of the Soil Bacterial Communities Associated to Native Plants from Desert Areas of North-Central Algeria. Microorganisms. 2021; 9(7):1359. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071359
Chicago/Turabian StyleBona, Elisa, Nadia Massa, Omrane Toumatia, Giorgia Novello, Patrizia Cesaro, Valeria Todeschini, Lara Boatti, Flavio Mignone, Houda Titouah, Abdelghani Zitouni, and et al. 2021. "Climatic Zone and Soil Properties Determine the Biodiversity of the Soil Bacterial Communities Associated to Native Plants from Desert Areas of North-Central Algeria" Microorganisms 9, no. 7: 1359. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071359